Скелетная мышечная ткань — Википедия
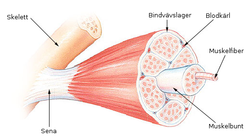 Схема скелетной мышцы в разрезе.
Схема скелетной мышцы в разрезе.  Строение скелетной мышцы
Строение скелетной мышцыСкелетная (поперечнополосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов: один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 70—75 % из воды.
Источником развития скелетной мускулатуры являются клетки миотомов — миобласты. Часть из них дифференцируется в местах образования так называемых аутохтонных мышц. Прочие же мигрируют из миотомов в мезенхиму; при этом они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникает 2 клеточные линии. Клетки первой сливаются, образуя симпласты — мышечные трубки (миотубы). Клетки второй группы остаются самостоятельными и дифференцируются в миосателлиты (миосателлитоциты).
В первой группе происходит дифференцировка специфических органелл миофибрилл, постепенно они занимают большую часть просвета миотубы, оттесняя ядра клеток к периферии.
Клетки второй группы остаются самостоятельными и располагаются на поверхности мышечных трубок.
Структурной единицей мышечной ткани является мышечное волокно. Оно состоит из миосимпласта и миосателлитоцитов (клеток-сателлитов), покрытых общей базальной мембраной. Длина мышечного волокна может достигать нескольких сантиметров при толщине в 50—100 микрометров.
Скелетные мышцы прикреплены к костям или друг к другу крепкими, гибкими сухожилиями.
Строение миосимпласта[править | править код]
Миосимпласт представляет собой совокупность слившихся клеток. В нем имеется большое количество ядер, расположенных по периферии мышечного волокна (их число может достигать десятков тысяч). Как и ядра, на периферии симпласта расположены другие органеллы, необходимые для работы мышечной клетки — эндоплазматическая сеть (саркоплазматический ретикулюм), митохондрии и др. Центральную часть симпласта занимают миофибриллы. Структурная единица миофибриллы — саркомер. Он состоит из молекул актина и миозина, именно их взаимодействие и обеспечивает изменение длины мышечного волокна и как следствие сокращение мышцы. В состав саркомера входят также многие вспомогательные белки — титин, тропонин, тропомиозин и др
Строение миосателлитов[править | править код]
Миосателлиты — одноядерные клетки, прилежащие к поверхности миосимпласта. Эти клетки отличаются низкой дифференцировкой и служат взрослыми стволовыми клетками мышечной ткани. В случае повреждения волокна или длительном увеличении нагрузки клетки начинают делиться, обеспечивая рост миосимпласта.
Функциональной единицей скелетной мышцы является моторная единица (МЕ). МЕ включает в себя группу мышечных волокон и иннервирующий их мотонейрон. Число мышечных волокон, входящих в состав одной МЕ, варьирует в разных мышцах. Например, там, где требуется тонкий контроль движений (в пальцах или в мышцах глаза), моторные единицы небольшие, они содержат не более 30 волокон. А в икроножной мышце, где тонкий контроль не нужен, в МЕ насчитывается более 1000 мышечных волокон.
Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на медленные (slow (S-МЕ)) и быстрые (fast (F-МЕ)). А F-МЕ в свою очередь делят по устойчивости к утомлению на устойчивые к утомлению (fast-fatigue-resistant (FR-МЕ)) и быстроутомляемые (fast-fatigable (FF-МЕ)).
Соответствующим образом подразделяют мотонейроны, иннервирующие данные МЕ. Существуют S-мотонейроны (S-МН), FF-мотонейроны (F-МН) и FR-мотонейроны (FR-МН).
S-МЕ характеризуются высоким содержанием белка миоглобина, который способен связывать кислород (О2). Мышцы, преимущественно состоящие из МЕ этого типа, за их темно-красный цвет называются красными. Красные мышцы выполняют функцию поддержания позы человека. Предельное утомление таких мышц наступает очень медленно, а восстановление функций происходит наоборот, очень быстро. Такая способность обуславливается наличием миоглобина и большого числа митохондрий. МЕ красных мышц, как правило, содержат большое количество мышечных волокон.
