Как действует креатин — статья avitasport.ru
Статьи >Поделиться
Поделиться
Твитнуть
Класснуть
Физический процесс
Универсальным источником энергии для всех процессов в организме является молекула АТФ (аденозина трифосфат). Все поступаемые с пищей источники энергии (жиры, белки и углеводы) сначала конвертируются путем различных биохимических реакций в АТФ, она является единственной молекулой, переносящей и передающей энергию в организме. Процесс высвобождения энергии из АТФ просходит при отщеплении фосфатной группы молекулы, формируется новая молекула называемая АДФ (аденозина дифосфат):
АТФ + H2O ⇒ АДФ + фосфорная кислота + энергия
В организме непрерывно происходит ресинез новых молекул АТФ для обеспечения энергозатрат различных постоянно протекающих процессов, обеспечивающих нормальную жизнедеятельность.
Потребность в энергии при больших нагрузках намного превышают энергоемкость того количества АТФ, которое синтезируется в организме за время их выполнения.
Затем в процесс отдачи энергии вступает креатинфосфат, содержащийся в мышечных тканях. Креатинфосфат является высокоэнергетическим соединением, подобно АТФ, и является определяющим фактором энергопродукции в мышечной ткани.
При отдаче энергии от креатинфосфата освобождается фосфатная группа, и он превращается в креатин. Далее фосфатная группа рекомбинируется с молекулой АДФ, образуя молекулу АТФ, являющуюся источником энергии:
креатинфосфат + АДФ ⇔ креатин + АТФ
Энергии креатинфосфата хватает на ~2 минуты. Далее вы доходите до состояния, когда все энергоресурсы мышечных клеток исчерпаны: и АТФ, и креатинфосфата. Выработка АТФ не успевает за энергетическими потребностями организма, ресурсы креатинфосфата исчерпаны – наступает мышечная усталость, делая невозможными мышечные сокращения. В перерывах между подходами креатин комбинируется с фосфатом в организме, используя синтезируемую во время отдыха АТФ, чтобы вновь образовать фосфокреатин. Именно поэтому обязательно делать нормальные перерывы между подходами, чтобы дать этому процессу рекомбинации креатинфосфата завершится полностью. Только после отдыха у вас снова появятся силы выкладываться на 100%.
Именно поэтому обязательно делать нормальные перерывы между подходами, чтобы дать этому процессу рекомбинации креатинфосфата завершится полностью. Только после отдыха у вас снова появятся силы выкладываться на 100%.
Что дает прием креатина?
Суточный расход креатина в спокойной жизни составляет примерно 2 г. Естественно, что при занятиях спортом расход увеличивается. Для покрытия этого расхода креатин может синтезироваться в организме (из 3-х аминокислот: глицина, аргинина и метионина), также креатин содержится в принимаемом с пищей мясе. Большая часть общего количества креатина запасается в скелетных мышцах.
Принимая креатин в качестве пищевой добавки, вы можете повысить уровень фосфокреатина, что приводит к увеличению количества запасенной в мышцах энергии. Это приводит к тому, что если при выполнении упражнений отказ мышц наступает, например, на 8 повторении, то энергетическая прибавка в АТФ в процессе выполнения упражнения позволит одолеть 9-10 повторений.
Почему необходимо принимать креатин, а не креатинфосфат или саму АТФ? Дело в том, что ни креатинфосфат, ни АТФ не усваиваются в кишечнике в своем исходном виде. А вот креатин, если он принимается орально, усваивается и успешно попадает в мышечные клетки. Образующийся креатин с током крови поступает в мышцы, где под влиянием фермента креатинкиназы превращается в креатинфосфат.
А вот креатин, если он принимается орально, усваивается и успешно попадает в мышечные клетки. Образующийся креатин с током крови поступает в мышцы, где под влиянием фермента креатинкиназы превращается в креатинфосфат.
Ultimate Nutrition 100% Creatine Monohydrate (300 g
Креатин является естественной составляющей мышечных и других тканей в нашем теле и играет жизненно важную роль в предоставлении и передачи энергии.
Микроизмельченный креатин устанавливает новые стандарты для всех добавок, входящие в категории Creatine Monohydrate, так как производимые при этом микрочастицы креатина в 20 раз меньше, чем в обычном креатине в порошковом виде. Несмотря на то, что процесс измельчения немного удорожает продукт, оно того стоит!
Преимущества:
- Мышцы становятся сильнее, более мощными и полны энергией.
- Креатин повышает прочность связок сухожилий, а также помогает увеличить размер мышц.
- Креатин – это биомолекула, помогающее ускорить процесс заправки мышечных клеток молекулами АТФ.

- Многочисленные исследования показали, что креатин не только повышает силу и работоспособность, но и увеличивает выносливость организма. Тренировки проходят дольше и эффективней.
- Микроизмельченный креатин лучше усваивается и быстрее всасыванию в кишечнике, чем обычный порошковый креатин.
- За счет увеличения общих этапов обработки и очистки, микроизмельченный креатин на конечном этапе представляет собой более чистый креатин.
- Эффект накачки, за счет клеточной гидротации, т.е. накапливания мышечными клетками воды.

Энергия, снабжающая мышечные сокращения, а также их релаксацию, предоставляется молекулами соединения аденозин трифосфата (АТФ). В нашем теле молекулы АТФ имеют ограниченный жизненный цикл, около 2 секунд при интенсивных мышечных нагрузках, поэтому нашим клеткам приходится постоянно производить их для поддержания энергией наши мышцы. Прилив энергии производится за счет распада молекул АТФ, когда одна фосфатная группа высвобождается, формируя значительную метаболическую энергию. Наш организм использует креатинфосфат для быстрого пополнения молекулами АТФ.
В нашем теле молекулы АТФ имеют ограниченный жизненный цикл, около 2 секунд при интенсивных мышечных нагрузках, поэтому нашим клеткам приходится постоянно производить их для поддержания энергией наши мышцы. Прилив энергии производится за счет распада молекул АТФ, когда одна фосфатная группа высвобождается, формируя значительную метаболическую энергию. Наш организм использует креатинфосфат для быстрого пополнения молекулами АТФ.
Таким образом, от того количества креатинфосфата, который храниться в наших клетках, зависит в каком количестве и как эффективно наш организм производит АТФ. Креатин – это биомолекула, являющаяся производителем АТФ. В свою очередь, в нашем организме креатин вырабатывается в печени и скапливается в местах хранения: в мозгу, в скелетный и сердечных мышцах, служащих готовыми «бассейнами» для производства АТФ. Креатин – ключевое вещество, помогающее ускорить процесс заправки мышц молекулами АТФ. Большее количество креатина, сосредоточенного внутри мышц, означает более высокое производство энергии, позволяющее мышцам работать дольше с максимальной интенсивностью.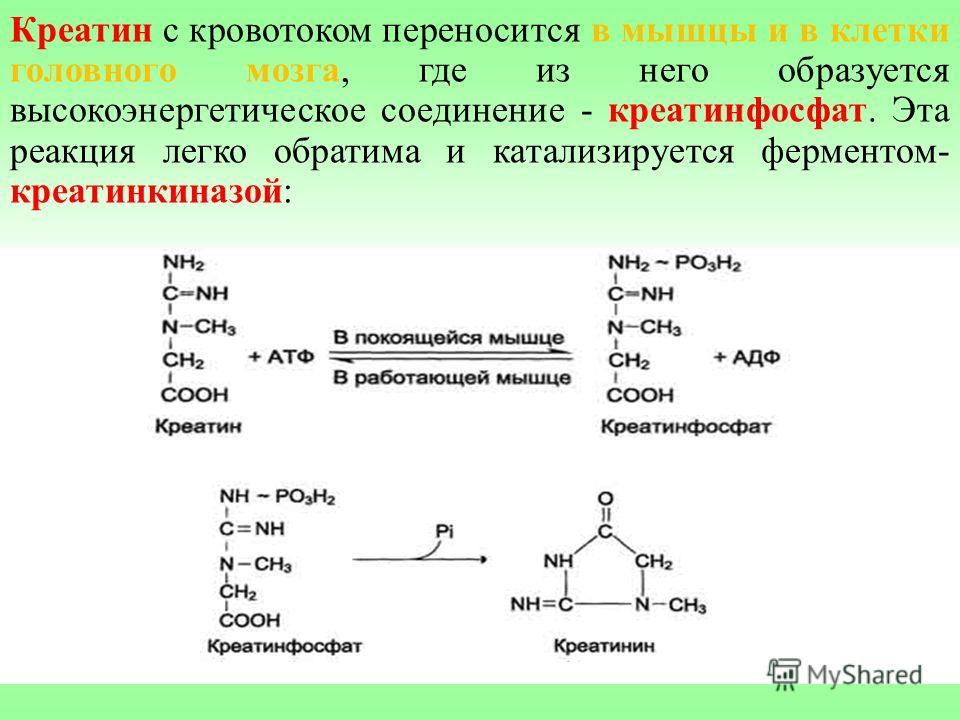
Количество питательных веществ в одной порции Ultimate Nutrition Creatine Monohydrate – 1 чайная ложка (примерно 5 г) продукта или 5 капсул:
- Креатин моногидрат – 4,5 г (в 5-и капсулах) или около 5 г (в одной чайной ложке)*
Рекомендации по применению:
- для порошка — 1 чайную ложку Ultimate Nutrition Creatine Monohydrate (примерно 5 г) развести на 150 мл воды, либо сока. Принимать преимущественно натощак.
- для капсул: порция 5 капсул. Принимать натощак.
1 неделя – ежедневная норма достигает 20 г креатина в день или по 1 порции (по 1-ой чайной ложке или по 5 капсул) 4 раза в день.
2-5 недели – ежедневная норма по 10 г креатина в день или по 1 порции (по 1-ой чайной ложке или по 5 капсул) 2 раза в день.
6-8 недели – разгрузка, никакого креатина.
Повторить курс.
Порций в упаковке:
1000 г — 200 порций.
300 г — 60 порций.
200 капсул — 40 порций.
КФК — Клиника 1
Креатинкиназа – фермент, который стимулирует превращение креатина в креатинфосфат и обеспечивает энергией мышечное сокращение.
Креатинкиназа – это фермент, который катализирует реакцию переноса фосфорильного остатка с АТФ на креатин с образованием креатинфосфата и АДФ. АТФ (аденозинтрифосфат) – молекула, являющаяся источником энергии в биохимических реакциях человеческого организма.
Для чего используется исследование?
- Для подтверждения диагнозов «инфаркт миокарда», «миокардит», «миокардиодистрофия».
- Для подтверждения диагнозов «полимиозит», «дерматомиозит», «миодистрофия».
- Чтобы проверить наличие заболеваний щитовидной железы.
- Чтобы убедиться в наличии опухолевого процесса и оценить его тяжесть.
- Чтобы оценить тяжесть течения полимиозита, дерматомиозита, миодистрофии, миопатии.
- Чтобы выявить носительство гена миопатии Дюшенна.
- Для диагностики и оценки тяжести поражения сердца и мышечной системы при интоксикации из-за инфекции, а также при отравлениях (угарным газом, ядом змеи, лекарственными средствами).

Когда назначается исследование?
- При симптомах ишемической болезни сердца.
- При симптомах инфаркта миокарда, в частности при стертой клинической картине, особенно при повторном инфаркте, атипичной локализации, болевом синдроме или ЭКГ-признаках, затруднении дифференциальной диагностики с другими формами ишемической болезни сердца.
- При гипотиреозе.
- При симптомах миозита, миодистрофии, миопатии.
- При планировании беременности женщиной, в семье которой были больные миопатией Дюшенна.
- При заболеваниях, которые могут привести к поражению сердца или мышечной системы.
Стоимость исследования
Креатин — для чего он нужен? — Статьи — Русскоязычный сайт болельщиков Депортиво
Откуда мышцы получают энергию во время фитнес тренировок?Для сокращения и совершения работы мышцам необходима энергия.
 Она поступает в мышечную клетку в «законсервированном» виде, а ее источником является молекула аденозинтрифосфорной кислоты (АТФ). Молекула АТФ образована аденозином и тремя фосфорными остатками. Когда от нее отщепляется один фосфат, происходит разрыв макроэргической связи, в результате чего высвобождается энергия. Утратившая фосфат АТФ преобразуется в АДФ (аденозиндифосфорная кислота). Чтобы снова стать носителем энергии, АДФ должна вернуть себе фосфорный остаток. Этим остатком АДФ снабжает креатинфосфат. Поделившись с АДФ фосфатом, молекула креатинфосфата превращается в креатин. Когда мышцы работают, тратится энергия, АТФ преобразуется в АДФ и, если в организме недостает креатинфосфата, АДФ накапливается. Слишком большое количество молекул АДФ вызывает состояние утомления и не позволяет мышцам сокращаться в полную силу.
Она поступает в мышечную клетку в «законсервированном» виде, а ее источником является молекула аденозинтрифосфорной кислоты (АТФ). Молекула АТФ образована аденозином и тремя фосфорными остатками. Когда от нее отщепляется один фосфат, происходит разрыв макроэргической связи, в результате чего высвобождается энергия. Утратившая фосфат АТФ преобразуется в АДФ (аденозиндифосфорная кислота). Чтобы снова стать носителем энергии, АДФ должна вернуть себе фосфорный остаток. Этим остатком АДФ снабжает креатинфосфат. Поделившись с АДФ фосфатом, молекула креатинфосфата превращается в креатин. Когда мышцы работают, тратится энергия, АТФ преобразуется в АДФ и, если в организме недостает креатинфосфата, АДФ накапливается. Слишком большое количество молекул АДФ вызывает состояние утомления и не позволяет мышцам сокращаться в полную силу.Зачем в программы спортивного питания включают креатин?
Во время интенсивных занятий фитнесом уровень креатинфосфата падает быстрее, чем АТФ.
 То есть возможности спортсмена ограничены не количеством АТФ в клетках, а в первую очередь дефицитом креатинфосфата. Это значит, что, если своевременно снабжать организм креатинфосфатом, можно усилить энергетический обмен в мышцах и добиться лучших силовых показателей, подробнее тут. Снижение уровней АТФ и креатинфосфата после физической нагрузки измеряли экспериментально. У бегунов, преодолевших 400 метров за 50 секунд, содержание АТФ в мышцах бедер уменьшилось на 27%, а креатинфосфата – на 90%. Чтобы уровень креатинфосфата восстановился, должно пройти не менее 5 минут. Разная скорость расходования креатинфосфата и молекул АТФ показывает, что именно нехватка креатинфосфата препятствует полному и своевременному обеспечению мышц энергий. Поскольку с помощью диеты снабдить организм спортсмена нужным количеством креатинфосфата невозможно, приходится принимать спортивные добавки. Но сам креатинфосфат расщепляется во время переваривания, поэтому употреблять его не имеет смысла. Вместо него тяжелоатлеты пьют креатин.
То есть возможности спортсмена ограничены не количеством АТФ в клетках, а в первую очередь дефицитом креатинфосфата. Это значит, что, если своевременно снабжать организм креатинфосфатом, можно усилить энергетический обмен в мышцах и добиться лучших силовых показателей, подробнее тут. Снижение уровней АТФ и креатинфосфата после физической нагрузки измеряли экспериментально. У бегунов, преодолевших 400 метров за 50 секунд, содержание АТФ в мышцах бедер уменьшилось на 27%, а креатинфосфата – на 90%. Чтобы уровень креатинфосфата восстановился, должно пройти не менее 5 минут. Разная скорость расходования креатинфосфата и молекул АТФ показывает, что именно нехватка креатинфосфата препятствует полному и своевременному обеспечению мышц энергий. Поскольку с помощью диеты снабдить организм спортсмена нужным количеством креатинфосфата невозможно, приходится принимать спортивные добавки. Но сам креатинфосфат расщепляется во время переваривания, поэтому употреблять его не имеет смысла. Вместо него тяжелоатлеты пьют креатин. Попав в кровоток, он переносится к мышцам и здесь под влиянием специального фермента превращается в креатинфосфат.
Попав в кровоток, он переносится к мышцам и здесь под влиянием специального фермента превращается в креатинфосфат.Какой эффект производит прием добавок с креатином?
Принимая дополнительные дозы креатина, спортсмен поддерживает свои мышцы в максимально рабочем состоянии. Креатин ускоряет образование АТФ, снижает скорость ее расходования во время силовых нагрузок, создает условия для расщепления АДФ и снижает утомляемость. Особенно сильно благотворное влияние креатина проявляется при интенсивных упражнениях, которые длятся не меньше десяти секунд. Помимо всего прочего, прием креатина позитивно влияет на состоянии сердечно-сосудистой и центральной нервной системы, снимает хронические и острые воспаления, например, при артрите, и, в силу стимуляции энергетических процессов, оказывает благотворное воздействие на организм в целом.
Источник: medaboutme.ru
Креатин Моногидрат Optimum System 100% Пюр 300г
Креатин моногидрат – это одна из ключевых добавок в силовых видах спорта, которая применяется для мощного повышения физической работоспособности. Креатин значительно повышает силовые показатели, выносливость, увеличивает мышечные объемы за счет повышенной гидратации клеток. Также добавка способна выступать буфером молочной кислоты, что позволяет мышцам дольше находится под нагрузкой до наступления отказа.
Креатин значительно повышает силовые показатели, выносливость, увеличивает мышечные объемы за счет повышенной гидратации клеток. Также добавка способна выступать буфером молочной кислоты, что позволяет мышцам дольше находится под нагрузкой до наступления отказа.
Креатин моногидрат – оптимальная форма добавки. Она отличается высокой эффективностью, низкой стоимостью и полной безопасностью. Вещество изучалось на протяжении десятков лет и за это время ученые не выявили каких-либо негативных воздействий на организм.
