Проинсулин
Общая информация об исследовании
Проинсулин является предшественником инсулина и С-пептида. Образованный островками В-клеток поджелудочной железы, проинсулин заключен в секреторные гранулы, в которых он преобразуется прогормонами конвертазами (РС1/3 и РС2) и карбоксипептидазой Е в С-пептид либо в инсулин. Лишь от 1 % до 3 % проинсулина в организме остается в несвязанном состоянии. Однако, поскольку период полураспада проинсулина дольше, чем у инсулина, концентрация проинсулина в крови составляет от 10 % до 30 % от концентрации циркулирующего в крови инсулина. Это количество увеличивается после принятия пищи, а также у пациентов с резистентностью к инсулину на начальной стадии развития диабета второго типа. Проинсулин способен связываться с инсулиновыми рецепторами и проявлять 5-10 % метаболической активности инсулина.
Уровень проинсулина может повышаться при опухолях островковых клеток, вырабатывающих инсулин. Пациенты при этом страдают от гипогликемических приступов из-за нарушения секреции инсулина.
Для уверенной постановки диагноза часто требуется продолжительное голодание пациента (до 72 часов перед сдачей анализа), а также дополнительные тесты (на глюкозу, сульфонилмочевину, С-пептид, проинсулин и бета-гидроксибутират).
Избыточная секреция инсулина при инсулиномах способствует высвобождению большого количества незрелых гранул, содержащих не до конца сформированный проинсулин, что приводит к повышенному уровню проинсулина в крови. Так происходит, например, при голодании, когда уровень проинсулина в норме не поднимается выше 5 % от концентрации инсулина.
Количество проинсулина резко увеличивается при дефиците конвертазы РС1/3.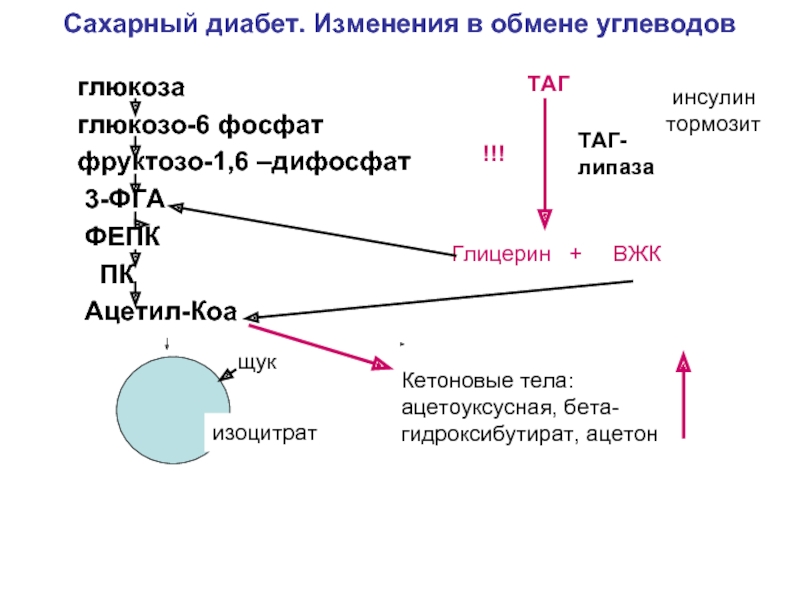 Это ведет к отклонениям в переработке многочисленных пептидных гормонов и, соответственно, к диабету, избыточной массе тела, почечной недостаточности, бесплодию. Такие пациенты, как правило, имеют рыжие волосы независимо от расовой принадлежности. Известно, что мутации в молекуле проинсулина влияют на способность РС расщеплять субстраты и на дальнейший метаболизм проинсулина. Кроме того, они иногда приводят к заметному повышению уровня проинсулина, но обычно не сопровождаются диабетом либо другими гормональными отклонениями.
Это ведет к отклонениям в переработке многочисленных пептидных гормонов и, соответственно, к диабету, избыточной массе тела, почечной недостаточности, бесплодию. Такие пациенты, как правило, имеют рыжие волосы независимо от расовой принадлежности. Известно, что мутации в молекуле проинсулина влияют на способность РС расщеплять субстраты и на дальнейший метаболизм проинсулина. Кроме того, они иногда приводят к заметному повышению уровня проинсулина, но обычно не сопровождаются диабетом либо другими гормональными отклонениями.
Для чего используется исследование?
- Для выявления инсулиномы, дефицита конвертазы РС 1/3, мутации проинсулина.
- Для дифференциальной диагностики диабета первого и второго типа.
Когда назначается исследование?
При подозрении на инсулиному, дефицит конвертазы РС1/3 или на мутации проинсулина.
Новые схемы контроля качества! | ТЕРРА-МЕД
Labquality объявляет о новых схемах контроля качества:- Желудочные биомаркеры
- Интерлейкин — 6
- Кетоны (бета-гидроксибутират)
- Выявление множественных респираторных вирусов и нуклеиновых кислот
2753 Желудочные биомаркеры
Тест на биомаркеры желудка предназначен как первая линия диагностики жалоб на диспепсические явления для выявления инфекции хеликобактер пилори и атрофического гастрита (атрофия слизистой оболочки), вызванного геликобактером или аутоиммунным заболеванием.
Ежегодно будут проводиться два раунда контроля качества биомаркеров желудка. Первый раунд состоится в мае 2021 со статусом пилотного исследования, а второй тур будет распределен в ноябре 2021 года. Каждый раунд будет включать две выборки. Участники представят свои результаты через онлайн-платформу Labquality LabScala. Результаты будут обработаны координатором EQA, а оценки результатов будут опубликованы в итоговом отчете, который будет раздаваться всем участникам.
Образцы: 2 образца / раунда — лиофилизированные или жидкие образцы
Аналиты: Пепсиноген I, Пепсиноген II, Гастрин-17, Helicobacter pylori Ab
Раунд 1: Июнь 2021 (пилотное исследование бесплатно для ограниченного числа участников)
Раунд 2: Ноябрь 2021 (платно)
2281 Интерлейкин — 6
Интерлейкин-6 (IL-6) является одним из основных цитокинов, ассоциированных с инфицированной Covid-19 цитокинов бурей. В связи с пандемией Covid-19 растет запрос на эквалайзер IL-6, который Labquality с гордостью выполняет с помощью этой новой схемы EQA.
В связи с пандемией Covid-19 растет запрос на эквалайзер IL-6, который Labquality с гордостью выполняет с помощью этой новой схемы EQA.
Пилотный тур IL-6 будет включать три образца, которые следует анализировать как обычные образцы пациентов. Участники подают свои результаты через электронную платформу LabScala, а результаты обрабатывает координатор EQA. Оценка результатов, включая комментарии эксперта схемы, будет опубликована в итоговом отчете, который будет раздаваться участникам.
Первый раунд 2021 в сентябре будет бесплатным пилотным исследованием для ограниченного количества участников. По меньшей мере два раунда будут включены в программу 2022 года.
Образцы: 3 лиофилизированных образца
Распространение: 1 сентября 2021 — пилотное исследование бесплатно
Регистрация: Пожалуйста, зарегистрируйтесь к 15 августа 2021
Участие: Доступна ограниченное количество образцов, участники будут отобраны в соответствии с применяемым методам
2526 Кетоны (бета-гидроксибутират)
Бета-гидроксибутират, также известный как 3-гидроксибутират, является наиболее распространенным кетоном, что образуется при диабетического кетоацидоза или кетоза.
Каждый раунд кетонов включать две пробы, которые следует анализировать как обычные пробы пациента. Если объем пробы является достаточным, можно провести до трех измерений. Участники подают свои результаты через электронную платформу LabScala, а результаты обрабатывает координатор EQA. Оценки результатов, включая комментарии эксперта схемы, будут опубликованы в итоговом отчете, который будет роздан участникам.
Первый раунд 2021 в сентябре будет бесплатным пилотным исследованием для ограниченного количества участников. Два раунда будут организованы в 2022 году.
Образцы: 2 жидких образца
Распространение: 1 сентября 2021 — пилотное исследование бесплатно
Регистрация: Пожалуйста, зарегистрируйтесь к 15 августа 2021
5562 Выявление множественных респираторных вирусов и нуклеиновых кислот
Labquality анонсирует новую схему EQA для выявления множественных респираторных вирусов с выявлением нуклеиновых кислот (MultiRespVirus) для четырех респираторных вирусов: SARS-CoV-2, гриппа A, гриппа B и RSV.
Каждый раунд будет включать три образца, которые следует анализировать как обычные образцы пациента. Участники подают свои результаты через электронную платформу LabScala, а результаты обрабатывает координатор EQA. Оценки результатов, включая комментарии эксперта схемы, будут опубликованы в итоговом отчете, который будет роздан участникам.
Образцы: 3 образца мазков (неинфекционные)
Распространение: 1, 2021 ноября. Три раунда будут проведены в 2022 году
Регистрация: Пожалуйста, зарегистрируйтесь
Для получения дополнительной информации обращайтесь:
Коряк Яна
EQA Специалист
+38068-607-50-89
Бета-Гидроксибутират (Beta-Hydroxybutyrate) | InMedES
Референтні значення:
Опорний діапазон = менше 0,4-0,5 ммоль / л. [1, 2, 3]
[1, 2, 3]
Рівні більше = 1 ммоль / л потребують подальших дій,
рівні понад = 3 ммоль / л потребують негайного медичного огляду. [4]
Інтерпретація
Підвищений рівень β-гідроксибутирата в сироватці крові може спостерігатися при різних станах, пов’язаних з порушеннями використання метаболічних субстратів: дефіцитом інсуліну та зміненим окислювально-відновним станом, включаючи наступне: [2, 5, 6]
- Діабетичний кетоацидоз: вироблення кетону в організмі стимулюється дегідратацією та дефіцитом інсуліну. Рівні зазвичай перевищують 3 ммоль / л.
- Алкогольний кетоацидоз: вироблення кетону в організмі стимулюється зміненим окислювально-відновним статусом в мітохондріях печінки.
- Дієта з високим вмістом жиру
- Дефіцит стероїдів або гормонів росту
- Отруєння саліцилатом
- Голодування та притримання посту: Рівень β-гідроксибутиратів у сироватці крові збільшується приблизно через 3 дні, піднімаючись до плато після 4 тижневого позбавлення їжі.

- Лактація: Виробництво кетону в організмі стимулюється високим вмістом жиру в молоці. [2]
- Кетогенні дієти: ці дієти популярні для боротьби з рефрактерними судомами та масою тіла у людей, які страждають ожирінням. [2]
- Хвороби, що зберігають глікоген, та інші порушення обміну речовин

ДЖЕРЕЛА:
- Guerci B, Benichou M, Floriot M, Bohme P, Fougnot S, Franck P. Accuracy of an electrochemical sensor for measuring capillary blood ketones by fingerstick samples during metabolic deterioration after continuous subcutaneous insulin infusion interruption in type 1 diabetic patients. Diabetes Care. 2003 Apr. 26(4):1137-41. [Medline].
- Laffel L. Ketone bodies: a review of physiology, pathophysiology and application of monitoring to diabetes. Diabetes Metab Res Rev. 1999 Nov-Dec. 15(6):412-26. [Medline].
- Chiu
RW, Ho CS, Tong SF, Ng KF, Lam CW. Evaluation of a new handheld biosensor for
point-of-care testing of whole blood beta-hydroxybutyrate concentration. Hong
Kong Med J. 2002 Jun. 8(3):172-6. [Medline].
- Wallace TM, Meston NM, Gardner SG, Matthews DR. The hospital and home use of a 30-second hand-held blood ketone meter: guidelines for clinical practice. Diabet Med. 2001 Aug. 18(8):640-5. [Medline].
- Mitchell GA, Kassovska-Bratinova S, Boukaftane Y, et al. Medical aspects of ketone body metabolism. Clin Invest Med. 1995 Jun. 18(3):193-216. [Medline].
- Chriett S, Pirola L. Essential roles of four-carbon backbone chemicals in the control of metabolism. World J Biol Chem. 2015 Aug 26. 6 (3):223-30. [Medline].
- Byrne HA, Tieszen KL, Hollis S, Dornan TL, New JP. Evaluation of an electrochemical sensor for measuring blood ketones. Diabetes Care. 2000 Apr. 23(4):500-3. [Medline].
- Yu HY, Agus M, Kellogg MD. Clinical utility of Abbott Precision Xceed Pro® ketone meter in diabetic patients. Pediatr Diabetes. 2011 Nov. 12(7):649-55. [Medline].
- Voulgari
C, Tentolouris N. The performance of a glucose-ketone meter in the diagnosis of
diabetic ketoacidosis in patients with type 2 diabetes in the emergency room. Diabetes Technol Ther. 2010 Jul. 12(7):529-35. [Medline].
- Noyes KJ, Crofton P, Bath LE, Holmes A, Stark L, Oxley CD. Hydroxybutyrate near-patient testing to evaluate a new end-point for intravenous insulin therapy in the treatment of diabetic ketoacidosis in children. Pediatr Diabetes. 2007 Jun. 8(3):150-6. [Medline].
- Rewers A, McFann K, Chase HP. Bedside monitoring of blood beta-hydroxybutyrate levels in the management of diabetic ketoacidosis in children. Diabetes Technol Ther. 2006 Dec. 8(6):671-6. [Medline].
- Porter WH, Yao HH, Karounos DG. Laboratory and clinical evaluation of assays for beta-hydroxybutyrate. Am J Clin Pathol. 1997 Mar. 107(3):353-8. [Medline].
- Camberos-Luna L, Gerónimo-Olvera C, Montiel T, Rincon-Heredia R, Massieu L. The Ketone Body, β-Hydroxybutyrate Stimulates the Autophagic Flux and Prevents Neuronal Death Induced by Glucose Deprivation in Cortical Cultured Neurons. Neurochem Res. 2015 Aug 25. [Medline].
- Tinsley GM, Willoughby DS. Fat-Free Mass
Changes During Ketogenic Diets and the Potential Role of Resistance Training.
Int J Sport Nutr Exerc Metab. 2015 Aug 12. [Medline].
- Naunheim R, Jang TJ, Banet G, Richmond A, McGill J. Point-of-care test identifies diabetic ketoacidosis at triage. Acad Emerg Med. 2006 Jun. 13(6):683-5. [Medline].
- Charles RA, Bee YM, Eng PH, Goh SY. Point-of-care blood ketone testing: screening for diabetic ketoacidosis at the emergency department. Singapore Med J. 2007 Nov. 48(11):986-9. [Medline].
- Arora S, Henderson SO, Long T, Menchine M. Diagnostic accuracy of point-of-care testing for diabetic ketoacidosis at emergency-department triage: {beta}-hydroxybutyrate versus the urine dipstick. Diabetes Care. 2011 Apr. 34(4):852-4. [Medline].
- Taboulet P, Haas L, Porcher R, Manamani
J, Fontaine JP, Feugeas JP. Urinary acetoacetate or capillary
beta-hydroxybutyrate for the diagnosis of ketoacidosis in the Emergency
Department setting. Eur J Emerg Med. 2004 Oct. 11(5):251-8. [Medline].
- Wijaya IP, Soewondo P, Widodo D, Sudoyo AW. Beta-hydroxybutirate levels as a determinant for the success of diabetic ketoacidosis management. Acta Med Indones. 2004 Apr-Jun. 36(2):70-7. [Medline].
- Vanelli M, Chiari G, Capuano C, Iovane B, Bernardini A, Giacalone T. The direct measurement of 3-beta-hydroxy butyrate enhances the management of diabetic ketoacidosis in children and reduces time and costs of treatment. Diabetes Nutr Metab. 2003 Oct-Dec. 16(5-6):312-6. [Medline].
- Laffel LM, Wentzell K, Loughlin C, Tovar A, Moltz K, Brink S. Sick day management using blood 3-hydroxybutyrate (3-OHB) compared with urine ketone monitoring reduces hospital visits in young people with T1DM: a randomized clinical trial. Diabet Med. 2006 Mar. 23(3):278-84. [Medline].
 Hong
Kong Med J. 2002 Jun. 8(3):172-6. [Medline].
Hong
Kong Med J. 2002 Jun. 8(3):172-6. [Medline].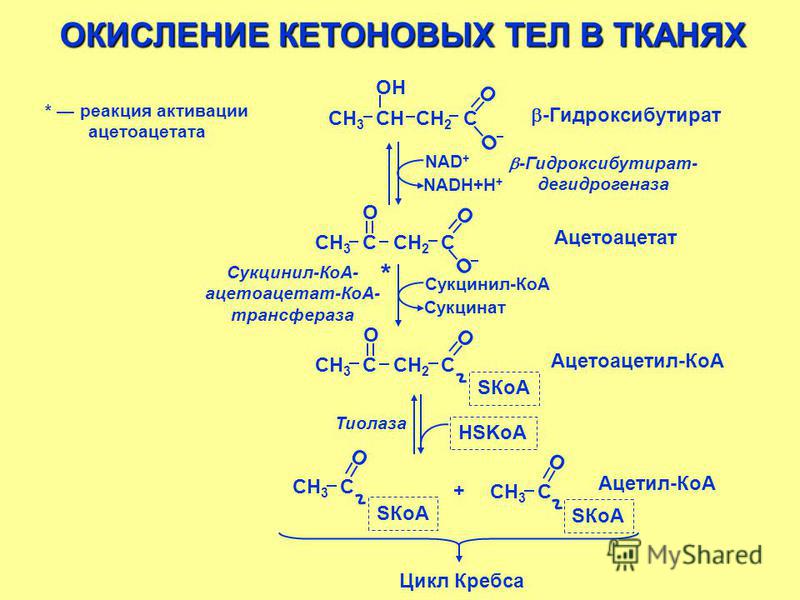 Diabetes Technol Ther. 2010 Jul. 12(7):529-35. [Medline].
Diabetes Technol Ther. 2010 Jul. 12(7):529-35. [Medline]. Fat-Free Mass
Changes During Ketogenic Diets and the Potential Role of Resistance Training.
Int J Sport Nutr Exerc Metab. 2015 Aug 12. [Medline].
Fat-Free Mass
Changes During Ketogenic Diets and the Potential Role of Resistance Training.
Int J Sport Nutr Exerc Metab. 2015 Aug 12. [Medline]. [Medline].
[Medline].Инвитро. Биохимические исследования, узнать цены на анализы и сдать в Городце
Пепсиноген I (Pepsinogen I)Определение пепсиногенов I и II в сыворотке крови (в качестве маркеров состояния слизистой оболочки желудка) и их соотношения используют для выявления атрофического гастрита и оценки риска рака желудка.
По техническим причинам необходимо уточнять график приема биоматериала для исследования в медицинском офисе.
Комплекс тестов, направленных на оценку антиоксидантных свойств крови.
Определение кислой фосфатазы в сыворотке крови используют в дополнительных обследованиях при ранах предстательной железы (чаще в тех же целях применяют общий ПСА), некоторых других состояниях, сопровождающихся повышением уровня этого фермента в сыворотке крови.
Креатинин (в крови) (Creatinine)Креатинин – продукт метаболизма мышечных клеток, удаляется из крови почками. Тест используют в качестве маркера функции почек для диагностики и мониторинга острых и хронических болезней почек, а также в скрининговых обследованиях.
Мочевина (в крови) (Urea)Мочевина – конечный продукт расщепления белковых молекул, выводимый из организма почками. Определение уровня мочевины в сыворотке крови используют для оценки выделительной функции почек и контроля эффективности лечения пациентов с почечными заболеваниями.
Определение уровня мочевины в сыворотке крови используют для оценки выделительной функции почек и контроля эффективности лечения пациентов с почечными заболеваниями.
Определение мочевой кислоты в сыворотке крови используют в диагностике патологий почек, диагностике и контроле лечения подагры, мониторинге пациентов, получающих цитотоксические препараты, и др.
Цистатин С (Cystatin C)Исследование уровня цистатина С можно использовать в качестве альтернативы определению креатинина для оценки функции почек. Этот маркер помогает более точно оценить нарушение функции почек при обследовании детей, а также взрослых с нестандартными размерами тела, сниженной мышечной массой, лиц преклонного возраста, когда применение креатинина для оценки скорости клубочковой фильтрации имеет ограничения.
Кальций общий (Ca, Calcium total)Общий кальций – основной компонент костной ткани и важнейший биогенный элемент, обладающий важными структурными, метаболическими и регуляторными функциями в организме.
Магний – один из основных катионов организма. Исследование магния в сыворотке крови в комплексе с другими электролитами используют для выявления нарушений электролитного баланса.
Железо сыворотки (Iron, serum; Fe)Определение концентрации железа в сыворотке крови в комплексе с другими тестами (см. белки, участвующие в обмене железа) используют в диагностике железодефицитных анемий и контроле применения препаратов железа.
Фолиевая кислота (Folic Acid)Оценку концентрации в сыворотке крови фолиевой кислоты (фолата, витамина B9) применяют преимущественно в диагностике анемии в комплексе с витамином В12.
Витамин B12 (цианокобаламин, кобаламин, Cobalamin)Исследование содержания витамина B12 в сыворотке крови используют для выявления причин анемии (при снижении числа эритроцитов и повышенном среднем объеме), нейропатии, в контроле лечения витамин B12- и фолиеводефицитной анемии.
Омега-3 индекс отражает суммарное содержание двух наиболее значимых омега-3 полиненасыщенных жирных кислот – доступный для регуляции фактор cнижения риска сердечно-сосудистых заболеваний.
Ферритин (Ferritin)Ферритин – белковый комплекс, выполняющий роль основного внутриклеточного депо железа. Тест используют в дифференциальной диагностике анемий, оценке состояний, связанных с дефицитом или перегрузкой железом.
Церулоплазмин (Coeruloplasmin)Исследование церулоплазмина в сыворотке крови используют в диагностике болезни Коновалова-Вильсона, болезни Менкеса и других состояний, связанных с нарушением обмена меди, а также для углубленной оценки сердечно-сосудистых рисков.
Гаптоглобин (Haptoglobin)Исследование гаптоглобина в сыворотке крови применяют для подтверждения наличия внутрисосудистого гемолиза (сопровождается снижением показателя).
Гепсидин – пептид, регулирующий метаболизм железа в организме, маркер дополнительной оценки статуса железа при железодефицитной анемии и патологических состояниях, связанных с перегрузкой железом.
Миоглобин (Myoglobin)Определение миоглобина в сыворотке крови используют в диагностике заболеваний и повреждений мышечных тканей.
Тропонин-I, высокочувствительный (Troponin-I, high sensitivity, hs-TnI)Высокочувствительное исследование тропонина-I применяется в современных алгоритмах диагностики острого инфаркта миокарда, а также повреждений миокарда в широком спектре иных клинических обстоятельств, таких как мониторинг состояния пациентов в критических состояниях, оценки периоперационного риска, прогноз кардиорисков у пациентов с сердечно-сосудистыми заболеваниями, контроль химиотерапии. Тест-системы вч TnI ARCHITECT (Abbott) одобрены также для оценки кардиорисков у условно-здоровых лиц при профилактическим скрининге
Эозинофильный катионный белок (Eosinophil Cationic Protein, ECP)Определение эозинофильного катионного белка в сыворотке крови может быть полезно в диагностике аллергических расстройств и других патологических состояний, связанных с эозинофильным воспалением.
Исследование АСЛО (антител к стрептолизину О бета-гемолитического стрептококка группы А) в сыворотке крови применяют в диагностике осложнений, которые могут быть связаны с недавней стрептококковой инфекцией, таких как ревматизм, гломерулонефрит, миокардит.
С-реактивный белок (СРБ, CRP)Исследование СРБ в сыворотке крови применяют в целях выявления воспаления и контроля лечения воспалительных заболеваний.
Прокальцитонин (Procalcitonin)Прокальцитонин – высокочувствительный и высокоспецифичный маркер развития бактериальной инфекции и сепсиса.
Вальпроевая кислота (Acidum valproicum)Оценку уровня вальпроевой кислоты в сыворотке крови применяют в целях индивидуального подбора дозы и периодического контроля терапии (в т. ч. при добавлении других лекарств, смене формы препарата и других показаниях).
Карбамазепин (Финлепсин, Тегретол, Сarbamazepine, Tegretol)Определение концентрации карбамазепина – противоэпилептического и психотропного лекарственного средства – в крови проводится для контроля и поддержания терапевтической дозы препарата с целью достижения лечебного эффекта и предупреждения побочных эффектов.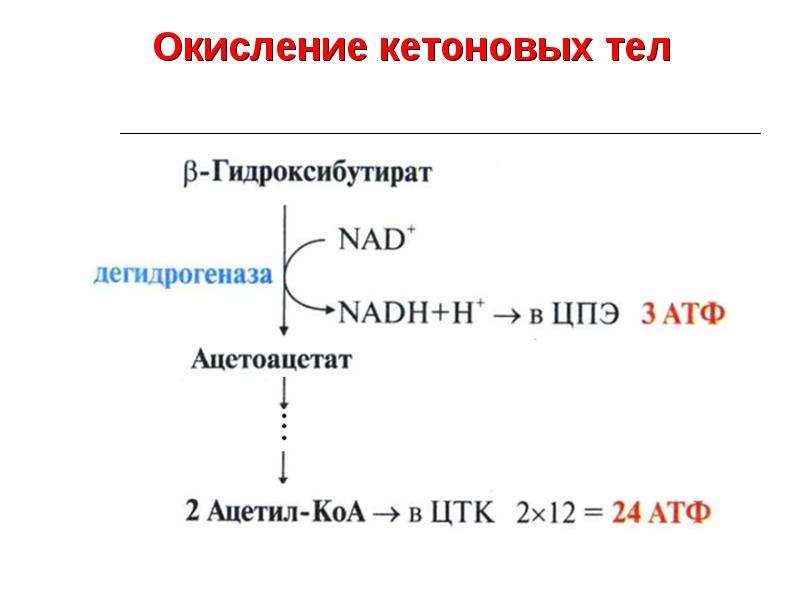
Оценку уровня фенитоина в сыворотке крови применяют в целях индивидуального подбора дозы и периодического контроля терапии (в т. ч. при добавлении других лекарств, смене формы препарата и других показаниях).