FR-МЕ составляют мышцы, способные выполнять быстрые сокращения без заметного утомления. Волокна FR-ME содержат большое количество митохондрий и способны образовывать АТФ путём окислительного фосфорилирования. Как правило, число волокон в FR-ME меньше, чем в S-ME.
Волокна FF-ME характеризуются меньшим содержанием митохондрий, чем в FR-ME, а также тем, что АТФ в них образуется за счет гликолиза. В них отсутствует миоглобин, поэтому мышцы, состоящие из МЕ этого типа, называют белыми. Белые мышцы развивают сильное и быстрое сокращение, но довольно быстро утомляются.
Функция[править | править код]
Скелетные мышцы обеспечивают возможность выполнения произвольных движений. Сокращающаяся мышца воздействует на кости или кожу, к которым она прикрепляется. При этом один из пунктов прикрепления остаётся неподвижным — так называемая
- Ю.И. Афанасьев, Н.А. Юрина, Е.Ф. Котовский. Гистология. — 5-е изд., перераб. и доп.. — Москва: Медицина, 2002. — 744 с. — ISBN 5-225-04523-5.
- [1]- Механизмы развития мышечной ткани (англ.)
Типы мышечных волокон
Если мы хотим заботиться о своем здоровье, хотим его сохранить на долгие годы, то нам необходимо заниматься физическими упражнениями. Но заниматься нужно правильно, добиваясь положительного эффекта. А сделать это, ничего не зная о том, что такое мышечные волокна и какие бывают типы этих самых волокон невозможно.
Чтобы ликвидировать безграмотность в области физической культуры, предлагаю ознакомиться с очень важной и крайне нужно информацией. Постараюсь всё передать максимально понятно и доходчиво. И хотя, наверняка, покажется сложным пробираться через большое количество новых терминов, это необходимо для плодотворной жизни и здоровья.
Что такое мышечное волокно
Представьте себе любую мышцу. Если не можете представить, просто посмотрите на какую-то часть тела.
Мы почти полностью покрыты мышцами. Некоторые из них мощные и большие, как мышцы ног и ягодиц. Другие – более мелкие, например, мышцы рук. Но каждая мышца состоит из определенного количество мышечных волокон.
Образно это можно представить следующим образом. Возьмите пачку спагетти, это и будет мышца. А каждая спагеттинка в пачке – это мышечное волокно. Вот так просто.
При этом, чем больше мышца от природы, тем больше в ней мышечных волокон.
Типы мышечных волокон
Мышечные волокна делятся по 3-м признакам: скорости сокращения, цвету и способу получению энергии.
По скорости сокращения мышечные волокна делятся на:
1. Быстрые мышечные волокна;
2. Медленные мышечные волокна.
Быстрота мышечного сокращения зависит от вырабатываемого в волокне фермента АТФаза, который воздействует на молекулу АТФ либо сравнительно быстро, либо сравнительно медленно. Это наследуемый признак, и особого значения для жизни не имеет. Но в спорте это один из основных элементов на этапе отбора.
По цвету мышечные волокна делятся на:
1. Красные;
2. Белые;
3. Розовые.
Цвет мышечному волокну придает миоглобин – белок, ответственный за доставку кислорода внутрь волокна.
Там, где миоглобина много, мышечное волокно окрашивается в красный цвет.
Там, где миоглобина меньше, но он есть, цвет становится розовым.
Там, где миоглобина вообще почти нет, мышечное волокно остается белым.
Цвет мышечных волокон более важен для жизни, чем скорость сокращения.
Но наиболее важным остается деление мышечных волокон по признаку получения энергии. С этой стороны мышечные волокна делятся на:
1. Окислительные;
2. Гликолитические;
3. Промежуточные.
В окислительных мышечных волокнах энергию для сокращения (произведения работы) получают с помощью окисления жиров (липолиз) или окисления глюкозы (аэробный гликолиз). Окисление подразумевает взаимодействие с кислородом, который доставляется внутрь при помощи уже упомянутого миоглобина. В идеале, окислительное мышечное волокно всегда красного цвета.
Получение энергии при взаимодействии с кислородом возможно благодаря обильному распространению в окислительных мышечных волокнах т.н. митохондрий – «энергетических станций» мышечных клеток. В них происходит образование энергии.