Принцип работы креатина Optimum System
- Креатин – это полностью натуральное вещество. Оно содержится в мышечных тканях и принимает непосредственное участие в энергетическом обмене. Без креатина невозможно было бы совершать мышечные сокращения и любую двигательную активность. Организм способен синтезировать вещество только в том количестве, которое необходимо для выполнения повседневной двигательной активности.
- При занятиях спортом происходит увеличенное высвобождение энергии. Это увеличивает потребность мышц в АТФ (аденозина трифосфат) в сотни раз. Креатин напрямую влияет на результативность атлета, увеличивая количество АТФ за счет уровня фосфокреатина. Это способствует росту силовой выносливости, силы, физической производительности.

Основные эффекты 100% Pure Creatine Monohydrate
- Креатин – добавка с широким спектром действия. Несмотря на то, что основной эффект достигается за счет повышения количества АТФ, дополнительный прием моногидрата способен оказывать комплексное повышение физической производительности.
Основные эффекты 100% Pure Creatine Monohydrate:
- Увеличение силовых показателей;
- Повышение аэробной и анаэробной выносливости;
- Увеличение мышечных объемов, улучшение качества и плотности мускулатуры;
- Повышение физической работоспособности.
Также некоторые исследования доказывают, что прием добавки способен незначительно повышать синтез анаболических гормонов и выступать буфером молочной кислоты.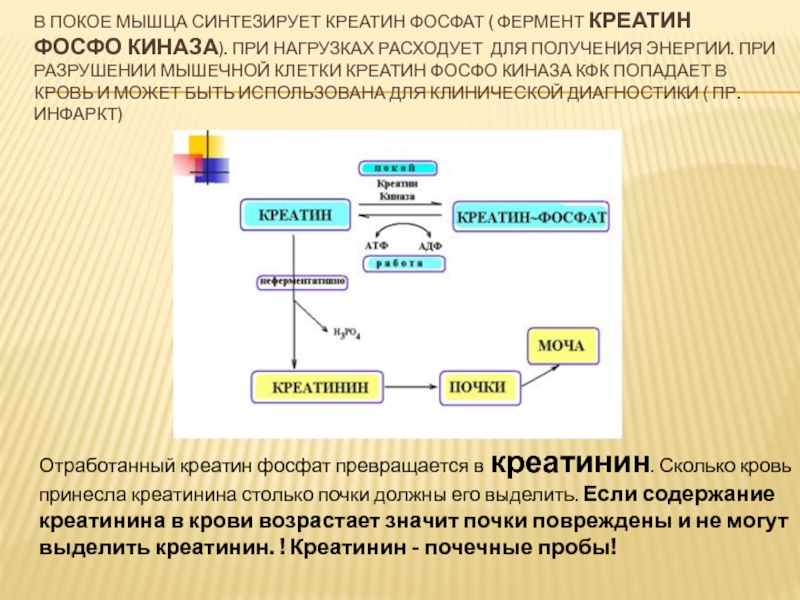 Это способствует ускорению роста мышечных тканей, восстановлению волокон, а также повышению пиковой производительности.
Это способствует ускорению роста мышечных тканей, восстановлению волокон, а также повышению пиковой производительности.
Способы приёма и дозировки
- Существует два способа употребления добавки: с фазой загрузки и фиксированным стабильным приёмом. Первый вариант подразумевает увеличение суточной дозировки вещества на первые 5 дней до 20 грамм (4 порции). После этого спортсмен переходит на поддерживающую дозировку в 2-3 грамма на весь последующий курс. Альтернативой является стабильный прием добавки в дозировке 5 грамм в сутки от первого до последнего дня курса.
- Оба способа применения одинаково эффективны, но фаза загрузки может повышать нагрузку на почки. Из-за этого большинство атлетов постепенно отказываются от неё, предпочитая стабильный прием в виде суточной дозировки в 5 грамм.
- Стандартный курс длится 1 месяц, после чего следует перерыв на 2-4 недели. Также существуют схемы приема 8/4 недель, где основная фаза приема продлевается до 2 месяцев.
- 100% Pure Creatine Monohydrate – это высококачественный креатин от известного бренда, который способен существенно ускорить прогрессирование в любых видах силового спорта. Добавка полностью безопасна для здоровья, а первый эффект в выраженном увеличении силовых показателей и выносливости отмечается уже к концу первой недели курса.
 Добавка полностью безопасна для здоровья, а первый эффект в выраженном увеличении силовых показателей и выносливости отмечается уже к концу первой недели курса.
Добавка полностью безопасна для здоровья, а первый эффект в выраженном увеличении силовых показателей и выносливости отмечается уже к концу первой недели курса.Креатин моногидрат – одна из основных добавок в силовых видах спорта. Способствует выраженному повышению силы, выносливости, увеличению мышечных объемов и гидратации клеток. Снижает «закисление» мышц во время нагрузки, отодвигая порог усталости и мышечного отказа. Вещество полностью безопасно для здоровья и не имеет каких-либо противопоказаний, за исключение периодов беременности, лактации, а также возраста до 18 лет. Применение креатина в виде добавки значительно увеличивает физическую производительность и качество тренировок для спортсменов любого уровня, способствует достижению лучших результатов за период времени в любых тренировочных циклах.
Срок годности: 24 месяца.
Принцип действия креатин моногидрата | Freestyle — парк экстремальных развлечений и горнолыжный центр в Москве
Анализируя спрос в интернет-магазине «Фиткульт», мы можем смело сказать, что среди креатиновых добавок самой популярной формулой для действующего вещества считается креатин моногидрат. Мы уверены, что такая ситуация не только в нашем, но и в других магазинах спортивного питания в Ростове-на-Дону.
Мы уверены, что такая ситуация не только в нашем, но и в других магазинах спортивного питания в Ростове-на-Дону.
Вот категория, где вы найдете самый большой выбор креатина. Кроме того, судя по самым актуальным исследованиям в мире спортивной медицины, аналогичная ситуация наблюдается во всем мире. В данной статье мы попробуем объяснить принцип действия моногидрата креатина в человеческом организме и эффект, оказываемый его приемом.
Креатин моногидрат является натуральным веществом, произведенным из естественных пищевых источников сырья. После абсорбции в человеческом организме в ходе обменных процессов моногидрат креатина превращается в креатин фосфат. При непосредственном участии креатин фосфата в клетках человеческого организма происходит синтез аденозин трифосфата, известного всем нам под аббревиатурой АТФ. Ну а АТФ — это главный источник энергии для наших мышц.
Человеческий организм в состоянии самостоятельно производить определенное количество креатина, но все же основным источником данного вещества являются пищевые продукты. Содержания креатина в том или ином продукте во многом зависит от способа его приготовления. Регулярные физические нагрузки увеличивают расход креатина человеческим организмом, а потому естественных пищевых источников для компенсации расходов недостаточно. Для того, чтобы мышц не страдали от недостатка энергии и рекомендуют принимать дополнительные спортивные добавки с креатином.
Содержания креатина в том или ином продукте во многом зависит от способа его приготовления. Регулярные физические нагрузки увеличивают расход креатина человеческим организмом, а потому естественных пищевых источников для компенсации расходов недостаточно. Для того, чтобы мышц не страдали от недостатка энергии и рекомендуют принимать дополнительные спортивные добавки с креатином.
Чтобы понять принцип работы креатина в нашем организме, необходимо понимать, как работает АТФ. Аденозин трифосфат является непосредственным источником энергии, которая необходима для того, чтобы мышцы сокращались. В мышечных волокнах содержится запас АТФ всего лишь для нескольких сокращений, дополнительные запасы вещества черпаются из организма. Креатин моногидрат является одним из источников для производства АТФ. На тренировке же больший запас энергии позволит вам брать большие веса, рассчитывать на большее количество повторений в подходах, улучшая таким образом вашу производительность.
Запись опубликована в рубрике Полезные статьи.Гребля на драконах,mdbc.ru.МОСКОВСКИЕДРАКОНЫ,dragonboat — Креатин
Креатин – азотсодержащая карбоновая кислота, которая участвует в энергетическом обмене в мышечных и нервных клетках. В мире бодибилдинга креатин широко используется как спортивная добавка для увеличения силы, мышечной массы и кратковременной анаэробной выносливости с доказанной безопасностью.
Биологическая роль креатина
Креатин – это натуральное вещество, которое содержится в мышцах человека и животных, и требуется для энергетического обмена и выполнения движений. В организме человека имеется около 100 -140 г этого вещества, выполняющего функцию источника энергии для мышц. Суточный расход креатина в обычных условиях составляет примерно 2 г. Креатин так же важен для жизни, как белок, углеводы, жиры, витамины и минералы. Креатин может синтезироваться организмом самостоятельно из 3-х аминокислот: глицина, аргинина и метионина. Эти аминокислоты – компоненты белка.
У людей ферменты, вовлеченные в синтез креатина, локализуются в печени, поджелудочной железе и почках. Креатин может быть произведен в любом из этих органов, и затем транспортирован кровью в мышцы. Приблизительно 95 % общего пула креатина запасается в тканях скелетной мускулатуры.
При увеличении физической нагрузки расход креатина тоже увеличивается, и его запас должен быть пополнен с помощью диеты или за счет собственного натурального производства организмом. Решающим фактором для достижения высоких результатов в спорте является способность организма высвобождать большое количество энергии за короткий промежуток времени. В принципе наш организм постоянно получает энергию, расщепляя углеводы и жир.
Непосредственным же источником энергии для сокращения скелетной мускулатуры является молекула, называемая АТФ (аденозина трифосфат). Количество АТФ имеющееся в непосредственном распоряжении, ограничено и является решающим для спортивной активности.
Все источники топлива – углеводы, жиры и белок – сначала конвертируются путем различных химических реакций в АТФ, которая затем становится доступной как единственная молекула, которую тело использует для энергии. Когда АТФ высвобождает энергию, чтобы обеспечить энергией мышечные сокращения, фосфатная группа отщепляется, и формируется новая молекула, называющаяся АДФ (аденозина дифосфат). Эта реакция обратима за счет креатин-фосфата, богатого энергией вещества. Креатин комбинируется с фосфатом в организме, чтобы образовать фосфокреатин, который является определяющим фактором энергопродукции в мышечной ткани.
Когда АТФ высвобождает энергию, чтобы обеспечить энергией мышечные сокращения, фосфатная группа отщепляется, и формируется новая молекула, называющаяся АДФ (аденозина дифосфат). Эта реакция обратима за счет креатин-фосфата, богатого энергией вещества. Креатин комбинируется с фосфатом в организме, чтобы образовать фосфокреатин, который является определяющим фактором энергопродукции в мышечной ткани.
Эффекты креатина
Увеличение силы:
В бодибилдинге, во время выполнения высокоинтенсивных упражнений потребность в АТФ в работающих мышцах значительно увеличивается – в сотни раз выше по сравнению с состоянием покоя. Истощенные запасы АТФ и фосфокреатина должны постоянно пополняться для того, чтобы мышечные сокращения могли продолжаться на пиковых уровнях частоты и интенсивности. Увеличивая фосфокреатин путем приема моногидрата креатина, вы можете увеличивать количество АТФ и, таким образом, повышается сила мышц.
Увеличение мышечной массы:
Нередки прибавления сухой массы тела от 2 до 5 кг за 1 месяц приема, при систематических тренировках и правильном питании. Недавние опыты доказывают, что креатин моногидрат способен увеличивать повторный максимум в жиме лежа на 10 кг уже через неделю приема, улучшать спринтерские способности. Увеличение силы позволяет добиться максимального ростостимулирующего воздействия на мышцы. Однако следует заметить, что прием креатина не всегда имеет подобный эффект. Он может проявляться гораздо слабее вследствие слабой чувствительности организма к креатину и выражаться лишь в некотором увеличении работоспособности без прироста силовых качеств.
Недавние опыты доказывают, что креатин моногидрат способен увеличивать повторный максимум в жиме лежа на 10 кг уже через неделю приема, улучшать спринтерские способности. Увеличение силы позволяет добиться максимального ростостимулирующего воздействия на мышцы. Однако следует заметить, что прием креатина не всегда имеет подобный эффект. Он может проявляться гораздо слабее вследствие слабой чувствительности организма к креатину и выражаться лишь в некотором увеличении работоспособности без прироста силовых качеств.
Качество мышц:
Ко всему прочему, креатин также улучшает рельефность мускулатуры. Моногидрат креатина связывается с водой, по мере того как он абсорбируется в мышечные клетки. Поскольку большее количество креатина запасается, большее количество воды привлекается в мышечную клетку. Это объясняет гидратирующее влияние креатина на мышечную клетку, которая состоит приблизительно на 75 % из воды. Бодибилдеры замечают: хорошо гидратированная мышца внешне выглядит более полной, более округлой и более накачанной. Научные исследования показывают, что когда клетки мышц увеличивают объем за счет сверхгидратации, синтез протеина увеличивается, а процесс его распада минимизируется (это также может усиливать синтез гликогена).
Научные исследования показывают, что когда клетки мышц увеличивают объем за счет сверхгидратации, синтез протеина увеличивается, а процесс его распада минимизируется (это также может усиливать синтез гликогена).
Увеличение секреции анаболических гормонов:
Исследования показывают, что креатин может увеличивать секрецию эндогенных анаболических гормонов в ответ на тренировочные нагрузки. Эти гормоны представлены соматотропином и тестостероном. Причем уровень соматотропина увеличивается только через 2 часа после приема креатина. Такая задержка может говорить о том, что выброс соматотропного гормона зависит не от самого креатина, а носит опосредованный характер и возникает в результате клеточного ответа.
Буфер молочной кислоты:
Есть свидетельства того, что креатин также работает как буфер молочной кислоты. Во время интенсивной работы в анаэробном режиме мышцы выделяют молочную кислоту, это вещество частично отвечает за то чувство жжения, которое возникает, когда мышца работает до отказа. Исследований показывают, что креатин может сдерживать выделение и действие молочной кислоты и улучшать время восстановления после недолгой интенсивной нагрузки (например, силовой тренировки).
Исследований показывают, что креатин может сдерживать выделение и действие молочной кислоты и улучшать время восстановления после недолгой интенсивной нагрузки (например, силовой тренировки).
Креатин для спортсменов
Главная ценность креатина, по-видимому, связана с усилением кратковременных спортивных показателей, например, в беге на короткие дистанции, велосипедном спринте, силовых видах спорта и, разумеется, в бодибилдинге. Креатин пригоден для видов спорта, в которых необходимо производить прыжки, ускорения или финишные рывки. В фазе рывка интенсивность нагрузки настолько велика, что при нем креатинфосфат (фосфокреатин) также используется в качестве источника энергии. Дополнение питания атлетов креатином может также приносить пользу, когда высокоинтенсивное упражнение чередуется с более низким по интенсивности упражнением или отдыхом. Для командных видов спорта, таких как баскетбол, футбол, хоккей, а также единоборств, тенниса, легкой атлетики и спринтерского бега также характерны короткие взрывчатые мышечные сокращения, сопровождаемые короткими периодами отдыха или восстановительными периодами.
Судя по всему, креатин помогает поддерживать высокий уровень быстрого снабжения тела энергией. Он также препятствует возрастанию содержания иона аммония в плазме крови, который в ином случае замедляет физическую деятельность.
Креатин для набора мышечной массы
Дополнение питания креатином может помочь атлету тренироваться тяжелее в течение более продолжительного периода времени. В свою очередь, возросшая интенсивность тренинга мышц генерирует более быстрый мышечный рост и силу. По наблюдениям одного шведского спортивного физиолога, креатин позволяет тяжелоатлетам добиваться увеличения мышечной массы на 2 кг (и более). Практические опыты показали, что увеличение общего пула креатина может способствовать увеличению веса тела. Многие атлеты, дополняющие диету креатином, отмечали увеличение задержки воды внутри мышечных клеток. Тем самым увеличивается объем клетки. Напряжение мышц, так называемый мышечный тонус, улучшается, и мышцы лучше тренировать. Спортсмен силовых видов спорта, весящий 75 кг может увеличить свой вес на 2-4 кг. После прекращения дополнительного приема креатина, прирост веса вновь уменьшается, благодаря повышенному выделению воды. Но за счет лучшей производительности во время тренировки, часть реального прироста мышечной массы остается. При этом в силовых видах спорта отмечается улучшение «рессорных» свойств мышц и их способности в силу этого преодолевать большие отягощения.
После прекращения дополнительного приема креатина, прирост веса вновь уменьшается, благодаря повышенному выделению воды. Но за счет лучшей производительности во время тренировки, часть реального прироста мышечной массы остается. При этом в силовых видах спорта отмечается улучшение «рессорных» свойств мышц и их способности в силу этого преодолевать большие отягощения.
Креатин: возраст и пол
Употребление креатина ассоциируется, как правило, с мужской половиной населения в возрастном диапазоне от 18 до 35 лет. Научные исследования, проведенные в группах с таким составом, точно подтверждают как эффективность креатина, так и его безопасность. Однако, эффекты креатина на детей, пожилых и женщин были изучены гораздо меньше. Теоретически, нет ни одной веской причины полагать, что основной механизм действия креатина разнится в зависимости от возраста или пола. Тем не менее, незначительные различия могут иметь место.
Несколько недавних научных исследований были посвящены рассмотрению действия креатина на женский организм при разном уровне физической подготовленности. Эти исследования показали, что хотя и креатин увеличивает силовые показатели женщин, но эффект этот меньше, чем у мужчин, участвующих в этих же исследованиях. Креатиновые добавки имели более выраженное действие, что объясняется более высоким уровнем тестостерона.
Эти исследования показали, что хотя и креатин увеличивает силовые показатели женщин, но эффект этот меньше, чем у мужчин, участвующих в этих же исследованиях. Креатиновые добавки имели более выраженное действие, что объясняется более высоким уровнем тестостерона.