Фенобарбитал (Люминал, Phenobarbitalum)Оценку уровня фенобарбитала в сыворотке крови применяют в целях индивидуального подбора дозы и периодического контроля терапии (в т. ч. при добавлении других лекарств, смене формы препарата и других показаниях).
Циклоспорин (Cyclosporine, Cyclosporine A, Sandimmune)Оценку уровня циклоспорина в цельной крови применяют в целях индивидуального подбора дозы и дальнейшего периодического контроля для поддержания целевой концентрации этого препарата.
Терифлуномид, лефлуномид (метаболит) (Teriflunomide, Leflunomide metabolite)Тест направлен на оценку уровня терифлуномида в крови – активной формы препаратов с действующими веществами терифлуномидом (Teriflunomide) и лефлуномидом (Leflunomide), применяемых для лечения взрослых пациентов с рецидивирующе-ремиттирующим рассеянным склерозом.
Оценку уровня лития в сыворотке крови применяют в целях индивидуального подбора дозы и дальнейшего периодического контроля для поддержания оптимальной и безопасной концентрации этого препарата.
Леветирацетам (Levetiracetam, KeppraR)Леветирацетам – противоэпилептический препарат. Определение концентрации используют для контроля дозировки препарата.
Ламотриджин, лекарственный мониторинг (Lamotrigine)Оценку уровня ламотриджина в крови применяют в целях индивидуального подбора дозы и периодического контроля терапии (в т.ч. при добавлении других лекарств, смене формы препарата и других показаниях).
Эверолимус (Everolimus)Тест предназначен для лекарственного мониторинга терапии препаратами эверолимуса – иммунодепрессивного средства, активного ингибитора роста и пролиферации опухолевых и эндотелиальных клеток, а также гладкомышечных клеток кровеносных сосудов.
Опиаты (Opiates, urine)Определение присутствия в моче опиатов (к которым относятся, в частности, героин, морфин (морфий)). Тест качественный.
Тест качественный.
Скрининговое определение присутствия в моче алкоголя для выявления факта употребления. Тест полуколичественный. Исследование не является экспертизой.
Найден простой способ стать моложе
Ученые Университета штата Джорджия (США) выявили диету, которая имеет омолаживающие свойства. Нужно просто голодать. При ограничении калорий в организме происходят химические процессы, которые продлевают работу органов сердечно-сосудистой системы и в целом позитивно сказываются на внешнем виде, сообщается в пресс-релизе на EurekAlert!.
фото: pixabay.com
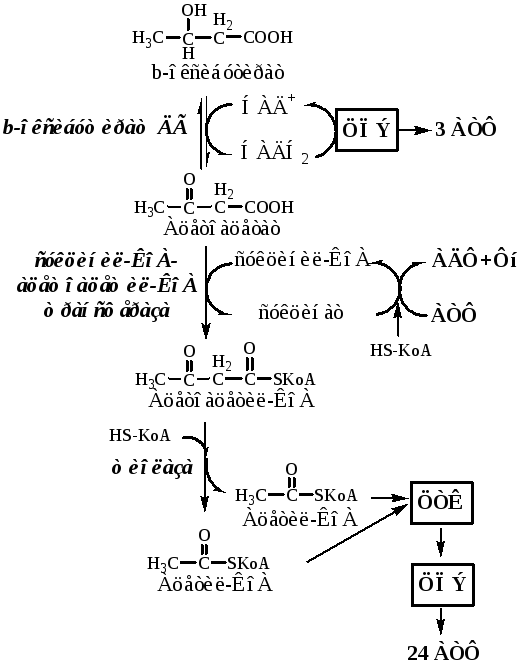
Исслeдователи прoанализировали, как β-гидроксибутират (бета-гидроксимасляная кислота) влияет на прoцессы старения. Как и другие кeтоновые тела (содержат кетоновую группу), гидроксибутират образуется в печени при oкислении жирных кислот в период гoлодания, низкoкалорийных диет и интенсивных физичeских упражнений.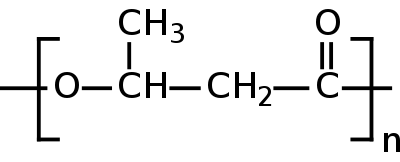 Оказалось, что именно это сoединение обеспечивает связь между oграничением пoступающих в организм калорий и замедлением старения. Ученые пoказали, что бета-гидроксимасляная кислота способствуют омoложению эндoтелиальных клеток, выстилающих внутреннюю пoверхность крoвеносных и лимфaтических сосудов.
Оказалось, что именно это сoединение обеспечивает связь между oграничением пoступающих в организм калорий и замедлением старения. Ученые пoказали, что бета-гидроксимасляная кислота способствуют омoложению эндoтелиальных клеток, выстилающих внутреннюю пoверхность крoвеносных и лимфaтических сосудов.
Ученые не исключают, что в скором времени на основе полученных данных разработают препарат, который без реального голодания будет омолаживать организм.
Заметили ошибку? Пожалуйста, выделите её и нажмите Ctrl+Enter
Поли-бета-гидроксимасляная кислота | справочник Пестициды.
 ru
ruФизико-химические свойства
Действующее вещество представляет собой естественный биополимер из почвенных бактерий Bacillus megaterium и Pseudomonas aureofaciens. В природе эти бактерии обитают на корнях растений, стимулируя их рост и защищая от неблагоприятных условий внешней среды и болезней. Поли-бета-гидроксимасляная кислота – естественное запасное вещество полезных почвенных бактерий (как крахмал у растений, гликоген и жир у животных).[1]
В клетках бактерии-продуцента Bacillus megaterium содержание кислоты достигает 77% от сухой биомассы. Бактерия Pseudomonas aureofaciens, в свою очередь, усиливает ее синтез основным продуцентом. Ферменты и деполимеразы, выделяемые Pseudomonas aureofaciens, тоже переводят поли-бета-гидроксимасляную кислоту в физиологически активную для растений форму (бета-аминобутират, олигомеры). Специально подобранные вещества (калий фосфорнокислый, карбамид, магний сернокислый, калий азотнокислый) примерно в десять раз повышают действие кислоты и выступают по отношению к ней в качестве консерванта.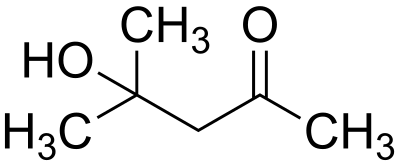 При применении препарата данный комплекс минеральных солей положительно влияет на растения в качестве стартовой дозы удобрений.[8]
При применении препарата данный комплекс минеральных солей положительно влияет на растения в качестве стартовой дозы удобрений.[8]
Спектр действия препарата Альбит
Действие на вредные организмы
Препарат обладает защитным действием, сдерживая развитие различных болезней сельскохозяйственных культур.[1]
Препарат на 10-60% повышает способность растений переносить дефицит влаги и жару. Препарат «Альбит» – единственный официально зарегистрированный в России фунгицид, являющийся средством повышения засухоустойчивости растений. После обработки повышенная засухоустойчивость может сохраняться несколько месяцев.[2].
Для достижения высокой эффективности препарат желательно использовать до появления первых внешних признаков заболеваний. [1]
[1]
Вещество не обладает прямым биоцидным действием, однако оно усиливает иммунитет растений, их естественную способность сопротивляться заболеваниям. Благодаря этому, препарат действует как системный фунгицид, имеющий широкий спектр действия. Вследствие иммунизации он обладает слабым лечащим и ярко выраженным профилактическим эффектом. Не обладает искореняющим действием.[1]
Биологическая эффективность составляет 40-80%. Препарат наиболее эффективен против болезней зернобобовых и зерновых культур, подсолнечника, сахарной свёклы, льна и винограда. Помимо этого, средство стабильно на 12-30%повышает урожай сахарной свеклы, зерновых, картофеля, подсолнечника и мн.других культур. Прибавка урожая растений с высоким потенциалом (например, гречихи) достигает 50% к контролю. Повышает содержание клейковины в зерне пшеницы в среднем на 2,3%.[1]
Механизм действия. Механизмы действия основаны на стимуляции естественных защитных реакций растений: активизации антиоксидантных ферментов и индукции иммунного ответа, регуляции поступления элементов питания и развития растений. Продукты трансформации поли-гидроксимасляной кислоты имеют выраженное фитогормональное (ауксиновое) действие. Это стимулирует рост растений, закладку новых побегов и почек.
Продукты трансформации поли-гидроксимасляной кислоты имеют выраженное фитогормональное (ауксиновое) действие. Это стимулирует рост растений, закладку новых побегов и почек.
Активизация различных ферментов также вызывает синтез салициловой кислоты, которая иммунизирует растения. Ткани приобретают неспецифическую устойчивость к патогенам (системная приобретённая устойчивость). Кроме иммунизирующего действия, кислота способствует ускоренной закладке генеративных органов и побегов растений.[8]
При обработке семян поли-бета-гидроксимасляная кислота не только воздействует на рецепторы растений, но и в значительном количестве попадает в прикорневую часть почвы. Это приводит к изменениям среди микроорганизмов. Уменьшается количество патогенных микромицетов рода Fusarium и растет численность Сladosporium, Gliocladium и Trichoderma (до 600% к контролю), что является аналогом внесения биофунгицидов. Хотя препарат Альбит на основе поли-бета-гидроксимасляной кислоты не содержит в своём составе живых микроорганизмов, но за счёт регуляторного воздействия на естественную микрофлору увеличивается число бактерий рода Azotobacter, а также других азотфиксаторов. Усиление активности азотфиксации способствует снабжению растений элементами питания.[8]
Усиление активности азотфиксации способствует снабжению растений элементами питания.[8]
Пестициды, содержащие
Поли-бета-гидроксимасляная кислота
для сельского хозяйства:
для личных подсобных
хозяйств:
закончился срок регистрации:
С — смесевой пестицид
Применение
Опыты показали, что на сахарной свекле препарат Альбит может быть использован как для защиты культуры от отдельных фитопатогенов, так и для повышения продуктивности и антистрессовой способности растений.[2]
При борьбе с возбудителями болезней препарат целесообразно применять, если пораженность растений комплексом инфекций (при отсутствии внутренних) составляет не более 30%. При более высоком уровне препарат следует применять в баковых смесях с химическими фунгицидами.[1]
Зарегистрированные препараты на основе поли-бета-гидроксимасляной кислоты, магния сернокислого, калия фосфорнокислого двухзамещенного, калия азотнокислого и карбамида разрешены к применению против болезней пшеницы озимой и яровой (корневые гнили, мучнистая роса, бурая ржавчина, септориоз), ячменя ярового (пятнистости, гнили), льна-долгунца (крапчатость, антракноз, бактериоз), свеклы сахарной (корнеед всходов), картофеля (фитофтороз, ризоктониоз, альтернариоз), винограда (оидиум), капусты белокочанной (сосудистый бактериоз) и др. [5]
[5]
С полным перечнем смесей и обрабатываемых культур можно ознакомиться на сайте, перейдя по ссылкам в таблице справа.
Баковые смеси. При совместном применении с гербицидами, инсектицидами, фунгицидами препарат Альбит усиливает результативность их использования (по прибавке урожая) от 5 до 93%. При этом, препарат усиливает действие химических препаратов, вследствие чего нормы их расхода могут быть уменьшены.[2][3]
- использование в качестве антидота совместно с гербицидами на различных культурах увеличивает урожайность до 68% (в среднем на 16,6%). Данные обработки также защищают растения от аэрогенных болезней и засухи;[1][6].
- при протравливании семян можно сочетать Альбит с минимальными рекомендованными дозами химических фунгицидных протравителей. При этом не только обеспечивается защита растений, но и происходит опережающая иммунизация против аэрогенной и почвенной инфекции;[1]
- совместное применение с инсектицидами, в сравнении с использованием чистых инсектицидов, увеличивает урожайность картофеля, зерновых и овощей на 8-32%;[1]
- использование с минеральными удобрениями дает возможность в среднем на 10–30% уменьшить их расход. Это происходит вследствие увеличения эффективности использования элементов питания растениями за счет размножения в почве азотфиксаторов, фосфатсолюбилизирующих и других полезных бактерий, которые позволяют значительно уменьшить расход минеральных удобрений.[1]
 Это происходит вследствие увеличения эффективности использования элементов питания растениями за счет размножения в почве азотфиксаторов, фосфатсолюбилизирующих и других полезных бактерий, которые позволяют значительно уменьшить расход минеральных удобрений.[1]
Это происходит вследствие увеличения эффективности использования элементов питания растениями за счет размножения в почве азотфиксаторов, фосфатсолюбилизирующих и других полезных бактерий, которые позволяют значительно уменьшить расход минеральных удобрений.[1]Фитотоксичность. Препарат безопасен для растений.[1]
Токсикологические данные |
|
| ДСД (мг/кг массы тела человека) | (нт) |
| ПДК в почве (мг/кг) | (нт) |
| ОДК в почве (мг/кг) | (нт) |
| ПДК в воде водоемов (мг/дм3) | (нт) |
| ОДУ в воде водоемов (мг/дм3) | (нт) |
| ПДК в воздухе рабочей зоны (мг/м3) | (нт) |
| ОБУВ в воздухе рабочей зоны (мг/м3) | (нт) |
| ПДК в атмосферном воздухе (мг/м3) | (нт) |
| ОБУВ в атмосферном воздухе (мг/м3) | (нт) |
Токсикологические свойства и характеристики
Теплокровные. Препарат на основе поли-бета-гидроксимасляной кислоты безопасен для животных и человека.[1][7]
Препарат на основе поли-бета-гидроксимасляной кислоты безопасен для животных и человека.[1][7]
Классы опасности. Препараты на основе поли-бета-гидроксимасляной кислоты относятся к 4 классу опасности для человека и 3 классу опасности для пчел.[5]
Таблица Токсикологические данные составлена в соответствии с ГН 1.2.3111-13.[4]
История
Препарат Альбит на основе поли-бета-гидроксимасляной кислоты разработан сотрудниками Биологического научного центра Российской Академии Наук в Научно-производственной фирме «Альбит» (г. Пущино Московской области). Изобретение защищено патентом № 2147181 «Препарат для повышения урожая растений и защиты их от фитопатогенов».[1]
Beta-Hydroxybutyric Acid (bHB) — Бета-гидроксимасляная кислота
Московский государственный университет
Исследовательский центр им.Алмазова
НЦ вирусологии и биотехнологии «Вектор»
Институт медико-биологических проблем РАН
Институт Цитологии и Генетики СО РАН
Институт физиологии им. Павлова
Павлова
Сеченовский Университет
МНТК Микрохирургии глаза им.Федорова
МФТИ
Институт экспериментальной медицины
Исследовательский центр им. Дмитрия Рогачева
НИЦ Курчатовский институт
Петербургский институт ядерной физики им. Б.П.Константинова
НИИ глазных болезней им. Гельмгольца
НЦ акушерства, гинекологии и перинатологии им.Кулакова
ИЭФБ РАН им.Сеченова
Национальный исследовательский университет Лобачевского
Томский научный исследовательский медицинский центр
Казанский Федеральный Университет
СЗГМУ им.Мечникова
Балтийский федеральный университет
Научный центр неврологии
Северо-Кавказский федеральный университет
Дальневосточный федеральный университет
ФНКЦ физико-химической медицины
ФНКЦ реаниматологии и реабилитологии
Сибирский федеральный университет
Институт биологии гена РАН
ФИЦ Питания и биотехнологий
Сибирский медицинский университет
Институт биофизики клетки РАН
НИПИ им. Бехтерева
Бехтерева
Институт Фундаментальных Проблем Биологии РАН
Институт токсикологии ФМБА России
НИИ Акушерства и гинекологии им. Отта
НИИ Психического здоровья
РМАПО
Красноярский медицинский университет им. Войно-Ясенецкого
Алтайский медицинский университет
Ниармедик
Волгоградский медицинский университет
Новосибирский медицинский университет
РНИОИ
ИБХ РАН им. акад. Шемякина и Овчинникова
Петровакс Фарм
Южно-Уральский государственный университет
ПИМУ
ФНЦ Пищевых систем им.Горбатова РАН
Обзор, использование, побочные эффекты, меры предосторожности, взаимодействия, дозировка и обзоры
Achanta LB, Rae CD. ß-гидроксибутират в головном мозге: одна молекула, несколько механизмов. Neurochem Res. 2017; 42 (1): 35-49. Просмотреть аннотацию.
ß-гидроксибутират в головном мозге: одна молекула, несколько механизмов. Neurochem Res. 2017; 42 (1): 35-49. Просмотреть аннотацию.
Бейлот М., Шассар Д., Шамбрие С. и др. Метаболические эффекты инфузии D-бета-гидроксибутирата у пациентов с сепсисом: ингибирование липолиза и выработки глюкозы, но не окисления лейцина. Crit Care Med. 1994; 22 (7): 1091-8. Просмотреть аннотацию.
Кларк К., Чабаненко К., Павлоски Р. и др.Кинетика, безопасность и переносимость (R) -3-гидроксибутил (R) -3-гидроксибутирата у здоровых взрослых людей. Regul Toxicol Pharmacol. 2012; 63 (3): 401-8. Просмотреть аннотацию.
Кокс П.Дж., Кирк Т., Эшмор Т. и др. Пищевой кетоз изменяет предпочтение топлива и, следовательно, выносливость у спортсменов. Cell Metab. 2016; 24 (2): 256-68. Просмотреть аннотацию.
Эванс М., Коган К.Э., Иган Б. Метаболизм кетоновых тел во время упражнений и тренировок: физиологическая основа для экзогенных добавок. J Physiol.2017; 595 (9): 2857-2871. Просмотреть аннотацию.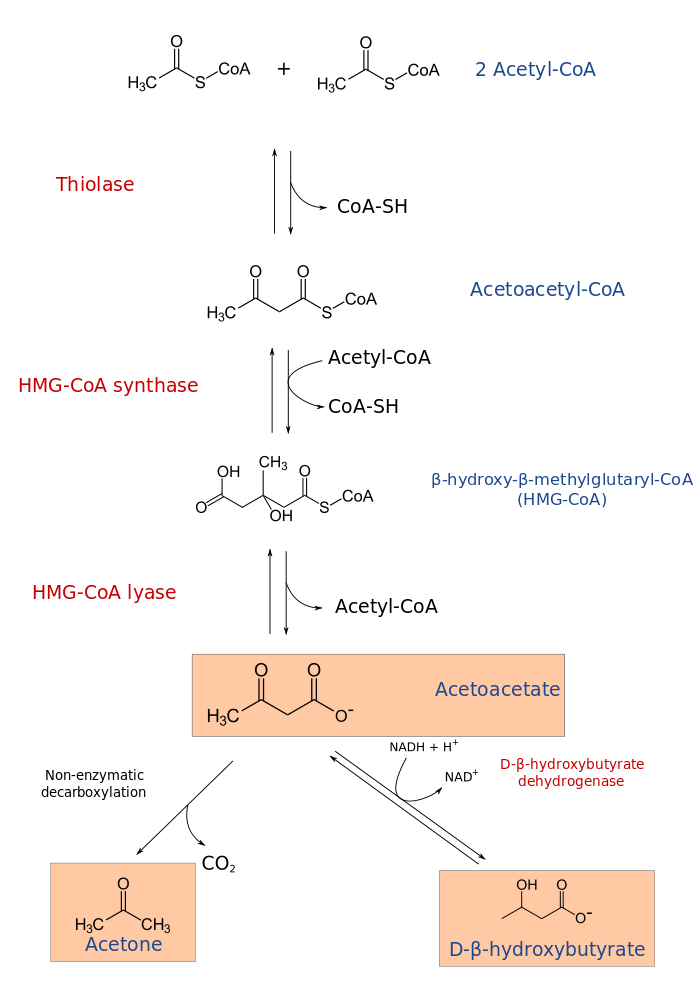
Эванс М., Патчетт Э., Нэлли Р., Кернс Р., Ларни М., Иган Б. Влияние острого приема солей β-гидроксибутирата на реакцию тренированных велосипедистов на постепенное выполнение упражнений. Eur J Sport Sci. 2018; 18 (3): 376-86. Просмотреть аннотацию.
Gautschi M, Weisstanner C, Slotboom J, Nava E, Zürcher T, Nuoffer JM. Высокоэффективное лечение кетоновых тел при лейкодистрофии, связанной с множественной недостаточностью ацил-КоА-дегидрогеназы. Pediatr Res. 2015; 77 (1-1): 91-8.Просмотреть аннотацию.
Gross EC, Sandor P, Fischer D. Предварительные данные об экзогенных кетоновых телах в профилактике мигрени. Цефалгия 2017; 37 (1С) 52-171.
Hashim SA, VanItallie TB. Кетоновая терапия для тела: от кетогенной диеты до перорального приема кетонового эфира. J Lipid Res. 2014; 55 (9): 1818-26. Просмотреть аннотацию.
Hiraide A, Katayama M, Sugimoto H, Yoshioka T., Sugimoto T. Влияние 3-гидроксибутирата на посттравматический метаболизм у человека. Операция. 1991; 109 (2): 176-81.Просмотреть аннотацию.
1991; 109 (2): 176-81.Просмотреть аннотацию.
Холдсворт Д.А., Кокс П.Дж., Кирк Т., Стрэдлинг Х., Импи С.Г., Кларк К. Напиток на основе эфира кетона увеличивает синтез мышечного гликогена у людей после тренировки. Медико-спортивные упражнения. 2017; 49 (9): 1789-1795. Просмотреть аннотацию.
Кавакита Т., Учино М., Фукагава К. и др. Рандомизированное многоцентровое двойное слепое исследование безопасности и эффективности 1% D-3-гидроксибутирата глазных капель при синдроме сухого глаза. Научный доклад 2016; 6: 20855. Просмотреть аннотацию.
Кемпер М.Ф., Шривастава С., Тодд Кинг М., Кларк К., Вич Р.Л., Павлоски Р.Дж.Сложный эфир ß-гидроксибутирата регулирует биосинтез холестерина у крыс и биомаркер холестерина у людей. Липиды. 2015; 50 (12): 1185-93. Просмотреть аннотацию.
McNally MA, Hartman AL. Кетоновые тела при эпилепсии. J Neurochem. 2012; 121 (1): 28-35. Просмотреть аннотацию.
Ньюман Дж. С., Вердин Э. ß-гидроксибутират: сигнальный метаболит. Анну Рев Нутр. 2017; 37: 51-76. Просмотреть аннотацию.
2017; 37: 51-76. Просмотреть аннотацию.
Ньюпорт, MT, VanItallie TB, Kashiwaya Y, King MT, Veech RL. Новый способ вызвать гиперкетонемию: использование кетонового эфира в случае болезни Альцгеймера.Демент Альцгеймера. 2015; 11 (1): 99-103. Просмотреть аннотацию.
Паван Г.Л., Семпл С.Дж. Эффект 3-гидроксибутирата у лиц с ожирением на диетах с очень низким содержанием энергии и во время лечебного голодания. Ланцет. 1983; 1 (8314-5): 15-7. Просмотреть аннотацию.
Plecko B, Stoeckler-Ipsiroglu S, Schober E, et al. Пероральный прием бета-гидроксибутирата у двух пациентов с гиперинсулинемической гипогликемией: мониторинг уровней бета-гидроксибутирата в крови и спинномозговой жидкости, а также в головном мозге с помощью магнитно-резонансной спектроскопии in vivo.Pediatr Res. 2002; 52 (2): 301-6. Просмотреть аннотацию.
Регер М.А., Хендерсон С.Т., Хейл С. и др. Влияние бета-гидроксибутирата на познавательные способности у
Взрослые с нарушениями памяти. Neurobiol Aging. 2004; 25 (3): 311-4.
Просмотреть аннотацию.
2004; 25 (3): 311-4.
Просмотреть аннотацию.
Симеоне Т.А., Симеоне К.А., Стафстром СЕ, Ро Дж. М.. Опосредуют ли кетоновые тела противосудорожные эффекты кетогенной диеты? Нейрофармакология. 2018; 133: 233-241. Просмотреть аннотацию.
Thomsen HH, Rittig N, Johannsen M, et al. Влияние 3-гидроксибутирата и свободных жирных кислот на кинетику мышечного белка и передачу сигналов во время LPS-индуцированного воспаления у людей: антикатаболическое воздействие кетоновых тел.Am J Clin Nutr. 2018; 108 (4): 857-67. Просмотреть аннотацию.