В гликолитических мышечных волокнах митохондрии почти отсутствуют. Поэтому энергию для сокращения такие волокна получают при помощи расщепления глюкозы путем анаэробного гликолиза – без кислорода. Такие мышечные волокна всегда белого цвета по причине почти полного отсутствия миоглобина.
Промежуточные мышечные волокна имеют некоторое количество митохондрий. Их больше, чем в гликолитических, но меньше, чем в окислительных волокнах. Поэтому таким мышечным волокнам дано название промежуточных. По цвету они розовые, т.е. миоглобин есть, не его немного.
Почему важны мышечные волокна
Когда мышца сокращается, то всегда делает это при помощи сокращения каждого мышечного волокна. Только при небольшой нагрузке начинают сокращаться сначала окислительные мышечные волокна. Потом подключаются промежуточные. А когда работа требует уже достаточных усилий, в работу включаются и гликолитические. Это т.н. правило Ханнемана – правило рекрутирования мышечных волокон. Рекрутирование всегда идет по нарастающей, от окислительных к гликолитическим.
А теперь самое важное, что нужно знать
Окислительные мышечные волокна способны производить работу небольшой интенсивности (приложения силы), но почти не уставая. Огромное количество митохондрий для такой работы используют сначала запасенные капельки жира, а после них – запасенный гликоген.
При повышении нагрузки, включаются промежуточные мышечные волокна. Интенсивность растет, но мышцы способны поддерживать и такой режим работы в течение некоторого времени, пока не закончится гликоген (как бензин в автомобиле). Все побочные продукты такой работы удаляются в митохондриях как самих промежуточных мышечных волокон, так и в окислительных, куда попадают с потоком крови.
Как только нагрузка возрастает настолько, что требуется включение гликолитических мышечных волокон, то работающая мышца становится обреченной на скорое прекращение работы. Побочных продуктов такого сильного сокращения в мышечных волокнах становится так много, что митохондрии не справляются. И в течение нескольких минут работа, скорее всего, прекратится.
На практике это выглядит так. Мы можем часами ходить, т.к. при ходьбе работают окислительные мышечные волокна.
Мы можем довольно долго бегать трусцой или идти быстро. Повышение нагрузки требует совместной работы и окислительных, и промежуточных мышечных волокон, но митохондрии справляются с утилизацией побочных веществ.
Но как только мы переходим на быстрый бег (спринт), то через короткое время будем вынуждены либо резко снизить скорость, либо вообще перейти на шаг или остановиться. В работу включились все три типа мышечных волокон, и гликолитические производят такое количество побочных веществ, что митохондрии перестают справляться. Мышца становится неспособной поддерживать заданную интенсивность работы.
Из вышесказанного следует вывод. Самыми важными для человека являются именно окислительные мышечные волокна, как способные производить работу (сокращения) в течение долгого времени без утомления.
Именно развитию окислительных мышечных волокон или превращению промежуточных и гликолитических в окислительные должны быть посвящены оздоровительные физические тренировки. Потому что в жизни крайне важно быть способным долго производить полезную работу любой направленности.
Ну, а о том, как это делать, поговорим в другой раз.
Тема сложная, поэтому должно возникать много вопросов. Задавайте, будем разбираться.
Понравилось? Поделитесь!
blogozdorovie.ru
Мышечное сокращение — Википедия
Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Это жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами.
Все виды произвольных движений — ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счёт скелетных мышц. Непроизвольные движения (кроме сокращения сердца) — перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря — обусловлены сокращением гладкой мускулатуры. Работа сердца обеспечивается сокращением сердечной мускулатуры.
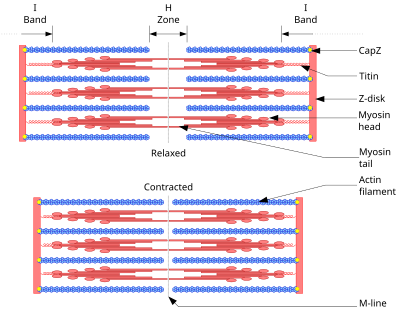 Схема, показывающая мышцы в расслабленном (выше) и сокращённом (ниже) положениях.