Таким образом, креатин можно рекомендовать женщинам, особенно при снижении веса. При похудении креатиновые добавки позволят повысить интенсивность тренировок, а, следовательно, ускорить процесс разрушения жира. Ко всему прочему, креатин максимально сохранит мышечную массу, которая начинает активно теряться при голодании. Отсюда становится понятно, что креатин полезен не только для набора мышечной массы, но и для формирования красивой женской фигуры.
креатин | Tastycraze.com
Креатин — одна из самых эффективных и известных добавок, которые используются в силовых видах спорта. Он был обнаружен в 1832 году французским ученым Мишелем Эженом Шеврёлем. Креатин в организме образуется из аминокислот глицина, аргинина и метионина. В организме содержится около 120 г креатина, который находится в форме креатинфосфата. Его основная функция — помогать клеткам получать необходимую энергию.
В организме содержится около 120 г креатина, который находится в форме креатинфосфата. Его основная функция — помогать клеткам получать необходимую энергию.
Функции креатина
Принятие креатиновых добавок увеличивает концентрацию креатинфосфата в организме.В результате аденозинтрифосфат в организме увеличивает свою концентрацию — он может восстанавливаться быстрее и в течение более длительного времени способен снабжать мышцы энергией. Весь процесс можно объяснить следующим образом: мышцы человеческого тела используют аденозин-трифосфат / ATF / для получения необходимой энергии. Во время восстановления АТФ распадается с образованием аденозиндифосфата.
Для продолжения работы мышц аденозиндифосфат снова становится АТФ, присоединяясь к молекуле фосфата.Он взят из соединения кретин-фосфат, который, как уже упоминалось, циркулирует в организме. Тем самым восстанавливается присутствие необходимого АТФ, и снова заряжается организм энергией.
Выбор креатина
Разнообразие добавок креатина чрезвычайно велико. Наиболее распространены моногидрат креатина, глюконат креатина, малат креатина, цитрат креатина и многие другие. Это лишь очень небольшая часть всех форм креатина, имеющихся в продаже.Учитывая разнообразие, очень сложно определить, какой товар подходит лучше всего. Ряд экспертов в данной области согласны с тем, что лучшим является моногидрат креатина, КМ, и, в частности, микронизированный КМ, предпочтительнее, потому что он легче для желудка и растворяет его.
Наиболее распространены моногидрат креатина, глюконат креатина, малат креатина, цитрат креатина и многие другие. Это лишь очень небольшая часть всех форм креатина, имеющихся в продаже.Учитывая разнообразие, очень сложно определить, какой товар подходит лучше всего. Ряд экспертов в данной области согласны с тем, что лучшим является моногидрат креатина, КМ, и, в частности, микронизированный КМ, предпочтительнее, потому что он легче для желудка и растворяет его.
Преимущества креатина
Прием креатина дает спортсменам дополнительные силы. Эта энергия, в свою очередь, будет способствовать более интенсивным тренировкам, увеличению используемых весов и увеличению веса. Все они обладают очень сильным психическим эффектом, что является очень хорошей наградой после тяжелых тренировок.
Среди доказанных преимуществ приема креатина — повышение общего количества креатина в организме, увеличение мышечной массы, увеличение веса в результате увеличения безжировой массы тела, меньшая утомляемость и большая выносливость во время интенсивных упражнений, а также улучшение когнитивных способностей. , имеет потенциальные преимущества при болезни Паркинсона и Хантингтона. Креатин помогает при мышечной дистрофии и в сочетании с тренировками может снизить уровень ЛПНП в крови.
, имеет потенциальные преимущества при болезни Паркинсона и Хантингтона. Креатин помогает при мышечной дистрофии и в сочетании с тренировками может снизить уровень ЛПНП в крови.
Преимущества моногидрата креатина
Моногидрат креатина имеет несколько преимуществ перед другими формами.Во-первых, почти все когда-либо проводились исследования моногидрата креатина. Это означает, что вышеупомянутые преимущества для УЗ полностью продемонстрированы. Кроме того, эта форма креатина полностью безопасна для вашего здоровья. Некоторые из других форм, доступных на рынке, разрешено продавать, но они в значительной степени неэффективны, а некоторые даже вредны. Кроме того, это самая дешевая форма, которая продается.
Суточные дозы креатина
Рекомендуется принимать от 3 до 5 граммов моногидрата креатина в день.Его можно пить в любое время дня — после тренировки между приемами пищи, утром, днем или вечером.
Единственное исключение — перед тренировкой — тогда его не следует принимать. Однако, поскольку его можно принимать в любое другое время, для спортсменов это не проблема.
Однако, поскольку его можно принимать в любое другое время, для спортсменов это не проблема.
Удобнее всего пить во время завтрака. Таким образом, маловероятно, что его забудут днем. Его можно принимать с кофе, соком, молоком, водой, протеиновым коктейлем и т. Д.
Что касается продолжительности приема, наиболее распространенной является практика приема периодов.Пример — два месяца приема, затем один месяц перерыва. Интервалы могут варьироваться в зависимости от планов спортсмена, но лучше всего, если период отдыха будет не менее 30 дней.
Источники креатина
Креатинможно купить отдельно или в составе других добавок. Он добавлен в различные протеиновые порошки, средства для отдыха, послетренировочные порошки, аминокислотные комплексы, гейнеры и многое другое.
Креатин естественным образом содержится в некоторых продуктах питания — в говядине, сельди и лососе очень высокая концентрация этого химического вещества.В 1 кг говядины содержится 4 грамма креатина.
Модуляция активности промотора мышечной креатинкиназы индуцибельным орфанным ядерным рецептором TIS1.
Biochem J. 1997, 15 января; 321 (Pt 2): 281–287.
Кафедра фармакологии, Университет Миссури-Колумбия 65212, США.
Abstract
TIS1, индуцибельный орфанный ядерный рецептор, первоначально был выделен в качестве гена, индуцируемого промотором опухоли в клетках 3T3 мыши, а позже было показано, что он индуцируется факторами роста и другими внеклеточными стимулами.Мы показываем здесь, что мРНК TIS1 экспрессировалась в пролиферирующих клетках скелетных мышц мышей C2C12, из того, что уровень экспрессии TIS1 увеличивался во время дифференцировки мышц. Сверхэкспрессия репортерных генов трансактивированной мышечной креатинкиназы (MCK), содержащих всего 80 п.н. проксимальной 5′-фланкирующей области. Напротив, конструкция TIS1 без промотора и конструкция TIS1 с мутантом со сдвигом рамки считывания были неспособны трансактивировать репортерный ген MCK. Более того, эффект, оказываемый TIS1, оказался селективным в отношении промотора MCK. Обработка клеток C2C12 форсколином, который, как известно, индуцирует экспрессию TIS1, также стимулировала активность репортерного гена MCK. Интересно, что транслированный in vitro белок TIS1 не смог связываться с промоторной областью MCK, что позволяет предположить, что эффект трансактивации TIS1 может быть опосредован без прямого взаимодействия белка с ДНК промотора MCK. В совокупности эти результаты предполагают, что изменение уровней TIS1 может помочь модулировать экспрессию MCK и, возможно, других специфичных для мышц генов в ответ на физиологические изменения.
Обработка клеток C2C12 форсколином, который, как известно, индуцирует экспрессию TIS1, также стимулировала активность репортерного гена MCK. Интересно, что транслированный in vitro белок TIS1 не смог связываться с промоторной областью MCK, что позволяет предположить, что эффект трансактивации TIS1 может быть опосредован без прямого взаимодействия белка с ДНК промотора MCK. В совокупности эти результаты предполагают, что изменение уровней TIS1 может помочь модулировать экспрессию MCK и, возможно, других специфичных для мышц генов в ответ на физиологические изменения.
Полный текст
Полный текст этой статьи доступен в формате PDF (316 КБ).
Избранные ссылки
Эти ссылки находятся в PubMed. Это может быть не полный список ссылок из этой статьи.
- Lim RW, Варнум БК, Herschman HR. Клонирование последовательностей «первичного ответа», индуцированных сложным эфиром тетрадеканоилфорбола, и их экспрессия в клетках Swiss 3T3 с задержкой плотности и непролиферативном варианте TPA. Онкоген. 1987. 1 (3): 263–270. [PubMed] [Google Scholar]
- Хейзел Т.Г., Натанс Д., Лау Л.Ф.Ген, индуцируемый факторами роста сыворотки, кодирует член суперсемейства рецепторов стероидов и гормонов щитовидной железы. Proc Natl Acad Sci U S. A. 1988, ноябрь; 85 (22): 8444–8448. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ryseck RP, Macdonald-Bravo H, Mattéi MG, Ruppert S, Bravo R. Структура, картирование и экспрессия гена, индуцируемого фактором роста, кодирующего предполагаемый ядерный гормональный рецептор. EMBO J. 1989, ноябрь; 8 (11): 3327–3335. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хейзел Т.Г., Мисра Р., Дэвис И.Дж., Гринберг М.Э., Лау Л.Ф.Nur77 дифференцированно модифицируется в клетках PC12 после деполяризации мембраны и лечения факторами роста. Mol Cell Biol. 1991 июн; 11 (6): 3239–3246. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Yoon JK, Lau LF. Активация транскрипции гена индуцибельного ядерного рецептора nur77 фактором роста нервов и деполяризация мембраны в клетках PC12. J Biol Chem. 1993 25 апреля; 268 (12): 9148–9155. [PubMed] [Google Scholar]
- Morgan JI, Curran T. Связь между стимулом и транскрипцией в нервной системе: участие индуцибельных протоонкогенов fos и jun.Annu Rev Neurosci. 1991; 14: 421–451. [PubMed] [Google Scholar]
- Милбрандт Дж. Фактор роста нервов индуцирует ген, гомологичный гену рецептора глюкокортикоидов. Нейрон. 1988 Май; 1 (3): 183–188. [PubMed] [Google Scholar]
- Дэвис И.Дж., Хейзел Т.Г., Лау Л.Ф. Активация транскрипции с помощью Nur77, члена суперсемейства рецепторов стероидных гормонов, индуцируемого фактором роста. Мол Эндокринол. 1991 июн; 5 (6): 854–859. [PubMed] [Google Scholar]
- Уилсон Т.Э., Фарнер Т.Дж., Джонстон М., Милбрандт Дж.Идентификация сайта связывания ДНК для NGFI-B путем генетической селекции у дрожжей. Наука. 1991 31 мая; 252 (5010): 1296–1300. [PubMed] [Google Scholar]
- Paulsen RE, Weaver CA, Fahrner TJ, Milbrandt J. Домены, регулирующие транскрипционную активность индуцибельного сиротского рецептора NGFI-B. J Biol Chem. 1992 15 августа; 267 (23): 16491–16496. [PubMed] [Google Scholar]
- Типпеттс М.Т., Варнум Британская Колумбия, Лим Р.В., Хершман Х.Р. Гены, индуцируемые промотором опухоли, по-разному экспрессируются в развивающихся мышах.Mol Cell Biol. 1988 Октябрь; 8 (10): 4570–4572. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Накаи А., Карта С., Сакураи А., Тобак Ф. Г., ДеГрут Л. Дж.. Ген раннего ответа человека, гомологичный мышиному nur77 и крысиному NGFI-B и относящийся к суперсемейству ядерных рецепторов. Мол Эндокринол. 1990 Октябрь; 4 (10): 1438–1443. [PubMed] [Google Scholar]
- Scearce LM, Laz TM, Hazel TG, Lau LF, Taub R. RNR-1, ядерный рецептор семейства NGFI-B / Nur77, который быстро индуцируется при регенерации печени. J Biol Chem.1993 25 апреля; 268 (12): 8855–8861. [PubMed] [Google Scholar]
- Wilson TE, Mouw AR, Weaver CA, Milbrandt J, Parker KL. Рецептор орфанного ядра NGFI-B регулирует экспрессию гена, кодирующего стероидную 21-гидроксилазу. Mol Cell Biol. 1993 Февраль; 13 (2): 861–868. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lim RW, Zhu CY, Stringer B. Дифференциальная регуляция экспрессии генов первичного ответа в клетках скелетных мышц посредством множественных путей передачи сигнала. Biochim Biophys Acta.1995 6 апреля; 1266 (1): 91–100. [PubMed] [Google Scholar]
- Лим Р.В., Ян В.Л., Ю.Х. Десенсибилизация экспрессии орфанного ядерного рецептора TIS1, специфичная для пути передачи сигнала. Biochem J. 1995, 15 июня; 308 (Pt 3): 785–789. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Джейнс Дж. Б., Джонсон Дж. Э., Бускин Дж. Н., Гартсайд К. Л., Хаушка С. Д.. Ген мышечной креатинкиназы регулируется множеством вышестоящих элементов, включая специфический для мышц усилитель. Mol Cell Biol. 1988, январь; 8 (1): 62–70. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Дэвис Р.Л., Вайнтрауб Х., Лассар А.Б.Экспрессия одной трансфицированной кДНК превращает фибробласты в миобласты. Клетка. 1987 24 декабря; 51 (6): 987–1000. [PubMed] [Google Scholar]
- Харпольд М.М., Эванс Р.М., Салдит-Джорджифф М., Дарнелл Дж. Э. Производство мРНК в клетках китайского хомячка: зависимость скорости синтеза от цитоплазматической концентрации девяти конкретных последовательностей мРНК. Клетка. 1979 августа; 17 (4): 1025–1035. [PubMed] [Google Scholar]
- Хомчинский П., Сакки Н. Одностадийный метод выделения РНК экстракцией кислотным тиоцианатом гуанидиния, фенолом и хлороформом.Анальная биохимия. 1987 Апрель; 162 (1): 156–159. [PubMed] [Google Scholar]
- Nishimoto I, Murayama Y, Katada T., Ui M, Ogata E. Возможная прямая связь рецептора инсулиноподобного фактора роста II с белками, связывающими гуанин-нуклеотид. J Biol Chem. 1989 25 августа; 264 (24): 14029–14038. [PubMed] [Google Scholar]
- Уотсон М.А., Милбрандт Дж. Ген NGFI-B, транскрипционно индуцибельный член суперсемейства генов стероидных рецепторов: геномная структура и экспрессия в мозге крысы после индукции судорожного припадка.Mol Cell Biol. 1989 Октябрь; 9 (10): 4213–4219. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Fontaine B, Klarsfeld A, Hökfelt T, Changeux JP. Связанный с геном кальцитонина пептид, пептид, присутствующий в мотонейронах спинного мозга, увеличивает количество рецепторов ацетилхолина в первичных культурах мышечных трубок куриного эмбриона. Neurosci Lett. 1986, 30 октября; 71 (1): 59–65. [PubMed] [Google Scholar]
- New HV, Mudge AW. Пептид, родственный гену кальцитонина, регулирует синтез мышечных рецепторов ацетилхолина.Природа. 323 (6091): 809–811. [PubMed] [Google Scholar]
- Силинский Э.М. О связи между секрецией медиатора и высвобождением адениновых нуклеотидов из нервных окончаний двигательных нервов млекопитающих. J Physiol. 1975 Май; 247 (1): 145–162. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Джейнс Дж. Б., Чемберлен Дж. С., Бускин Дж. Н., Джонсон Дж. Э., Хаушка С. Д.. Транскрипционная регуляция гена мышечной креатинкиназы и регулируемая экспрессия в трансфицированных миобластах мыши. Mol Cell Biol. 1986 август; 6 (8): 2855–2864.[Бесплатная статья PMC] [PubMed] [Google Scholar]
- Schwarz JJ, Chakraborty T., Martin J, Zhou JM, Olson EN. Основная область миогенина взаимодействует с двумя доменами активации транскрипции, чтобы индуцировать специфичную для мышц транскрипцию. Mol Cell Biol. 1992 Янв; 12 (1): 266–275. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sternberg EA, Spizz G, Perry WM, Vizard D, Weil T., Olson EN. Идентификация вышестоящих и внутригенных регуляторных элементов, которые обеспечивают экспрессию гена мышечной креатинкиназы, ограниченную типом клеток и специфичную для дифференцировки.Mol Cell Biol. Июль 1988 г .; 8 (7): 2896–2909. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тапскотт С.Дж., Вайнтрауб Х. MyoD и регуляция миогенеза с помощью белков спираль-петля-спираль. J Clin Invest. 1991 апр; 87 (4): 1133–1138. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Вайнтрауб Х., Дэвис Р., Локшон Д., Лассар А. MyoD кооперативно связывается с двумя сайтами в целевой последовательности энхансера: для активации требуется занятость двух сайтов. Proc Natl Acad Sci U S. A., 1990, август; 87 (15): 5623–5627.[Бесплатная статья PMC] [PubMed] [Google Scholar]
- Weintraub H, Dwarki VJ, Verma I, Davis R, Hollenberg S, Snider L, Lassar A, Tapscott SJ. Активация транскрипции, специфичная для мышц, MyoD. Genes Dev. 1991 август; 5 (8): 1377–1386. [PubMed] [Google Scholar]
- Райт В.Е., Сассун Д.А., Лин В.К. Миогенин, фактор, регулирующий миогенез, имеет домен, гомологичный MyoD. Клетка. 1989 24 февраля; 56 (4): 607–617. [PubMed] [Google Scholar]
- Адамсон Э.Д. Онкогены в разработке. Разработка.1987 апр; 99 (4): 449–471. [PubMed] [Google Scholar]
- Ковари К., Браво Р. Существование различных комплексов Fos / Jun во время перехода от G0 к G1 и во время экспоненциального роста фибробластов мыши: дифференциальная роль белков Fos. Mol Cell Biol. 1992 ноя; 12 (11): 5015–5023. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Hai T., Curran T. Межсемейная димеризация транскрипционных факторов Fos / Jun и ATF / CREB изменяет специфичность связывания ДНК. Proc Natl Acad Sci U S. A. 1991, 1 мая; 88 (9): 3720–3724.[Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kerppola TK, Curran T. Гетеродимеры Fos-Jun и гомодимеры Jun изгибают ДНК в противоположных направлениях: последствия для кооперативности факторов транскрипции. Клетка. 26 июля 1991 г .; 66 (2): 317–326. [PubMed] [Google Scholar]
- Шариф М., Привальский М.Л. Белки V-erbA и c-erbA усиливают активацию транскрипции c-jun. Онкоген. 1992 Май; 7 (5): 953–960. [PubMed] [Google Scholar]
- Stein B, Cogswell PC, Baldwin AS., Jr. Функциональные и физические ассоциации между членами семейства NF-kappa B и C / EBP: взаимодействие между доменом Rel и bZIP.Mol Cell Biol. Июль 1993 г .; 13 (7): 3964–3974. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Сукович Д.А., Мукерджи Р., Бенфилд, Пенсильвания. Новый, специфичный для клеточного типа механизм активации гена, опосредованной рецептором эстрогена, в отсутствие элемента, чувствительного к эстрогену. Mol Cell Biol. 1994 ноя; 14 (11): 7134–7143. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Baniahmad A, Tsai MJ. Механизмы активации транскрипции рецепторами стероидных гормонов. J Cell Biochem. 1993 Февраль; 51 (2): 151–156.[PubMed] [Google Scholar]
- Молькентин Дж. Д., Блэк Б. Л., Мартин Дж. Ф., Олсон Э. Н.. Совместная активация экспрессии мышечных генов с помощью MEF2 и миогенных белков bHLH. Клетка. 1995, 29 декабря; 83 (7): 1125–1136. [PubMed] [Google Scholar]
- Muscat GE, Mynett-Johnson L, Dowhan D, Downes M, Griggs R. Активация транскрипции гена myoD 3,5,3′-трийод-L-тиронином: прямая роль гормоны щитовидной железы и рецепторы ретиноидов X. Nucleic Acids Res. 1994 25 февраля; 22 (4): 583–591. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Downes M, Griggs R, Atkins A, Olson EN, Muscat GE.Идентификация элемента ответа на гормон щитовидной железы в гене миогенина мыши: характеристика гетеродимерного сайта связывания гормона щитовидной железы и рецептора ретиноида X. Рост клеток различается. 1993 ноябрь; 4 (11): 901–909. [PubMed] [Google Scholar]
 Онкоген. 1987. 1 (3): 263–270. [PubMed] [Google Scholar]
Онкоген. 1987. 1 (3): 263–270. [PubMed] [Google Scholar]Статьи из биохимического журнала любезно предоставлены Биохимическое общество
Что означает креатин? Бесплатный словарь
Супернатант использовали для анализа активности гексокиназы (HK), пируваткиназы (PK) и креатинкиназы (CK) с коммерческими наборами (Nanjing Jiancheng Bioengineering Institute, Китай).исследовали влияние низких и постоянных доз креатина и продемонстрировали, что приема низкой дозы (7,7 г / день) в течение 21 дня достаточно для повышения силы, потенции и устойчивости к утомлению у спортсменов мужского пола колледжа по сравнению с контрольной группой (14). .Таблица 1 содержит фазы лечения, измеряемые параметры, продолжительность приема креатина и время измерений Синдромы церебральной недостаточности креатина: клинические аспекты, лечение и патофизиология. (36) взаимодействие между кофеином и креатином может уменьшить снабжение креатином и фармакокинетика, которые могут отрицательно влиять на процесс синтеза белка, действие Caf в высвобождении саркоплазматического ретикулума кальция связано с хроническим истощением внутриклеточного кальция, которое изменяет процесс утомления, но нарушает синтез белка (46).Концентрация креатиназы была получена с использованием метода Брэдфорда (32), а активность креатиназы была измерена путем оценки количества образования мочевины в результате гидролиза креатина в колориметрическом анализе. После гибели рыбы также выделялись химические вещества N-соединений, состоящие из аминокислот, мочевая кислота, пурины, метиламин, таурин, имидазолы, креатин и креатинин на стадии постригора [19]. Даже не считая хорошо известных опасностей использования стероидов, креатин является одной из самых популярных добавок для набора веса в этой возрастной группе; в нескольких исследованиях сообщается о 8.8% и 21% атлетов старшеклассников признают употребление креатина. Влияние добавок креатина на состав тела и работоспособность: метаанализ. У людей, принимавших 400 мкг FA отдельно или в сочетании с креатином, уровень мышьяка в крови был аналогичен таковому. группы плацебо, (3) возможно потому, что у немногих участников исследования был дефицит фолиевой кислоты по сравнению со всеми участниками более ранней работы. Ключевые слова: гипоксическая ишемическая энцефалопатия (ГИЭ) моногидрат креатина интерлейкин-6 (ИЛ-6) интерлейкин-18 (IL-18) иммуноферментный анализ (ELISA).Самая низкая стоимость получения эквивалента 5 граммов высококачественного моногидрата креатина из добавки составляла 9 центов, а 5 граммов BCAA — 31 цент, в то время как стоимость получения тех же ингредиентов из некоторых других продуктов была намного выше 1 доллара. .Дифференциация и специфическая активность волокон интронного усилителя мышечной креатинкиназы | Skeletal Muscle
Welle S, Bhatt K, Thornton CA: Перечень мРНК с высоким содержанием мРНК в скелетных мышцах нормальных мужчин. Genome Res.1999, 9: 506-513.