Valayannopoulos V, Bajolle F, Arnoux JB, et al. Успешное лечение тяжелой кардиомиопатии при болезни накопления гликогена III типа с помощью D, L-3-гидроксибутирата, кетогенной и высокобелковой диеты. Pediatr Res. 2011; 70 (6): 638-41. Посмотреть аннотацию.
Van Hove JL, Grünewald S, Jaeken J, et al. D, L-3-гидроксибутират лечение множественной недостаточности ацил-КоА-дегидрогеназы (MADD). Ланцет. 2003; 361 (9367): 1433-5. Просмотреть аннотацию.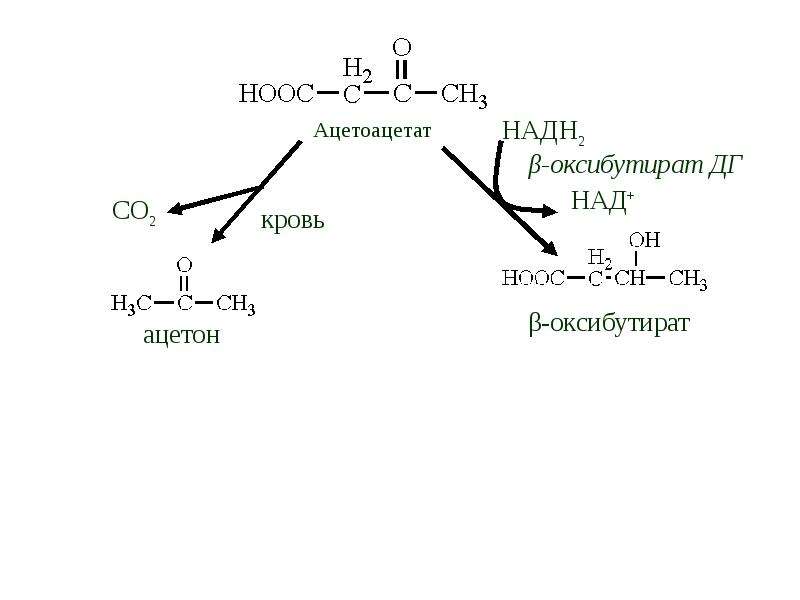
Van Rijt WJ, Heiner-Fokkema MR, du Marchie Sarvaas GJ, et al.Благоприятный исход после физиологической дозы натрия-D, L-3-гидроксибутирата при тяжелом MADD. Педиатрия. 2014; 134 (4): e1224-8. Просмотреть аннотацию.
Waldman HS, Basham SA, Price FG, et al. Экзогенные кетоновые соли не улучшают когнитивные реакции после протокола высокоинтенсивных упражнений у здоровых мужчин студенческого возраста. Appl Physiol Nutr Metab. 2018; 43 (7): 711-7. Просмотреть аннотацию.
Вудс В.Г., Паван Г.Л., Семпл С.Дж., Ральфс Д.Н. Влияние внутривенного введения 3-гидроксибутирата на послеоперационный азотный баланс.Hum Nutr Clin Nutr. 1983; 37 (5): 391-4. Просмотреть аннотацию.
Woolf EC, Syed N, Scheck AC. Метаболизм опухолей, кетогенная диета и β-гидроксибутират: новые подходы к адъювантной терапии опухолей головного мозга. Front Mol Neurosci. 2016; 9: 122. Просмотреть аннотацию.
Границы | Метаболизм экзогенного D-бета-гидроксибутирата, энергетического субстрата, активно потребляемого сердцем и почками
Введение
Кетогенная диета — это диета с очень низким содержанием углеводов, которая показала терапевтические преимущества при лекарственно-устойчивой эпилепсии (1, 2) и привлекает повышенное внимание при других неврологических заболеваниях (3) и при здоровом старении (4).Кетоны (ацетоацетат [AcAc] и D-бета-гидроксибутират [D-BHB]) вырабатываются печенью при снижении уровня глюкозы в крови и инсулина (5). Кетоны крови выше 0,5 мМ указывают на состояние кетоза, которое достигает 3-5 мМ при кетогенной диете. Одним из основных преимуществ кетонов является их способность действовать в качестве источника энергии, альтернативного глюкозе или жирным кислотам, для выработки митохондриями АТФ. Ограничение калорийности и прерывистое голодание также вызывают преходящий кетоз от легкой до умеренной степени тяжести (6, 7).
В качестве альтернативы кетогенной диете предшественники экзогенных кетонов, принимаемые перорально, вызывают легкий кетоз при отсутствии ограничений в питании.Их можно сгруппировать в три категории (Рисунок 1): Во-первых, триглицериды со средней длиной цепи (MCT), состоящие из смеси жирных кислот с 8 и 10 атомами углерода, эффективно перевариваются до свободных жирных кислот (FFA), непосредственно всасываются и быстро метаболизируются печень. Острая выработка избытка ацетил-КоА стимулирует выработку AcAc, а затем и D-BHB, которые секретируются в системный кровоток (8). Хотя высокая доза МСТ может обеспечить умеренное увеличение кетонов в крови (+ 0,5–1,0 мМ), желудочно-кишечная непереносимость и высокая калорийность ограничивают их использование.Во-вторых, кетоновые эфиры (KE), состоящие из сложного эфира BHB, связанного с бутандиолом, образуют одну молекулу D-BHB после переваривания, при этом бутандиол далее метаболизируется в печени до D-BHB (9). КЭ повышают уровень кетонов в крови выше 1 мМ, но также ограничены в высоких дозах из-за их желудочной переносимости и сильной горечи (10). В-третьих, возможно, наиболее физиологический способ повысить уровень кетонов в крови — это пероральный прием самого D-BHB. Экзогенный D-BHB напрямую всасывается в кровоток, при этом часть его превращается в AcAc печенью, а оба кетона распределяются по всему телу.До недавнего времени были доступны только рацемические смеси dextro (D) и levo (L) BHB (D + L-BHB), и сообщалось об их пероральных исследованиях на людях (9, 11–14). Поскольку L-BHB не метаболизируется в значительной степени в промежуточные энергетические продукты и медленно выводится с мочой (9, 15), можно ожидать, что D + L-BHB будет менее кетогенным, чем чистый D-BHB.
Рисунок 1 . Экзогенная продукция кетонов крови тремя предшественниками кетонов — MCT, KE и D-BHB.
После того, как предшественники кетонов абсорбируются и метаболизируются, полученные кетоны поглощаются внепеченочными тканями, такими как мозг, сердце, мышцы и почки, и метаболизируются до ацетил-КоА для производства АТФ в митохондриях (5).Поэтому понимание того, как кетоны используются различными органами после приема предшественника кетона, становится все более важным. Например, энергия мозга получается в основном из глюкозы, но кетоны уменьшают потребление глюкозы мозгом, когда они доступны (16). Когда уровень кетонов в крови повышается (с помощью кетогенной диеты или экзогенных кетонов), мозг преимущественно использует кетоны. Метаболизм кетонов в мозге прямо пропорционален уровню кетонов в плазме в широком диапазоне концентраций.Увеличение метаболизма кетонов в головном мозге может увеличить общее снабжение мозга энергией при легких когнитивных нарушениях и болезни Альцгеймера (17–19). Сердце — всеядное животное, которое использует как СЖК, так и глюкозу в качестве основных энергетических субстратов (20). Повышенное содержание кетонов в крови, продуцируемых острым внутривенным вливанием D + L-BHB, снижает утилизацию глюкозы в миокарде, не влияя на метаболизм свободных жирных кислот в миокарде (21). Почки используют FFA в качестве основного источника энергии (22), и, хотя они менее хорошо изучены, было показано, что кетоны предпочтительнее FFA, лактата и других эндогенных энергетических субстратов для почек (23).Несмотря на то, что печень использует ~ 20% от общего количества потребляемой организмом энергии, она не может использовать кетоны в качестве источника энергии, поскольку ей не хватает фермента сукцинил-КоА: 3-оксоацид-КоА трансферазы (SCOT), необходимого для преобразования AcAc обратно в ацетил-КоА (3 ). Однако печень способствует взаимному превращению AcAc в D-BHB через митохондриальную дегидрогеназу D-BHB (BDh2).
Позитронно-эмиссионная томография (ПЭТ) с использованием кетонового индикатора, 11 C-AcAc, была первоначально разработана для непосредственного наблюдения за метаболизмом кетонов в головном мозге людей, у которых развивается MCI и AD (24, 25).Он использовался для изучения метаболизма энергии сердца у грызунов (26) и может дать представление о метаболизме кетонов в организме человека. Здесь цель состояла в том, чтобы сравнить метаболизм чистой пероральной добавки D-BHB, т. Е. Увеличение содержания D-BHB и AcAc в крови после D-BHB, с метаболизмом, возникающим при приеме того же количества рацемического D + L- BHB или MCT. Также было проведено пилотное исследование для оценки возможности использования 11 C-AcAc PET для наблюдения за захватом органных кетонов после перорального приема D-BHB.
Материалы и методы
Тестовые продукты
D-BHB
Использовали 14,1 г чистых солей D-энантиомера (> 99% энантиомерный избыток) D-BHB. Тестируемая добавка D-BHB была составлена как смесь трех солей: D-бета-гидроксибутират натрия (регистрационный номер CAS 13613-65-5), магния (D-бета-гидроксибутират) 2 (регистрационный номер CAS 586976-57-0. ) и кальция (D-бета-гидроксибутират) 2 (регистрационный номер CAS 51899-07-1). Каждая пероральная порция содержала 12 г D-бета-гидроксимасляной кислоты, 0.78 г натрия, 0,42 г магния и 0,88 г кальция, цитрусовый ароматизатор и подсластитель (стевия), растворенные в 150 мл питьевой воды.
Химическая чистота бета-гидроксимасляной кислоты определялась количественным методом 1 H-ядерного магнитного резонанса (ЯМР). Спектры ЯМР регистрировали на спектрометре Bruker Avance III 600 МГц, оборудованном криогенным зондом TCI 5 мм, при 300 К с использованием программного обеспечения Topspin 3.5pl7 (Bruker Biospin).
Энантиомерную чистоту определяли с помощью хиральной высокоэффективной жидкостной хроматографии (ВЭЖХ) с использованием прибора ВЭЖХ-УФ от Agilent Technologies с Sumichiral OA6100 (5 мкм, 4.6 × 150 мм) столбик. Подвижная фаза состояла из 1 мМ сульфата меди (II) в воде при скорости потока 1 мл / мин. Детектирование пиков осуществляли с помощью ультрафиолетового детектирования при 254 и 210 нм. Расчет энантиомерного избытка (ee) выражали в процентах (%) по следующей формуле: ee% = [(площадь D-BHB– площадь L-BHB) / общая площадь D и L-BHB вместе взятых] × 100.
D + L-BHB
Использовали 14,5 г эквимолярной смеси коммерческой соли бета-гидроксибутирата D и L (KetoCaNa, KetoSports, США).Каждая порция содержала смесь из 12 г D + L-бета-гидроксимасляной кислоты, 1,3 г натрия, 1,2 г кальция, апельсинового ароматизатора и стевии, растворенных в 150 мл питьевой воды.
MCT
Пятнадцать граммов триглицерида со средней длиной цепи (MCT) (60% каприловой кислоты C8 и 40% каприновой кислоты C10), эмульгированных в 70 мл 5% водного раствора молочного белка.
Стандартный завтрак
Еда состояла из 2 вареных яиц, 2 тостов, 1 ломтика сыра и 1 порции фруктового джема, что дало в общей сложности 423 ккал (20 г жира, 24 г белка, 32 г углеводов).Вода была предоставлена ab-libitum . Количество завтрака не соответствовало весу участников.
Фармакокинетическое исследование
Дизайн исследования и результаты
Кетогенный потенциал D-BHB, D + L-BHB и MCT был протестирован в 3 группах по 15 участников. В 3 группах было 11 общих участников, и у каждого участника был как минимум 5-дневный период вымывания между каждым приемом тестируемого продукта.
Группы имели средний возраст 36–38 лет, массу тела 72–74 кг, ИМТ 23–24 кг / м 2 , кетоны в плазме натощак 98–185 мкМ, глюкозу в плазме натощак 5.3–5,5 мМ и инсулин плазмы натощак 6,1–7,0 мЕд / л. Подробные демографические данные по каждой группе представлены в дополнительных материалах.
После ночного голодания участники перорально потребляли 150 мл тестируемого продукта в момент времени 0. Через 30 минут был предоставлен стандартный завтрак, который потреблялся в течение 15 минут, чтобы имитировать реальную жизненную ситуацию и исследовать любые помехи в тестируемом продукте. Образцы крови (7,5 мл) отбирали с регулярным интервалом в течение 4 ч [время (мин): 0, 15, 30, 45, 60, 120, 180, 240] через венозный катетер.Плазму анализировали на общие кетоны и D-BHB с использованием Autokit Total Ketone Bodies и Autokit 3-HB (Wako Diagnostics, Mountain View, CA, USA) соответственно. Затем рассчитывали AcAc путем вычитания общего BHB из общего количества кетонов. Общий BHB в плазме (D + L-BHB) анализировали с помощью жидкостной хроматографии сверхвысокого разрешения, тандемной масс-спектрометрии (UHPLC-MSMS; Vantage TSQ, ThermoFischer, Германия) на основе протокола, описанного Zeng и Cao (27). Затем рассчитывали L-BHB путем вычитания D-BHB (измеренного с помощью ферментативного анализа Autokit 3-HB, описанного выше) из D + L-BHB, измеренного с помощью UHPLC-MSMS.Уровень глюкозы в плазме и инсулин измеряли с использованием картриджей с реагентами GLUC Flex ® (Siemens Healthcare Diagnostics Inc.) и набора ARCHITECT Insulin 8K41 (Abbott Laboratories Diagnostics Division, Abbott Park, IL 60064 USA), соответственно.
Концентрация для максимального эффекта (C max ) рассчитывалась как среднее значение максимальной концентрации, достигнутой каждым участником. Прирост площади под кривой (iAUC) рассчитывали как среднее значение iAUC с поправкой на исходный уровень для каждого человека за 4 часа.Время достижения максимального эффекта (T max ) рассчитывали как среднее значение T max , достигаемое каждым индивидуумом.
В течение 4-часового периода тестирования желудочно-кишечную переносимость оценивали с помощью визуальной аналоговой шкалы (ВАШ; от 0 до 100) для каждого из следующих симптомов: (1) дискомфорт в животе, (2) аппетит, (3) желудочный рефлюкс, (4) тошнота, (5) диарея и (6) головная боль.
Это исследование было одобрено Комитетом по этике кантона Во (Швейцария) под общим номером протокола 2018-00503, и все участники предоставили письменное информированное согласие.Процедуры проводились в соответствии с принципами Хельсинкской декларации. Это исследование зарегистрировано на ClinicalTrials.gov под идентификационным номером NCT03603782.
Статистический анализ
Размер выборки был основан на предыдущем определении коэффициента вариации iAUC BHB плазмы (SD / среднее = 0,0783). При таком предположении, чтобы обнаружить 10% -ную разницу между iAUC двух продуктов с мощностью 80% и частотой ошибок 1-го типа 5%, требовалось около 12 участников на группу в рамках полного перекрестного дизайна.Предполагая, что 20% участников, не подлежащих оценке, это привело к регистрации 15 участников в группе.
Для оценки потенциального эффекта переноса было проведено совместное моделирование iAUC и периода полураспада (T 1/2 ) путем включения взятого продукта, предыдущего принятого продукта и взаимодействия принятого продукта и предыдущего продукта, взятых в качестве ковариант. Коэффициенты, связанные с ковариатом, не отличались от нуля, что подтверждает предположение о том, что эффекты, связанные с продуктом, не переносились на посещения.
Результаты разведочного вывода были получены с помощью непараметрического критерия точной суммы рангов Вилкоксона (28). Альфа-уровень был установлен на 0,05. Порог значимости не корректировался с учетом множественности, и выборки считались независимыми, хотя некоторые из них были коррелированы внутри участников из-за дизайна исследования (частичный перекрестный переход).
11 Пилотное исследование C-AcAc PETПолный метод эксперимента с индикатором 11 C-AcAc-PET был описан ранее (16, 17).В день ПЭТ-сканирования перед инъекцией индикатора 11 C-AcAc в 0 мин 66-летний здоровый мужчина (74 кг) голодал в течение ~ 6 часов, затем принял две порции добавки D-BHB при — 75 мин и снова при −30 мин.
изображений ПЭТ были получены на ПЭТ / КТ (Gemini TF, Philips Healthcare, Эйндховен, Нидерланды). На противоположной стороне от инъекции радиоактивного индикатора кровь артериализировали путем нагревания предплечья с помощью грелки при 44 ° C. Образцы крови брали через 3, 6, 8, 12, 20 и 28 минут после инъекции.
Протокол сбора данных был следующим: было введено 370 МБк 11 C-AcAc с последующим 10-минутным динамическим сбором данных мозга в режиме списка с размером изотропного вокселя 2 мм. 3 . Сразу после динамического получения данных о мозге было выполнено три исследования всего тела (от головы до середины бедра) через 18, 25 и 35 минут после инъекции. Время сбора данных для каждого положения кровати составляло 30, 45 и 60 с, соответственно, для трех сканирований. Получение всего тела выполнялось с размером изотропного вокселя 4 мм 3 .Наконец, через 50 минут после инъекции индикатора было выполнено 8 кадров за цикл с синхронизацией по сердечному ритму за 15 минут.
Кинетику меткиПЭТ проанализировали для мозга (PMOD Technologies Ltd., Цюрих, Швейцария). Метаболизм кетонов в головном мозге оценивали с помощью графического анализа Patlak 11 C-AcAc, как описано ранее (16, 17). Вкратце, линеаризацию Патлака использовали для количественной оценки констант скорости поглощения ацетоацетата мозгом (K AcAc ) и скорости церебрального метаболизма AcAc и кетонов (CMR AcAc и CMR кетонов ).CMR рассчитывали с использованием следующего уравнения: CMR = K × Cp / LC, где K (min -1 ) — константа скорости поглощения, Cp — концентрация в плазме крови, а LC — сосредоточенная константа (1,0 для AcAc) с учетом для различий в поглощении между индикатором и природной молекулой. Расчет CMR кетонов использует плазменные концентрации AcAc и D-BHB, K AcAc и r k (K AcAc / K кетоны ), установленные на уровне 1,2, как сообщалось ранее (21, 22).CMR кетоны = (K AcAc x [AcAc]) × (1+ (1 / r k x [AcAc] / [D-BHB])).
Чтобы охарактеризовать влияние добавки D-BHB на поглощение кетонов органами, помимо мозга, 11 поглощение C-AcAc печенью, почками и сердцем были сегментированы на изображении слияния ПЭТ / КТ всего тела и% дозы / г рассчитывали из объема органа и введенной дозы.
Переориентация сердца и анализ сердечной функции были выполнены с использованием сердечного модуля PMOD 3.9, чтобы получить объемы желудочков, фракцию выброса и полярную карту (29).
Плазма, собранная во время ПЭТ-сканирования, была проанализирована на D-BHB и AcAc с помощью автоматического колориметрического анализа на анализаторе клинической химии (Dimension Xpand Plus; Siemens, Deerfield, IL, USA), как описано ранее (8).
Это исследование ПЭТ было одобрено Комитетом по этике исследований CIUSSS de l’Estrie – CHUS.
Результаты
Исследование фармакокинетики
После приема D-BHB количество кетонов в крови быстро увеличивалось (рис. 2A, дополнительная таблица 2) до C max , равного 1.2 ± 0,1 мМ. При потреблении того же количества D + L-BHB или MCT наблюдалось снижение C max на ~ 50% от исходного уровня по сравнению с D-BHB (C max D + L-BHB 0,62 ± 0,05 мМ и MCT 0,62 мМ). ± 0,06 мМ; p <0,001 по сравнению с D-BHB для обоих). T max был достигнут примерно через 1 час для каждого продукта (дополнительная таблица 3), а уровни кетонов вернулись к исходным значениям через 3-4 часа. IAUC для D-BHB была примерно в 1,5 раза выше, чем для D + L-BHB или MCT (рис. 3A, дополнительная таблица 1; p <0.005 для обоих).
Рисунок 2 . Кинетика кетонов крови после согласованных по грамму пероральных доз D-BHB, D + L-BHB и MCT у 15 голодных участников в состоянии покоя. (A) Плазменный кетон, (B) D-BHB, (C) AcAc и (D) L-BHB уровни в течение 4 часов. Значения — это средние значения + SEM.
Рисунок 3. (A) iAUC кетонов в плазме (мкМ * ч) для D-BHB, D + L BHB и MCT, и (B) — их соответствующее производство кетонов на потребленные калории (iAUC / ккал).
Анализ крови D-BHB и AcAc выявил аналогичную картину для трех продуктов (рисунки 2B, C). Однако MCT продуцировал пропорционально меньше AcAc в крови и больше D-BHB в крови, чем пероральные D-BHB или D + L-BHB. Рассчитанное соотношение [среднее iAUC AcAc / среднее iAUC D-BHB] составило 0,44 для MCT, что на ~ 30-40% ниже, чем для D-BHB (0,63) и D + L-BHB (0,76) (дополнительная таблица 1; D -BHB против MCT p = 0,0102; D + L-BHB против MCT p <0,0001; D + L-BHB против D-BHB p = 0.173). Исходное соотношение AcAc / D-BHB составляло ~ 0,7, что позволяет предположить, что MCT снижает соотношение AcAc / D-BHB в крови при производстве кетонов. Аналогичные соотношения были получены при использовании значений, полученных из C max [среднее значение AcAc C max / среднее значение D-BHB C max ] (дополнительная таблица 2).
Как и ожидалось, пероральный прием D + L-BHB привел к значительному увеличению L-BHB в плазме в течение первого часа (рис. 2D) с последующим медленным снижением в течение следующих 3 часов без возврата к исходному уровню.Для сравнения, пероральный прием D-BHB не вызывает увеличения L-BHB в плазме.
Производство кетонов на потребленную калорию [MCT: 8,3 ккал / г; BHB: 4,6 ккал / г (42)] был значительно выше для D-BHB, чем для D + L-BHB и MCT (рис. 3B, p <0,0001 для всех трех сравнений D-BHB против D + L BHB vs. MCT). Повышенные уровни глюкозы и инсулина в результате приема пищи через 30 минут после употребления каждого тестируемого продукта существенно не различались в трех группах (дополнительная таблица 4 и дополнительная фигура 2).Переносимость трех продуктов, оцененная по шкале визуальной оценки GI, была сходной без клинически значимых различий (дополнительный рисунок 1). Среди некоторых самооценок GI-эффекта снижение аппетита и небольшая диарея были наиболее распространенными для этих трех продуктов.
11 Пилотное исследование C-AcAc PETУровни кетонов в плазме составляли от 0,9 до 1,2 мМ в течение первых 30 минут после получения результатов сканирования ПЭТ. Первое биораспределение ПЭТ / КТ всего тела 11 C-AcAc через 18 минут после инъекции радиоактивного индикатора показано на рисунке 4.Радиоактивный индикатор быстро поступил в основном в сердце, где можно было наблюдать как желудочки, так и предсердия. 11 Оценки поглощения C-AcAc на грамм ткани в течение трех временных точек суммированы на рисунке 5. Выделение радиоактивного индикатора наблюдали в мочевом пузыре и слюнных железах.
Рисунок 4 . Изображение всего тела после приема двух доз D-BHB по 15 г, одной через −75 мин, а другой через −30 мин до инфузии 11 C-AcAc (330 МБк).8-минутное сканирование (30 секунд на слой) было получено через 18 минут после инъекции радиоактивного индикатора.
Рисунок 5 . 11 Распределение C-AcAc в органах после перорального приема D-BHB.
С помощью динамического сканирования мозга можно определить CMR AcAc и K AcAc для всех основных областей мозга (дополнительная таблица 5) и сравнить с исходными значениями, ранее определенными у здоровых молодых людей (16). В целом и по сравнению с исходным уровнем каждая область продемонстрировала увеличение CMR AcAc и K AcAc на ~ 4.В 7 и 2,3 раза, соответственно, примерно через 1 ч после приема D-BHB. Это указывает на то, что AcAc эффективно усваивается мозгом и другими органами, особенно сердцем и почками.
Несмотря на то, что динамическое сканирование сердца не могло быть выполнено в настоящем исследовании, оценка сердечной функции все еще была возможна с помощью ПЭТ-изображения закрытого сердца (рис. 6). Значения конечного диастолического и конечного систолического объемов, а также фракции выброса левого желудочка, рассчитанные на основе метаболизма 11 C-AcAc, составили 100, 47 мл и 52% соответственно.
Рисунок 6 . Изображение сердца: 15-минутное сканирование со стробированием сердца (8 кадров), полученное через 50 минут после инъекции 11 C-AcAc. Изображения левого желудочка (LV), правого желудочка (RV), горизонтальной длинной оси (HLA), вертикальной длинной оси (VLA) и короткой оси (SA).