Схема, показывающая мышцы в расслабленном (выше) и сокращённом (ниже) положениях.Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечают миофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1-2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновыми филаментами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновых филаментов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела[1].
Изометрическое сокращение[править | править код]
- Основная статья: Изометрическое сокращение[en]
Изометрическое сокращение мышцы создаёт напряжение без изменения длины. Пример можно найти, когда мышцы руки и предплечья захватывают объект; суставы руки не двигаются, но мышцы генерируют достаточную силу, чтобы предотвратить падение объекта.
Изотоническое сокращение[править | править код]
- Основная статья: Изотоническое сокращение[en]
При изотоническом сокращении напряжение в мышцах остаётся постоянным, несмотря на изменение длины мышц. Это происходит, когда сила сокращения мышц соответствует общей нагрузке на мышцы.
Концентрическое сокращение[править | править код]
При концентрическом сокращении мышечное напряжение является достаточным для преодоления нагрузки, а мышца укорачивается при сокращении. Это происходит, когда сила, создаваемая мышцей, превышает нагрузку, противодействующую её сокращению.
Эксцентрическое сокращение[править | править код]
- См. также: Эксцентрическая тренировка[en]
При эксцентрическом сокращении напряжение недостаточно для преодоления внешней нагрузки на мышцы и мышечные волокна, удлиняются при их сокращении. Вместо того, чтобы тянуть сустав в направлении сокращения мышц, мышца действует так, чтобы замедлить сустав в конце движения или иным образом контролировать перемещение груза. Это может происходить невольно (например, при попытке переместить вес, слишком тяжёлый для подъёма мышцы) или добровольно (например, когда мышца «сглаживает» движение или сопротивляется гравитации, как, например, во время ходьбы вниз). В краткосрочной перспективе силовые тренировки с участием как эксцентрических, так и концентрических сокращений, по-видимому, увеличивают мышечную силу больше, чем тренировки только с концентрическими сокращениями. Однако вызванное физическими упражнениями повреждение мышц также больше при удлинении сокращений.
Эксцентрические сокращения в движении[править | править код]
Эксцентрические сокращения обычно возникают как тормозящее усилие в противоположность концентрическому сжатию, чтобы защитить суставы от повреждения. Во время практически любого обычного движения эксцентрические сокращения помогают сохранять плавность движений, но также могут замедлять быстрые движения, такие как удар или бросок. Часть тренировки для быстрых движений, таких как качки во время бейсбола, включает в себя уменьшение эксцентрического торможения, позволяющего развивать большую мощность во время движения.
Скольжение миозина относительно актина[править | править код]
Головки миозина расщепляют АТФ и за счет высвобождающейся энергии меняют конформацию, скользя по актиновым филаментам. Цикл можно разделить на 4 стадии:
- Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата и остаётся связанной с ними. (Обратимый процесс — энергия, выделившаяся в результате гидролиза, запасается в изменённой конформации миозина).
- Головки слабо связываются со следующей субъединицей актина, фосфат отделяется, и это приводит к прочному связыванию головки миозина с актиновым филаментом. Эта реакция уже необратима.
- Головка претерпевает конформационное изменение, производящее подтягивание толстого филамента к Z-диску (или, что эквивалентно, свободных концов тонких филаментов друг к другу).
- Отделяется АДФ, за счёт этого головка отделяется от актинового филамента. Присоединяется новая молекула АТФ.
Далее цикл повторяется до уменьшения концентрации ионов Ca2+ или исчерпании запаса АТФ (в результате смерти клетки). Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновом филаменте много (около 500) молекул миозина и, следовательно, при сокращении цикл повторяется сотнями головок сразу, что и приводит к быстрому и сильному сокращению. Следует заметить, что миозин ведёт себя как фермент — актин-зависимая АТФаза. Так как каждое повторение цикла связано с гидролизом АТФ, а следовательно, с положительным изменением свободной энергии, то процесс однонаправленный. Миозин движется по актину только в сторону плюс-конца.