PubMed Central CAS PubMed Google ученый
Чемберлен Дж. С., Джейнес Дж. Б., Хаушка С. Д.: Регулирование индукции креатинкиназы в дифференцирующихся миобластах мыши. Mol Cell Biol. 1985, 5: 484-492.
PubMed Central CAS Статья PubMed Google ученый
Tapscott SJ: Схема главного переключателя: Myod и регуляция транскрипции генов скелетных мышц.Разработка. 2005, 132: 2685-2695. 10.1242 / dev.01874.
CAS Статья PubMed Google ученый
Lyons GE, Muhlebach S, Moser A, Masood R, Paterson BM, Buckingham ME, Perriard JC: Регулирование развития экспрессии гена креатинкиназы миогенными факторами в скелетных мышцах эмбрионов мышей и кур. Разработка. 1991, 113: 1017-1029.
CAS PubMed Google ученый
Ямасита К., Йошиока Т. Профили изоферментных композиций креатинкиназы в отдельных мышечных волокнах разных типов. J Muscle Res Cell Motil. 1991, 12: 37-44. 10.1007 / BF01781172.
CAS Статья PubMed Google ученый
Кушмерик М.Дж., Мёрланд Т.С., Вайзман Р.В.: Волокна скелетных мышц млекопитающих, различающиеся по содержанию фосфокреатина, АТФ и Pi. Proc Natl Acad Sci USA. 1992, 89: 7521-7525. 10.1073 / pnas.89.16.7521.
PubMed Central CAS Статья PubMed Google ученый
Wentworth BM, Donoghue M, Engert JC, Berglund EB, Rosenthal N: парные MyoD-связывающие сайты регулируют экспрессию гена легкой цепи миозина. Proc Natl Acad Sci USA. 1991, 88: 1242-1246. 10.1073 / pnas.88.4.1242.
PubMed Central CAS Статья PubMed Google ученый
Salminen M, Lopez S, Maire P, Kahn A, Daegelen D: ДНК-белковые взаимодействия, специфичные для быстрых мышц, происходящие in vivo на промоторе альдолазы A M человека, необходимы для правильной активности промотора у трансгенных мышей. Mol Cell Biol. 1996, 16: 76-85.
PubMed Central CAS Статья PubMed Google ученый
Лупа-Кимбалл В.А., Эссер К.А.: Использование инъекции ДНК для идентификации медленных нервно-зависимых областей гена MLC2s.Am J Physiol. 1998, 274: C229-235.
CAS PubMed Google ученый
Esser K, Nelson T, Lupa-Kimball V, Blough E: САСС-бокс и сайты фактора-2 усилителя миоцитов в медленном промоторе легкой цепи миозина 2 взаимодействуют в регуляции нервно-специфической транскрипции в скелетных мышцах. J Biol Chem. 1999, 274: 12095-12102. 10.1074 / jbc.274.17.12095.
CAS Статья PubMed Google ученый
Vullhorst D, Buonanno A: Множественные GTF2I-подобные повторы общего фактора транскрипции 3 проявляют свойства связывания ДНК. Доказательства общего происхождения как модуля взаимодействия с последовательностью ДНК. J Biol Chem. 2005, 280: 31722-31731. 10.1074 / jbc.M500593200.
CAS Статья PubMed Google ученый
Рана З.А., Гундерсен К., Буонанно А: Взлеты и падения регуляции генов посредством электрической активности в скелетных мышцах.J Muscle Res Cell Motil. 2009, 30: 255-260. 10.1007 / s10974-010-9200-2.
PubMed Central Статья PubMed Google ученый
Issa LL, Палмер SJ, Guven KL, Santucci N, Hodgson VR, Popovic K, Joya JE, Hardeman EC: MusTRD может регулировать постнатальную экспрессию волокон. Dev Biol. 2006, 293: 104-115. 10.1016 / j.ydbio.2006.01.019.
CAS Статья PubMed Google ученый
Кальво С., Вулхорст Д., Венепалли П., Ченг Дж., Караванова И., Буонанно А. Молекулярное вскрытие последовательностей ДНК и факторов, участвующих в медленной мышечно-специфической транскрипции. Mol Cell Biol. 2001, 21: 8490-8503. 10.1128 / MCB.21.24.8490-8503.2001.
PubMed Central CAS Статья PubMed Google ученый
Халлауэр П.Л., Брэдшоу Х.Л., Гастингс К.Е.: Комплексная специфическая для типа волокна экспрессия генных конструкций тропонина I быстрых скелетных мышц у трансгенных мышей.Разработка. 1993, 119: 691-701.
CAS PubMed Google ученый
Hallauer PL, Hastings KE: TnIfast IRE-энхансер: многоступенчатая регуляция развития во время дифференциации типа волокон скелетных мышц. Dev Dyn. 2002, 224: 422-431. 10.1002 / dvdy.10122.
CAS Статья PubMed Google ученый
Бускин Дж. Н., Хаушка С. Д.: Идентификация ядерного фактора миоцитов, который связывается с мышечным энхансером гена мышечной креатинкиназы мыши.Mol Cell Biol. 1989, 9: 2627-2640.
PubMed Central CAS Статья PubMed Google ученый
Доновил Д.Б., Шилд М.А., Бускин Дж. Н., Хауген Х.С., Клегг С.Н., Хаушка С.Д.: Анализ регуляторных элементов гена креатинкиназы в мышцах в скелетных и сердечных мышцах трансгенных мышей. Mol Cell Biol. 1996, 16: 1649-1658.
PubMed Central CAS Статья PubMed Google ученый
Джейнс Дж. Б., Чемберлен Дж. С., Бускин Дж. Н., Джонсон Дж., Хаушка С. Д.: Транскрипционная регуляция гена креатинкиназы в мышцах и регулируемая экспрессия в трансфицированных миобластах мыши. Mol Cell Biol. 1986, 6: 2855-2864.
PubMed Central CAS Статья PubMed Google ученый
Jaynes JB, Johnson JE, Buskin JN, Gartside CL, Hauschka SD: Ген мышечной креатинкиназы регулируется множеством вышестоящих элементов, включая специфический для мышц энхансер.Mol Cell Biol. 1988, 8: 62-70.
PubMed Central CAS Статья PubMed Google ученый
Johnson JE, Gartside CL, Jaynes JB, Hauschka SD: Экспрессия трансфицированного гена мышечной креатинкиназы мыши индуцируется при лишении фактора роста миогенных, но не немиогенных клеток. Dev Biol. 1989, 134: 258-262. 10.1016 / 0012-1606 (89)
CAS Статья PubMed Google ученый
Johnson JE, Wold BJ, Hauschka SD: Элементы последовательности мышечной креатинкиназы, регулирующие экспрессию скелетных и сердечных мышц у трансгенных мышей. Mol Cell Biol. 1989, 9: 3393-3399.
PubMed Central CAS Статья PubMed Google ученый
Amacher SL, Buskin JN, Hauschka SD: Множественные регуляторные элементы по-разному влияют на активность усилителя мышечной креатинкиназы в скелетных и сердечных мышцах.Mol Cell Biol. 1993, 13: 2753-2764.
PubMed Central CAS Статья PubMed Google ученый
Himeda CL, Ranish JA, Angello JC, Maire P, Aebersold R, Hauschka SD: Количественная протеомная идентификация six4 как фактора связывания trex в усилителе креатинкиназы мышц. Mol Cell Biol. 2004, 24: 2132-2143. 10.1128 / MCB.24.5.2132-2143.2004.
PubMed Central CAS Статья PubMed Google ученый
Nguyen QG, Buskin JN, Himeda CL, Fabre-Suver C, Hauschka SD: Трансгенные и тканевые анализы контролирующего элемента Trex усилителя креатинкиназы мышц в скелетных и сердечных мышцах указывают на различия в экспрессии генов между типами мышц. Transgenic Res. 2003, 12: 337-349. 10.1023 / А: 1023369225799.
CAS Статья PubMed Google ученый
Nguyen QG, Buskin JN, Himeda CL, Shield MA, Hauschka SD: Различия в функции трех консервативных E-блоков гена креатинкиназы мышц в культивируемых миоцитах и в скелетных и сердечных мышцах трансгенных мышей.J Biol Chem. 2003, 278: 46494-46505. 10.1074 / jbc.M308194200.
CAS Статья PubMed Google ученый
Shield MA, Haugen HS, Clegg CH, Hauschka SD: E-box сайты и проксимальная регуляторная область гена мышечной креатинкиназы по-разному регулируют экспрессию в различных скелетных мышцах и сердечных мышцах трансгенных мышей. Mol Cell Biol. 1996, 16: 5058-5068.
PubMed Central CAS Статья PubMed Google ученый
Mueller PR, Wold B: Поступление in vivo мышечно-специфичного энхансера с помощью лигирования опосредованной ПЦР. Наука. 1989, 246: 780-786. 10.1126 / science.2814500.
CAS Статья PubMed Google ученый
Джейнс Дж .: Регуляция транскрипции гена мышечной креатинкиназы. Диссертация. 1986, Вашингтонский университет, факультет биохимии
Google ученый
Tamir Y, Bengal E: белок p53 активируется во время дифференцировки мышц и вместе с MyoD участвует в транскрипции гена мышечной креатинкиназы. Онкоген. 1998, 17: 347-356. 10.1038 / sj.onc.1201929.
CAS Статья PubMed Google ученый
Винсент К.К., Гуальберто А., Патель К.В., Уолш К. Различные регуляторные последовательности контролируют экспрессию гена креатинкиназы-М в скелетных и сердечных мышцах, непосредственно подвергнутых инъекции.Mol Cell Biol. 1993, 13: 1264-1272.
PubMed Central CAS Статья PubMed Google ученый
Himeda CL, Ranish JA, Hauschka SD: Количественная протеомная идентификация MAZ как регулятора транскрипции специфичных для мышц генов в скелетных и сердечных миоцитах. Mol Cell Biol. 2008, 28: 6521-6535. 10.1128 / MCB.00306-08.
PubMed Central CAS Статья PubMed Google ученый
Himeda CL, Ranish JA, Pearson RCM, Crossley M, Hauschka SD: KLF3 регулирует экспрессию генов, специфичных для мышц, и взаимодействует с SRF на сайтах связывания KLF. Молекулярная клеточная биология (в обзоре). 2010
Google ученый
Dunant P, Larochelle N, Thirion C, Stucka R, Ursu D, Petrof BJ, Wolf E, Lochmuller H: Экспрессия дистрофина, управляемая промотором MCK 1,35 kb, быстро улучшает мышечную дистрофию, но не медленные мышцы трансгенных мышей MDX.Mol Ther. 2003, 8: 80-89. 10.1016 / S1525-0016 (03) 00129-1.
CAS Статья PubMed Google ученый
Salva MZ, Himeda CL, Tai PW, Nishiuchi E, Gregorevic P, Allen JM, Finn EE, Nguyen QG, Blankinship MJ, Meuse L, et al: Дизайн тканеспецифичных регуляторных кассет для высокого уровня rAAV-опосредованная экспрессия в скелетных и сердечных мышцах. Mol Ther. 2007, 15: 320-329. 10.1038 / sj.mt.6300027.
CAS Статья PubMed Google ученый
Eppenberger HM, Eppenberger M, Richterich R, Aebi H: Онтогенез изоферментов креатинкиназы. Dev Biol. 1964, 10: 1-16. 10.1016 / 0012-1606 (64)
-8.
CAS Статья PubMed Google ученый
Cox GA, Cole NM, Matsumura K, Phelps SF, Hauschka SD, Campbell KP, Faulkner JA, Chamberlain JS: Сверхэкспрессия дистрофина у трансгенных мышей mdx устраняет симптомы дистрофии без токсичности. Природа. 1993, 364: 725-729.10.1038 / 364725a0.
CAS Статья PubMed Google ученый
Sternberg EA, Spizz G, Perry WM, Vizard D, Weil T., Olson EN: Идентификация вышестоящих и внутригенных регуляторных элементов, которые обеспечивают ограниченную по типу клеток и специфичную для дифференцировки экспрессию гена мышечной креатинкиназы. Mol Cell Biol. 1988, 8: 2896-2909.
PubMed Central CAS Статья PubMed Google ученый
Polly P, Haddadi LM, Issa LL, Subramaniam N, Palmer SJ, Tay ES, Hardeman EC: hMusTRD1alpha1 репрессирует активацию MEF2 медленного энхансера тропонина I. J Biol Chem. 2003, 278: 36603-36610. 10.1074 / jbc.M212814200.