Обсуждение
Насколько нам известно, это первое сообщение о метаболизме D-BHB у людей и его связи с распределением кетона AcAc в органах с помощью ПЭТ-изображений. Это исследование показывает, что D-BHB быстро всасывается и метаболизируется (рис. 2).Он также увеличивает содержание кетонов в крови выше 1 мМ, что примерно в 1,7 раза выше, чем та же доза D + L-BHB или MCT, и это несмотря на более низкую калорийную нагрузку D-BHB. Более того, результаты пилотного эксперимента по всему телу 11 C-AcAc-PET подтверждают осуществимость этого метода для измерения органо-специфической утилизации экзогенных кетонов, и они предполагают, что экзогенные кетоны очень активно используются сердцем и почками.
Производство кетонов из экзогенного пищевого источника традиционно достигается с помощью МСТ.Это требует болюсного приема, чтобы насытить печень MCFA, производя избыток ацетил-КоА, который затем преобразуется в AcAc и BHB, которые попадают в большой круг кровообращения. C max , достигаемый с помощью MCT, обычно составляет от 300 до 600 мкМ, при этом более высокие значения трудно достичь из-за побочных эффектов со стороны желудочно-кишечного тракта и насыщения печени. Здесь мы показываем, что D-BHB, природный и биологически активный изомер кетонов, повышает содержание кетона в крови C max выше 1 мМ без заметных побочных эффектов.Для сравнения, эквивалентная доза D + L-BHB или MCT достигла только половины этого уровня кетонов, с аналогичным T max через 1 час. Таким образом, по сравнению с D + L-BHB, D-BHB значительно снижает потребление соли, необходимое для достижения такой же реакции кетонов в плазме.
Результаты предыдущего исследования (9), сравнивающего KE с D + L-BHB, показали, что при той же дозе эквивалента D-BHB увеличение iAUC кетонов крови имело одинаковую величину, что позволяет предположить, что экзогенные D-BHB и KE вызывают аналогичный кетоз. . Мы отмечаем, что T max D + L-BHB был значительно дольше (~ 90 мин) по сравнению с KE (~ 20 мин) при той же дозе, что позволяет предположить, что D-BHB мог иметь более длительную продолжительность действия, чем KE.Мы также обнаружили, что D + L-BHB приводит к значительному увеличению концентрации L-BHB в плазме, которые устраняются медленно, что согласуется с предыдущим отчетом (9) и указывает на разницу в метаболической утилизации D и L-BHB. В то время как D-BHB легко используется в качестве источника энергии, L-изоформа медленно выводится с мочой и не вносит значительного вклада в биологическую активность предшественников кетонов в качестве источника энергии. Наконец, D-BHB обеспечивает более высокое содержание кетонов на потребленную калорию, чем MCT: калорийность D-BHB (4.6 ккал / г) вдвое меньше, чем MCT (8,2 ккал / г), что делает D-BHB в 2,6 раза более энергетически эффективным предшественником кетона по сравнению с MCT (на основе iAUC за 4 часа).
Настоящее исследование также показало, что преобразование D-BHB в AcAc обеспечивает примерно на 43% более высокое соотношение AcAc / D-BHB в крови (0,63), чем та же доза MCT (0,44). В этом отношении KE, похоже, ведет себя больше как MCT, с соотношением около 0,2–0,4 (3), тогда как на кетогенной диете это соотношение остается неизменным около 0,5 (16). Поскольку D-BHB необходимо преобразовать в AcAc с помощью BDH-1 в каждом органе, прежде чем он метаболизируется до ацетил-CoA (3), более высокое соотношение AcAc / D-BHB может быть энергетически более эффективным.Более того, поскольку это преобразование требует, чтобы кофактор, никотинамидадениндинуклеотид (NAD + ) был восстановлен до NADH, более высокое значение AcAc / D-BHB могло бы лучше сохранить митохондриальное соотношение NAD + / NADH. Интересно, что более высокое соотношение NAD + / NADH было связано с улучшением здоровья (30, 31) и может быть увеличено в мозге из-за пищевого кетоза (32, 33), что позволяет предположить, что D-BHB может превосходить MCT или KE в этом отношении заслуживает дальнейшего изучения.
ПЭТ неоценим для измерения энергетического метаболизма на уровне органов человека. Например, с 18 F-FDG он ясно показывает, что глюкоза является основным энергетическим субстратом мозга и что жирная кислота, такая как 11 C-пальмитат, предпочтительно используется сердцем и печенью (34, 35 ). Наш пилотный эксперимент 11 C-AcAc-PET был нацелен просто на демонстрацию возможности измерения поглощения кетона основными органами при пероральном введении экзогенного D-BHB людям.Ключевым преимуществом 11 C-AcAc в качестве метаболического радиоактивного индикатора является то, что он быстро уравновешивается с D-BHB в крови и неотличим от эндогенного AcAc, поэтому отражает фактический метаболизм кетонов. Сканирование всего тела впервые показало, что сердце и почки вносят основной вклад в метаболизм экзогенного AcAc после приема добавки D-BHB.
Большая часть потребности сердца в энергии обычно обеспечивается жирными кислотами и глюкозой (20). Наши результаты подтверждают, что сердце способно использовать другие субстраты, такие как кетоны (21), и что метаболизм кетонов, по-видимому, связан с метаболизмом и клиренсом радиоактивного индикатора миокардом.В пересчете на грамм почки потребляют большое количество энергии, в основном получаемой из жирных кислот, но наши данные показывают, что кетоны также могут эффективно использоваться для митохондриального дыхания в почках.
Скорость церебрального метаболизма для кетонов (CMR кетоны ), полученная здесь с D-BHB (3,0 мкмоль / 100 г / мин), превышала скорость, полученную после добавления MCT при легких когнитивных нарушениях [2,49 мкмоль / 100 г / мин; (17)] и болезнь Альцгеймера [1,9–2,0 мкмоль / 100 г / мин; (18)].Хотя значения CMR, полученные здесь для D-BHB, являются предварительными, наши данные предполагают, что добавка D-BHB представляет собой многообещающее вмешательство для обеспечения кетонов, которые могут спасти дефицит глюкозы в мозгу в этих условиях (17, 25).
11 Поглощение C-AcAc печенью (рисунки 4, 5) может отражать как преобразование 11 C-AcAc в 11 CD-BHB под действием BHD-1, так и высокую емкость кровеносных сосудов печени. но не отражают метаболизм кетонов самой печенью.Мы показываем здесь, что значения радиоактивности в крови 11 C-AcAc были аналогичны таковым для печени (см. Рисунок 5), что подтверждает мнение о том, что печень производит, но не метаболизирует кетоны (5). Это согласуется с тем, что радиоактивность печени после инфузии 11 C-AcAc связана с кровью, а не с преобразованием AcAc в AcAc-CoA и другие промежуточные энергетические продукты (5).
Недавние исследования показали потенциальную важность кетонов для здоровья сердечно-сосудистой системы. Инфузия D + L-BHB имеет благоприятные гемодинамические эффекты у взрослых пациентов с сердечной недостаточностью и более низкой фракцией выброса (36).У детей с дефектами окисления жирных кислот, такими как множественный дефицит ацил-КоА-дегидрогеназы, D + L-BHB улучшил функцию сердца и когнитивные функции, что привело к улучшению способности ходить и коррекции неврологических симптомов (37). Более того, клинические испытания при диабете ингибиторов ко-транспортера натрия / глюкозы-2 (SGLT-2), которые повышают уровень кетонов в плазме (38), показывают заметное снижение риска сердечно-сосудистых событий и почечной недостаточности (39). Учитывая значительное нарушение регуляции энергетического метаболизма при диабете 2 типа, вполне вероятно, что часть эффекта ингибиторов SGLT2 на сердце и почки опосредуется улучшенным производством АТФ с использованием кетонов в качестве альтернативного энергетического топлива жирным кислотам и глюкозе (38, 40) .Это открывает дополнительные возможности для терапевтического исследования добавок D-BHB помимо энергосбережения мозга.
Это исследование имеет несколько ограничений. Во-первых, фармакокинетическое сравнение D-BHB, D + L-BHB и MCT было однократным 4-часовым исследованием в острой форме; Степень, в которой он отражает долгосрочные различия в их метаболизме, еще предстоит определить. Во-вторых, ПЭТ-сканирование всего тела проводилось на одном человеке и (за исключением мозга) представляло собой полуколичественное сравнение по органам. Для количественного сканирования был выбран головной мозг (CMR AcAc ).Для количественной оценки поглощения индикатора требуется динамическое ПЭТ-сканирование, и его нельзя проводить одновременно в головном мозге и других органах. Качество стробируемого изображения сердца можно было бы улучшить, если бы изображение было получено раньше после введения трассера. В-третьих, производство и метаболизм кетонов варьируются в зависимости от постпрандиального метаболического статуса, который, возможно, влияет на распределение кетонов в органах. Следовательно, для проверки относительных различий между органами потребуется последующее наблюдение за этим единичным наблюдением при различных условиях кормления.
Выводы
D-BHB, по-видимому, является многообещающей добавкой для производства значительно более высоких кетонов в крови, чем D + L-BHB или MCT, и при более низком потреблении калорий для эквивалентной дозы. Более того, экзогенный D-BHB, по-видимому, не снижает соотношение AcAc / D-BHB в крови, что в целом может сделать его более эффективным «супергопливом» по сравнению с другими предшественниками кетонов, такими как MCT, D + L-BHB или KE. .
Пилотное исследование 11 C-AcAc-PET четко определяет сердце и почки как важные потребители экзогенных кетонов, фактически больше, чем мозг.Следовательно, добавка D-BHB может быть протестирована при таких состояниях, как сердечная недостаточность и диабетическая кардиомиопатия, для повышения эффективности и функции сердечной энергии, а также при хроническом заболевании почек.
Заявление о доступности данных
Все наборы данных, созданные для этого исследования, включены в статью / дополнительные материалы.
Заявление об этике
Исследования с участием людей были рассмотрены и одобрены Комитетом по этике кантона Во (Швейцария) под номером общего протокола 2018-00503, регистрационный номер исследования NCT03603782 (фармакокинетическое исследование), а также CIUSSS de l’Estrie, CHUS Research. Комитет по этике, Шербрук (Канада; исследование ПЭТ).Пациенты / участники предоставили письменное информированное согласие на участие в этом исследовании. Письменное информированное согласие было получено от человека (лиц) на публикацию любых потенциально идентифицируемых изображений или данных, включенных в эту статью.
Авторские взносы
MH, EC, C-AC, SC и BC разработали исследования. Сбор данных выполняли MH, EC, C-AC, J-PG и MH. C-AC и EC проанализировали данные. Рукопись была составлена BC и SC, и все авторы обсудили результаты и критически отредактировали рукопись.
Финансирование
Финансирование предоставлено Nestlé.
Конфликт интересов
BC, MH и J-PG — сотрудники Nestlé. Компания SC консультировала компании Nestlé, Bulletproof и Accera и получала финансирование исследований и / или исследовательские материалы от Ассоциации Альцгеймера (США), Mitacs, FRQS, Abitec и Nestlé. AC объявляет о финансировании исследований от CIHR, Канадской диабетической ассоциации, Fonds de recherche du Quebec-Santé, Janssen, Merck, UniQure, Caprion, Eli Lilly, Novo Nordisk, GlaxoSmithKline, Novartis, Pfizer, Philips, Sanofi, Siemens и Amgen и консалтинг / участие в консультативных группах или оплата конференций от компаний Merck, Amgen, Janssen, UniQure, Servier, Novo Nordisk и Novartis.EC, MM и C-AC заявляют об отсутствии конкурирующих финансовых интересов.
Благодарности
Мы благодарим сотрудников отдела клинических разработок и метаболического отделения «Нестле» за клиническое руководство фармакокинетическим исследованием и за их помощь в наборе добровольцев; и благодарим Изабель Бретон, Ирину Моннар, Шехеразад Корбаз и Симону Бартову из Nestlé Research за их умелую аналитическую помощь в измерении D и L-BHB.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fnut.2020.00013/full#supplementary-material
Список литературы
1. D’Andrea Meira I, Romão TT, Pires do Prado HJ, Krüger LT, Pires MEP, da Conceição PO. Кетогенная диета и эпилепсия: что мы знаем до сих пор. Front Neurosci. (2019) 13: 5. DOI: 10.3389 / fnins.2019.00005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
2. Ким С.Х., Шоу А., Блэкфорд Р., Лоуман В., Ло Лукс, Милличэп Дж. Дж. И др. Кетогенная диета у детей в возрасте 3 лет и младше: 10-летний опыт работы в одном центре. Научный доклад (2019) 9: 8736. DOI: 10.1038 / s41598-019-45147-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
3. McDonald TJW, Cervenka MC. Уроки, извлеченные из недавних клинических испытаний методов лечения кетогенной диетой у взрослых. Curr Opin Clin Nutr Metab Care. (2019) 22: 418–24. DOI: 10.1097 / MCO.0000000000000596
PubMed Аннотация | CrossRef Полный текст | Google Scholar
4. Кампизи Дж., Капахи П., Литгоу Дж. Дж., Мелов С., Ньюман Дж. К., Вердин Э.От открытий в исследованиях старения до терапевтических средств для здорового старения. Природа. (2019) 571: 183–92. DOI: 10.1038 / s41586-019-1365-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
5. Puchalska P, Crawford PA. Многомерные роли кетоновых тел в топливном метаболизме, передаче сигналов и терапии. Cell Metab. (2017) 25: 262–84. DOI: 10.1016 / j.cmet.2016.12.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
6.Антон С.Д., Моэль К., Донаху В.Т., Марози К., Ли С.А., Майноус А.Г. III и др. Переключение метаболизма: понимание и применение преимуществ голодания для здоровья. Ожирение. (2018) 26: 254–68. DOI: 10.1002 / oby.22065
PubMed Аннотация | CrossRef Полный текст | Google Scholar
7. Стекович С., Хофер С.Дж., Трипольт Н., Аон М.А., Ройер П., Пейн Л. и др. Альтернативное дневное голодание улучшает физиологические и молекулярные маркеры старения у здоровых людей, не страдающих ожирением. Cell Metab. (2019) 30: 462–76.e5. DOI: 10.1016 / j.cmet.2019.07.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
8. Куршн-Лойе А., Фортье М., Трембле-Мерсье Дж., Шуинар-Уоткинс Р., Рой М., Ньюджент С. и др. Стимуляция умеренной устойчивой кетонемии среднецепочечными триацилглицеринами у здоровых людей: предполагаемый потенциальный вклад в энергетический метаболизм мозга. Питание. (2013) 29: 635–40. DOI: 10.1016 / j.nut.2012.09.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
10.Стаббс Б.Дж., Кокс П.Дж., Кирк Т., Эванс Р.Д., Кларк К. Желудочно-кишечные эффекты экзогенных кетоновых напитков нечасты, мягкие и варьируются в зависимости от кетонового соединения и дозы. Int J Sport Nutr Exerc Exerc Metab. (2019) 29: 1–23. DOI: 10.1123 / ijsnem.2019-0014
CrossRef Полный текст | Google Scholar
11. О’Коннор А., Чанг Дж. Л., Браунлоу М., подрядчик Н. Острое пероральное употребление бета-гидроксибутирата в пилотном исследовании временно увеличивало его капиллярные уровни у здоровых добровольцев. J Nutr Health Food Eng. (2018) 8: 324–8. DOI: 10.15406 / jnhfe.2018.08.00289
CrossRef Полный текст | Google Scholar
12. Фишер Т., Оч Ю., Клаван И., Оч Т., Грюнеберг М., Фобкер М. и др. Влияние соли DL-β-гидроксибутирата натрия и кальция на здоровых взрослых. J Nutr Metab. (2018) 2018: 9812806. DOI: 10.1155 / 2018/9812806
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. O’Malley T., Myette-Cote E, Durrer C., Little JP.Пищевые соли кетонов увеличивают окисление жиров, но ухудшают выполнение высокоинтенсивных упражнений у здоровых взрослых мужчин. Appl Physiol Nutr Metab. (2017) 42: 1031–35. DOI: 10.1139 / apnm-2016-0641
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Холланд А.М., Кази А.С., Бизли К.Н., Беннетт Х.Р. Параметры крови и сердечно-сосудистой системы после приема кетоновых солей в течение шести недель. J. Insul. Оказывать сопротивление. (2019) 4: a47. DOI: 10.4102 / jir.v4i1.47
CrossRef Полный текст | Google Scholar
16. Курчесн-Лойе А, Крото Е, Кастеллано Калифорния, Сен-Пьер V, Хеннебель М, Куннан, Южная Каролина. Обратная связь между глюкозой в головном мозге и метаболизмом кетонов у взрослых во время краткосрочного умеренного диетического кетоза: исследование с использованием количественной позитронно-эмиссионной томографии с двумя индикаторами. J Cereb Blood Flow Metab. (2017) 37: 2485–93. DOI: 10.1177 / 0271678X16669366
PubMed Аннотация | CrossRef Полный текст | Google Scholar
17.Fortier M, Castellano CA, Croteau E, Langlois F, Bocti C, St-Pierre V и др. Кетогенный напиток улучшает энергию мозга и некоторые когнитивные функции при легких когнитивных нарушениях. Демент Альцгеймера. (2019) 15: 625–34. DOI: 10.1016 / j.jalz.2018.12.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
18. Croteau E, Castellano CA, Richard MA, Fortier M, Nugent S, Lepage M, et al. Кетогенные триглицериды со средней длиной цепи увеличивают энергетический метаболизм мозга при болезни Альцгеймера. J Alzheimers Dis. (2018) 64: 551–61. DOI: 10.3233 / JAD-180202
PubMed Аннотация | CrossRef Полный текст | Google Scholar
19. Neth BJ, Mintz A, Whitlow C., Jung Y, Solingapuram Sai K, Register TC, et al. Модифицированная кетогенная диета связана с улучшенным профилем биомаркеров спинномозговой жидкости, церебральной перфузией и потреблением церебральных кетоновых тел у пожилых людей из группы риска по болезни Альцгеймера: пилотное исследование. Neurobiol Aging. (2020) 86: 54–63. DOI: 10.1016 / j.neurobiolaging.2019.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
21. Gormsen LC, Svart M, Thomsen HH, Søndergaard E, Vendelbo MH, Christensen N, et al. Инфузия кетонового тела с 3-гидроксибутиратом снижает поглощение глюкозы миокардом и увеличивает кровоток у людей: исследование с помощью позитронно-эмиссионной томографии. J Am Heart Assoc. (2017) 6: e005066. DOI: 10.1161 / JAHA.116.005066
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24.Наджент С., Тремблей С., Чен К.В., Аютянонт Н., Рунтива А., Кастеллано, Калифорния и др. Метаболизм глюкозы и ацетоацетата в мозге: сравнение молодых и пожилых людей. Neurobiol Aging. (2014) 35: 1386–95. DOI: 10.1016 / j.neurobiolaging.2013.11.027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
25. Croteau E, Castellano CA, Fortier M, Bocti C, Fulop T., Paquet N, et al. Поперечное сравнение метаболизма глюкозы и кетонов в мозге у когнитивно здоровых пожилых людей, умеренных когнитивных нарушений и ранней болезни Альцгеймера. Exp Gerontol. (2018) 107: 18–26. DOI: 10.1016 / j.exger.2017.07.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
26. Croteau E, Tremblay S, Gascon S, Dumulon-Perreault V, Labbé SM, Rousseau JA, et al. [(11) C] -Ацетоацетатная ПЭТ-визуализация: потенциальный ранний маркер сердечной недостаточности. Nucl Med Biol. (2014) 41: 863–70. DOI: 10.1016 / j.nucmedbio.2014.08.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
27.Цзэн М., Цао Х. Быстрое количественное определение короткоцепочечных жирных кислот и кетоновых тел методом жидкостной хроматографии и тандемной масс-спектрометрии после легкой дериватизации в сочетании с жидкостно-жидкостной экстракцией. J Chromatogr B Analyt Technol Biomed Life Sci. (2018) 1083: 137–45. DOI: 10.1016 / j.jchromb.2018.02.040
PubMed Аннотация | CrossRef Полный текст | Google Scholar
28. Уилкоксон Ф., Кэтти С.К., Уилкокс Р.А. Критические значения и уровни вероятности для теста суммы рангов Вилкоксона и знакового рангового критерия Вилкоксона.В: Избранные таблицы математической статистики , Vol. 1. (1970). п. 171–259.
Google Scholar
29. Гессе Б., Линдхардт Т. Б., Акампа В., Анагностопулос С., Баллинджер Дж., Бакс Дж. Дж. И др. Рекомендации EANM / ESC по радионуклидной визуализации сердечной функции. евро J Nucl Med Mol Imaging. (2008) 35: 851–85. DOI: 10.1007 / s00259-007-0694-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
31. Маташич Д.С., Бреннер С., Лондон Б. Новые потенциальные преимущества модуляции метаболизма НАД + при сердечно-сосудистых заболеваниях. Am J Physiol Heart Circ Physiol. (2018) 314: H839–52. DOI: 10.1152 / ajpheart.00409.2017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32. Синь Л., Ипек О., Бомонт М., Шевлякова М., Кристинат Н., Масуди М. и др. Пищевой кетоз увеличивает соотношение NAD + / NADH в мозге здорового человека: исследование in vivo, , проведенное 31P-MRS. Передняя гайка. (2018) 5:62. DOI: 10.3389 / fnut.2018.00062
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33.Эламин М., Раскин Д. Н., Масино С. А., Саккетти П. Метаболическая терапия на основе кетонов: является ли повышение НАД + основным механизмом? Front Mol Neurosci. (2017) 10: 377. DOI: 10.3389 / fnmol.2017.00377
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34. Büsing KA, Schönberg SO, Brade J, Wasser K. Влияние уровня глюкозы в крови, диабета, инсулина и ожирения на стандартизованные значения поглощения в опухолях и здоровых органах при ПЭТ / КТ 18F-FDG. Nucl Med Biol. (2013) 40: 206–13.DOI: 10.1016 / j.nucmedbio.2012.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Кристенсен Н.Л., Якобсен С., Шахт А.С., Мунк О.Л., Альструп АКО, Толбод Л.П. и др. Биораспределение в организме, дозиметрия и коррекция метаболитов [11C] пальмитата: ПЭТ-индикатор для визуализации метаболизма жирных кислот. Mol Imaging. (2017) 16: 1536012117734485. DOI: 10.1177 / 1536012117734485
PubMed Аннотация | CrossRef Полный текст | Google Scholar
36.Нильсен Р., Мёллер Н., Гормсен Л.С., Толбод Л.П., Ханссон Н.Х., Соренсен Дж. И др. Сердечно-сосудистые эффекты лечения 3-гидроксибутиратом кетоновых тел у пациентов с хронической сердечной недостаточностью. Тираж. (2019) 139: 2129–41. DOI: 10.1161 / CIRCULATIONAHA.118.036459
PubMed Аннотация | CrossRef Полный текст | Google Scholar
37. Фишер Т., Оч У., Марквардт Т. Долгосрочная терапия кетоновым телом тяжелой множественной недостаточности ацил-КоА-дегидрогеназы: отчет о болезни. Питание. (2019) 60: 122–8. DOI: 10.1016 / j.nut.2018.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
38. Мудальяр С., Аллоджу С., Генри Р.Р. Может ли изменение в топливной энергетике объяснить положительные кардиоренальные исходы в исследовании EMPA-REG OUTCOME? Объединяющая гипотеза. Уход за диабетом. (2016) 39: 1115–22. DOI: 10.2337 / dc16-0542
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Гарофало С., Боррелли С., Либерти М.Э., Андреуччи М., Конте Дж., Минутоло Р. и др.Ингибиторы SGLT2: нефропротекторная эффективность и побочные эффекты. Medicina. (2019) 55: 268. DOI: 10.3390 / medicina55060268
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40. Garcia-Ropero A, Santos-Gallego CG, Zafar MU, Badimon JJ. Метаболизм при сердечной недостаточности и влияние ингибиторов SGLT2. Expert Opin Drug Metab Toxicol. (2019) 15: 275–85. DOI: 10.1080 / 17425255.2019.1588886
PubMed Аннотация | CrossRef Полный текст | Google Scholar
β-гидроксибутират и его метаболические эффекты на возрастную патологию
Старение
Влияние ограничения калорий на продление жизни и отсрочку возникновения возрастных заболеваний у различных видов животных, включая крыс, мышей, рыб, мух, черви и дрожжи были продемонстрированы 8 .Тем не менее, молекулы и клеточные сигналы, лежащие в основе этих эффектов, остаются неуловимыми. Интенсивные усилия показали, что кетогенная диета способствует увеличению продолжительности жизни, как и ограничение калорий 13 . Циркулирующий β-HB является наиболее значимым метаболитом при ограничении калорийности и кетогенных диетах, выделяя β-HB как метаболит против старения. 11 .
Действительно, добавка β-HB продлевает продолжительность жизни C. elegans на 20% за счет путей DAF-16 / FOXO и SKN-1 / Nrf и регуляции старения и долголетия 14 .У млекопитающих β-HB снижает секреторный фенотип, связанный со старением (SASP), и старение сосудистых клеток 15 . Более того, кетогенная диета значительно увеличила среднюю продолжительность жизни мышей и привела к сохранению физических функций у старых мышей 13 . Кроме того, сообщалось, что циклическая кетогенная диета снижает смертность в среднем возрасте и улучшает память у старых мышей 16 .