Последовательные стадииДля сокращения мышцы используется энергия гидролиза АТФ, но мышечная клетка имеет крайне эффективную систему регенерации запаса АТФ, так что в расслабленной и работающей мышце содержание АТФ примерно равно. Фермент фосфокреатинкиназа катализирует реакцию между АДФ и креатинфосфатом, продукты которой — АТФ и креатин. Креатинфосфат содержит больше запасённой энергии, чем АТФ. Благодаря этому механизму при вспышке активности в мышечной клетке падает содержание именно креатинфосфата, а количество универсального источника энергии — АТФ — не изменяется. Механизмы регенерации запаса АТФ могут различаться в зависимости от парциального давления кислорода в окружающих тканях (см. Анаэробные организмы).
В основном в регуляции мышечной активности участвуют нейроны, но есть случаи, когда сокращением гладкой мускулатуры управляют и гормоны (например, адреналин и окситоцин). Сигнал о сокращении можно разделить на несколько этапов:
От клеточной мембраны до саркоплазматического ретикулума[править | править код]
Воздействие медиатора, выделившегося из мотонейрона, вызывает потенциал действия на клеточной мембране мышечной клетки, который передаётся далее с помощью специальных впячиваний мембраны, называемых Т-трубочками, которые отходят от мембраны внутрь клетки. От Т-трубочек сигнал передаётся саркоплазматическому ретикулуму — особому компартменту из уплощенных мембранных пузырьков (эндоплазматической сети мышечной клетки), окружающих каждую миофибриллу. Этот сигнал вызывает открытие Ca2+-каналов в мембране ретикулума. Обратно ионы Ca2+ попадают в ретикулум с помощью мембранных кальциевых насосов — Ca2+-АТФазы.
От выделения ионов Ca2+ до сокращения миофибрилл[править | править код]
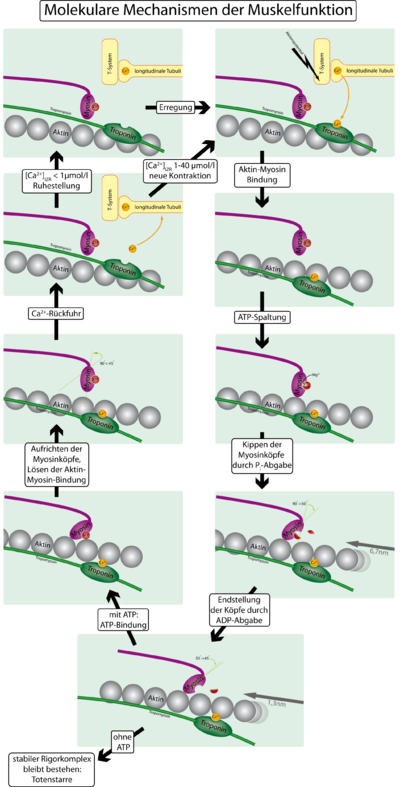 Механизм сокращения мышц с учётом тропонина и тропомиозина
Механизм сокращения мышц с учётом тропонина и тропомиозинаДля того, чтобы контролировать сокращение, к актиновому филаменту прикрепляется белок тропомиозин и комплекс из трёх белков — тропонин (субъединицы этого комплекса называются тропонинами T,I и C). Тропонин C — близкий гомолог другого белка, кальмодулина. Через каждые семь субъединиц актина расположен только один тропониновый комплекс. Связь актина с тропонином I перемещает тропомиозин в положение, мешающее связи миозина с актином. Тропонин C связывается с четырьмя ионами Ca2+ и ослабляет действие тропонина I на актин, и тропомиозин занимает положение, не препятствующее связи актина с миозином. Источником энергии для сокращения мышечных волокон служит АТФ. При связывании тропонина с ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. За счет ферментативной активности головок миозина гидролизуется АТФ, расположенный на головке миозина, что обеспечивает энергией изменение конформации головок и скольжение нитей. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. К миозиновой головке присоединяется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снизится до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
| Белок | Доля белка % | Его мол. масса, kDa | Его функция |
|---|---|---|---|
| Миозин | 44 | 510 | Главный компонент толстых филаментов. Образует связи с актином. Движется по актину за счёт гидролиза АТФ. |
| Актин | 22 | 42 | Главный компонент тонких филаментов. Во время сокращения мышцы по нему движется миозин. |
| Титин | 9 | 2500 | Большой гибкий белок, образующий цепь для связывания миозина с Z-диском. |
| Тропонин | 5 | 78 | Комплекс из трёх белков, регулирующий сокращение при связывании с ионами Ca2+. |
| Тропомиозин | 5 | 64 | Связанный с актиновыми филаментами стержневидный белок, блокирующий движение миозина. |
| Небулин[en] | 3 | 600 | Длинный нерастяжимый белок, связанный с Z-диском и идущий параллельно актиновым филаментам. |
- Б. Альбертс, Д. Брей, Дж. Льюис, М. Рефф, К. Робертс, Дж. Уотсон, Молекулярная биология клетки — В 3-х т. — Пер. с англ. — Т.2. — М.: Мир, 1994. — 540 с.