CAS Статья PubMed Google ученый
Wingender E: Проект TRANSFAC как пример рамочной технологии, которая поддерживает анализ геномной регуляции. Краткий биоинформ. 2008, 9: 326-332.10.1093 / bib / bbn016.
CAS Статья PubMed Google ученый
Wright WE, Binder M, Funk W: Циклическая амплификация и отбор мишеней (CASTing) для консенсусного сайта связывания миогенина. Mol Cell Biol. 1991, 11: 4104-4110.
PubMed Central CAS Статья PubMed Google ученый
Blackwell TK, Weintraub H: различия и сходства в предпочтениях связывания ДНК комплексов белков MyoD и E2A, выявленные путем выбора сайта связывания.Наука. 1990, 250: 1104-1110. 10.1126 / science.2174572.
CAS Статья PubMed Google ученый
Фикетт JW: Количественная дискриминация сайтов MEF2. Mol Cell Biol. 1996, 16: 437-441.
PubMed Central CAS Статья PubMed Google ученый
van Dam H, Castellazzi M: Различная роль димеров Jun: Fos и Jun: ATF в онкогенезе.Онкоген. 2001, 20: 2453-2464. 10.1038 / sj.onc.1204239.
CAS Статья PubMed Google ученый
Berkes CA, Tapscott SJ: MyoD и транскрипционный контроль миогенеза. Semin Cell Dev Biol. 2005, 16: 585-595. 10.1016 / j.semcdb.2005.07.006.
CAS Статья PubMed Google ученый
Молкентин Дж. Д., Блэк Б. Л., Мартин Дж. Ф., Олсон Е. Н.: Совместная активация экспрессии мышечных генов с помощью MEF2 и миогенных белков bHLH.Клетка. 1995, 83: 1125-1136. 10.1016 / 0092-8674 (95)
CAS Статья PubMed Google ученый
Игараси К., Ито К., Мотохаши Н., Хаяси Н., Матудзаки Ю., Накаучи Н., Нишизава М., Ямамото М.: Активность и экспрессия мышиного малого белка семейства Maf MafK. J Biol Chem. 1995, 270: 7615-7624. 10.1074 / jbc.270.13.7615.
CAS Статья PubMed Google ученый
Toki T, Itoh J, Kitazawa J, Arai K, Hatakeyama K, Akasaka J, Igarashi K, Nomura N, Yokoyama M, Yamamoto M, Ito E: маленькие белки Maf человека образуют гетеродимеры с факторами транскрипции семейства CNC и распознают NF- Мотив E2. Онкоген. 1997, 14: 1901-1910. 10.1038 / sj.onc.1201024.
CAS Статья PubMed Google ученый
Miskimins R, Miskimins WK: роль AP-1-подобного сайта в экспрессии гена основного белка миелина во время дифференцировки.Int J Dev Neurosci. 2001, 19: 85-91. 10.1016 / S0736-5748 (00) 00066-6.
CAS Статья PubMed Google ученый
Zimprich A, Kraus J, Woltje M, Mayer P, Rauch E, Hollt V: Аллельная вариация в промоторе гена продинорфина человека изменяет индуцированную стимулом экспрессию. J Neurochem. 2000, 74: 472-477. 10.1046 / j.1471-4159.2000.740472.x.
CAS Статья PubMed Google ученый
Ohkawa Y, Marfella CG, Imbalzano AN: Спецификация скелетных мышц миогенином и Mef2D через SWI / SNF-АТФазу Brg1. Эмбо Дж. 2006, 25: 490-501. 10.1038 / sj.emboj.7600943.
PubMed Central CAS Статья PubMed Google ученый
Cao Y, Yao Z, Sarkar D, Lawrence M, Sanchez GJ, Parker MH, MacQuarrie KL, Davison J, Morgan MT, Ruzzo WL и др.: Связывание MyoD по всему геному в клетках скелетных мышц: a потенциал для широкого клеточного перепрограммирования.Dev Cell. 2010, 18: 662-674. 10.1016 / j.devcel.2010.02.014.
PubMed Central CAS Статья PubMed Google ученый
Блэк Б.Л., Олсон EN: Транскрипционный контроль развития мышц с помощью белков фактора-2 энхансера миоцитов (MEF2). Annu Rev Cell Dev Biol. 1998, 14: 167-196. 10.1146 / annurev.cellbio.14.1.167.
CAS Статья PubMed Google ученый
Lakich MM, Diagana TT, North DL, Whalen RG: MEF-2 и Oct-1 связываются с двумя гомологичными элементами промоторной последовательности и участвуют в экспрессии гена, специфичного для скелетных мышц. J Biol Chem. 1998, 273: 15217-15226. 10.1074 / jbc.273.24.15217.
CAS Статья PubMed Google ученый
Карасева Н., Цика Дж., Джи Дж., Чжан А., Мао X, Цика Р.: Фактор усилителя транскрипции 1 связывает несколько мышечных MEF2 и A / T-богатые элементы во время перехода от быстрого к медленному типу волокон скелетных мышц. .Mol Cell Biol. 2003, 23: 5143-5164. 10.1128 / MCB.23.15.5143-5164.2003.
PubMed Central CAS Статья PubMed Google ученый
Блэк Б.Л., Молкентин Дж. Д., Олсон EN: Множественные роли основной области MyoD в передаче сигналов активации транскрипции и взаимодействии с MEF2. Mol Cell Biol. 1998, 18: 69-77.
PubMed Central CAS Статья PubMed Google ученый
Грегоревич П., Мезнарич Н.А., Бланкиншип М.Дж., Кроуфорд Р.В., Чемберлен Дж.С.: Меченные флуорофором миозин-специфические антитела упрощают фенотипирование мышечных волокон. Мышечный нерв. 2008, 37: 104-106. 10.1002 / mus.20877.
PubMed Central Статья PubMed Google ученый
Скьяффино С., Горца Л., Сарторе С., Саггин Л., Аусони С., Вианелло М., Гундерсен К., Ломо Т. Три изоформы тяжелых цепей миозина в волокнах скелетных мышц 2 типа.J Muscle Res Cell Motil. 1989, 10: 197-205. 10.1007 / BF01739810.
CAS Статья PubMed Google ученый
Kunarso G, Chia NY, Jeyakani J, Hwang C, Lu X, Chan YS, Ng HH, Bourque G: Мобильные элементы перестроили основную регуляторную сеть человеческих эмбриональных стволовых клеток. Нат Жене. 42: 631-634.
Schmidt D, Wilson MD, Ballester B, Schwalie PC, Brown GD, Marshall A, Kutter C, Watt S, Martinez-Jimenez CP, Mackay S, et al: ChIP-seq пяти позвоночных раскрывает эволюцию динамика связывания факторов транскрипции.Наука. 328: 1036-1040.
Mal AK: гистон-метилтрансфераза Suv39h2 репрессирует миогенную дифференцировку, стимулированную MyoD. Эмбо Дж. 2006, 25: 3323-3334. 10.1038 / sj.emboj.7601229.
PubMed Central CAS Статья PubMed Google ученый
Родс С.Дж., Конечны С.Ф .: Идентификация MRF4: нового члена семейства генов фактора регуляции мышц. Genes Dev. 1989, 3: 2050-2061. 10.1101 / гад.3.12b.2050.
CAS Статья PubMed Google ученый
Hinterberger TJ, Sassoon DA, Rhodes SJ, Konieczny SF: Экспрессия фактора регуляции мышц MRF4 во время развития сомитов и скелетных миофибрилл. Dev Biol. 1991, 147: 144-156. 10.1016 / S0012-1606 (05) 80014-4.
CAS Статья PubMed Google ученый
Fullwood MJ, Ruan Y: Методы на основе ChIP для идентификации дальнодействующих взаимодействий хроматина.J Cell Biochem. 2009, 107: 30-39. 10.1002 / jcb.22116.
PubMed Central CAS Статья PubMed Google ученый
Олсон Э.Н., Перри М., Шульц Р.А.: Регуляция дифференцировки мышц семейством MEF2 факторов транскрипции MADS-бокса. Dev Biol. 1995, 172: 2-14. 10.1006 / dbio.1995.0002.
CAS Статья PubMed Google ученый
Morisaki T, Sermsuvitayawong K, Byun SH, Matsuda Y, Hidaka K, Morisaki H, Mukai T: Ген Mef2b мыши: уникальный член семейства генов MEF2. J Biochem. 1997, 122: 939-946.
CAS Статья PubMed Google ученый
Молкентин JD, Firulli AB, Black BL, Martin JF, Hustad CM, Copeland N, Jenkins N., Lyons G, Olson EN: MEF2B является мощным трансактиватором, экспрессирующимся в ранних миогенных клонах. Mol Cell Biol. 1996, 16: 3814-3824.
PubMed Central CAS Статья PubMed Google ученый
Chang PS, Li L, McAnally J, Olson EN: мышечная специфичность, кодируемая сайтами связывания специфических сывороточных факторов ответа. J Biol Chem. 2001, 276: 17206-17212. 10.1074 / jbc.M010983200.
CAS Статья PubMed Google ученый
Cserjesi P, Lilly B, Bryson L, Wang Y, Sassoon DA, Olson EN: MHox: мезодермально ограниченный гомеодоменный белок, который связывает важный сайт в усилителе креатинкиназы в мышцах.Разработка. 1992, 115: 1087-1101.
CAS PubMed Google ученый
Cserjesi P, Lilly B, Hinkley C, Perry M, Olson EN: гомеодоменный белок MHox и MADS-белок, усиливающий фактор связывания миоцитов-2, сходятся на общем элементе в усилителе мышечной креатинкиназы. J Biol Chem. 1994, 269: 16740-16745.
CAS PubMed Google ученый
Gossett LA, Kelvin DJ, Sternberg EA, Olson EN: новый фактор связывания энхансера, специфичный для миоцитов, который распознает консервативный элемент, связанный с множеством специфичных для мышц генов.Mol Cell Biol. 1989, 9: 5022-5033.
PubMed Central CAS Статья PubMed Google ученый
Amacher SL: Регулирующие элементы миокардиоцитов и транс-действующие факторы гена мышечной креатинкиназы. 1993, Вашингтонский университет, биохимия
Google ученый
Tsika RW, Schramm C, Simmer G, Fitzsimons DP, Moss RL, Ji J: Сверхэкспрессия TEAD-1 в поперечно-полосатых мышцах трансгенных мышей вызывает более медленный сократительный фенотип скелетных мышц.J Biol Chem. 2008, 283: 36154-36167. 10.1074 / jbc.M807461200.
PubMed Central CAS Статья PubMed Google ученый
Watchko JF, Daood MJ, LaBella JJ: Активность креатинкиназы в скелетных мышцах крысы связана с фенотипом миозина во время развития. Pediatr Res. 1996, 40: 53-58. 10.1203 / 00006450-199607000-00010.
CAS Статья PubMed Google ученый
Вентура-Клапир Р., Кузнецов А.В., д’Албис А., Ван Дерсен Дж., Виринга Б., Векслер В.И.: Мыши с дефицитом креатинкиназы. I. Нарушения миофибриллярной функции. J Biol Chem. 1995, 270: 19914-19920. 10.1074 / jbc.270.34.19914.
CAS Статья PubMed Google ученый
Парк С.К., Гунаван А.М., Шеффлер Т.Л., Грант А.Л., Джеррард Д.Е.: Содержание изоформы тяжелой цепи миозина и энергетический метаболизм могут быть разделены в скелетных мышцах свиней.J Anim Sci. 2009, 87: 522-531.
CAS Статья PubMed Google ученый
Spitz F, Salminen M, Demignon J, Kahn A, Daegelen D, Maire P: Комбинация белков MEF3 и NFI активирует транскрипцию в подмножестве быстро сокращающихся мышц. Mol Cell Biol. 1997, 17: 656-666.
PubMed Central CAS Статья PubMed Google ученый
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, et al: Clustal W и Clustal × версия 2.0. Биоинформатика. 2007, 23: 2947-2948. 10.1093 / биоинформатика / btm404.
CAS Статья PubMed Google ученый
Clegg CH, Linkhart TA, Olwin BB, Hauschka SD: Контроль фактора роста дифференцировки скелетных мышц: приверженность терминальной дифференцировке происходит в фазе G1 и подавляется фактором роста фибробластов.J Cell Biol. 1987, 105: 949-956. 10.1083 / jcb.105.2.949.
CAS Статья PubMed Google ученый
Нельсон Дж. Д., Денисенко О., Сова П., Бомштык К.: Быстрый анализ иммунопреципитации хроматина. Nucleic Acids Res. 2006, 34: e2-10.1093 / nar / gnj004.
PubMed Central Статья PubMed Google ученый
Dignam JD, Lebovitz RM, Roeder RG: Точная инициация транскрипции с помощью РНК-полимеразы II в растворимом экстракте из изолированных ядер млекопитающих.Nucleic Acids Res. 1983, 11: 1475-1489. 10.1093 / nar / 11.5.1475.
PubMed Central CAS Статья PubMed Google ученый
Bradford MM: Быстрый и чувствительный метод количественного определения количества белка в микрограммах, использующий принцип связывания белок-краситель. Анальная биохимия. 1976, 72: 248-254. 10.1016 / 0003-2697 (76)
-3.
CAS Статья PubMed Google ученый
Джонсон Д.С., Мортазави А., Майерс Р.М., Уолд Б.: Полногеномное картирование взаимодействий белок-ДНК in vivo. Наука. 2007, 316: 1497-1502. 10.1126 / science.1141319.
CAS Статья PubMed Google ученый
Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ, Pachter L: Сборка транскриптов и количественная оценка с помощью RNA-Seq выявляют неаннотированные транскрипты и переключение изоформ во время клетки дифференциация.Nat Biotechnol. 2010, 28: 511-515. 10.1038 / nbt.1621.
PubMed Central CAS Статья PubMed Google ученый
Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B: Картирование и количественная оценка транскриптомов млекопитающих с помощью RNA-Seq. Нат методы. 2008, 5: 621-628. 10.1038 / nmeth.1226.
CAS Статья PubMed Google ученый
Zhang Y, Liu T, Meyer CA, Eeckhoute J, Johnson DS, Bernstein BE, Nussbaum C, Myers RM, Brown M, Li W, Liu XS: модельный анализ ChIP-Seq (MACS) .Genome Biol. 2008, 9: R137-10.1186 / gb-2008-9-9-r137.
PubMed Central Статья PubMed Google ученый
Sambrook JFE, Maniatis T: Molecular Cloning: A Laboratory Manual. 1989, Колд-Спринг-Харбор: Лаборатория Колд-Спринг-Харбор, 2
Google ученый
Длительное лечение бета-блокаторами предотвращает хронические изменения системы креатинкиназы и лактатдегидрогеназы в сердце крыс после инфаркта миокарда
% PDF-1.7 % 1 0 объект > / Тип / Каталог / PageLabels> >> эндобдж 14 0 объект > эндобдж 2 0 obj > эндобдж 3 0 obj > транслировать Acrobat Distiller 7.0.5 для MacintoshUnknownElsevier2016-07-18T10: 51: 53 + 05: 302016-07-18T10: 51: 53 + 05: 302016-07-18T10: 51: 53 + 05: 30application / pdfdoi: 10.1016 / 0735- 1097 (95) 00458-0
Депрессия связана со снижением 25-гидроксивитамина D и повышением уровня паратироидного гормона у пожилых людей | Депрессивные расстройства | JAMA Psychiatry
Контекст Депрессия случайно была связана с изменением уровня 25-гидроксивитамина D [25 (OH) D] и паратироидного гормона (ПТГ), но эта связь никогда не изучалась систематически.
Цель Чтобы определить в большой когорте населения, существует ли связь между депрессией и измененными уровнями 25 (OH) D и ПТГ.
Дизайн Популяционное когортное исследование (Longitudinal Aging Study, Амстердам).
Участники Одна тысяча двести восемьдесят два жителя общины в возрасте от 65 до 95 лет.
Настройка Нидерланды.
Основная мера результата Депрессия измерялась с помощью самоотчетов (Центр эпидемиологических исследований — шкала депрессии) и диагностических интервью (Диагностический график интервью).Оценивали уровни 25 (OH) D и ПТГ. Также измерялись потенциально смешивающие факторы (например, возраст, пол, статус курения, индекс массы тела, количество хронических состояний и концентрация креатинина в сыворотке) и объясняющие факторы (например, время сбора данных, уровень урбанизации и физическая активность).
Результаты Уровни 25 (OH) D были на 14% ниже у 169 человек с малой депрессией и на 14% ниже у 26 человек с большим депрессивным расстройством по сравнению с уровнями у 1087 контрольных лиц ( P <.001). Уровни ПТГ были на 5% и 33% выше, соответственно ( P = 0,003). Тяжесть депрессии (Шкала депрессии Центра эпидемиологических исследований) была значительно связана со снижением уровней 25 (OH) D в сыворотке ( P = 0,03) и повышенными уровнями ПТГ в сыворотке ( P = 0,008).
Заключение Результаты этого крупного популяционного исследования показывают связь депрессивного статуса и тяжести со снижением уровня 25 (OH) D в сыворотке крови и повышением уровня ПТГ в сыворотке у пожилых людей.