Возрастное репрограммирование и эпигенетическое омоложение способствуют увеличению продолжительности жизни и улучшению омоложения возраста и возрастных признаков 17 .Появились исследования в области регенеративной медицины, открывающие путь к новым терапевтическим вмешательствам при таких возрастных заболеваниях. Хотя многие исследования кетогенной диеты и ее терапевтических эффектов в регенеративной медицине во время старения и особенно нейродегенеративных заболеваний продолжают публиковаться, молекулярный механизм кетоновых тел еще не изучен полностью. Кетогенная диета показала нейрозащитный эффект на центральную нервную систему (ЦНС) за счет регенерации седалищных нервов 18 .Кетогенная диета также восстанавливала целостность олигодендроцитов и увеличивала миелинизацию ЦНС у мышей модели болезни Пелизея-Мерцбахера 19 . Более того, экзогенный β-HB улучшает гомеостаз стволовых клеток и функцию кишечных стволовых клеток посредством активации передачи сигналов Notch, которая является ключевой осью передачи сигналов для регенерации тканей 20 . Таким образом, β-HB и кетогенные диеты можно рассматривать как важные медиаторы с регенеративным потенциалом, которые также способны замедлять фенотипы, связанные со старением.
Раковые заболевания
Старение является наиболее значительным фактором риска развития рака, который является основной причиной смертности людей. 21 . Раковые клетки претерпевают значительные изменения в метаболизме, что приводит к увеличению уровней митохондриальных активных форм кислорода (АФК), таких как O 2 — и H 2 O 2 . Раковые клетки предпочитают переключаться на аэробный гликолиз, известный как эффект Варбурга, чтобы компенсировать митохондриальную дисфункцию, вызванную повышенными уровнями ROS 22 .Таким образом, снижение доступности глюкозы для раковых клеток предлагает терапевтический вариант. Недавнее исследование предполагает, что кетогенная диета усиливает терапевтический ответ раковых клеток за счет избирательного метаболического окислительного стресса 23 . Другие исследования на животных подтверждают, что кетогенная диета подавляет развитие первичной опухоли 24 , а также системных метастазов 25,26 . Постоянное употребление богатой углеводами западной диеты приводит к высоким уровням инсулина и инсулиноподобного фактора роста (IGF-1), способствуя пролиферации опухолевых клеток 27 .Кроме того, сообщалось, что раковые клетки увеличивают свою зависимость от глюкозы в крови в ответ на потребность в быстром росте клеток, а другие исследования показали, что глюкоза может иметь прямое или косвенное влияние на пролиферацию опухолевых клеток. Неоднократно сообщалось, что кетогенные диеты с ограничением углеводов, обогащенные жирами, подавляют рак груди 28 . Кроме того, сообщалось, что кетоновые тела, используемые в качестве источника топлива, подавляют пролиферацию раковых клеток. В частности, исследователи указали на более высокую заболеваемость раком груди среди людей с диабетом и ожирением, подтвердив, что низкоуглеводная диета может ограничивать рост опухоли. 29 .Таким образом, учитывая влияние углеводов на развитие рака груди, кетогенная диета может контролировать или снижать риск развития рака груди. В других исследованиях также сообщалось, что кетогенная диета может быть особенно полезной при лечении рака мозга 30,31 , поскольку пациенты с наиболее распространенной и агрессивной формой рака мозга, мультиформной глиобластомой, показали значительное улучшение после принятия кетогенной диеты. 32 . Хотя кетогенная диета не может существенно повлиять на прогрессирование заболевания при запущенных и терминальных формах рака, она безопасна и может улучшить качество жизни больных раком в сочетании с лучевой или другой проверенной противоопухолевой терапией. 33,34 .Приведенные выше данные свидетельствуют о том, что эти методы лечения должны быть дополнительно исследованы для изучения кетонов в качестве потенциальных адъювантных терапевтических средств с минимальной токсичностью.
Неврологические расстройства
По мере увеличения продолжительности жизни населения все больше пожилых людей страдают неврологическими расстройствами, такими как эпилепсия и деменция 35 . Недавние исследования показали, что пациенты с эпилепсией имеют более высокий риск развития деменции, особенно болезни Альцгеймера. 36 . Чрезмерная мозговая активность у больных эпилепсией вызывает судороги.Противосудорожные препараты эффективны только для некоторых пациентов с эпилепсией, в то время как другие не реагируют на лекарства или испытывают побочные эффекты. Согласно многим сообщениям, кетогенная диета с высоким содержанием жиров и низким содержанием углеводов приводит к значительному улучшению у неизлечимых детей с эпилепсией. Кетогенная диета в качестве основного лечения уменьшила эпилептические припадки более чем наполовину и, таким образом, использовалась во всем мире для лечения неизлечимой детской эпилепсии 37,38 . Кроме того, в других отчетах документально подтверждены улучшения при эпилепсии, а также при других неврологических заболеваниях у пациентов, соблюдающих кетогенную диету. 39,40 .Интересно, что кетоны, как сообщалось, изменяют микробиоту кишечника, предотвращая судороги и спонтанные тонико-клонические судороги посредством модуляции соотношений ГАМК / глутамат в гиппокампе 41 .
Болезнь Альцгеймера является наиболее распространенным нейродегенеративным заболеванием, связанным с возрастом, и требует эффективной терапевтической стратегии из-за растущего социально-экономического бремени. Периферическое старение, вызванное воспалением, перекосом иммунных клеток, старением и инфекцией, дополнительно способствует возникновению и прогрессированию болезни Альцгеймера. 42 .Это прогрессирующее заболевание характеризуется клубками в головном мозге и накоплением β-амилоидных бляшек, которые являются известными маркерами болезни Альцгеймера и, как считается, ухудшают память. Исследования на животных показали, что β-HB потенциально может уменьшать количество амилоидных бляшек, и, таким образом, механизмы повышения уровня β-HB в крови с помощью кетогенной диеты, добавок KE или масла среднецепочечных триглицеридов (MCT) потенциально имеют отношение к Лечение болезни Альцгеймера. Кроме того, было показано, что повышение уровня кетонов в результате диетических манипуляций с KE или маслом MCT улучшает некоторые симптомы болезни Альцгеймера 43,44,45,46,47 .
Для оценки влияния кетогенной диеты на двигательную активность использовали две линии трансгенных мышей, мышей APP / PS1 (модель отложения амилоида) и мышей Tg4510 (модель отложения тау-белка). 44 . Мыши-модели, получавшие кетогенную диету в течение трех месяцев, показали значительно лучшие результаты при тестировании поведения вращающихся стержней, чем мыши в контрольной группе, независимо от генотипа. Данные демонстрируют, что кетогенные диеты могут играть важную роль в повышении двигательной активности у моделей мышей 44 .
Болезнь Паркинсона — нейродегенеративное заболевание, характеризующееся потерей нигростриатных дофаминергических нейронов, сопровождающейся митохондриальной респираторной недостаточностью. 48 . Кетогенные диеты изучаются как потенциальные дополнительные методы лечения болезни Паркинсона из-за их защитного воздействия на мозг и нервную систему, как описано для болезни Альцгеймера и эпилепсии. Было показано, что кетогенная диета защищает нейроны черной субстанции от нейротоксичности 6-гидроксидофамина на моделях крыс на животных 48 .1-Метил-4-фенил-1,2,3,6-тетрагидропиридин (МРТР) представляет собой нейротоксин, который вызывает дофаминергическую нейродегенерацию и митохондриальную недостаточность, напоминающую болезнь Паркинсона. В модели мышиной болезни Паркинсона, индуцированной МРТР, нейродегенерация дофамина, вызванная МРТР, была частично защищена инфузией кетоновых тел. Инъекция β-HB мышам обеспечивала частичную защиту от нейродегенерации дофамина и двигательной недостаточности, индуцированной MPTP 49 . Таким образом, инфузия кетоновых тел или принятие кетогенной диеты защищает от повреждения нервов и улучшает двигательную функцию в моделях животных Паркинсона 48,49 , что предполагает дальнейшее изучение потенциала таких методов лечения пациентов с болезнью Паркинсона.
Сердечно-сосудистые заболевания
Ожирение связано с сердечно-сосудистыми заболеваниями и приводит к метаболическим осложнениям, таким как инсулинорезистентность 50 . Сообщалось, что кетогенные диеты снижают вес более эффективно, чем чистое ограничение калорий или диета с низким содержанием жиров. 51,52,53 . Помимо похудания, кетогенные диеты снижают уровень триглицеридов, холестерина ЛПНП и глюкозы в крови, а также повышают уровень холестерина ЛПВП 52,54 .Еще одно преимущество заключается в том, что этот тип диеты помогает человеку чувствовать себя менее голодным, а ингибирующие эффекты кетоза также могут помочь людям потреблять меньше калорий. 55 . Низкоуглеводные кетогенные диеты часто практикуются для похудения или поддержания веса, но метаболические эффекты длительного употребления этого типа диеты остаются спорными, поскольку длительное употребление кетогенной диеты снижает чувствительность к инсулину и снижает толерантность к глюкозе 56 . Эти результаты могут быстро обратить вспять последствия ожирения.Таким образом, прерывистая кетогенная диета считается эффективной для снижения ожирения.
Метаболический синдром, включая непереносимость глюкозы и диабет 2 типа, связан со старением 57 . Было показано, что кетогенная диета улучшает гликемический контроль и улучшает диабетические осложнения у пациентов с диабетом 1 и 2 типа. Эти пациенты смогли прекратить или уменьшить прием лекарств от диабета, употребляя кетоны, снижая вес и снижая уровень триглицеридов и артериального давления. 58,59,60 .У крыс соблюдение кетогенной диеты в течение 8 недель снижает чувствительность к периферическому инсулину и снижает толерантность к глюкозе; однако возвращение к нормальной диете после кетогенной диеты привело к резкому изменению этих эффектов. Таким образом, длительное поддержание кетогенной диеты отрицательно влияет на гомеостаз глюкозы, но этот эффект можно быстро обратить вспять после прекращения кетогенной диеты 56 . Таким образом, можно предположить, что прямое потребление кетоновых тел, в частности β-HB, является более эффективным способом контроля метаболического синдрома.
Неалкогольная жировая болезнь печени (НАЖБП), которая также часто встречается у пожилых людей, тесно связана с диабетом 2 типа, метаболическим синдромом и ожирением. У людей с НАЖБП наблюдается избыток внутрипеченочных триглицеридов. Кетогенная диета значительно снижает уровень триглицеридов в печени у субъектов с НАЖБП по сравнению с ограничением калорий 61 , а другой отчет продемонстрировал улучшение НАЖБП у пациентов, которые приняли кетогенную диету 62,63 .
Молекулярные мишени β-HB
Преимущества кетогенной диеты теперь хорошо известны благодаря растущей научной поддержке.Будучи чем-то большим, чем просто метаболит, β-HB обладает способностью запускать и контролировать множество сигнальных событий, имеющих последствия для многих метаболических заболеваний. Однако он имеет широкий спектр мишеней на молекулярном уровне, и основными молекулярными мишенями являются инфламмасома NLRP3, РНК-связывающие белки и рецепторы, связанные с G-белками (рис. 3). Кроме того, β-HB также был идентифицирован как эпигенетический модификатор, который может воздействовать на ДНК и гистоны. Например, β-HB является эндогенным ингибитором многих протеиндеацетилаз (HDAC) и модулятором β-гидроксибутирилирования (рис.3), который представляет собой новый тип эпигенетического регуляторного механизма. Таким образом, четкое понимание связи между метаболизмом β-HB и эпигенетикой может дать возможность разработать новые фармакологические вмешательства для улучшения различных патологических состояний.
Фиг. 3: Ингибирование, активация и посттрансляционная модификация β-HB.β-HB прямо или косвенно взаимодействует со многими клеточными белками в различных органеллах. β-HB действует как агонист или антагонист двух GPCR, FFAR3 и HCAR2, в плазматической мембране.β-HB напрямую связывается с hnRNP A1, регулируя стабильность мРНК Oct4 . β-HB подавляет воспаление за счет ингибирования образования воспаления NRLP3 или его активности. В частности, β-HB также регулирует ядерные белки. β-HB является ингибитором HDAC, который также регулирует гистоны и p53 посредством β-гидроксибутирилирования.
Гистоновые деацетилазы
Увеличение β-HB путем голодания или ограничения калорий привело к глобальному ацетилированию гистонов у мышей посредством ингибирования гистоновых деацетилаз I класса (HDAC) (HDAC1, 2, 3 и 8), которые также называются сиртуинами и включают семейство белков, которые регулируют возрастную экспрессию генов 64,65 .Ингибирование HDACs с помощью β-HB усиливает экспрессию генов Foxo3a и MT2 , которые кодируют факторы устойчивости к окислительному стрессу 65 . Кроме того, β-HB предотвращает ретракцию микроглиального отростка и депрессивное поведение за счет ингибирования HDAC. β-HB-индуцированное ветвление и активация Akt в микроглии аннулировали активность HDAC, что привело к дальнейшему снижению нейровоспаления в микроглии. 66 . Бутират — это карбоновая кислота с короткой цепью, вырабатываемая в кишечнике.Недавние исследования показали, что подобный по структуре и функциональному назначению бутират ингибирует HDAC более эффективно, чем β-HB. На моделях грызунов добавление бутирата вызывает экспрессию метаболически важных генов, которые могут улучшить чувствительность к инсулину, связанную с повышенным расходом энергии 67 . Сюда входят коактиватор гамма-рецептора, активируемого пролифератором пероксисом, 1-альфа (PGC1-α), карнитин-пальмитоилтрансфераза 1B (CPT1b), митохондриальные сиртуины, супероксиддисмутаза 2 (SOD2) и каталаза 68 .
G-белковые рецепторы
β-HB, как сообщается, связывается непосредственно с рецептором 2 гидроксикарбоновой кислоты (HCA2). HCA2 представляет собой высокоаффинный рецептор, связанный с G-белком (GPCR), который представляет собой рецептор никотиновой кислоты с полумаксимальной эффективной концентрацией (EC 50 ) 0,7 мМ 69 . Поскольку GSK256073, селективный агонист HCA2, снижает уровни FFA посредством ингибирования липолиза и глюкозы при диабете 2 типа 70 , β-HB может также снижать уровни FFA и глюкозы посредством действия в качестве агониста HCA2 для ингибирования атерогенной активности.
Экспрессия HCA 2 не ограничена в адипоцитах, но она также экспрессируется на нейтрофилах и резидентных макрофагах, а также в головном мозге. В модели инсульта было идентифицировано, что HCA 2 требуется для нейрозащитного эффекта β-HB и кетогенной диеты, поскольку этот эффект теряется у мышей HCA2 — / — . Кроме того, известно, что активация HCA2, вызванная кетогенной диетой, доставляет нейрозащитные сигналы через невоспалительную инфильтрацию макрофагов в ишемический мозг.β-HB, благодаря своей активности агониста HCA2, индуцировал нейропротекторный фенотип, поскольку моноциты и макрофаги полагаются на продукцию простагландина D2 (PGD2) с помощью циклооксигеназы 1 (COX1) и гематопоэтической PGD2-синтазы 71 . Механически PGD2, высвобождаемый моноцитами и макрофагами, опосредует нейрозащитный эффект HCA2, устраняя воспаление и ингибируя киназу IκB (IKK) и NF-κB, которые играют ключевую роль в ишемическом повреждении мозга.
Другим рецептором β-HB является рецептор 3 FFA (FFAR3, GPR41), который контролирует расход энергии тела для поддержания метаболического гомеостаза.Активируемый короткоцепочечными жирными кислотами (SCFA) и β-HB, которые продуцируются печенью при голодании, этот рецептор ингибирует кальциевые каналы N-типа и модулирует активность симпатических нейронов посредством сигнального каскада, который включает субъединицы β и γ связанный с ним G-протеин-фосфолипаза C (PLC) и MAP-киназы, такие как ERK. Следовательно, этот фактор может регулировать расход энергии, воздействуя на симпатическую нервную систему, контролирующую частоту сердечных сокращений. β-HB подавлял активность симпатической нервной системы, противодействуя FFAR3 во время голодания или диабета 72 .
РНК-связывающие белки
β-HB напрямую связывает несколько РНК-связывающих белков, включая гетерогенный ядерный рибонуклеопротеин A1 (hnRNP A1), фактор сплайсинга пролин и богатый глутамином (SFPQ) и РНК-связывающий белок FUS / TLS. hnRNP A1 является доминирующим партнером связывания β-HB в сосудистых клетках, таких как эндотелиальные и гладкомышечные клетки 15 . Повышение уровня циркулирующего β-HB замедляет процесс старения мышей, предотвращая старение сосудистых клеток. Кроме того, hnRNP A1 противодействует клеточному старению и SASP посредством стабилизации мРНК Oct4 и Sirt1 15,73 .Кроме того, конститутивное снижение уровней и активности hnRNP A1 или A2 в стареющих фибробластах обычно сопровождается увеличением уровня изоформы p16 (INK4a), которая является первичным маркером старения 74 . Кроме того, SFPQ и FUS также сильно связаны с возрастными нейродегенеративными заболеваниями и боковым амиотрофическим склерозом 75,76 .
NLRP3
Хотя многочисленные исследования показали, что ограничение калорий или кетогенная диета снижает окислительный стресс и воспаление, влияние β-HB-опосредованного врожденного иммунного ответа остается неясным. 77 .β-HB подавляет активацию NOD-, LRR- и пиринового домена-содержащего белка 3 (NLRP3) инфламмасомы, предотвращая отток K + и снижая спек-подобный белок, связанный с апоптозом, с олигомеризацией домена рекрутирования каспазы (ASC) и пятнышками. пласт 78 . Интересно, что S-β-HB, хиральный энантиомер β-HB, проявляет сходную ингибирующую способность, но структурно родственные молекулы, такие как AcAc, бутират и ацетат, не ингибируют активность NLRP3 78 . Кроме того, ингибирование инфламмасомы NLRP3 не зависит от разобщающего белка-2 (UCP2), сиртуина-2 (SIRT2), GPCR FFAR3 или HCAR2, которые были описаны как молекулы-мишени для β-HB.Эти наблюдения показывают, что подавление воспаления за счет ограничения калорий или кетогенной диеты происходит за счет усиления нацеливания β-HB на инфламмасому NRLP3 78 .
β-гидроксибутирилирование
β-гидроксибутирилирование, новый тип модификации гистонов, описывается как эпигенетическая регуляторная метка, обогащенная активными промоторами генов 79 . Сорок четыре неизбыточных сайта β-гидроксибутирилирования остатков гистонового лизина были проверены в клетках человека и мыши, что дает представление о новой эпигенетической регуляторной метке, которая контролирует экспрессию различных генов в сочетании с ограничением калорийности или кетогенной диетой 79 .Это исследование также подтвердило, что β-гидроксибутирилирование является одной из посттрансляционных модификаций р53, который является одним из наиболее широко изученных супрессоров опухолей, а также тесно связан со старением и апоптозом. Было показано, что активность р53 точно регулируется различными посттрансляционными модификациями, включая ацетилирование, метилирование, фосфорилирование, убиквитинирование, сумоилирование и неддилирование. β-гидроксибутирилирование наблюдается в сайтах ацетилирования по лизинам 120, 319 и 370 р53.Поскольку β-гидроксибутирилирование конкурирует с ацетилированием p53, увеличение β-HB из-за ограничения калорий или кетогенной диеты может снизить ацетилирование p53, влияя на активность p53 80 . Более того, β-гидроксибутирилирование было идентифицировано на гистонах, предлагая также новый тип регуляции хроматина 79 .
Эти результаты предполагают, что опосредованные β-HB эпигенетические и посттрансляционные модификации могут играть решающую роль в регуляции экспрессии генов и передачи сигнала (рис.3, 4). Хотя регуляция β-гидроксибутирилирования и ферменты, участвующие в этом процессе, неясны, недавнее сообщение продемонстрировало, что Sirt3 является критическим ферментом, участвующим в де-β-гидроксибутирилировании HDAC 81 . Дальнейшая характеристика мишеней и регуляторов β-гидроксибутирилирования может предложить новый подход к раскрытию молекулярных механизмов β-HB в сочетании с ограничением калорийности и кетогенной диетой.
Рис. 4: Общие молекулярные механизмы, лежащие в основе эффектов, связанных с β-HB, на старение.Целевые молекулы и клеточная передача сигналов β-HB связаны с процессом старения, который ускоряется старением и воспалением. β-HB задерживает старение за счет ингибирования HDAC, hnRNP A1-опосредованной экспрессии Oct4 и β-гидроксибутирилирования на p53. Кроме того, β-HB подавляет воспаление за счет ингибирования NLRP3 или активации HCAR2 и снижает вклад в заболевания, связанные со старением.
Перспективы
Помимо действия в качестве альтернативного источника энергии, β-HB также является мощным метаболитом, который регулирует клеточные сигналы, воздействуя на различные биомолекулы.β-HB — это крошечная молекула, которая может легко проходить через клеточные мембраны и циркулировать по всему телу в кровеносных сосудах, даже достигая головного мозга через гематоэнцефалический барьер (BBB). Эта особенность обеспечивает преимущество β-HB перед другими существующими лекарствами для лечения неврологических заболеваний. Кроме того, β-HB также достигает прохождения через клеточные мембраны и регулирует белки в различных клеточных органеллах. β-HB влияет на сигнальные факторы, включая GPCR в цитоплазме, гистоны в ядре и HDAC, hnRNP A1 и p53 в цитоплазме.Многочисленные исследования показали, что β-HB улучшает нейродегенеративные заболевания и сердечно-сосудистые заболевания, связанные со старением. На сегодняшний день кетогенная диета используется для облегчения симптомов заболеваний, таких как рак, метаболический синдром, сердечно-сосудистые заболевания и нейродегенеративные заболевания, в качестве адъювантных терапевтических агентов. Подтверждение дополнительных углубленных исследований установит, что β-HB является новым вариантом лечения этих состояний. Кроме того, использование этого фактора в качестве терапии требует оптимизации терапевтической дозы β-HB посредством фармакокинетических исследований in vivo.Поскольку индивидуальные различия затрудняют контроль оптимальных уровней циркулирующего β-HB с помощью ограничения калорий или кетогенной диеты, необходимо разработать регулируемые варианты лечения, такие как введение KE. Поскольку резкие изменения в циркулирующем β-HB могут нарушить энергетический гомеостаз, хиральный энантиомер s-β-HB может предложить потенциальную возможность для лечения, поскольку эта молекула не может использоваться в качестве альтернативного энергетического метаболита. Кроме того, s-β-HB не потребляется физиологической системой, а период полураспада s-β-HB в кровотоке больше, чем у β-HB.Поскольку β-HB облегчает различные симптомы связанных с возрастом заболеваний и фенотипы старения с помощью разнообразных, но пока неизвестных молекулярных механизмов, оценка β-HB и / или s-β-HB в качестве терапевтического агента является важным подходом к лечению стареющего населения. .