- М. Б. Беркинблит, С. М. Глаголев, В. А. Фуралев, Общая биология — В 2-х ч. — Ч.1. — М.:МИРОС, 1999. — 224 с.: ил.
ru.wikipedia.org
Гипертрофия скелетных мышц — Википедия
Гипертрофия скелетных мышц (греч. hyper – больше и греч. trophe – питание, пища) – увеличение объёма или массы скелетной мышцы . Уменьшение объёма или массы скелетной мышцы называется атрофией. Уменьшение объёма или массы скелетной мышцы в пожилом возрасте называется саркопенией.
Методика оценки степени гипертрофии[править | править код]
Для того, чтобы оценить степень гипертрофии скелетной мышцы, необходимо измерить изменение её объёма или массы. Современные методы исследования (компьютерная или магнито-резонансная томография) позволяют оценить изменение объёма скелетных мышц человека и животных. С этой целью выполняются многократные «срезы» поперечного сечения мышцы, что позволяет вычислить её объём. Однако, до настоящего времени о степени гипертрофии скелетных мышц достаточно часто судят по изменению максимального значения поперечного сечения мышцы, полученного посредством компьютерной или магниторезонансной томографии.
Показатели, определяющие объём скелетных мышц[править | править код]
Основным компонентом скелетных мышц являются мышечные волокна, которые составляют приблизительно 87% от её объёма (J.D. MacDougall et al., 1984). Этот компонент мышцы называют сократительным, так как сокращение мышечных волокон позволяет мышце изменять свою длину и перемещать звенья опорно-двигательного аппарата, осуществляя движение звеньев тела человека. Остальной объём мышцы (13%) занимают несократительные элементы (соединительно-тканные образования, кровеносные и лимфатические сосуды, нервы, тканевая жидкость и др.). В первом приближении[1] объём всей мышцы (Vм) можно выразить формулой: Vм = Vмв nмв +Vнс, где: Vмв – объём мышечного волокна; nмв – количество мышечных волокон; Vнс – объём несократительной части мышцы (то есть тот объём, который занимают все компоненты мышцы, кроме мышечных волокон).
Влияние тренировки на параметры, определяющие объём скелетных мышц[править | править код]
Доказано, что под влиянием силовой тренировки и тренировки на выносливость возрастает объём мышечных волокон (Vмв) и объём несократительной части мышцы (Vнс). Не доказано увеличения количества мышечных волокон (гиперплазии мышечных волокон) у человека под влиянием силовой тренировки, хотя у животных (млекопитающих и птиц) гиперплазия мышечных волокон доказана[2].
Можно выделить два крайних типа гипертрофии мышечных волокон[3][4]: миофибриллярную гипертрофию и саркоплазматическую гипертрофию.
Миофибриллярная гипертрофия мышечных волокон – увеличение объёма мышечных волокон за счёт увеличения объёма миофибрилл. При этом возрастает плотность укладки миофибрилл в мышечном волокне. Гипертрофия мышечных волокон ведёт к значительному росту максимальной силы мышцы. По-видимому, наиболее предрасположены к миофибриллярной гипертрофии быстрые (IIB тип) мышечные волокна (Я.М. Коц, 1998).