Значительные депрессивные симптомы широко распространены у пожилых людей (13%) 1 и приводят к высокой заболеваемости и смертности. 2 Schneider et al. 3 предположили, что психические расстройства, в частности депрессия, могут быть связаны с низкими концентрациями 25-гидроксивитамина D [25 (OH) D] в сыворотке крови. Основные причины дефицита витамина D, такие как меньшее пребывание на солнце в результате снижения активности на свежем воздухе, различных привычек в жилище или одежде, а также снижение потребления витаминов 4 могут быть вторичными по отношению к депрессии, но депрессия также может быть следствием плохого статуса витамина D. .Более того, низкий уровень витамина D вызывает повышение уровня паратироидного гормона (ПТГ) в сыворотке крови. Первичный гиперпаратиреоз, в свою очередь, часто сопровождается депрессивными расстройствами, и настроение обычно нормализуется после лечения гиперпаратиреоза. 5 , 6
Нарушения сывороточного 25 (OH) D и ПТГ, хотя и встречаются часто, хорошо поддаются лечению, что может способствовать предотвращению депрессии. Поэтому удивительно, что исследований этих аномалий в отношении депрессии было немного.Было проведено всего несколько небольших исследований с противоречивыми данными у лиц с большим депрессивным расстройством (БДР). 3 -5,7 , 8 Насколько нам известно, связь аномалий 25 (OH) D и ПТГ с небольшой депрессией или тяжестью депрессии не оценивалась. Таким образом, настоящее исследование оценивает связь между большой и малой депрессией и тяжестью депрессии и уровнями 25 (OH) D и ПТГ в сыворотке крови в крупном популяционном исследовании пожилых людей.
Амстердамское лонгитюдное исследование старения — это продолжающееся когортное исследование предикторов и последствий изменений настроения, автономии и благополучия у стареющего населения в Нидерландах. Случайная выборка пожилых мужчин и женщин (возраст 55-85 лет), стратифицированная по возрасту, полу, урбанизации и ожидаемой 5-летней смертности, была взята из регистров населения 11 муниципалитетов в районах на западе, юге и к северо-востоку от Нидерландов.Всего в базовом обследовании (1992–1993 гг.) Приняли участие 3107 человек, 99% белых. Процедуры отбора проб и данных были подробно описаны ранее. 9 , 10
Настоящее исследование было ограничено участниками в возрасте 65 лет и старше (т. Е. Родившихся в течение или до 1930 г.) (n = 2525), которые участвовали в первом наблюдении в 1995-1996 гг. (N = 1720) и у которых могла быть взята кровь. (n = 1285) и уровни 25 (OH) D и ПТГ были доступны (n = 1282).Между первым и вторым циклами истощение (n = 805 [51,7%]) было в основном из-за смерти. Респонденты в 1995-1996 годах, которые не участвовали в процедуре забора крови, были старше ( P, <.000), чаще женщины ( P, = 0,003) и чаще страдали депрессией ( P = 0,008). . Информированное согласие было получено от всех респондентов, и исследование было одобрено комитетом по медицинской этике Vrije Universiteit Amsterdam, Амстердам, Нидерланды.
При первом наблюдении в 1995–1996 годах депрессивный статус и тяжесть депрессии оценивались с использованием шкалы Центра эпидемиологических исследований – депрессии (CES-D). 11 Лица, набравшие баллы выше общепринятого порогового значения для клинически значимых симптомов (оценка CES-D ≥16), были приглашены на диагностическое психиатрическое обследование с использованием расписания диагностического интервью. 12
Образцы крови были взяты и центрифугированы утром. Субъектам разрешалось есть чай и тосты, но не молочные продукты. Образцы сыворотки хранили при -25 ° C до проведения анализов в 1997–1998 годах. Повторение некоторых измерений в 2005 г. дало аналогичные результаты.Концентрация 25 (OH) D в сыворотке определялась с помощью анализа конкурентного связывания белков (Nichols Institute Diagnostics Inc, Сан-Хуан-Капистрано, Калифорния) с коэффициентом вариации между анализами 10% и нормальным диапазоном от 10 до 40 нг / мл ( чтобы преобразовать в наномоли на литр, умножьте на 2,496). Уровень ПТГ в сыворотке измеряли с помощью иммуно-радиометрического анализа (Incstar Corp, Стиллуотер, Миннесота), как описано Pluijm et al, 13 , с коэффициентом вариации между исследованиями 7% и нормальным диапазоном 0.7–7 пг / мл (чтобы преобразовать в нанограммы на литр, умножьте на 1).
Также были измерены потенциально мешающие факторы. Статус курения классифицировался как курящий или некурящий. Мы контролировали индекс массы тела (ИМТ), потому что Wortsman et al 14 показали, что повышение уровня витамина D 3 в сыворотке после пребывания на солнце было на 57% меньше у людей с ожирением по сравнению с пациентами, не страдающими ожирением. Наиболее распространенные хронические соматические заболевания, включая инсульт, рак, заболевания легких, сердечные заболевания, сахарный диабет, остеоартрит и заболевание периферических артерий, были измерены с использованием самоотчетов и данных врачей общей практики. 15 Поскольку легкая почечная недостаточность может повлиять на гомеостаз витамина D и ПТГ, мы скорректировали концентрацию креатинина в сыворотке.
Было определено несколько возможных объясняющих факторов, которые могут объяснить связь между депрессией и уровнями витамина D и ПТГ. Поскольку статус витамина D частично зависит от воздействия солнечного света, регистрировали, собирались ли данные зимой, весной, летом или осенью. Кроме того, уровни физической активности (общая активность или мероприятия на свежем воздухе только в минутах в день) и урбанизация (количество жителей на квадратный километр) оценивались, как описано Snijder et al. 10 Использование антидепрессантов было определено, потому что Zhou et al. 16 описали усиление катаболизма витамина D с помощью лекарств.
Все анализы были выполнены с использованием имеющегося в продаже программного обеспечения (SPSS для Windows версии 14.0; SPSS Inc, Чикаго, Иллинойс). χ 2 Были проведены тесты для определения различий в возрасте, поле, ИМТ, статусе курения, физической активности, количестве хронических состояний, концентрации креатинина в сыворотке, сезоне, когда были собраны данные, и уровне урбанизации между людьми, не страдающими депрессией, небольшой депрессией. , или MDD.Был проведен множественный линейный регрессионный анализ для изучения связи между переменными депрессии (независимые переменные) и уровнями 25 (OH) D или ПТГ (зависимые переменные) в сыворотке крови. Поскольку распределение сывороточного ПТГ было искажено, мы использовали логарифмически преобразованные значения. Кроме того, мы скорректировали регрессионные модели для потенциальных мешающих переменных и, на отдельном этапе, для тех потенциальных объясняющих переменных, которые были однозначно связаны с депрессивным статусом. Возможное изменение эффекта в зависимости от пола было исследовано путем дополнительного включения в скорректированный регрессионный анализ продуктовых терминов, состоящих из уровней гормонов × пол.
У 26 человек был текущий (1 месяц назад) диагноз БДР в соответствии с графиком диагностического интервью, у 169 была легкая депрессия (по шкале CES-D ≥16, но график диагностического интервью отрицательный для БДР), и 1087 человек были не подавлен (таблица 1). По сравнению с людьми без депрессии, депрессивные люди были значительно ( P <0,05) старше, чаще были женщинами, чаще курили (только MDD), имели более высокий ИМТ (только MDD), имели больше хронических заболеваний и чаще жили в сильно урбанизированных районах, что подчеркивает необходимость учета этих факторов при анализе.Концентрация креатинина, сезон оценки и уровни физической активности существенно не различались между депрессивными и не депрессивными людьми. Только 1% участников принимали низкие дозы витамина D или кальция. Судя по уровням 25 (OH) D и ПТГ в сыворотке крови, это не повлияло на наши результаты. Менее 1% участников (т.е. 7 человек с легкой депрессией и 4 с БДР) использовали антидепрессанты, которые не повлияли на наши результаты при добавлении в модель (данные не показаны).
Средний (SD) уровень 25 (OH) D в сыворотке был 21 (10) нг / мл (рисунок, A), а средний уровень ПТГ в сыворотке был 3.6 (1,7) пг / мл (рисунок, Б). Коэффициент корреляции Пирсона между уровнями 25 (OH) D и ПТГ в сыворотке составлял -0,30 ( P <0,001). У 4,5% мужчин и 7,7% женщин уровень 25 (OH) D в сыворотке был менее 10 нг / мл, что указывало на состояние дефицита, определенное Липсом. 17 У 38,8% мужчин и 56,9% женщин уровень 25 (OH) D в сыворотке был менее 20 нг / мл, что указывает на недостаточный статус.
Уровни 25 (OH) D были на 14% ниже у 169 человек с небольшой депрессией (среднее значение 19 нг / мл) и на 14% ниже у 26 человек с БДР (среднее значение 19 нг / мл) по сравнению с таковыми в группе. 1087 человек без депрессии (среднее значение 22 нг / мл) ( P <.001). Уровни ПТГ были на 5% выше у лиц с незначительной депрессией (в среднем 3,72 пг / мл) и на 33% выше у лиц с БДР (в среднем 4,69 пг / мл) по сравнению с таковыми у лиц без депрессии (в среднем 3,53 пг / мл). ( P = 0,003).
Тяжесть депрессии (оценка по шкале CES-D) была связана со снижением уровней 25 (OH) D в сыворотке ( P <0,001) и повышением уровней ПТГ в сыворотке в соответствии с квартилями 25 (OH) D и уровней ПТГ ( P ). <0,001; Таблица 2). После корректировки на возраст, пол, ИМТ, статус курения и количество хронических состояний, баллы по шкале CES-D все еще были связаны со снижением 25 (OH) D ( P =.01) и повышенный уровень ПТГ ( P = 0,002). После дополнительной корректировки уровня урбанизации, которая была единственной объясняющей переменной, которая различалась в зависимости от депрессивного статуса, эти ассоциации оставались значимыми ( P = 0,03 и 0,008, соответственно). Что касается связи с депрессивными симптомами, не было обнаружено значимых условий взаимодействия для пола × 25 (OH) D ( P = 0,70) или пола × PTH ( P = 0,82), что указывает на то, что ассоциации были последовательными для разных полов. .Дополнительная корректировка уровней ПТГ не ослабляла связь между оценкой шкалы CES-D и уровнем 25 (OH) D, тогда как ПТГ оставался значимо связанным с оценкой шкалы CES-D после корректировки на уровень 25 (OH) D.
Это крупное популяционное исследование впервые показывает связь депрессивного статуса и тяжести депрессии со снижением уровня 25 (OH) D в сыворотке крови и повышенным уровнем ПТГ в сыворотке крови у пожилых людей.Эта связь была скорректирована с учетом потенциально смешивающих факторов, таких как возраст, пол, ИМТ, статус курения и количество хронических состояний, и не объясняется различиями в сезоне сбора данных, уровне физической активности или использовании антидепрессантов. Только уровень урбанизации имел небольшой объяснительный эффект, который подтверждает, что уменьшение воздействия света способствует снижению уровня 25 (OH) D в сыворотке крови и депрессии. То, что уровень ПТГ также был связан с депрессией после корректировки на уровень 25 (ОН) D, предполагает, что ПТГ также может играть роль в патогенезе депрессии.Это согласуется с высокой распространенностью депрессии при первичном гиперпаратиреозе.
Низкий уровень 25 (OH) D может быть вовлечен в патогенез депрессии несколькими путями. Авторадиография и иммуногистохимия на грызунах показали, что ткани-мишени 25 (OH) D связаны с экзокринными и эндокринными секреторными и соматотрофическими процессами, а не только с метаболизмом кальция. Более того, распределение нейронов-мишеней 25 (OH) D предполагает влияние уровней синтеза фактора роста нервов, 18 ацетилхолинацетилазы, серотонина, тестостерона, тироидного гормона и матричной РНК тирозингидроксилазы, 19 , 20 которые все были вовлечены в патогенез депрессии у людей.Недавно было обнаружено, что дефицит витамина D, ограниченный поздними сроками беременности, приводит к гиперлокомоции у взрослых крыс, что предполагает роль в патогенезе нервно-психических расстройств. 21 У людей распределение рецептора витамина D похоже на таковое у грызунов, с наиболее сильным окрашиванием в гипоталамусе, 22 , что указывает на его роль в нейроэндокринном функционировании. В более ранней работе мы обнаружили, что гипоталамус человека, вероятно, вовлечен в патогенез депрессии, потому что ряд типов нейронов показывает измененные уровни нейропептидов и соответствующую экспрессию генов в посмертной ткани мозга пациентов с депрессией по сравнению с контрольными субъектами. 11 23 26 Однако необходимы дополнительные патофизиологические исследования рецептора витамина D в гипоталамусе человека и возможных изменений при депрессии, чтобы подтвердить его значимость для патогенеза расстройства.
С точки зрения пациента, наши результаты могут иметь клиническое значение, поскольку распространенность малой депрессии у пожилых людей высока (13%) 1 , и как снижение уровня 25 (OH) D, так и повышение уровня ПТГ в сыворотке теоретически могут , лечиться более высоким потреблением с пищей витамина D 3 или кальция и повышенным воздействием дневного света. 27 Кроме того, было обнаружено, что статус витамина D предопределяет физическую работоспособность и ее снижение у пожилых людей. 28 Статус витамина D и ПТГ также может быть улучшен косвенно путем адекватного лечения антидепрессантами, что приводит к увеличению потребления пищи и повышению активности. 4 Более того, клиническая значимость настоящего исследования подчеркивается нашим выводом о том, что 38,8% мужчин и 56,9% женщин в нашей когорте на уровне общины имели недостаточный статус витамина D.
На сегодняшний день отсутствуют надежные данные о результатах исследований витамина D или легких интервенционных исследований у лиц с легкой депрессией или БДР, и результаты у субъектов с сезонным аффективным расстройством или здоровых субъектов с сезонными изменениями настроения противоречивы. 29 , 30 Мы не обнаружили влияния сезона на уровень 25 (OH) D. Только одно испытание 31 показало значительный положительный эффект фототерапии на 25 (OH) D. В этом исследовании препараты витамина D оказали положительное влияние как на уровень 25 (OH) D, так и на тяжесть депрессии, но количество субъектов, получавших витамин D, было небольшим (n = 8), и данные заслуживают последующего наблюдения. Поэтому мы предлагаем, чтобы в будущих крупномасштабных интервенционных исследованиях, например, по остеопорозу 32 у пожилых людей, использованию витамина D 3 и воздействию дневного света, также использовалось настроение в качестве критерия результата.
В заключение мы обнаружили, что тяжесть депрессии и депрессии, измеренная по шкале CES-D, тесно связана с более низкими уровнями 25 (OH) D в сыворотке крови и более высокими уровнями ПТГ, даже после корректировки на возраст, пол, ИМТ, статус курения. , состояние здоровья, уровень физической активности и уровень урбанизации. Следует провести долгосрочные продольные исследования с повторными оценками, чтобы выяснить, предшествуют ли снижение уровня 25 (OH) D и повышение уровня ПТГ депрессии или следуют за ней.Другими словами, являются ли эти биологические изменения причиной или следствием депрессии?
Для корреспонденции: Витте Дж. Г. Хоогендейк, доктор медицинских наук, Медицинский центр Университета VU, Амстердамский университет Vrije, A. J. Ernststraat 887, 1081 HL, Амстердам, Нидерланды ([email protected]).
Представлено для публикации: 24 июля 2007 г .; окончательная редакция получена 5 декабря 2007 г .; принята к печати 5 декабря 2007 г.
Раскрытие финансовой информации: Это исследование основано на данных, собранных в рамках Амстердамского лонгитюдного исследования старения, финансируемого Министерством здравоохранения Нидерландов, Гаага.
Финансирование / поддержка: Это исследование было поддержано грантом 907-00-012 (W.J.G.H.) ZonMw от Нидерландской организации научных исследований.
Роль спонсора: Нидерландская организация научных исследований не принимала участия в разработке и проведении исследования; в сборе, анализе и интерпретации данных; или при подготовке, рецензировании или утверждении рукописи.