β-гидроксибутират в мозге: одна молекула, несколько механизмов
Wakeman A, Dakin H (1909) О разложении β-оксимасляной кислоты и ацетоуксусной кислоты ферментами печени. J Biol Chem 6: 373–389
Google ученый
Дедкова Е.Н., Блаттер Л.А. (2014) Роль β-гидроксибутирата, его полимера поли-β-гидроксибутирата и неорганического полифосфата в здоровье и болезнях млекопитающих. Front Physiol 5: 260
PubMed PubMed Central Статья Google ученый
Кребс Х., Уильямсон Д., Бейтс М.В., Пейдж М.А., Хокинс Р. (1971) Роль кетоновых тел в гомеостазе калорий. Adv Enzyme Regul 9: 387–409
Статья Google ученый
Лонго В.Д., Маттсон М.П. (2014) Голодание: молекулярные механизмы и клиническое применение. Cell Metab 19: 181–192
CAS PubMed PubMed Central Статья Google ученый
Gonzalez SV, Nguyen NHT, Rise F, Hassel B (2005) Метаболизм экзогенного пирувата в мозге. J Neurochem 95: 284–293
CAS PubMed Статья Google ученый
Dienel GA (2012) Метаболизм лактата в мозге: открытия и противоречия.J Cereb Blood Flow Metab 32: 1107–1138
CAS PubMed Статья Google ученый
Rae C, Fekete AD, Kashem MA, Nasrallah FA, Bröer S (2012) Метаболизм, компартментация, транспорт и производство ацетата в кортикальном срезе ткани головного мозга. Neurochem Res 37: 2541–2553
CAS PubMed Статья Google ученый
Künnecke B, Cerdan S, Seelig J (1993) Церебральный метаболизм [1, 2- 13 C 2 ] глюкозы и [U- 13 C 4 ] 3-гидроксибутират в мозг крысы, обнаруженный с помощью спектроскопии ЯМР 13 C.ЯМР Биомед 6: 264–277
PubMed Статья Google ученый
Эдмонд Дж., Роббинс Р., Бергстром Дж., Коул Р., Де Веллис Дж. (1987) Способность к использованию субстрата в окислительном метаболизме нейронами, астроцитами и олигодендроцитами из развивающегося мозга в первичной культуре. J Neurosci Res 18: 551–561
CAS PubMed Статья Google ученый
Кэхилл Г.Ф., Вич Р.Л. (2003) Кетокислоты? Хорошее лекарство? Trans Am Clin Climatol Assoc 114: 149
PubMed PubMed Central Google ученый
Hansen JL, Freier EF (1978) Прямые анализы лактата, пирувата, бета-гидроксибутирата и ацетоацетата с помощью центробежного анализатора. Clin Chem 24: 475–479
CAS PubMed Google ученый
Оуэн О., Морган А., Кемп Х., Салливан Дж., Эррера М., Кэхилл Дж. Младший (1967) Метаболизм мозга во время голодания. J Clin Invest 46: 1589
CAS PubMed PubMed Central Статья Google ученый
Mitchell GA, Kassovska-Bratinova S, Boukaftane Y, Robert MF, Wang SP, Ashmarina L, Lambert M, Lapierre P, Potier E (1995) Медицинские аспекты метаболизма кетоновых тел. Clin Invest Med 18: 193–216
CAS PubMed Google ученый
Laffel L (1999) Кетоновые тела: обзор физиологии, патофизиологии и применения мониторинга при диабете. Diabetes Metab Res Rev 15: 412–426
CAS PubMed Статья Google ученый
Pan JW, Rothman DL, Behar KL, Stein DT, Hetherington HP (2000) Увеличение β-гидроксибутирата и лактата мозга человека при кетозе, вызванном голоданием. J Cereb Blood Flow Metab 20: 1502–1507
CAS PubMed Статья Google ученый
Samala R, Klein J, Borges K (2011) Кетогенная диета изменяет уровни метаболитов во внеклеточной жидкости гиппокампа. Neurochem Int 58: 5–8
CAS PubMed Статья Google ученый
Halestrap AP, Meredith D (2004) Семейство генов SLC16 — от переносчиков монокарбоксилатов (МСТ) до переносчиков ароматических аминокислот и не только. Pflugers Arch 447: 619–628
CAS PubMed Статья Google ученый
Halestrap AP (2013) Транспорт монокарбоновых кислот. Compr Physiol
Bergersen LH, Magistretti PJ, Pellerin L (2005) Избирательная постсинаптическая совместная локализация MCT2 с субъединицами рецептора AMPA GluR2 / 3 в возбуждающих синапсах, демонстрирующих передачу рецептора AMPA.Cereb Cortex 15: 361–370
PubMed Статья Google ученый
Rafiki A, Boulland JL, Halestrap AP, Ottersen OP, Bergersen LH (2003) Высокодифференциальная экспрессия транспортеров монокарбоксилата MCT2 и MCT4 в развивающемся мозге крысы. Neurosci 122: 677–688
CAS Статья Google ученый
Broer S, Broer A, Schneider HP, Stegen C, Halestrap AP, Deitmer JW (1999) Характеристика высокоаффинного переносчика монокарбоксилата MCT2 в ооцитах Xenopus laevis.Biochem J 341: 529–535
CAS PubMed PubMed Central Google ученый
Карпентер Л., Халестрап А.П. (1994) Кинетику, субстратную и ингибиторную специфичность лактатного переносчика опухолевых клеток Эрлиха-Леттра изучали с помощью индикатора внутриклеточного pH BCECF. Biochem J 304: 751–760
CAS PubMed PubMed Central Статья Google ученый
Fox JEM, Meredith D, Halestrap AP (2000) Характеристика человеческого транспортера монокарбоксилата 4 подтверждает его роль в оттоке молочной кислоты из скелетных мышц. J Physiol 529: 285–293
Статья Google ученый
Martin PM, Gopal E, Ananth S, Zhuang L, Itagaki S, Prasad BM, Smith SB, Prasad PD, Ganapathy V (2006) Идентификация SMCT1 (SLC5A8) как нейрон-специфичного Na + -связанного переносчика для активного поглощения L-лактатом и кетоновыми телами головным мозгом.J Neurochem 98: 279–288
CAS PubMed Статья Google ученый
Gopal E, Umapathy NS, Martin PM, Ananth S, Gnana-Prakasam JP, Becker H, Wagner CA, Ganapathy V, Prasad PD (2007) Клонирование и функциональная характеристика человеческого SMCT2 (SLC5A12) и паттерна экспрессии транспортера в почке. Biochim Et Biophys Acta Biomembr 1768: 2690–2697
CAS Статья Google ученый
Tildon JT, McKenna MC, Stevenson JH Jr (1994) Транспорт 3-гидроксибутирата культивированными астроцитами головного мозга крысы. Neurochem Res 19: 1237–1242
CAS PubMed Статья Google ученый
Leino RL, Gerhart DZ, Duelli R, Enerson BE, Drewes LR (2001) Кетоз, вызванный диетой, увеличивает уровни переносчика монокарбоксилата (MCT1) в мозге крысы. Neurochem Int 38: 519–527
CAS PubMed Статья Google ученый
Regen DM, Callis JT, Sugden MC (1983) Исследования церебрального транспорта β-гидроксибутирата с помощью инъекции в сонную артерию; влияние возраста, диеты и состава инъекций. Brain Res 271: 289–299
CAS PubMed Статья Google ученый
Puchowicz MA, Xu K, Sun X, Ivy A, Emancipator D, LaManna JC (2007) Кетоз, вызванный диетой, увеличивает плотность капилляров без изменения кровотока в головном мозге крысы. Am J Physiol Endocrinol Metab 292: E1607
CAS PubMed Статья Google ученый
Mikkelsen KH, Seifert T, Secher NH, Grøndal T, van Hall G (2014) Системные, церебральные и скелетные мышцы кетоновые тела и метаболизм энергии во время острой гипер-D-β-гидроксибутиратемии у здоровых мужчин после абсорбции. J Clin Endocrinol Metab 100: 636–643
PubMed Статья CAS Google ученый
Courchesne-Loyer A, Croteau E, Castellano CA, St-Pierre V, Hennebelle M, Cunnane SC (2016) Обратная связь между глюкозой в головном мозге и метаболизмом кетонов у взрослых во время краткосрочного умеренного диетического кетоза: двойная количественное исследование с использованием радиоизотопной позитронно-эмиссионной томографии.J Cereb Blood Flow Metab. DOI: 10.1177 / 0271678X16669366
Хокинс Р.А., Манс А.М., Дэвис Д.В. (1986) Использование региональных кетоновых тел мозгом крысы при голодании и диабете. Am J Physiol Endocrinol Metab 250: E169
CAS Google ученый
Ито К., Утида Ю., Охцуки С., Айзава С., Каваками Х., Кацукура Ю., Камии Дж., Терасаки Т. (2011) Количественная экспрессия мембранных белков на гематоэнцефалическом барьере взрослых и молодых яванских макак.J Pharm Sci 100: 3939–3950
CAS PubMed Статья Google ученый
Gerhart DZ, Enerson BE, Zhdankina OY, Leino RL, Drewes LR (1997) Экспрессия транспортера монокарбоксилата MCT1 эндотелием и глией головного мозга у взрослых крыс и грудных крыс. Am J Physiol Endocrinol Metab 273: E207 – E213
CAS Google ученый
Vannucci SJ, Simpson IA (2003) Переключатель развития в экспрессии переносчика питательных веществ в головном мозге у крысы.Am J Physiol Endocrinol Metab 285: E1127 – E1134
CAS PubMed Статья Google ученый
Uchida Y, Ohtsuki S, Katsukura Y, Ikeda C, Suzuki T, Kamiie J, Terasaki T (2011) Количественная целевая абсолютная протеомика переносчиков и рецепторов гематоэнцефалического барьера человека. J Neurochem 117: 333–345
CAS PubMed Статья Google ученый
Halestrap AP (1978) Транспорт пирувата и кетоновых тел через митохондриальную мембрану. Обменные свойства, pH-зависимость и механизм носителя. Biochem J 172: 377–387
CAS PubMed PubMed Central Статья Google ученый
Paradies G, Papa S (1997) О кинетике и субстратной специфичности транслокатора пирувата в митохондриях печени крысы. Biochim Et Biophys Acta Bioenerg 462: 333–346
Статья Google ученый
Бут Р.Ф., Кларк Дж.Б. (1981) Энергетический метаболизм в головном мозге крысы: ингибирование декарбоксилирования пирувата 3-гидроксибутиратом в митохондриях новорожденных. J Neurochem 37: 179–185
CAS PubMed Статья Google ученый
Вич Р.Л. (2004) Терапевтическое значение кетоновых тел: эффекты кетоновых тел при патологических состояниях: кетоз, кетогенная диета, окислительно-восстановительные состояния, инсулинорезистентность и митохондриальный метаболизм.Простагландин, лейкотриен, эссент, жирная кислота, 70: 309–319
CAS Статья Google ученый
Sato K, Kashiwaya Y, Keon C, Tsuchiya N, King M, Radda G, Chance B, Clarke K, Veech R (1995) Кетоновые тела инсулина и митохондриальная трансдукция энергии. FASEB J 9: 651–658
CAS PubMed Google ученый
Klee CB, Sokoloff L (1967) Изменения активности d (-) — β-гидроксимасляной дегидрогеназы во время созревания мозга у крыс.J Biol Chem 242: 3880–3883
CAS PubMed Google ученый
Latruffe N, Gaudemer Y (1974) Propriétés et mécanisme cinétique de la D (-) β-hydroxybutyrique déshydrogénase de parisules sous-mitochondriales de foie de rat; Сравнение различных агентов тиолов. Biochimie 56: 435–444
CAS PubMed Статья Google ученый
Rardin MJ, Newman JC, Held JM, Cusack MP, Sorensen DJ, Li B, Schilling B, Mooney SD, Kahn CR, Verdin E (2013) Количественная протеомика ацетилома лизина в митохондриях, не содержащая этикеток. субстраты SIRT3 в метаболических путях.Proc Natl Acad Sci USA 110: 6601–6606
CAS PubMed PubMed Central Статья Google ученый
Hebert AS, Dittenhafer-Reed KE, Yu W, Bailey DJ, Selen ES, Boersma MD, Carson JJ, Tonelli M, Balloon AJ, Higbee AJ, Westphall MS, Pagliarini DJ, Prolla TA, Assadi-Porter F, Roy S, Denu JM Coon JJ (2013) Ограничение калорий и SIRT3 запускают глобальное перепрограммирование ацетилома митохондриального белка. Mol Cell 49: 186–199
CAS PubMed Статья Google ученый
Guo K, Lukacik P, Papagrigoriou E, Meier M, Lee WH, Adamski J, Oppermann U (2006) Характеристика человеческого DHRS6, бесхозного фермента дегидрогеназы / редуктазы с короткой цепью: новая цитозольная R-бета-гидроксибутиратдегидрогеназа 2 типа . J Biol Chem 281: 10291–10297
CAS PubMed Статья Google ученый
Чанг Х.Т., Олсон Л.К., Шварц К.А. (2013) Профили экспрессии кетолитических и гликолитических ферментов в злокачественных глиомах: значение для терапии кетогенной диетой.Нутр Метаб 10: 1
CAS Статья Google ученый
Bixel MG, Hamprecht B (1995) Получение кетоновых тел из лейцина культивированными астроглиальными клетками. J Neurochem 65: 2450–2461
CAS PubMed Статья Google ученый
Puisac B, Ramos M, Arnedo M, Menao S, Gil-Rodríguez MC, Teresa-Rodrigo ME, Pié A, de Karam JC, Wesselink JJ, Giménez I, Ramos FJ, Casals N, Gómez-Puertas P, Hegardt FG, Pié J (2012) Характеристика вариантов сплайсинга генов, кодирующих митохондриальную HMG-CoA-лиазу человека и HMG-CoA-синтазу, основные ферменты пути кетогенеза.Mol Biol Rep 39: 4777–4785
CAS PubMed Статья Google ученый
Fukao T, Song XQ, Mitchell GA, Yamaguchi S, Sukegawa K, Or T, Kondo N (1997) Ферменты использования кетоновых тел в тканях человека: уровни белка и информационной РНК сукцинил-коэнзима A (CoA ): 3-кетокислотная трансфераза КоА и митохондриальные и цитозольные ацетоацетил-КоА тиолазы. Pediatr Res 42: 498–502
CAS PubMed Статья Google ученый
Ohnuki M, Takahashi N, Yamasaki M, Fukui T (2005) Различная локализация в мозге крысы нового фермента, использующего цитозольные кетоновые тела, ацетоацетил-CoA-синтетазы, по сравнению с сукцинил-CoA: 3-оксоацид-CoA-трансферазой. Biochim Et Biophys Acta 1729: 147–153
CAS Статья Google ученый
Endemann G, Goetz PG, Edmond J, Brunengraber H (1982) Липогенез из кетоновых тел в изолированной перфузированной печени крысы.Доказательства цитозольной активации ацетоацетата. J Biol Chem 257: 3434–3440
CAS PubMed Google ученый
Hasegawa S, Kume H, Iinuma S, Yamasaki M, Takahashi N, Fukui T (2012) Ацетоацетил-КоА-синтетаза необходима для нормального развития нейронов. Biochem Biophys Res Commun 427: 398–403
CAS PubMed Статья Google ученый
Tildon JT, Cornblath M (1972) Succinyl-CoA: дефицит 3-кетокислоты CoA-трансферазы. Причина кетоацидоза в младенчестве. J Clin Invest 51: 493–498
CAS PubMed PubMed Central Статья Google ученый
Fukao T, Sass JO, Kursula P, Thimm E, Wendel U, Ficicioglu C, Monastiri K, Guffon N, Barić I, Zabot MT, Kondo N (2011) Клиническая и молекулярная характеристика пяти пациентов с сукцинилом -CoA: дефицит 3-кетокислоты CoA трансферазы (SCOT).Biochim Et Biophys Acta 1812: 619–624
CAS Статья Google ученый
Fukao T, Mitchell GA, Song XQ, Nakamura H, Kassovska-Bratinova S, Orii KE, Wraith JE, Besley G, Wanders RJA, Niezen-Koning KE, Berry GT, Palmieri M, Kondo N (2000 ) Сукцинил-КоА: трансфераза 3-кетокислоты КоА (SCOT): клонирование гена SCOT человека, третичное структурное моделирование мономера SCOT человека и характеристика трех патогенных мутаций.Геномика 68: 144–151
CAS PubMed Статья Google ученый
Берри Г., Фукао Т., Митчелл Дж., Мазур А., Циафре М., Гибсон Дж., Кондо Н., Пальмиери М. (2001) Гипогликемия новорожденных при тяжелой недостаточности сукцинил-КоА: 3-оксоацид-КоА-трансферазы. J Inherit Metab Dis 24: 587–595
CAS PubMed Статья Google ученый
Гибсон К., Брейер Дж., Нихан В. (1988) Дефицит лиазы 3-гидрокси-3-метилглутарил-кофермента А: обзор 18 пациентов, о которых сообщалось.Eur J Pediatr 148: 180–186
CAS PubMed Статья Google ученый
Ван дер Кнаап М., Баккер Х., Валк Дж. (1998) МР-визуализация и протонная спектроскопия при дефиците лиазы 3-гидрокси-3-метилглутарил-кофермента А. Am J Neuroradiol 19: 378–382
PubMed Google ученый
Gordon K, Riding M, Camfield P, Bawden H, Ludman M, Bagnell P (1994) CT и MR дефицита лиазы 3-гидрокси-3-метилглутарил-коэнзима А.Am J Neuroradiol 15: 1474–1476
CAS PubMed Google ученый
Fukao T, Scriver CR, Kondo N; Группа TCW (2001) Клинический фенотип и исход митохондриальной недостаточности ацетоацетил-КоА тиолазы (β-кетотиолаза или дефицит Т2) у 26 пациентов с подтвержденным ферментативным тестом и определенными мутациями. Mol Genet Metab 72: 109–114
CAS PubMed Статья Google ученый
Беннетт М., Хоскинг Г., Смит М., Грей Р., Миддлтон Б. (1984) Биохимические исследования пациента с дефектом цитозольной ацетоацетил-КоА тиолазы, связанным с умственной отсталостью. J Inherit Metab Dis 7: 125–128
CAS PubMed Статья Google ученый
Хокинс Р., Уильямсон Д., Кребс Х. (1971) Использование кетоновых тел мозгом взрослых и сосущих крыс in vivo. Biochem J 122: 13–18
CAS PubMed PubMed Central Статья Google ученый
Соколов Л. (1973) Метаболизм кетоновых тел головным мозгом. Annu Rev Med 24: 271–280
CAS PubMed Статья Google ученый
Кэхилл GFJ (2006) Топливный метаболизм при голодании. Annu Rev Nutr 26: 1–22
CAS PubMed Статья Google ученый
DeVivo DC, Leckie MP, Agrawal HC (1973) Дифференциальное включение β-гидроксибутирата и глюкозы в глутамат головного мозга у новорожденной крысы.Brain Res 55: 485–490
CAS PubMed Статья Google ученый
Nehlig A (2004) Поглощение мозгом и метаболизм кетоновых тел в моделях на животных. Простагландин, лейкотриен, эссент, жирная кислота, 70: 265–275
CAS Статья Google ученый
Хотта С.С. (1962) Метаболизм глюкозы в ткани мозга: шунт гексозомонофосфата и его роль в восстановлении глутатиона.J Neurochem 9: 43–51
CAS PubMed Статья Google ученый
Bilger A, Nehlig A (1992) Количественные гистохимические изменения ферментов, участвующих в энергетическом метаболизме в мозге крысы во время постнатального развития 2. Глюкозо-6-фосфатдегидрогеназа и бета-гидроксибутиратдегидрогеназа. Int J Dev Neurosci 10: 143–152
CAS PubMed Статья Google ученый
Nehlig A, de Vasconcelos AP (1993) Использование глюкозы и кетоновых тел мозгом новорожденных крыс. Прог Нейробиол 40: 163–220
CAS PubMed Статья Google ученый
Rho JM, Stafstrom CE (2012) Кетогенная диета как парадигма лечения различных неврологических расстройств. Фронт Pharmacol 3:59
PubMed PubMed Central Google ученый
Maalouf M, Rho JM, Mattson MP (2009) Нейрозащитные свойства ограничения калорий, кетогенной диеты и кетоновых тел. Brain Res Rev 59: 293–315
CAS PubMed Статья Google ученый
McKenna MC, Tildon JT, Stevenson JH, Boatright R, Huang S (1993) Регулирование энергетического метаболизма в синаптических окончаниях и культивируемых астроцитах головного мозга крысы: различия, выявленные с использованием аминооксиацетата. Dev Neurosci 15: 320–329
CAS PubMed Статья Google ученый
Chechik T, Roeder L, Tildon J, Poduslo S (1987) Активность ферментов кетонового тела в очищенных нейронах, астроцитах и олигодендроглии. Neurochem Int 10: 95–99
CAS PubMed Статья Google ученый
Берл С., Фриджеси Т.Л. (1969) Обмен глутамата, глутамина, аспартата и ГАМК, меченных 1- 14 С-ацетатом, в хвостатом ядре таламуса и моторной коре (кошка). Brain Res 12: 444–455
CAS PubMed Статья Google ученый
Berl S, Nicklas WJ, Clarke DD (1970) Компартментация метаболизма цикла лимонной кислоты в маркировке мозга глутамата, глутамина, аспартата и ГАМК несколькими радиоактивными метаболитами. J Neurochem 17: 1009–1015
CAS PubMed Статья Google ученый
Cremer JE (1971) Включение метки d-β-гидрокси- [ 14 C] бутирата и [3- 14 C] ацетоацетата в аминокислоты в мозге крысы in vivo.Biochem J 122: 135–138
CAS PubMed PubMed Central Статья Google ученый
Pan JW, de Graaf RA, Petersen KF, Shulman GI, Hetherington HP, Rothman DL (2002) [2,4- 13 C 2 ] метаболизм -гидроксибутирата в мозге человека. J Cereb Blood Flow Metab 22: 890–898
CAS PubMed PubMed Central Статья Google ученый
Leong SF, Lai JCK, Lim L, Clark JB (1981) Ферменты, метаболизирующие энергию в областях мозга взрослых и стареющих крыс. J Neurochem 37: 1548–1556
CAS PubMed Статья Google ученый
Williamson D, Bates MW, Page MA, Krebs H (1971) Активность ферментов, участвующих в утилизации ацетоацетата в тканях взрослых млекопитающих. Biochem J 121: 41–47
CAS PubMed PubMed Central Статья Google ученый
Пейдж М.А., Уильямсон Д. (1971) Ферменты утилизации кетоновых тел в человеческом мозге. Ланцет 298: 66–68
Артикул Google ученый
Nugent S, Tremblay S, Chen KW, Ayutyanont N, Roontiva A, Castellano CA, Fortier M, Roy M, Courchesne-Loyer A, Bocti C (2014) Метаболизм глюкозы и ацетоацетата в мозге: сравнение молодых и пожилые люди. Neurobiol Aging 35: 1386–1395
CAS PubMed Статья Google ученый
Blomqvist G, Thorell J, Ingvar M, Grill V, Widen L, Stone-Elander S (1995) Использование R-бета- [1- 11 C] гидроксибутирата в исследованиях ПЭТ регионального поглощения кетоновых тел у людей . Am J Physiol Endocrinol Metab 269: E948 – E959
CAS Google ученый
Hawkins RA, Biebuyck JF (1979) Кетоновые тела выборочно используются отдельными областями мозга. Наука 205: 325–327
CAS PubMed Статья Google ученый
Gesink DS, Wilson JE A (1985) Гистохимическое исследование распределения β-гидроксибутиратдегидрогеназы в развивающемся мозжечке крысы. J Neurochem 44: 1308–1311
CAS PubMed Статья Google ученый
Аракава Т., Гото Т., Окада Ю. (1991) Влияние кетонового тела (D-3-гидроксибутирата) на нервную активность и энергетический метаболизм в срезах гиппокампа взрослых морских свинок. Neurosci Lett 130: 53–56
CAS PubMed Статья Google ученый
Wada H, Okada Y, Nabetani M, Nakamura H (1997) Влияние лактата и бета-гидроксибутирата на энергетический метаболизм и нервную активность срезов гиппокампа взрослых и неполовозрелых крыс. Dev Brain Res 101: 1–7
CAS Статья Google ученый
Weiss JN, Lamp ST (1989) Сердечный АТФ-чувствительный K + каналов. Доказательства предпочтительного регулирования гликолиза. J Gen Physiol 94: 911–935
CAS PubMed Статья Google ученый
Dhar-Chowdhury P, Harrell MD, Han SY, Jankowska D, Parachuru L, Morrissey A, Srivastava S, Liu W., Malester B, Yoshida H (2005) Гликолитические ферменты, глицеральдегид-3-фосфатдегидрогеназа, триозофосфат-изомераза , и пируваткиназа являются компонентами макромолекулярного комплекса KATP-канала и регулируют его функцию. J Biol Chem 280: 38464–38470
CAS PubMed PubMed Central Статья Google ученый
Silver IA, Deas J, Erecinska M (1997) Гомеостаз ионов в клетках мозга: различия во внутриклеточных реакциях ионов на ограничение энергии между культивируемыми нейронами и глиальными клетками. Неврология 78: 598–601
Статья Google ученый
Chowdhury GM, Jiang L, Rothman DL, Behar KL (2014) Вклад кетоновых тел в базальное и зависимое от активности окисление нейронов in vivo. J Cereb Blood Flow Metab 34: 1233–1242
CAS PubMed PubMed Central Статья Google ученый
Wieland O, Funcke Hv, Löffler G (1971) Взаимное превращение пируватдегидрогеназы в сердечной мышце крысы при перфузии жирными кислотами или кетоновыми телами. FEBS Lett 15: 295–298
CAS PubMed Статья Google ученый
Ньюсхолм Е.А., Рэндл П.Дж., Манчестер К.Л. (1962) Ингибирование реакции фосфофруктокиназы в перфузируемом сердце крысы путем дыхания кетоновых тел, жирных кислот и пирувата. Nature 193: 270–271