1.Бикман ATCopeland Младший принц MJ Обзор распространенности депрессии в более позднем возрасте. Br J Psychiatry 1999; 174307-311PubMedGoogle ScholarCrossref 2. Penninx BWBeekman ATHonig ADeeg DJSchoevers Раван Эйк JTvan Тилбург W Депрессия и сердечная смертность: результаты длительного исследования на уровне сообщества. Arch Gen Psychiatry 2001; 58 (3) 221-227PubMedGoogle ScholarCrossref 3.Шнайдер BWeber Б.Френш AStein JFritz J Витамин D при шизофрении, большой депрессии и алкоголизме. J Neural Transm 2000; 107 (7) 839-842PubMedGoogle ScholarCrossref 4.Michelson DStratakis Холод LReynolds JGalliven EChrousos GGold P Минеральная плотность костей у женщин с депрессией. N Engl J Med 1996; 335 (16) 1176–1181PubMedGoogle ScholarCrossref 6.Watson LCMarx CE Новое начало психоневрологических симптомов у пожилых людей: возможный первичный гиперпаратиреоз. Психосоматика 2002; 43 (5) 413-417PubMedGoogle ScholarCrossref 7.Herrán AAmado Я.А.Гарсия-Унзуэта MTVázquez-Barquero JLPerera LGonzález-Macias J Усиление ремоделирования костей при первом эпизоде большого депрессивного расстройства. Psychosom Med 2000; 62 (6) 779-782PubMedGoogle ScholarCrossref 8.Уилкинс CHSheline YIRoe CMBirge SJMorris Дефицит витамина D JC связан с плохим настроением и ухудшением когнитивных функций у пожилых людей. Am J Geriatr Psychiatry 2006; 14 (12) 1032-1040PubMedGoogle ScholarCrossref 9.Deeg DJvan Тилбург TSmit JHde Leeuw ED Отсев в продольном исследовании старения в Амстердаме: влияние дифференциального включения в побочные исследования. J Clin Epidemiol 2002; 55 (4) 319-328PubMedGoogle ScholarCrossref 10.Снайдер Плотина МБван RMVisser MDeeg DJDekker JMBouter LMSeidell JCLips P Ожирение в зависимости от статуса витамина D и уровней паратиреоидного гормона: популяционное исследование у пожилых мужчин и женщин. J Clin Endocrinol Metab 2005; 90 (7) 4119-4123PubMedGoogle ScholarCrossref 11.Alkemade AUnmehopa UABrouwer JPHoogendijk WJWiersinga WMSwaab DFFliers E Снижение экспрессии гена тиреотропин-рилизинг-гормона в паравентрикулярном ядре гипоталамуса у пациентов с большой депрессией. Mol Psychiatry 2003; 8 (10) 838-839PubMedGoogle ScholarCrossref 12. Робинс LNHelzer Дж.Круган JRadcliff График диагностического интервью Национального института психического здоровья штата Канзас: его история, характеристики и достоверность. Arch Gen Psychiatry 1981; 38 (4) 381- 389PubMedGoogle ScholarCrossref 13.Pluijm SMFVisser MSmit Дж. Х. Попп-Снайдерс CRoos JCLips P Детерминанты минеральной плотности костей у пожилых мужчин и женщин: состав тела как посредник. J Bone Miner Res 2001; 16 (11) 2142 — 2151PubMedGoogle ScholarCrossref 14.Wortsman JMatsuoka LYChen TCLu Жолик MF Снижение биодоступности витамина D при ожирении [опубликованная поправка появилась в Am J Clin Nutr . 2003; 77 (5): 1342]. Am J Clin Nutr 2000; 72 (3) 690-693PubMedGoogle Scholar15.Kriegsman DMDeeg DJStalman WA Коморбидность соматических хронических заболеваний и ухудшение физического функционирования: Долгосрочное исследование старения, Амстердам. J Clin Epidemiol 2004; 57 (1) 55-65PubMedGoogle ScholarCrossref 16. Чжоу CAssem MTay Дж. К. Уоткинс PBBlumberg BSchuetz EGThummel KE Перекрестные помехи между рецепторами стероидов и ксенобиотиков и рецепторами витамина D опосредуют экспрессию CYP24 и лекарственную остеомаляцию. J Clin Invest 2006; 116 (6) 1703-1712PubMedGoogle ScholarCrossref 17. Губы P Дефицит витамина D и вторичный гиперпаратиреоз у пожилых людей: последствия потери костной массы и переломов и терапевтические последствия. Endocr Ред. 2001; 22 (4) 477-501PubMedGoogle ScholarCrossref 18.Wion D MacGrogan DNeveu IJehan FHoulgatte RBrachet P 1,25-Дигидроксивитамин D 3 является мощным индуктором синтеза фактора роста нервов. J Neurosci Res 1991; 28 (1) 110–114PubMedGoogle ScholarCrossref 19.Stumpf WE Сайты и механизмы действия витамина D: гистохимическая перспектива. Размышления о полезности авторадиографии и цитофармакологии для нацеливания лекарств. Histochem Cell Biol 1995; 104 (6) 417- 427PubMedGoogle ScholarCrossref 20.Stumpf WEO’Brien LP 1,25 (OH) 2 витамин D 3 участков действия в головном мозге: авторадиографическое исследование. Гистохимия 1987; 87 (5) 393-406PubMedGoogle ScholarCrossref 21.O’Loan Джейлс DWKesby JKo PMcGrath JJBurne TH Дефицит витамина D на разных стадиях беременности у крыс: его влияние на развитие и поведение взрослого потомства. Психонейроэндокринология 2007; 32 (3) 227-234PubMedGoogle ScholarCrossref 22.Eyles DWSmith Скинобе RHewison MMcGrath JJ Распределение рецептора витамина D и 1-альфа-гидроксилазы в мозге человека. J Chem Neuroanat 2005; 29 (1) 21-30PubMedGoogle ScholarCrossref 23.Meynen Гунмехопа Грнофман MASwaab DFHoogendijk WJ Гипоталамическая экспрессия мРНК окситоцина и меланхолическая депрессия. Mol Psychiatry 2007; 12 (2) 118-119PubMedGoogle ScholarCrossref 24.Purba JSHoogendijk WJHofman MASwaab DF Повышенное количество нейронов, экспрессирующих вазопрессин и окситоцин, в паравентрикулярном ядре гипоталамуса при депрессии. Arch Gen Psychiatry 1996; 53 (2) 137-143PubMedGoogle ScholarCrossref 25. Раадшир FCHoogendijk WJStam FCTilders FJSwaab DF Увеличение количества нейронов, экспрессирующих кортикотропин-рилизинг-гормон, в паравентрикулярном ядре гипоталамуса у пациентов с депрессией. Нейроэндокринология 1994; 60 (4) 436-444PubMedGoogle ScholarCrossref 26.Zhou JNRiemersma РФ UAHoogendijk WJvan Heerikhuize JJHofman MASwaab DF Изменения в нейронах аргинина вазопрессина в супрахиазматическом ядре при депрессии. Arch Gen Psychiatry 2001; 58 (7) 655-662PubMedGoogle ScholarCrossref 27.Stumpf WEPrivette TH Light, витамин D и психиатрия: роль 1,25-дигидроксивитамина D 3 (солтриол) в этиологии и терапии сезонного аффективного расстройства и других психических процессов. Психофармакология (Берл) 1989; 97 (3) 285-294PubMedGoogle ScholarCrossref 28.Wicherts И.С.ван Шур NMBoeke AJVisser MDeeg DJSmit JKnol DLLips Статус витамина P P предсказывает физическую работоспособность и ее снижение у пожилых людей. J Clin Endocrinol Metab 2007; 92 (6) 2058-2065PubMedGoogle ScholarCrossref 30.Oren Д.А.Щулкин JRosenthal NE 1,25 (OH) 2 витамин D 3 уровней при сезонном аффективном расстройстве: эффекты света. Психофармакология (Берл) 1994; 116 (4) 515-516PubMedGoogle ScholarCrossref 31.Gloth FM IIIAlam Уоллис Витамин D против фототерапии широкого спектра в лечении сезонного аффективного расстройства. J Nutr Health Aging 1999; 3 (1) 5-7PubMedGoogle Scholar32.Jackson RD LaCroix AZGass MWallace RBRobbins Дж. Льюис CEBassford TBeresford SABlack HRBlanchette PBonds DEBrunner Р.Л.Бжиски RGCaan BCauley Я.Хлебовски RTCummings С.Р.Гранек IHays JHeiss GHendrix SLHoward BVHsia Дж. Хаббелл Ф.А.Джонсон KCJudd HKotchen Дж. М. Куллер LHLanger RDLasser Н.Л. Лимахер MCLudlam SManson Дж. Э. Марголис KLMcGowan JOckene JKO’Sullivan MJPhillips LPrentice Р.Л.Сарто GEStefanick MLVan Horn LWactawski-Wende JWhitlock Эандерсон GLAssaf АРБарад Инициатива DWomen’s Health, добавление кальция и витамина D и риск переломов [опубликованное исправление опубликовано в N Engl J Med .2006; 354 (10): 1102]. N Engl J Med 2006; 354 (7) 669-683PubMedGoogle ScholarCrossrefКреатин — питание и травматическое повреждение головного мозга
Креатин (N- [аминоиминометил] -N-метилглицин) представляет собой аминокислотное соединение, которое эндогенно вырабатывается в печени, почках, поджелудочной железе и, возможно, в мозге в результате биосинтеза незаменимые аминокислоты метионин, глицин и аргинин или полученные из пищевых источников. Основными источниками питания являются продукты с высоким содержанием белка, включая мясо, рыбу и птицу.После синтеза или проглатывания креатин переносится из плазмы через стенку кишечника в другие ткани с помощью определенных переносчиков креатина, расположенных в скелетных мышцах, почках, сердце, печени и головном мозге.
Креатин и система креатинкиназа / фосфокреатин играют важную роль в качестве резервных источников энергии в тканях с высокими и непостоянными потребностями в энергии (например, в мышцах и головном мозге). Роль креатина в энергетическом обмене включает перенос N-фосфорильных групп от фосфорилкреатина к аденозиндифосфату (АДФ) для регенерации аденозинтрифосфата (АТФ) посредством обратимой реакции, катализируемой фосфорилкреатинкиназой (Andres et al., 2008; Броснан и Броснан, 2007).
И креатин, и фосфокреатин спонтанно расщепляются до креатинина, который выводится из организма с мочой. Скорость потери креатина составляет примерно 1,7 процента от общего запаса креатина в организме в день. Поскольку более 90 процентов креатина и фосфокреатина находится в скелетных мышцах, потери креатина и экскреция креатина варьируются в зависимости от различий в мышечной массе в зависимости от возраста, пола и уровня повседневной активности. Экскреция креатинина наиболее высока у молодых мужчин в возрасте от 18 до 29 лет — типичный возраст большинства военнослужащих в бою (Brosnan and Brosnan, 2007).
КРЕАТИН И МОЗГ
Хотя мозг составляет всего 2 процента от общей массы тела, он использует примерно 20 процентов энергии тела. Поскольку мозгу требуется значительный оборот АТФ для поддержания мембранного потенциала и сигнальной способности, метаболизм креатина и система креатинкиназа / фосфокреатин важны для нормальной функции мозга и могут быть нарушены при заболеваниях центральной нервной системы (Andres et al., 2008) (см. также статьи Салливана и Холла в Приложении C).
Доказательства важности креатиновой системы для функции мозга получены в исследованиях на животных с использованием мышей с нокаутом креатинкиназы, которые демонстрируют значительное снижение функционирования гиппокампа и нарушения как обучения, так и памяти по сравнению с генетически модифицированными мышами. У мышей, которым вводят препарат, который конкурентно ингибирует переносчик креатина и приводит к уменьшению запасов креатина в мышцах и головном мозге, дополнительно ухудшается обучение (Andres et al., 2008). Напротив, мыши, получавшие диету с добавлением 1% креатина, живут дольше, имеют меньше активных форм кислорода в мозгу и демонстрируют лучшую функцию памяти, чем однопометники без добавок (Bender et al., 2008).
У людей люди, рожденные с ошибками в синтезе креатина или дефектами х-связанного транспортера креатина, страдают умственной отсталостью, аутичным поведением, серьезными нарушениями речи и речи, эпилепсией и атрофией мозга. У некоторых людей когнитивные нарушения, возникающие в результате этих врожденных ошибок, могут быть улучшены, хотя и не полностью устранены, с помощью хронического приема больших доз креатина (Andres et al., 2008; Gualano et al., 2010).
Исследования с использованием спектроскопии ядерного магнитного резонанса показали, что добавка креатина увеличивает уровень креатина и фосфокреатина в мозге (Dechent et al., 1999; Пан и Такахаши, 2007). Такое повышение уровня креатина в мозге может привести к улучшению когнитивного поведения, особенно в стрессовых или неблагоприятных условиях. Например, молодые люди, получавшие креатиновые добавки до 36-часового недосыпания, лучше справились с тестом на управляющее функционирование, чем люди, не получавшие добавок (McMorris et al., 2006). Кроме того, было показано, что креатиновые добавки улучшают рабочую память у вегетарианцев, у которых уровень креатина обычно ниже, чем у всеядных (Rae et al., 2003), а также для улучшения результатов тестов вербальной и пространственной памяти у пожилых людей (McMorris et al., 2007).
Появляется все больше доказательств того, что креатин может иметь ценность при лечении ряда неврологических состояний, включая синдромы врожденного дефицита креатина, возрастное снижение когнитивных функций (например, болезнь Альцгеймера) и нейродегенеративные заболевания (например, болезнь Паркинсона, болезнь Хантингтона). болезнь, боковой амиотрофический склероз), все из которых связаны с дисфункциональным энергетическим обменом (Andres et al., 2008; Gualano et al., 2010). Добавки креатина также начинают привлекать внимание в качестве дополнительной стратегии лечения психических расстройств (например, депрессии, посттравматического стрессового расстройства и шизофрении) (Allen et al., 2010; Amital et al., 2006b; Roitman et al., 2007).
Как описано в главе 3, повреждение после черепно-мозговой травмы (ЧМТ) можно разделить на первичные, вторичные и долгосрочные. Хотя понимание механизмов, лежащих в основе вторичного повреждения, все еще находится на ранней стадии, данные показывают, что нарушения функции митохондрий играют роль в опосредовании отсроченных последствий ЧМТ (Andres et al., 2008; Шефф и Диллон, 2004; Салливан и др., 2000; Zhu et al., 2004). Была выдвинута гипотеза, что нейропротективные эффекты креатина после ЧМТ могут включать вызванное креатином поддержание биоэнергетики митохондрий (Sullivan et al., 2000). Подтверждением этой гипотезы является открытие Салливана и его коллег (2000) о том, что крысы, получавшие диету, содержащую один процент креатина в течение четырех недель, демонстрировали значительно более высокие потенциалы митохондриальной мембраны и поддерживали уровни АТФ лучше, чем крысы без добавок после контролируемого ушиба коры.Другие признаки улучшения функции митохондрий у крыс, получавших креатин, включали снижение уровней реактивных окислительных посредников и внутримитохондриального кальция.
Уровни лактата и свободных жирных кислот в мозге повышаются после очаговой травмы головного мозга. Считается, что это увеличение является следствием вторичного повреждения мозга в результате накопления эксайтотоксических уровней нейромедиатора глутамата. Scheff и Dhillon (2004) продемонстрировали, что уровни лактата и свободных жирных кислот в кортикальном слое и гиппокампе ниже у крыс, получавших креатин, чем у крыс без добавок (Scheff and Dhillon, 2004).Эти результаты подтверждают гипотезу о том, что нейропротекторное действие креатина, по крайней мере, частично связано с уменьшением процессов, связанных с вторичным повреждением головного мозга.
Также было высказано предположение, что нейропротекторные эффекты креатина могут отражать его способность улучшать цереброваскулярную функцию (Prass et al., 2007). Это предложение подтверждается недавним исследованием, показывающим, что кормление креатином мышей, подвергшихся окклюзии средней артерии, привело к уменьшению объемов инфаркта.Хотя не было изменений в креатине, фосфокреатине, АТФ, АДФ или аденозинмонофосфате (АМФ) в головном мозге, такие добавки действительно улучшили церебральный кровоток у этих животных, что позволяет предположить, что креатин может оказывать благотворное влияние на функционирование сосудов головного мозга.
Список исследований на людях (1990 г. и позже), оценивающих эффективность креатина в обеспечении устойчивости или лечении ЧМТ или связанных заболеваний или состояний (например, субарахноидального кровоизлияния, внутричерепной аневризмы, инсульта, аноксической или гипоксической ишемии, эпилепсии) у людей: представлен в.Это также включает подтверждающие данные, полученные на животных моделях ЧМТ. Хотя в этот отчет обычно не включаются исследования эффективности диетических вмешательств в отношении долгосрочных последствий последствий ЧМТ, в эту главу включена депрессия как эффект ЧМТ. Наличие или отсутствие побочных эффектов в исследованиях на людях включается, если сообщается авторами.
ИСПОЛЬЗОВАНИЕ И БЕЗОПАСНОСТЬ
Креатин — одна из наиболее широко используемых пищевых добавок. Атлеты, культуристы и военнослужащие используют креатин для увеличения мышечной массы и силы.Креатин также используется в качестве эргогенного средства для повышения эффективности краткосрочных высокоинтенсивных упражнений (Bemben and Lamont, 2005; Branch, 2003; IOM, 2008). Популярность креатина в качестве пищевой добавки еще более возросла в результате исследования 2006 года, продемонстрировавшего его положительное влияние на когнитивные и психомоторные функции (McMorris et al., 2006).
Поскольку он может синтезироваться в организме, Рекомендуемая диета для креатина отсутствует; однако в результате ежедневных потерь запасы креатина должны поддерживаться либо диетой, либо синтезом.Исследования показывают, что добавление креатина увеличивает запасы креатина и фосфокреатина в мышцах, особенно у молодых людей, которые занимаются высокой физической активностью, и у вегетарианцев, у которых может быть меньше, чем оптимальный запас фосфокреатина (Brosnan and Brosnan, 2007; Burke et al. ., 2003; Rawson et al., 2002).
Эксперименты среди спортсменов и военнослужащих показывают, что креатин, принимаемый в количествах, обычно доступных в добавках, вызывает минимальные побочные эффекты, если таковые имеются (IOM, 2008; Shao and Hathcock, 2006).Используя данные хорошо спланированных рандомизированных контролируемых клинических испытаний креатина на людях, Шао и Хэткок (2006) пришли к выводу, что постоянное потребление 5 г креатина в день безопасно и не представляет значительного риска для здоровья.
ДОКАЗАТЕЛЬСТВА, ПОКАЗЫВАЮЩИЕ ВЛИЯНИЕ НА УСТОЙЧИВОСТЬ
Исследования на людях
Как и в случае с другими питательными веществами или компонентами пищевых продуктов, комитет не обнаружил исследований на людях, в которых проверялось бы потенциальное преимущество креатина при ЧМТ или других связанных заболеваниях или состояниях, включенных в обзор литературы (субарахноидальный кровоизлияние, внутричерепная аневризма, инсульт, аноксическая или гипоксическая ишемия, эпилепсия).
Исследования на животных
Исследования на животных позволяют определить влияние добавок креатина на биохимические и физиологические, а также на поведенческие последствия ЧМТ. В большинстве исследований, оценивающих нейропротекторные эффекты креатина, в качестве модели ЧМТ использовались легкие ушибы коры головного мозга. Эти ушибы приводят к значительному сокращению корковой ткани, нарушению гематоэнцефалического барьера, потере нейронов гиппокампа и серьезным нарушениям поведения.Используя эту модель, Салливан и его коллеги (2000) продемонстрировали дозозависимое снижение коркового повреждения после ушибов у мышей, которым внутрибрюшинно вводили креатин за 1, 3 или 5 дней до индукции повреждения мозга. Точно так же крысы, получавшие стандартную диету для грызунов с добавлением 1% креатина в течение четырех недель до индукции ЧМТ, демонстрировали на 50% меньшее повреждение коры, чем крысы без добавок. Дальнейшие доказательства потенциально нейропротекторного действия креатина получены из исследования Scheff и Dhillon (2004), которые обнаружили, что крысы, получавшие диету, содержащую 1% креатина, до того, как испытали контролируемые корковые ушибы, имели значительно большую щадь кортикальной ткани, чем крысы, не получавшие креатин.