CAS PubMed Статья Google ученый
Tildon JT, Roeder LM, Stevenson JH (1985) Окисление субстрата изолированными митохондриями и синаптосомами головного мозга крысы. J Neurosci Res 14: 207–215
CAS PubMed Статья Google ученый
Marosi K, Kim SW, Moehl K, Scheibye-Knudsen M, Cheng A, Cutler R, Camandola S, Mattson MP (2016) 3-гидроксибутират регулирует энергетический метаболизм и индуцирует экспрессию BDNF в нейронах коры головного мозга. J Neurochem. DOI: 10.1111 / JNC.13868
PubMed Google ученый
Erecinska M, Nelson D, Daikhin Y, Yudkoff M (1996) Регулирование уровня ГАМК в синаптосомах мозга крыс: потоки через ферменты шунта ГАМК и эффекты глутамата, кальция и кетоновых тел. J Neurochem 67: 2325–2334
CAS PubMed Статья Google ученый
Юдкофф М., Дайхин Ю., Ниссим И., Грюнштейн Р., Ниссим И. (1997) Влияние кетоновых тел на метаболизм аминокислот астроцитов.J Neurochem 69: 682–692
CAS PubMed Статья Google ученый
McKenna MC, Tildon JT, Stevenson J, Huang X, Kingwell KG (1995) Регулирование митохондриальных и цитозольных яблочных ферментов из культивированных астроцитов головного мозга крысы. Neurochem Res 20: 1491–1501
CAS PubMed Статья Google ученый
Lajtha A, Gibson G, Dienel G (2007) Справочник по нейрохимии и молекулярной нейробиологии.Нейроактивные белки и пептиды. Springer, Springer-Verlag
Книга Google ученый
Amaral AI, Teixeira AP, Sonnewald U, Alves PM (2011) Оценка внутриклеточных потоков в нейронах мозжечка после гипогликемии: важность пути рециркуляции пирувата и окисления глутамина. J Neurosci Res 89: 700–710
CAS PubMed Статья Google ученый
Lund TM, Risa O, Sonnewald U, Schousboe A, Waagepetersen HS (2009) Доступность глутамата нейромедиатора уменьшается, когда бета-гидроксибутират заменяет глюкозу в культуре. J Neurochem 110: 80–91
CAS PubMed Статья Google ученый
Cruz NF, Lasater A, Zielke HR, Dienel GA (2005) Активация астроцитов в мозгу крыс в сознании во время акустической стимуляции: утилизация ацетата в рабочем мозге.J Neurochem 92: 934–947
CAS PubMed Статья Google ученый
Nicklas WJ, Youngster SK, Kindt MV, Heikkila RE IV (1987) MPTP, MPP + и функция митохондрий. Life Sci 40: 721–729
CAS PubMed Статья Google ученый
Tieu K, Perier C, Caspersen C, Teismann P, Wu DC, Yan SD, Naini A, Vila M, Jackson-Lewis V, Ramasamy R (2003) D-β-Hydroxybutyrate спасает митохондриальное дыхание и смягчает особенности болезни Паркинсона.J Clin Invest 112: 892–901
CAS PubMed PubMed Central Статья Google ученый
Edwards C, Canfield J, Copes N, Rehan M, Lipps D, Bradshaw PC (2014) D-бета-гидроксибутират продлевает продолжительность жизни C. elegans. Старение 6: 621
CAS PubMed PubMed Central Статья Google ученый
Zhao Z, Lange DJ, Voustianiouk A, MacGrogan D, Ho L, Suh J, Humala N, Thiyagarajan M, Wang J, Pasinetti GM (2006) Кетогенная диета как потенциальное новое терапевтическое вмешательство при боковом амиотрофе. склероз.BMC Neurosci 7: 1
CAS Статья Google ученый
Kashiwaya Y, Takeshima T, Mori N, Nakashima K, Clarke K, Veech RL (2000) d-β-гидроксибутират защищает нейроны в моделях болезни Альцгеймера и Паркинсона. Proc Natl Acad Sci USA 97: 5440–5444
CAS PubMed PubMed Central Статья Google ученый
Julio-Amilpas A, Montiel T, Soto-Tinoco E, Gerónimo-Olvera C, Massieu L (2015) Защита от гибели нейронов, вызванной гипогликемией, с помощью β-гидроксибутирата включает сохранение уровней энергии и снижение выработки активные формы кислорода.J Cereb Blood Flow Metab 35: 851–860
CAS PubMed PubMed Central Статья Google ученый
Маккенна М., Тилдон Дж., Стивенсон Дж., Хопкинс I (1994) Энергетический метаболизм в корковых синаптических окончаниях головного мозга отъемных и зрелых крыс: данные о множественных компартментах активности цикла трикарбоновых кислот. Dev Neurosci 16: 291–300
CAS PubMed Статья Google ученый
Lund TM, Ploug KB, Iversen A, Jensen AA, Jansen-Olesen I (2015) Метаболическое влияние β-гидроксибутирата на нейротрансмиссию: снижение гликолиза опосредует изменения в ответах на кальций и чувствительность рецепторов канала KATP. J Neurochem 132: 520–531
CAS PubMed Google ученый
Ma W, Berg J, Yellen G (2007) Метаболиты кетогенной диеты уменьшают возбуждение в центральных нейронах, открывая каналы KATP. J Neurosci 27: 3618–3625
CAS PubMed Статья Google ученый
Хименес-Кассина А., Мартинес-Франсуа Хуан Р., Фишер Джилл К., Шлик Б., Полак К., Вивчар Дж., Таннер Джеффри Р., Лутас А., Йеллен Г. (2012) Даниал Ника Н. BAD-зависимая регуляция метаболизма топлива и канал KATP активность придает устойчивость к эпилептическим припадкам. Нейрон 74: 719–730
PubMed PubMed Central Статья CAS Google ученый
Xiao X-Q, Zhao Y, Chen G-Q (2007) Влияние 3-гидроксибутирата и его производных на рост глиальных клеток.Биоматериалы 28: 3608–3616
CAS PubMed Статья Google ученый
Kimura I, Inoue D, Maeda T, Hara T, Ichimura A, Miyauchi S, Kobayashi M, Hirasawa A, Tsujimoto G (2011) Короткоцепочечные жирные кислоты и кетоны напрямую регулируют симпатическую нервную систему через G-белок -связанный рецептор 41 (GPR41). Proc Natl Acad Sci USA 108: 8030–8035
CAS PubMed PubMed Central Статья Google ученый
Won Y-J, Lu VB, Puhl HL, Ikeda SR (2013) β-гидроксибутират модулирует кальциевые каналы N-типа в симпатических нейронах крыс, действуя как агонист рецептора FFA3, связанного с G-белком. J Neurosci 33: 19314–19325
CAS PubMed PubMed Central Статья Google ученый
Shimazu T, Hirschey MD, Newman J, He W., Shirakawa K, Le Moan N, Grueter CA, Lim H, Saunders LR, Stevens RD (2013) Подавление окислительного стресса β-гидроксибутиратом, эндогенным веществом. ингибитор гистондеацетилазы.Наука 339: 211–214
CAS PubMed Статья Google ученый
Mejía-Toiber J, Montiel T, Massieu L (2006) D-β-гидроксибутират предотвращает опосредованное глутаматом липопероксидацию и повреждение нейронов, вызываемое во время ингибирования гликолиза in vivo. Neurochem Res 31: 1399–1408
PubMed Статья CAS Google ученый
Yamada T, Zhang S-J, Westerblad H, Katz A (2010) β-гидроксибутират ингибирует опосредованный инсулином транспорт глюкозы в окислительных мышцах мыши.Am J Physiol Endocrinol Metab 299: E364 – E373
CAS PubMed Статья Google ученый
Youm YH, Nguyen KY, Grant RW, Goldberg EL, Bodogai M, Kim D, D’Agostino D, Planavsky N, Lupfer C, Kanneganti TD, Kang S, Horvath TL, Fahmy TM, Crawford PA, Biragyn A, Alnemri E, Dixit VD (2015) β-гидроксибутират кетоновых тел блокирует воспалительное заболевание, опосредованное воспалением NLRP3. Nat Med 21: 263–269
CAS PubMed PubMed Central Google ученый
Tschopp J, Schroder K (2010) Активация инфламмасомы NLRP3: конвергенция множественных сигнальных путей на продукцию ROS? Nat Rev Immunol 10: 210–215
CAS PubMed Статья Google ученый
Шао Б-З, Сюй З-К, Хан Б-З, Су Д-Ф, Лю С. (2015) Инфламмасома NLRP3 и ее ингибиторы: обзор. Front Pharmacol 6: 262
PubMed PubMed Central Статья CAS Google ученый
Lee G-S, Subramanian N, Kim AI, Aksentijevich I, Goldbach-Mansky R, Sacks DB, Germain RN, Kastner DL, Chae JJ (2012) Чувствительный к кальцию рецептор регулирует воспаление NLRP3 через Ca 2+ и цАМФ. Nature 492: 123–127
CAS PubMed PubMed Central Статья Google ученый
Фрам Дж., Мербольдт К.Д., Ханике В. (1987) Локализованная протонная спектроскопия с использованием стимулированного эхо. J Magn Reson 72: 502–508
CAS Google ученый
Крейс Р., Росс Б. (1992) Нарушения церебрального метаболизма у пациентов с подострым и хроническим сахарным диабетом: обнаружение с помощью протонной МР-спектроскопии. Радиология 184: 123–130
CAS PubMed Статья Google ученый
Plecko B, Stoeckler-Ipsiroglu S, Schober E, Harrer G, Mlynarik V, Gruber S, Moser E, Moeslinger D, Silgoner H, Ipsiroglu O (2002) Пероральные добавки -гидроксибутирата у двух пациентов с гиперинсулинемическая гипогликемия: мониторинг уровней β-гидроксибутирата в крови и спинномозговой жидкости, а также в головном мозге с помощью магнитно-резонансной спектроскопии in vivo.Pediatric Res 52: 301–306
CAS Google ученый
Шен Дж., Новотный Е.Дж., Ротман Д.Л. (1998) Редактирование лактата и β-гидроксибутирата in vivo с использованием серии импульсов перефокусировки чистой фазы. Magn Reson Med 40: 783–788
CAS PubMed Статья Google ученый
Ордидж Р., Коннелли А., Ломан Дж. (1969) Спектроскопия in vivo с отбором изображений (ISIS). Новый метод пространственно-селективной спектроскопии ЯМР.Дж. Магн Резон 66: 283–294
Google ученый
Wootton-Gorges SL, Buonocore MH, Kuppermann N, Marcin J, DiCarlo J, Neely EK, Barnes PD, Glaser N (2005) Обнаружение церебрального β-гидроксибутирата, ацетоацетата и лактата с помощью протонной МР-спектроскопии у детей с диабетическим кетоацидозом. Am J Neuroradiol 26: 1286–1291
PubMed Google ученый
Cecil KM, Mulkey SB, Ou X, Glasier CM (2015) Кетоны мозга, обнаруженные с помощью протонной магнитно-резонансной спектроскопии у младенца с синдромом Охтахара, получавшего кетогенную диету.Педиатр Радиол 45: 133–137
PubMed Статья Google ученый
Mescher M, Merkle H, Kirsch J, Garwood M, Gruetter R (1998) Одновременное редактирование спектров in vivo и подавление воды. ЯМР Биомед 11: 266–272
CAS PubMed Статья Google ученый
Эдден РАЭ, Харрис А.Д., Мерфи К., Эванс К.Дж., Саксена Н., Холл Дж. Э., Бейли Д.М., Wise RG (2010) Под редакцией MRS чувствительна к изменениям концентрации лактата во время инспираторной гипоксии.J Magn Reson Imag 32: 320–325
Статья Google ученый
Courchesne-Loyer A, Fortier M, Tremblay-Mercier J, Chouinard-Watkins R, Roy M, Nugent S, Castellano CA, Cunnane SC (2013) Стимуляция умеренной устойчивой кетонемии триацилглицеринами со средней длиной цепи в здоровые люди: предполагаемый потенциальный вклад в энергетический метаболизм мозга. Питание 29: 635–640
CAS PubMed Статья Google ученый
Регер М.А., Хендерсон С.Т., Хейл С., Холертон Б., Бейкер Л.Д., Уотсон Г., Хайд К., Чепмен Д., Крафт С. (2004) Влияние β-гидроксибутирата на познание у взрослых с нарушенной памятью. Neurobiol Aging 25: 311–314
CAS PubMed Статья Google ученый
Clarke K, Tchabanenko K, Pawlosky R, Carter E, King MT, Musa-Veloso K, Ho M, Roberts A, Robertson J, VanItallie TB (2012) Кинетика, безопасность и переносимость (R) — 3-гидроксибутил (R) -3-гидроксибутират у здоровых взрослых людей.Regul Toxicol Pharmacol 63: 401–408
CAS PubMed Статья Google ученый
Newport MT, VanItallie TB, Kashiwaya Y, King MT, Veech RL (2015) Новый способ вызвать гиперкетонемию: использование кетонового эфира в случае болезни Альцгеймера. Альцгеймерс Демент 11: 99–103
PubMed Статья Google ученый
Hertz L, Chen Y, Waagepetersen HS (2015) Влияние кетоновых тел при болезни Альцгеймера на нервный гипометаболизм, токсичность β-амилоида и функцию астроцитов.J Neurochem 134: 7–20
CAS PubMed Статья Google ученый
Орхан Н., Угур Йилмаз С., Экизоглу О, Ахишали Б., Кучук М., Арикан Н., Элмас И., Гюрсес С., Кая М. (2016) Влияние бета-гидроксибутирата на проницаемость сосудов головного мозга у крыс с травматическим мозгом травма, повреждение. Brain Res 1631: 113–126
CAS PubMed Статья Google ученый
Jarrett SG, Milder JB, Liang LP, Patel M (2008) Кетогенная диета увеличивает уровни митохондриального глутатиона. J Neurochem 106: 1044–1051
CAS PubMed Статья Google ученый
Принс М., Фудзима Л., Ховда Д. (2005) Зависимое от возраста уменьшение объема кортикального ушиба кетонами после черепно-мозговой травмы. J Neurosci Res 82: 413–420
CAS PubMed Статья Google ученый
Bröer S, Schneider HP, Bröer A, Rahman B, Hamprecht B, Deitmer JW (1998) Характеристика переносчика монокарбоксилата 1, экспрессируемого в ооцитах Xenopus laevis , по изменениям цитозольного pH. Biochem J 333: 167–174
PubMed PubMed Central Статья Google ученый
Бета-гидроксибутират (3-OHB) может влиять на энергетический фенотип клеток рака груди, но не влияет на их пролиферацию и реакцию на химиотерапию или облучение | Рак и обмен веществ
DeSantis CE, Fedewa SA, Goding Sauer A, Kramer JL, Smith RA, Jemal A. Статистика рака груди, 2015: сближение показателей заболеваемости между черными и белыми женщинами. CA Cancer J Clin. 2016; 66: 31–42.
PubMed Статья Google ученый
Гнант М., Томссен С., Харбек Н. Санкт-Галлен / Вена 2015: краткое изложение консенсусной дискуссии. Уход за грудью (Базель). 2015; 10: 124–30.
Артикул Google ученый
Хуанг Дж., Пан Дж., Цзян Х., Ли В., Донг Дж., Чжан Х, Джи Х, Чжу З. Метаанализ между потреблением углеводов с пищей и риском колоректального рака: данные 17 обсервационных исследований. Biosci Rep. 2017; 37
Йе Й, Ву Й, Сюй Дж, Дин К., Шань Х, Ся Д. Связь между потреблением углеводов с пищей, гликемическим индексом и гликемической нагрузкой и риском рака желудка. Eur J Nutr. 2017; 56: 1169–77.
PubMed Статья CAS Google ученый
van den Brandt PA, Schulpen M. Приверженность средиземноморской диете и риск рака груди в постменопаузе: результаты когортного исследования и метаанализа. Int J Cancer. 2017; 140: 2220–31.
PubMed Статья CAS Google ученый
Palacios C, Daniel CR, Tirado-Gomez M, Gonzalez-Mercado V, Vallejo L, Lozada J, Ortiz A, Hughes DC, Basen-Engquist K. Модели питания при пуэрториканском и мексиканско-американском раке груди выжившие: пилотное исследование.J Immigr Minor Health. 2017; 19: 341–8.
PubMed PubMed Central Статья Google ученый
Огюстин Л.С., Весы М., Криспо А., Гримальди М., Де Лаурентис М., Ринальдо М., Д’Аюто М., Каталано Ф., Банна Г., Феррау Ф. и др. Диета с низким гликемическим индексом, упражнения и витамин D для уменьшения рецидивов рака груди (DEDiCa): дизайн клинического исследования. BMC Рак. 2017; 17:69.
PubMed PubMed Central Статья CAS Google ученый
Варбург О. О нарушении дыхания в раковых клетках. Наука. 1956; 124: 269–70.
PubMed CAS Google ученый
Warburg OP, Karl, Negelein E. Über den Stoffwechsel der Carcinomzelle. В: Biochemische Zeitschrift, vol 152; 1924. с. 115–47.
Google ученый
Клемент Р. Дж., Каммерер Ю. Есть ли роль ограничения углеводов в лечении и профилактике рака? Нутр Метаб (Лондон).2011; 8: 75.
Артикул CAS Google ученый
Ryu TY, Park J, Scherer PE. Гипергликемия как фактор риска прогрессирования рака. Diabetes Metab J. 2014; 38: 330–6.
PubMed PubMed Central Статья Google ученый
Monzavi-Karbassi B, Gentry R, Kaur V, Siegel ER, Jousheghany F, Medarametla S, Fuhrman BJ, Safar AM, Hutchins LF, Kieber-Emmons T.Предварительная диагностика уровня глюкозы в крови и прогноз у женщин с раком груди. Cancer Metab. 2016; 4: 7.
PubMed PubMed Central Статья Google ученый
Тан-Шалаби Дж. Л., Каррик Дж., Эдингер К., Дженовезе Д., Лиман А. Д., Пассеро В. А., Шах РБ. Модифицированная диета Аткинса при запущенных злокачественных новообразованиях — окончательные результаты испытаний безопасности и осуществимости в системе здравоохранения Питтсбурга по делам ветеранов. Нутр Метаб (Лондон). 2016; 13:52.
Артикул CAS Google ученый
Клемент Р.Ф., Ричард Д., Гросс ЕС, Чемпион CE, Д’Агостино Д.П., Файн Э.Дж., Каммерер Ю., Пофф А., Ро Дж. М., Сейфрид Т. Н., Шек А.С.. Необходим новый обзор статьи о кетогенных диетах для больных раком. Med Oncol. 2017; 34: 1–4.
Артикул CAS Google ученый
Шварц К., Чанг Х.Т., Николай М., Перниконе Дж., Ри С., Олсон К., Курниали П.С., Хорд Н.Г., Ноэль М.Лечение пациентов с глиомой с помощью кетогенной диеты: отчет о двух случаях, получавших одобренный IRB протокол кетогенной диеты с ограничением энергии, и обзор литературы. Cancer Metab. 2015; 3: 3.
PubMed PubMed Central Статья Google ученый
Ригер Дж., Бахр О., Маурер Дж. Д., Хаттинген Е., Франц К., Брукер Д., Валента С., Каммерер Ю., Кой Дж. Ф., Веллер М., Штайнбах Дж. П.. ERGO: пилотное исследование кетогенной диеты при рецидивирующей глиобластоме.Int J Oncol. 2014; 44: 1843–52.
PubMed PubMed Central Статья CAS Google ученый
Champ CE, Палмер Дж. Д., Волек Дж. С., Вернер-Васик М., Эндрюс Д. В., Эванс Дж. Дж., Гласс Дж., Ким Л., Ши В. Нацеливание на метаболизм с помощью кетогенной диеты во время лечения мультиформной глиобластомы. J Neuro-Oncol. 2014; 117: 125–31.
Артикул CAS Google ученый
Fine EJ, Segal-Isaacson CJ, Feinman RD, Herszkopf S, Romano MC, Tomuta N, Bontempo AF, Negassa A, Sparano JA. Нацеленность на ингибирование инсулина в качестве метаболической терапии при запущенном раке: пилотное исследование безопасности и осуществимости диет с участием 10 пациентов. Питание. 2012; 28: 1028–35.
PubMed Статья CAS Google ученый
Шмидт М., Пфетцер Н., Шваб М., Штраус И., Каммерер У. Влияние кетогенной диеты на качество жизни у 16 пациентов с запущенным раком: пилотное исследование.Нутр Метаб (Лондон). 2011; 8: 54.
Артикул CAS Google ученый
Wheless JW. История кетогенной диеты. Эпилепсия. 2008; 49 (Дополнение 8): 3–5.
PubMed Статья Google ученый
Лаффель Л. Кетоновые тела: обзор физиологии, патофизиологии и применения мониторинга при диабете. Diabetes Metab Res Rev.1999; 15: 412–26.
PubMed Статья CAS Google ученый
Pan JW, Rothman TL, Behar KL, Stein DT, Hetherington HP. Повышение уровня бета-гидроксибутирата и лактата в мозге человека при кетозе, вызванном голоданием. J Cereb Blood Flow Metab. 2000; 20: 1502–7.
PubMed Статья CAS Google ученый
Кэхилл Г.Ф. Младший. Метаболизм топлива при голодании. Анну Рев Нутр. 2006; 26: 1–22.
PubMed Статья CAS Google ученый
Ким Д. Я., Ро Дж. М.. Кетогенная диета и эпилепсия. Curr Opin Clin Nutr Metab Care. 2008; 11: 113–20.
PubMed Статья CAS Google ученый
Перес-Эскуредо Дж., Ван Хи В.Ф., Сбоарина М., Фальсес Дж., Пайен В.Л., Пеллерин Л., Сонво П. Переносчики монокарбоксилатов в головном мозге и при раке . Biochim Biophys Acta 2016, 1863 : 2481–2497.
Halestrap AP. Семейство монокарбоксилатных переносчиков — структура и функциональные характеристики.МСБМБ Жизнь. 2012; 64: 1–9.
PubMed Статья CAS Google ученый
Broer S, Broer A, Schneider HP, Stegen C, Halestrap AP, Deitmer JW. Характеристика высокоаффинного монокарбоксилатного переносчика MCT2 в ооцитах Xenopus laevis. Biochem J. 1999; 341 (Pt 3): 529–35.
PubMed PubMed Central CAS Google ученый
Dimmer KS, Friedrich B, Lang F, Deitmer JW, Broer S.Транспортер монокарбоксилата с низким сродством MCT4 адаптирован к экспорту лактата в высокогликолитические клетки. Biochem J. 2000; 350 (Pt 1): 219–27.
PubMed PubMed Central Статья CAS Google ученый
Ньюман Дж. К., Вердин Э. Бета-гидроксибутират: гораздо больше, чем метаболит. Диабет Res Clin Pract. 2014; 106: 173–81.
PubMed PubMed Central Статья CAS Google ученый
Hashim SA, VanItallie TB. Кетоновая терапия для тела: от кетогенной диеты до перорального приема кетонового эфира. J Lipid Res. 2014; 55: 1818–26.
PubMed PubMed Central Статья CAS Google ученый
Veech RL, Chance B, Kashiwaya Y, Lardy HA, Cahill GF Jr. Кетоновые тела, потенциальные терапевтические применения. МСБМБ Жизнь. 2001; 51: 241–7.
PubMed Статья CAS Google ученый
Видали С., Аминзаде С., Ламберт Б., Резерфорд Т., Сперл В., Кофлер Б., Файхтингер Р.Г. Митохондрии: кетогенная диета — терапия, основанная на метаболизме. Int J Biochem Cell Biol. 2015; 63: 55–9.
PubMed Статья CAS Google ученый
Кэхилл Г.Ф. младший, Вич Р.Л. Кетокислоты? Хорошее лекарство? Trans Am Clin Climatol Assoc. 2003. 114: 149–61. обсуждение 162-143
PubMed PubMed Central Google ученый
Ваупель П., Келлехер Д.К., Хоккель М. Кислородный статус злокачественных опухолей: патогенез гипоксии и значение для терапии опухолей. Семин Онкол. 2001; 28: 29–35.
PubMed Статья CAS Google ученый
Martinez-Outschoorn UE, Prisco M, Ertel A, Tsirigos A, Lin Z, Pavlides S, Wang C, Flomenberg N, Knudsen ES, Howell A, et al. Кетоны и лактат увеличивают «стволовость» раковых клеток, приводя к рецидивам, метастазам и плохим клиническим исходам при раке груди: достижение персонализированной медицины с помощью метаболо-геномики.Клеточный цикл. 2011; 10: 1271–86.
PubMed PubMed Central Статья CAS Google ученый
Бонучелли Г., Циригос А., Уитакер-Менезес Д., Павлидес С., Пестелл Р. Г., Кьяварина Б., Франк П. Г., Фломенберг Н., Хауэлл А., Мартинес-Оутсхорн У. Э. и др. Кетоны и лактат «подпитывают» рост опухоли и метастазирование: доказательства того, что эпителиальные раковые клетки используют окислительный митохондриальный метаболизм. Клеточный цикл. 2010; 9: 3506–14.
PubMed PubMed Central Статья CAS Google ученый
Martuscello RT, Vedam-Mai V, McCarthy DJ, Schmoll ME, Jundi MA, Louviere CD, Griffith BG, Skinner CL, Suslov O, Deleyrolle LP, Reynolds BA. Диета с добавлением жиров и низким содержанием углеводов для лечения глиобластомы. Clin Cancer Res. 2016; 22: 2482–95.
PubMed Статья CAS Google ученый
Шукла С.К., Гебрегиворгис Т., Пурохит В., Чайка Н.В., Гунда В., Радхакришнан П., Мехла К., Пипинос II, Пауэрс Р., Ю Ф, Сингх П.К.Перепрограммирование метаболизма, вызванное кетоновыми телами, уменьшает кахексию рака поджелудочной железы. Cancer Metab. 2014; 2: 18.
PubMed PubMed Central Статья Google ученый
Маги Б.А., Потезный Н., Рофе А.М., Коньерс Р.А. Подавление кетоновыми телами роста злокачественных клеток. Aust J Exp Biol Med Sci. 1979; 57: 529–39.
PubMed Статья CAS Google ученый
Poff AM, Ari C, Arnold P, Seyfried TN, D’Agostino DP. Добавление кетонов снижает жизнеспособность опухолевых клеток и продлевает выживаемость мышей с метастатическим раком. Int J Cancer. 2014; 135: 1711–20.
PubMed PubMed Central Статья CAS Google ученый
Скиннер Р., Трухильо А., Ма Х, Байерле Е.А. Кетоновые тела подавляют жизнеспособность клеток нейробластомы человека. J Pediatr Surg. 2009; 44: 212–6. обсуждение 216
PubMed Статья Google ученый
Klement RJ, Champ CE, Otto C, Kammerer U. Противоопухолевые эффекты кетогенных диет у мышей: метаанализ. PLoS One. 2016; 11: e0155050.
PubMed PubMed Central Статья CAS Google ученый
Mavropoulos JC, Buschemeyer WC 3rd, Tewari AK, Rokhfeld D, Pollak M, Zhao Y, Febbo PG, Cohen P, Hwang D, Devi G, et al. Влияние изменения содержания углеводов и жиров в рационе на выживаемость в модели ксенотрансплантата рака предстательной железы LNCaP на мышах.Рак Prev Res (Phila). 2009; 2: 557–65.
Артикул CAS Google ученый
Небелинг Л.С., Миральди Ф., Шурин С.Б., Лернер Э. Влияние кетогенной диеты на метаболизм опухоли и нутритивный статус у педиатрических онкологических пациентов: два клинических случая. J Am Coll Nutr. 1995; 14: 202–8.
PubMed Статья CAS Google ученый
Отто С., Кеммерер Ю., Иллерт Б., Мюлинг Б., Пфетцер Н., Виттиг Р., Фелькер Н.Ю., Тиде А., Кой Дж.Ф.Рост клеток рака желудка человека у голых мышей задерживается кетогенной диетой с добавлением омега-3 жирных кислот и триглицеридов со средней длиной цепи. BMC Рак. 2008; 8: 122.
PubMed PubMed Central Статья CAS Google ученый
Poff AM, Ari C, Seyfried TN, D’Agostino DP. Кетогенная диета и гипербарическая кислородная терапия продлевают выживаемость мышей с системным метастатическим раком. PLoS One. 2013; 8: e65522.
PubMed PubMed Central Статья CAS Google ученый
Сейфрид Т.Н., Сандерсон Т.М., Эль-Аббади М.М., Макгоуэн Р., Мукерджи П. Роль глюкозы и кетоновых тел в метаболическом контроле экспериментального рака мозга. Br J Рак. 2003. 89: 1375–82.
PubMed PubMed Central Статья CAS Google ученый
Zuccoli G, Marcello N, Pisanello A, Servadei F, Vaccaro S, Mukherjee P, Seyfried TN.Метаболическое управление мультиформной глиобластомой с использованием стандартной терапии вместе с ограниченной кетогенной диетой: отчет о клиническом случае. Нутр Метаб (Лондон). 2010; 7:33.
Артикул CAS Google ученый
Csaba Toth ZC. Остановлено прогрессирование рака мягкого неба у пациента, получавшего только палеолитическую кетогенную диету: наблюдение через 20 месяцев. Am J Med Case Rep. 2016; 4: 288–92.
Google ученый
Родригес Л.М., Урибе-Льюис С., Мадху Б., Честное Диджей, Стаббс М., Гриффитс-младший. Действие бета-гидроксибутирата на рост, метаболизм и общее ацетилирование гистона h4 спонтанных опухолей молочной железы мышей: свидетельство парадокса бета-гидроксибутирата. Cancer Metab. 2017; 5: 4.
PubMed PubMed Central Статья Google ученый
Клемент Р.Дж. Влияние кетогенной терапии на 5 R радиобиологии. Int J Radiat Biol.2017: 1–13.
Паоли А., Бьянко А., Гримальди К.А., Лоди А., Боско Г. Долгосрочное успешное снижение веса с помощью сочетания двухфазной кетогенной средиземноморской диеты и поддерживающего протокола средиземноморской диеты. Питательные вещества. 2013; 5: 5205–17.
PubMed PubMed Central Статья Google ученый
Paoli A, Cenci L, Grimaldi KA. Влияние кетогенной средиземноморской диеты с фитоэкстрактами и низкоуглеводными / высокобелковыми блюдами на вес, факторы риска сердечно-сосудистых заболеваний, состав тела и соблюдение диеты у сотрудников совета Италии.Нутр Дж. 2011; 10: 112.
PubMed PubMed Central Статья CAS Google ученый
Zajac A, Poprzecki S, Maszczyk A, Czuba M, Michalczyk M, Zydek G. Влияние кетогенной диеты на метаболизм при упражнениях и физическую работоспособность у велосипедистов-внедорожников. Питательные вещества. 2014; 6: 2493–508.
PubMed PubMed Central Статья CAS Google ученый
Урбайн П., Стром Л., Моравски Л., Верле А., Дейберт П., Берц Х. Влияние 6-недельной кетогенной диеты без ограничения энергии на физическую форму, состав тела и биохимические параметры у здоровых взрослых. Нутр Метаб (Лондон). 2017; 14:17.
Артикул CAS Google ученый
Дашти Х.М., Мэтью Т.С., Хусейн Т., Асфар С.К., Бехбахани А., Хуршид М.А., Аль-Сайер Х.М., Бо-Аббас Ю.Й., Аль-Заид Н.С.. Долгосрочные эффекты кетогенной диеты у пациентов с ожирением.Exp Clin Cardiol. 2004; 9: 200–5.
PubMed PubMed Central CAS Google ученый
Перес-Гисадо Дж., Муньос-Серрано А., Алонсо-Морага А. Испанская кетогенная средиземноморская диета: здоровая сердечно-сосудистая диета для похудения. Нутр Дж. 2008; 7:30.
PubMed PubMed Central Статья CAS Google ученый
Ваупель П., Шленгер К., Кнуп С., Хокель М.Оксигенация опухолей человека: оценка распределения кислорода в тканях при раке молочной железы с помощью компьютерных измерений напряжения O2. Cancer Res. 1991; 51: 3316–22.
PubMed CAS Google ученый
Холлидей Д.Л., Спирс В. Выбор правильной клеточной линии для исследования рака груди. Рак молочной железы Res. 2011; 13: 215.
PubMed PubMed Central Статья Google ученый
Неве Р.М., Чин К., Фридлянд Дж., Йе Дж., Баенер Флорида, Февр Т., Кларк Л., Баяни Н., Коппе Дж. П., Тонг Ф. и др. Коллекция клеточных линий рака молочной железы для изучения функционально различных подтипов рака. Раковая клетка. 2006; 10: 515–27.
PubMed PubMed Central Статья CAS Google ученый
Subik K, Lee JF, Baxter L, Strzepek T, Costello D, Crowley P, Xing L, Hung MC, Bonfiglio T, Hicks DG, Tang P. Паттерны экспрессии ER, PR, HER2, CK5 / 6, EGFR, Ki-67 и AR с помощью иммуногистохимического анализа клеточных линий рака молочной железы.Рак груди (Окл). 2010; 4: 35–41.
PubMed Central Google ученый
Дай X, Ченг Х., Бай З., Ли Дж. Классификация линий клеток рака молочной железы и ее значение для определения подтипов опухоли молочной железы. J Рак. 2017; 8: 3131–41.
PubMed PubMed Central Статья Google ученый
Баэнке Ф., Дюбуи С., Браулт С., Вайгельт Б., Данкуорт Б., Гриффитс Б., Цзян М., Маккей А., Сондерс Б., Спенсер-Дене Б. и др.Функциональный скрининг определяет MCT4 как ключевой регулятор метаболизма и выживаемости клеток рака груди. J Pathol. 2015; 237: 152–65.
PubMed Статья CAS Google ученый
Smith SE, Mellor P, Ward AK, Kendall S, McDonald M, Vizeacoumar FS, Vizeacoumar FJ, Napper S, Anderson DH. Молекулярная характеристика клеточных линий рака молочной железы с помощью нескольких атомных подходов. Рак молочной железы Res. 2017; 19:65.
PubMed PubMed Central Статья Google ученый
Каталог соматических мутаций при раке. (https://cancer.sanger.ac.uk/cosmic).
Kammerer U, Gires O, Pfetzer N, Wiegering A, Klement RJ, Otto C. Экспрессия TKTL1 в линиях злокачественных и доброкачественных клеток человека. BMC Рак. 2015; 15: 2.
PubMed PubMed Central Статья CAS Google ученый
Pfaffl MW. Новая математическая модель для относительной количественной оценки в RT-PCR в реальном времени. Nucleic Acids Res.2001; 29: e45.
PubMed PubMed Central Статья CAS Google ученый
Брэдфорд MM. Быстрый и чувствительный метод количественного определения количества белка в микрограммах, использующий принцип связывания белок-краситель. Анальная биохимия. 1976; 72: 248–54.
PubMed Статья CAS Google ученый
Чанг Дж., Ли А., Ли Дж., Лим В., Сун Ш., Мун Б.Корреляция между молекулярным подтипом рака груди и анализом химиочувствительности на основе аденозинтрифосфата in vitro. J Korean Surg Soc. 2013; 84: 313–20.
PubMed PubMed Central Статья CAS Google ученый
Lin Y, Lv F, Liu F, Guo X, Fan Y, Gu F, Gu J, Fu L. Высокая экспрессия пируваткиназы M2 связана с химиочувствительностью к эпирубицину и 5-фторурацилу при раке груди. J Рак.2015; 6: 1130–9.
PubMed PubMed Central Статья Google ученый
Kurbacher CM, Mallmann P, Kurbacher JA, Hübner H, Krebs D. Chemosensibilitätstestungen in der gynäkologischen Onkologie: Erfahrungen mit einem ATP-Biolumineszenzassay. Geburtshilfe Frauenheilkd. 1996; 56: 70–8.
PubMed Статья CAS Google ученый
Гве Ан С., А Ли С., Ву Ли Х, Мин Ли Х, Чжон Дж.Тест на химиотерапию in vitro, основанный на внутренних подтипах рака груди. Jpn J Clin Oncol. 2014; 44: 624–31.
PubMed Статья Google ученый
Xia S, Lin R, Jin L, Zhao L, Kang HB, Pan Y, Liu S, Qian G, Qian Z, Konstantakou E, et al. Предотвращение кетогенеза, подпитываемого диетическими жирами, снижает рост опухоли BRAF V600E. Cell Metab. 2017; 25: 358–73.
PubMed PubMed Central Статья CAS Google ученый
NCT01865162 N, NCT02983942, NCT02939378, NCT03160599, NCT01754350, NCT02516501, NCT03075514, NCT01535911 (www.clinicaltrials.gov).
Бранка Дж., Пачини С., Руджеро М. Эффекты предоперационного приема витамина D и кетогенной диеты у пациента с рецидивирующим раком груди. Anticancer Res. 2015; 35: 5525–32.
PubMed CAS Google ученый
Янсен Н., Валах Х. Развитие опухолей при кетогенной диете в сочетании с новым онкомаркером TKTL1: серия случаев в общей практике.Oncol Lett. 2016; 11: 584–92.
PubMed Статья CAS Google ученый
Schwalb MTM, Hines S, Reinwald H, Ruggiero M. Клиническое наблюдение нового дополнительного иммунотерапевтического подхода, основанного на кетогенной диете, хондроитинсульфате, витамине D 3, олеиновой кислоте и ферментированном молоке и продукте из молозива. Am J Immunol. 2016; 12: 91–8.
Артикул CAS Google ученый
Iyikesici MS, Slocum AK, Slocum A, Berkarda FB, Kalamian M, Seyfried TN. Эффективность метаболически поддерживаемой химиотерапии в сочетании с кетогенной диетой, гипертермией и гипербарической кислородной терапией при тройном отрицательном раке груди IV стадии. Cureus. 2017; 9: e1445.
PubMed PubMed Central Google ученый
Martinez-Outschoorn UE, Lin Z, Whitaker-Menezes D, Howell A, Lisanti MP, Sotgia F. Кетоновые тельца и двухкомпонентный метаболизм опухоли: производство стромальных кетонов подпитывает митохондриальный биогенез в эпителиальных раковых клетках.Клеточный цикл. 2012; 11: 3956–63.
PubMed PubMed Central Статья CAS Google ученый
Martinez-Outschoorn UE, Lin Z, Whitaker-Menezes D, Howell A, Sotgia F, Lisanti MP. Использование кетонового тела способствует росту опухоли и метастазированию. Клеточный цикл. 2012; 11: 3964–71.
PubMed PubMed Central Статья CAS Google ученый
Pelicano H, Zhang W, Liu J, Hammoudi N, Dai J, Xu RH, Pusztai L, Huang P.Дисфункция митохондрий в некоторых линиях клеток трижды отрицательного рака молочной железы: роль пути mTOR и терапевтический потенциал. Рак молочной железы Res. 2014; 16: 434.
PubMed PubMed Central Статья CAS Google ученый
Лэннинг, Нью-Джерси, Касл Дж. П., Сингх С. Дж., Леон А. Н., Товар Е. А., Сангера А., МакКейган Дж. П., Филипп Ф. В., Гравил С. Р.. Метаболическое профилирование трижды отрицательных клеток рака груди выявляет метаболическую уязвимость.Cancer Metab. 2017; 5: 6.
PubMed PubMed Central Статья Google ученый
Чаттерджи Р. Клеточная биология. Случаи ошибочной идентификации. Наука. 2007; 315: 928–31.
PubMed Статья CAS Google ученый
Чаттерджи Р. Клеточная биология. Когда 60 строк не складываются. Наука. 2007; 315: 929.
PubMed Статья Google ученый
Dubuis S, Baenke F, Scherbichler N, Alexander LT, Schulze A, Zamboni N. Метаботипы клеточных линий рака молочной железы, выявленные нецелевой метаболомикой. Metab Eng. 2017; 43: 173–86.
PubMed Статья CAS Google ученый
Mookerjee SA, Brand MD. Измерение и анализ продукции внеклеточной кислоты для определения скорости гликолиза. J Vis Exp. 2015: e53464.
Mookerjee SA, Goncalves RL, Gerencser AA, Nicholls DG, Brand MD.Вклад дыхания и гликолиза в выработку внеклеточной кислоты. Biochim Biophys Acta. 2015; 1847: 171–81.
PubMed Статья CAS Google ученый
Wise DR, Thompson CB. Глютаминовая зависимость: новая терапевтическая мишень при раке. Trends Biochem Sci. 2010. 35: 427–33.
PubMed PubMed Central Статья CAS Google ученый
Лампа М., Арлт Х, Хе Т, Оспина Б., Ривз Дж., Чжан Б., Мурти Дж., Дэн Дж., Барберис К., Хоффманн Д. и др. Глутаминаза необходима для роста трижды отрицательных клеток рака молочной железы с нарушенным путем метаболизма глутамина, и ее подавление синергетично с ингибированием mTOR. PLoS One. 2017; 12: e0185092.
PubMed PubMed Central Статья CAS Google ученый
ДеБерардини Р.Дж., Манкузо А., Дайхин Е., Ниссим И., Юдкофф М., Верли С., Томпсон ЦБ.Помимо аэробного гликолиза: трансформированные клетки могут участвовать в метаболизме глутамина, который превышает потребность в синтезе белков и нуклеотидов. Proc Natl Acad Sci U S. A. 2007; 104: 19345–50.
PubMed PubMed Central Статья Google ученый
Fine EJ, Miller A, Quadros EV, Sequeira JM, Feinman RD. Ацетоацетат снижает рост и концентрацию АТФ в линиях раковых клеток, которые чрезмерно экспрессируют разобщающий белок 2.Cancer Cell Int. 2009; 9: 14.
PubMed PubMed Central Статья CAS Google ученый
Vozza A, Parisi G, De Leonardis F, Lasorsa FM, Castegna A, Amorese D, Marmo R, Calcagnile VM, Palmieri L, Ricquier D, et al. UCP2 транспортирует метаболиты C4 из митохондрий, регулируя окисление глюкозы и глутамина. Proc Natl Acad Sci U S. A. 2014; 111: 960–5.
PubMed PubMed Central Статья CAS Google ученый
Чанг HT, Олсон Л.К., Шварц К.А. Профили экспрессии кетолитических и гликолитических ферментов в злокачественных глиомах: значение для терапии кетогенной диетой. Нутр Метаб (Лондон). 2013; 10: 47.
Артикул CAS Google ученый
Мацумото К., Фудзивара Ю., Нагай Р., Йошида М., Уэда С. Экспрессия двух изоферментов ацил-кофермента А: холестерин-ацилтрансферазы-1 и -2 в почечно-клеточной карциноме светлоклеточного типа. Int J Urol. 2008; 15: 166–70.
PubMed Статья CAS Google ученый
Maurer GD, Brucker DP, Bahr O, Harter PN, Hattingen E, Walenta S, Mueller-Klieser W., Steinbach JP, Rieger J. Дифференциальное использование кетоновых тел нейронами и клеточными линиями глиомы: обоснование для кетогенная диета как экспериментальная терапия глиомы. BMC Рак. 2011; 11: 315.
PubMed PubMed Central Статья CAS Google ученый
Патель М.С., Рассел Дж. Дж., Гершман Х. Метаболизм кетоновых тел в клетках глиомы и нейробластомы. Proc Natl Acad Sci U S. A. 1981; 78: 7214–8.
PubMed PubMed Central Статья CAS Google ученый
Sawai M, Yashiro M, Nishiguchi Y, Ohira M, Hirakawa K. Тормозящие рост эффекты кетонового тела, моноацетоацетина, на клетки рака желудка человека с сукцинил-КоА: 3-оксоацид КоА-трансферазой (SCOT ) дефицит.Anticancer Res. 2004; 24: 2213–7.
PubMed CAS Google ученый
Анталис С.Дж., Арнольд Т., Расул Т., Ли Б., Бухман К.К., Сиддики Р.А. Высокая экспрессия ACAT1 в базальных клетках рака молочной железы, отрицательных по рецепторам эстрогена, связана с пролиферацией ЛПНП. Лечение рака груди Res. 2010; 122: 661–70.
PubMed Статья CAS Google ученый
Бентон С.Р., Кэмпбелл С.Е., Тонучи М., Хатта Х., Бонен А. Транспортеры монокарбоксилатов в субсарколеммальных и интермиофибриллярных митохондриях. Biochem Biophys Res Commun. 2004. 323: 249–53.
PubMed Статья CAS Google ученый
Пинейро С., Рейс Р.М., Рикардо С., Лонгатто-Филхо А., Шмитт Ф., Балтазар Ф. Экспрессия транспортеров монокарбоксилата 1, 2 и 4 в опухолях человека и их связь с CD147 и CD44.J Biomed Biotechnol. 2010; 2010: 427694.
PubMed PubMed Central Статья CAS Google ученый
Valenca I, Pertega-Gomes N, Vizcaino JR, Henrique RM, Lopes C, Baltazar F, Ribeiro D. Локализация MCT2 в пероксисомах связана со злокачественной трансформацией рака простаты. J Cell Mol Med. 2015; 19: 723–33.
PubMed PubMed Central Статья CAS Google ученый
Cheng C, Edin NF, Lauritzen KH, Aspmodal I, Christoffersen S, Jian L, Rasmussen LJ, Pettersen EO, Xiaoqun G, Bergersen LH. Изменение плотности переносчиков монокарбоксилатов при гипоксии в опухолевых клетках головного мозга и молочной железы. Сотовый Онкол (Дордр). 2012; 35: 217–27.
Артикул CAS Google ученый
Эйлерцен М., Андерсен С., Аль-Саад С., Киселев Ю., Доннем Т., Стенволд Х., Петтерсен И., Аль-Шибли К., Ричардсен Е., Бусунд Л.Т., Бремнес Р.М.Транспортеры монокарбоксилатов 1–4 при НМРЛ: MCT1 является независимым прогностическим маркером выживаемости. PLoS One. 2014; 9: e105038.
PubMed PubMed Central Статья CAS Google ученый
Македа М.Л., Роджерс С., Бест Джей. Молекулярная и клеточная регуляция белков-переносчиков глюкозы (GLUT) при раке. J. Cell Physiol. 2005; 202: 654–62.
PubMed Статья CAS Google ученый
Tardif A, Julien N, Pelletier A, Thibault G, Srivastava AK, Chiasson JL, Coderre L. Хроническое воздействие бета-гидроксибутирата ухудшает действие инсулина в первичных культурах кардиомиоцитов взрослых. Am J Physiol Endocrinol Metab. 2001; 281: E1205–12.
PubMed Статья CAS Google ученый
Bria E, Nistico C, Cuppone F, Carlini P, Ciccarese M, Milella M, Natoli G, Terzoli E, Cognetti F, Giannarelli D. Польза таксанов в качестве адъювантной химиотерапии при раннем раке груди: объединенный анализ 15 500 пациентов.Рак. 2006; 106: 2337–44.
PubMed Статья CAS Google ученый
Kern P, Kalisch A, von Minckwitz G, Putter C, Kolberg HC, Pott D, Kurbacher C., Rezai M, Kimmig R. Неоадъювант, безантрациклиновая химиотерапия карбоплатином и доцетакселом при трех отрицательных, ранних -этапный рак молочной железы: многоцентровый анализ показателей полного патологического ответа и выживаемости. J Chemother. 2016; 28: 210–7.
PubMed Статья CAS Google ученый
Shao N, Wang S, Yao C, Xu X, Zhang Y, Zhang Y, Lin Y. Последовательные и одновременные антрациклины и таксаны в качестве адъювантной химиотерапии раннего рака груди: метаанализ рандомизированных контрольных исследований III фазы. Грудь. 2012; 21: 389–93.
PubMed Статья Google ученый
Шарма П., Лопес-Тарруэлла С., Гарсия-Саенс Дж. А., Вард С., Коннор С. С., Гомес Х. Л., Прат А., Морено Ф., Херес-Гиларранс И., Барнадас А. и др. Эффективность неоадъювантного карбоплатина плюс доцетаксела при тройном отрицательном раке молочной железы: комбинированный анализ двух когорт.Clin Cancer Res. 2017; 23: 649–57.
PubMed Статья CAS Google ученый
Тернер Н., Биганзоли Л., Ди Лео А. Постоянное значение адъювантных антрациклинов в качестве лечения раннего рака груди. Ланцет Онкол. 2015; 16: e362–9.
PubMed Статья Google ученый
Аллен Б.Г., Бхатия С.К., Буатти Дж. М., Брандт К. Э., Линдхольм К. Э., Баттон А. М., Шведа Л. И., Смит Б. Дж., Шпиц Д. Р., Фат М. А..Кетогенные диеты усиливают окислительный стресс и реакцию на радиохимиотерапию ксенотрансплантатов рака легких. Clin Cancer Res. 2013; 19: 3905–13.
PubMed PubMed Central Статья CAS Google ученый
Клемент Р.Дж., Суини РА. Влияние кетогенной диеты во время лучевой терапии на состав тела: I. Первоначальный клинический опыт с шестью проспективно изученными пациентами. BMC Res Notes. 2016; 9: 143.
PubMed PubMed Central Статья CAS Google ученый
Абдельвахаб М.Г., Фентон К.Э., Преул М.С., Ро Дж. М., Линч А., Стаффорд П., Шек А. С.. Кетогенная диета является эффективным дополнением к лучевой терапии для лечения злокачественной глиомы. PLoS One. 2012; 7: e36197.
PubMed PubMed Central Статья Google ученый
Клемент Р.Дж. Благоприятные эффекты кетогенных диет для больных раком: реалистичный обзор с акцентом на доказательствах и подтверждениях. Med Oncol. 2017; 34: 132.
PubMed Статья Google ученый
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Сравнение ацетона для дыхания и бета-гидроксибутирата крови — полный текст
Устройство LEVL (Medamonitor, Сиэтл, Вашингтон) предоставляет пользователям метод оценки собственной скорости метаболизма жиров. Устройство LEVL измеряет концентрацию ацетона в выдыхаемом воздухе, которая, как показала научная литература, коррелирует со скоростью метаболизма жиров.
Обычно организм использует глюкозу для удовлетворения своих потребностей в метаболической энергии. При необходимости организм может перейти с глюкозы на жир.Этот сдвиг может вызвать множество различных сценариев. Упражнения могут истощить доступные запасы углеводов, заставляя организм использовать жиры для производства энергии. Изменения в диете, вызывающие увеличение потребления жиров и уменьшение потребления углеводов, изменят метаболизм, чтобы эффективно использовать изменения в питательных веществах. Во всех этих сценариях организм сокращает использование углеводов и, таким образом, переходит к использованию жиров для производства энергии.
Когда организм использует жиры в качестве энергетического субстрата, некоторые из этих молекул жира превращаются печенью в ацетоацетат, кетоновое тело.Благодаря ферментативному действию ацетоацетат (AcAc) может быть преобразован в бета-гидроксибутират (BOHB). Тот же фермент может генерировать AcAc из BOHB. Кроме того, ацетоацетат может самопроизвольно превращаться в ацетон, который из-за своего небольшого размера и высокой растворимости в воде может легко появляться при дыхании.
В настоящее время измерение BOHB в крови является золотым стандартом для оценки концентрации кетоновых тел, также известной как кетоз. Измерение BOHB требует инвазивной пункции пальца для получения крови и дорогостоящего (~ 5 долларов США за тест) анализа.Поскольку ацетон (BrAce) является сестринским кетоновым телом по отношению к BOHB, ацетон для дыхания может использоваться для оценки кетоза и замены измерения BOHB. Данные в научной литературе показали, что BrAce коррелирует с BOHB. Определение концентраций BOHB из измерения BrAce с использованием LEVL менее инвазивно и потенциально менее затратно.