Эти положительные результаты подтверждаются исследованиями, проведенными на других моделях черепно-мозговой травмы. Например, Прасс и его коллеги (2007) сообщили, что у мышей, получавших диету, содержащую 1 или 2 процента креатина в течение трех недель, предшествовавших окклюзии средней мозговой артерии, модели ишемии, было значительно меньше повреждений мозга, чем у мышей, не получавших креатин. В этом исследовании, однако, нейропротекторные эффекты креатина не наблюдались, когда большая доза креатина давалась сразу после начала ишемии или когда давалась меньшая доза в течение 12 месяцев.
ДОКАЗАТЕЛЬСТВА, ПОКАЗЫВАЮЩИЕ ВЛИЯНИЕ НА ЛЕЧЕНИЕ
Исследования на людях
Не проводилось исследований по изучению воздействия добавок креатина на ЧМТ у взрослых. Результаты предварительных исследований Сакелларис и его коллег (2006; 2008) показывают, что добавление креатина может быть полезным при лечении вторичных и долгосрочных симптомов ЧМТ, которые не являются непосредственными последствиями травмы, но развиваются в течение минут, часов, или через несколько дней после травмы. Постановка задачи для этого исследования не включала систематический обзор эффективности вмешательств по питанию в отношении долгосрочных последствий ЧМТ.Исследования креатина и долгосрочных эффектов, таких как депрессия, включены сюда как прелюдия к обзору, который, по мнению комитета, должен быть проведен в будущем. В первом исследовании нейрозащитные эффекты пероральной суспензии 0,4 г / кг креатина, вводимой сначала в течение 4 часов с момента травмы, а затем один раз в день в течение 6 месяцев, изучались у пациентов с ЧМТ в возрасте от 1 до 18 лет. Дети и подростки, получавшие креатин, проводили меньше времени в отделении интенсивной терапии и им требовалось кормление через зонд в течение более короткого периода времени, чем в контрольной группе, не получавшей креатин.При обследовании через три и шесть месяцев после травмы люди, получавшие креатин, показали большее улучшение когнитивных функций, самообслуживания, коммуникабельности и коммуникативных навыков, чем в контрольной группе (Sakellaris et al., 2006). Во второй части исследования с той же группой пациентов и с использованием того же режима дозирования креатина доля детей с головными болями, головокружением и утомляемостью в течение шестимесячного периода наблюдения была значительно ниже в группе, получавшей креатин, чем в группе, получавшей креатин. контрольная группа (Sakellaris et al., 2008). О побочных эффектах приема креатина не сообщалось.
Депрессия — заметное долгосрочное последствие ЧМТ. Появляется все больше свидетельств того, что нарушения клеточной устойчивости, нейрональной пластичности и биоэнергетической функции мозга связаны с патогенезом депрессии. Была высказана гипотеза, что, обращая вспять эти нарушения, креатин может быть полезен для предотвращения или лечения депрессии. Результаты ряда исследований указывают на связь между депрессией, частым сопутствующим ЧМТ (Bombardier et al., 2010; Хорхе и Старкштейн, 2005; Masel and DeWitt, 2010) и креатин, и предполагают, что креатин может иметь ценность при лечении депрессивных симптомов, наблюдаемых у пациентов с ЧМТ. Первые доказательства роли креатина в депрессии были получены в исследованиях, демонстрирующих значительную отрицательную корреляцию между метаболитами креатина и самооценкой суицидальных мыслей у пациентов, страдающих серьезными депрессивными расстройствами (Agren and Niklasson, 1988). Исследования, проводимые с 2000 года, также показали, что уровни креатина в мозге обратно пропорциональны тяжести депрессивного эпизода (Dager et al., 2004; Segal et al., 2007). Подтверждение потенциальной полезности креатина в лечении депрессии основано на открытых исследованиях, демонстрирующих, что ежедневный пероральный прием от трех до пяти граммов креатина улучшает настроение у пациентов с депрессией, устойчивых к антидепрессантам, и у пациентов с коморбидным посттравматическим стрессовым расстройством (Amital et al. др., 2006б; Ройтман и др., 2007).
Пациенты с фибромиалгией часто страдают от хронической боли, усталости, проблем со сном и депрессии, симптомы также часто возникают после травм головного мозга; Сообщалось о сильном совпадении между фибромиалгией и посттравматическим стрессовым расстройством (Amital et al., 2006а). В ходе восьминедельного открытого исследования креатина было отмечено значительное улучшение качества жизни, режима сна и боли у пациентов с фибромиалгией. Эти улучшения ухудшились через четыре недели после прекращения терапии креатином. Во всех предыдущих исследованиях побочные реакции на добавление креатина, если таковые имели место, были умеренными (например, тошнота) и со временем уменьшались.
Хотя результаты предыдущих исследований предполагают многообещающее использование креатина при лечении ЧМТ, они ограничены небольшим количеством участников и отсутствием двойных слепых процедур.
Исследования на животных
Добавки креатина также могут уменьшить ряд пагубных последствий церебральной ишемии. После окклюзии средней мозговой артерии мыши, получавшие диету с добавлением двух процентов креатина в течение одного месяца, показали значительно лучшую неврологическую функцию и значительно меньшее повреждение мозга, чем контрольная группа, не получавшая креатин. Креатин также значительно снижает вызванное ишемией истощение АТФ, а также активацию каспазы-3 и высвобождение цитохрома c , индикаторов повреждения клеток (Zhu et al., 2004).
Как отмечалось выше, депрессия — один из наиболее распространенных симптомов ЧМТ. В поддержку исследований креатина и депрессии на людях, хроническое потребление диет с добавлением одного или двух процентов креатина уменьшало депрессивно-подобное поведение дозозависимым образом у самок крыс в тесте принудительного плавания, модели депрессии на животных. Однако у самцов крыс потребление креатиновой диеты не уменьшало депрессивное поведение (Allen et al., 2010). Наблюдение за поло-зависимыми эффектами креатина интересно, учитывая тот факт, что депрессия чаще диагностируется у женщин, чем у мужчин.
ВЫВОДЫ И РЕКОМЕНДАЦИИ
Основываясь на результатах исследований на животных и людях, креатин представляет собой многообещающую пищевую добавку для повышения устойчивости к ЧМТ и ее лечения. Взятые вместе, результаты представленных исследований показывают, что нейрозащита, возникающая в результате приема диет с добавлением креатина, обусловлена, по крайней мере частично, подавлением вторичного повреждения мозга. Хотя необходимы дополнительные исследования как на животных, так и на людях, эти результаты показывают, что профилактическое лечение креатином может быть полезным для людей с повышенным потенциалом развития ЧМТ; однако существует ряд вопросов, которые необходимо решить до начала использования креатина в вооруженных силах.
Вопрос о том, как экспериментально оценить профилактическое действие креатина на устойчивость, остается без ответа. Исследователи могут давать креатин группе лиц с высоким риском (например, людям с высоким риском инсульта или повышенным, чем обычно, риском травмы головы) и сравнивать функционирование и поведение мозга у тех, кто в конечном итоге получает травму мозга, с таковым у людей. в аналогичных группах, которым не давали креатин. Однако есть проблемы с проведением такого исследования.Во-первых, получение статистически адекватного числа участников для долгосрочного проспективного исследования может оказаться трудным. Дополнительной проблемой является отсутствие в настоящее время окончательных рекомендаций о том, сколько креатина следует давать и как долго он должен подаваться. Действительно, исследования на животных показывают, что длительное употребление креатина может снизить его способность защищать от последствий травмы головного мозга (Prass et al., 2007). Эти проблемы с осуществимостью проспективных исследований будут общими для любого интересующего питательного или пищевого компонента.Чтобы преодолеть эти проблемы, комитет рекомендует в Главе 5 провести исследование статуса приема пищи до и после травмы (например, использование пищевых добавок) у лиц с ЧМТ, чтобы определить любую связь с исходом ЧМТ, включая анализ возможных синергетический эффект между питательными веществами, компонентами пищи и биологически активными добавками. Креатин должен быть включен в это исследование.
Что касается лечения, то предварительные исследования Сакеллариса и его коллег (2006, 2008) предоставляют доказательства потенциально положительного терапевтического воздействия креатина на функцию и поведение мозга после травмы головного мозга.Эти положительные эффекты должны быть подтверждены у взрослого населения.
РЕКОМЕНДАЦИЯ 10-1. Основываясь на доказательствах, подтверждающих влияние креатина на функцию и поведение мозга у детей и подростков после травм головного мозга, Министерство обороны должно начать исследования на взрослых, чтобы оценить ценность креатина для лечения пациентов с ЧМТ.
СПИСОК ЛИТЕРАТУРЫ
- Agren, H., and F. Niklasson. 1988. Креатинин и креатин в спинномозговой жидкости — показатели энергообмена мозга при депрессии. Журнал нейронной передачи 74 (1): 55–59. [PubMed: 3171573]
- Аллен П. Дж., К. Э. Д’Анси, Р. Б. Канарек и П. Ф. Реншоу. 2010. Хронический прием креатина изменяет депрессивное поведение у грызунов в зависимости от пола. Нейропсихофармакология 35 (2): 534–546. [Бесплатная статья PMC: PMC2794979] [PubMed: 19829292]
- Амиталь, Д., Л. Фостик, М. Л. Поллиак, С. Сегев, Дж. Зохар, А. Рубинов и Х. Амитал. 2006. а. Посттравматическое стрессовое расстройство, болезненность и синдром фибромиалгии: это разные сущности? Психосоматический журнал Re поиск 61 (5): 663–669.[PubMed: 17084145]
- Амиталь Д., Т. Вишне, С. Ройтман, М. Котлер и Дж. Левин. 2006 г. б. Открытое исследование моногидрата креатина при резистентном к лечению посттравматическом стрессовом расстройстве. Журнал клинической психиатрии 67 (5): 836–837. [PubMed: 16841637]
- Андрес, Р. Х., А. Д. Дюкрей, У. Шлаттнер, Т. Валлиман и Х. Р. Видмер. 2008. Функции и влияние креатина на центральную нервную систему. Бюллетень исследований мозга 76 (4): 329–343. [PubMed: 18502307]
- Бембен, М.Г. и Х. С. Ламонт. 2005. Креатиновые добавки и упражнения — недавние открытия. Спортивная медицина 35 (2): 107–125. [PubMed: 15707376]
- Бендер, А., Дж. Бекерс, И. Шнайдер, С. М. Холтер, Т. Хаак, Т. Рутзац, Д. М. Фогт-Вайзенхорн, Л. Беккер, Дж. Гениус, Д. Руджеску, М. Ирмлер, Т. Мияльски, М. Мадер, Л. Кинтанилья-Мартинес, Х. Фукс, В. Гайлус-Думер, М. Х. де Ангелис, В. Вурст, Дж. Шмид и Т. Клопсток. 2008. Креатин улучшает здоровье и выживаемость мышей. Нейробиология старения 29 (9): 1404–1411. [PubMed: 17416441]
- Бомбардье, К. Х., Дж. Р. Фанн, Н. Р. Темкин, П. К. Эссельман, Дж. Барбер и С. С. Дикмен. 2010. Показатели большого депрессивного расстройства и клинические исходы после черепно-мозговой травмы. Журнал Американской медицинской ассоциации Ассоциация 303 (19): 1938–1945. [Бесплатная статья PMC: PMC30
] [PubMed: 20483970]
- Branch, J. D. 2003. Влияние креатина на состав тела и работоспособность: метаанализ. Международный журнал спортивного питания и метаболизма физических упражнений 13 (2): 198–226. [PubMed: 12945830]
- Броснан, Дж. Т. и М. Э. Броснан. 2007. Креатин: эндогенный метаболит, пищевая и терапевтическая добавка. Ежегодный обзор питания 27: 241–261. [PubMed: 17430086]
- Берк, Д. Г., П. Д. Чилибек, Г. Париз, Д. Г. Кандоу, Д. Махони и М. Тарнопольски. 2003. Влияние креатина и силовых тренировок на креатин в мышцах и производительность у вегетарианцев. Медицина и наука в спорте и Упражнение 35 (11): 1946–1955. [PubMed: 14600563]
- Дагер, С. Р., С. Д. Фридман, А. Паров, К. Демопулос, А. Л. Столл, К. Лью, Д. Л. Даннер и П. Ф. Реншоу. 2004. Изменения метаболизма мозга у пациентов с биполярным расстройством, не принимающих лекарства. Архив общей психиатрии 61 (5): 450–458. [PubMed: 15123489]
- Dechent, P., P. J. Pouwels, B. Wilken, F. Hanefeld и J. Frahm. 1999. Увеличение общего креатина в мозге человека после перорального приема моногидрата креатина. Американский журнал физиологии 277 (3, часть 2): R698–704. [PubMed: 10484486]
- Гуалано Б., Г. Г. Артиоли, Дж. Р. Портманс и А. Х. Ланча. 2010. Изучение терапевтической роли добавок креатина. Аминокислоты 38 (1): 31–44. [PubMed: 19253023]
- МОМ (Институт медицины). 2008. Употребление БАД военнослужащими . Вашингтон: Издательство национальных академий. [PubMed: 20669410]
- Хорхе, Р.Э., Старкштейн С. 2005. Патофизиологические аспекты большой депрессии после черепно-мозговой травмы. Журнал реабилитации после травм головы 20 (6): 475–487. [PubMed: 16304485]
- Masel, B.E., and D. S. DeWitt. 2010. Черепно-мозговая травма: заболевание, а не событие. Журнал Neu ротравма 27 (8): 1529–1540. [PubMed: 20504161]
- МакМоррис, Т., Р. К. Харрис, Дж. Суэйн, Дж. Корбетт, К. Коллард, Р. Дж. Дайсон, Л. Дай, К. Ходжсон и Н.Дрейпер. 2006. Влияние добавок креатина и лишения сна при легких физических упражнениях на когнитивные и психомоторные функции, настроение и концентрацию катехоламинов и кортизола в плазме. Psychopharma cology 185 (1): 93–103. [PubMed: 16416332]
- МакМоррис, Т., Р. К. Харрис, А. Н. Ховард, Г. Лэнгридж, Б. Холл, Дж. Корбетт, М. Дикс и К. Ходжсон. 2007. Добавки креатина, лишение сна, кортизол, мелатонин и поведение. Физиология и поведение 90 (1): 21–28.[PubMed: 17046034]
- Пан, Дж. У. и К. Такахаши. 2007. Церебральные энергетические эффекты добавок креатина у людей. Американский Журнал физиологии-регулятивной интегративной и сравнительной физиологии 292 (4): R1745 – R1750. [Бесплатная статья PMC: PMC3570026] [PubMed: 17185404]
- Prass, K., G. Royl, U. Lindauer, D. Freyer, D. Megow, U. Dirnagl, G. Stockler-Ipsiroglu, T. Wallimann, and Дж. Приллер. 2007. Улучшенная реперфузия и нейрозащита креатином на мышиной модели инсульта. Церебральный журнал Кровоток и метаболизм 27 (3): 452–459. [PubMed: 16773141]
- Рэй, К., А. Л. Дигни, С. Р. Макьюэн и Т. К. Бейтс. 2003. Пероральный прием моногидрата креатина улучшает работу мозга: двойное слепое плацебо-контролируемое перекрестное исследование. Труды Королевского общества Лондон Серия B-Биологические науки 270 (1529): 2147–2150. [Бесплатная статья PMC: PMC1691485] [PubMed: 14561278]
- Rawson, E.С., П. М. Кларксон, Т. Б. Прайс и М. П. Майлз. 2002. Дифференциальный ответ мышечного фосфокреатина на добавку креатина у молодых и старых субъектов. Acta Physiologica Scandinavica 174 (1): 57–65. [PubMed: 11851597]
- Ройтман, С., Т. Грин, Ю. Ошер, Н. Карни и Дж. Левин. 2007. Моногидрат креатина при устойчивой депрессии: предварительное исследование. Биполярное расстройство 9: 754–758. [PubMed: 17988366]
- Сакелларис, Г., М. Котсиу, М. Тамиолаки, Г.Калостос, Э. Цапаки, М. Спанаки, М. Спилиоти, Г. Чариссис и А. Евангелиу. 2006. Профилактика осложнений, связанных с черепно-мозговой травмой у детей и подростков с применением креатина: открытое рандомизированное пилотное исследование. Журнал травм-травм и инфекций Критическая помощь 61 (2): 322–329. [PubMed: 16917445]
- Сакелларис, Г., Г. Насис, М. Котсиу, М. Тамиолаки, Г. Чариссис и А. Евангелиу. 2008. Профилактика травматической головной боли, головокружения и утомляемости с помощью креатина.Пилотное исследование. Acta Paediatrica 97 (1): 31–34. [Бесплатная статья PMC: PMC2583396] [PubMed: 18053002]
- Scheff, S. W. и H. S. Dhillon. 2004. Креатиновая диета изменяет уровни лактата и свободных жирных кислот после экспериментальной травмы головного мозга. Нейрохимические исследования 29 (2): 469–479. [PubMed: 15002746]
- Сегал, М., А. Авиталь, М. Дробот, А. Луканин, А. Деревенский, С. Сандбэнк и А. Вейцман. 2007. Уровень креатинкиназы в сыворотке крови у пациентов с непсихотической, психотической, биполярной и шизоаффективной депрессией без лечения. Европейский Нейропсихофармакология 17 (3): 194–198. [PubMed: 17049435]
- Шао А. и Дж. Н. Хэткок. 2006. Оценка риска моногидрата креатина. Нормативная токсикология и Phar макология 45 (3): 242–251.