Саркоплазматическая гипертрофия мышечных волокон – увеличение объёма мышечных волокон за счёт преимущественного увеличения объёма саркоплазмы, т. е. несократительной их части. Гипертрофия этого типа происходит за счёт повышения содержания в мышечных волокнах митохондрий, а также: креатинфосфата, гликогена, миоглобина и др. Наиболее предрасположены к саркоплазматической гипертрофии, по-видимому, медленные (I) и быстрые окислительные (IIА) мышечные волокна (Я. М. Коц, 1998). Саркоплазматическая гипертрофия мышечных волокон мало влияет на рост силы мышц, но зато значительно повышает способность к продолжительной работе, т. е. увеличивает их выносливость.
В реальных ситуациях гипертрофия мышечных волокон представляет собой комбинацию двух названных типов с преобладанием одного из них. Преимущественное развитие того или иного типа гипертрофии мышечных волокон определяется характером тренировки. Упражнения со значительными внешними отягощениями (более 70%-от максимума), способствуют развитию миофибриллярной гипертрофии мышечных волокон. Такой тип гипертрофии характерен для силовых видов спорта (тяжёлая атлетика, пауэрлифтинг). Длительное выполнение двигательных действий, развивающих выносливость, с относительно небольшой силовой нагрузкой на мышцы вызывает, главным образом, саркоплазматическую гипертрофию мышечных волокон. Такая гипертрофия свойственна бегунам на средние и длинные дистанции. Спортсменам, занимающихся бодибилдингом, свойственна как миофибриллярная, так и саркоплазматическая гипертрофия мышечных волокон[1][5].
Механизмы гипертрофии скелетных мышц[править | править код]
В основе миофибриллярной гипертрофии мышечных волокон лежит интенсивный синтез и уменьшенный распад мышечных белков. Существует несколько гипотез миофибриллярной гипертрофии:
- гипотеза ацидоза;
- гипотеза гипоксии;
- гипотеза механического повреждения мышечных волокон.
Гипотеза ацидоза предполагает, что пусковым стимулом для повышенного синтеза белка в скелетных мышцах является накопление в них молочной кислоты (лактата). Увеличение лактата в мышечных волокнах вызывает повреждение сарколеммы мышечных волокон и мембран органелл, появление в саркоплазме мышечных волокон ионов кальция, что вызывает активизацию протеолитических ферментов, расщепляющих мышечные белки. Увеличение синтеза белка в этой гипотезе связано с активацией и последующим делением клеток-сателлитов.
Гипотеза гипоксии предполагает, что пусковым стимулом для повышенного синтеза белка в скелетных мышцах является временное ограничение поступления кислорода (гипоксия) к скелетным мышцам, что происходит при выполнении силовых упражнений с большими отягощениями. Гипоксия и последующая реперфузия (восстановление притока кислорода к скелетным мышцам) вызывает повреждение мембран мышечных волокон и органоидов, появление в саркоплазме мышечных волокон ионов кальция, что вызывает активизацию протеолитических ферментов, расщепляющих мышечные белки. Увеличение синтеза белка в этой гипотезе связано с активизацией и последующим делением клеток-сателлитов.
Гипотеза механического повреждения мышечных волокон предполагает, что пусковым стимулом для повышенного синтеза белка является большое мышечное напряжение, что приводит к сильным повреждениям сократительных белков и белков цитоскелета мышечного волокна. Доказано[6], что даже однократная силовая тренировка может привести к повреждению более 80 % мышечных волокон. Повреждение саркоплазматического ретикулума вызывает увеличение в саркоплазме мышечного волокна ионов кальция и последующим процессам, описанным выше.
Согласно вышеописанным гипотезам повреждение мышечного волокна вызывает запаздывающие болезненные ощущения в мышцах (DOMS), что связывается с их воспалением.
Очень важную роль в регуляции объёма мышечной массы, в частности в развитии гипертрофии мышц, играют андрогены (мужские половые гормоны). У мужчин они вырабатываются половыми железами (семенниками) и в коре надпочечников, а у женщин — только в коре надпочечников. Соответственно у мужчин количество андрогенов в организме больше, чем у женщин.
Возрастное развитие мышечной массы идёт параллельно с увеличением продукции андрогенных гормонов. Первое заметное увеличение объёма мышечных волокон наблюдается в 6-7-летнем возрасте, когда усиливается образование андрогенов. С наступлением полового созревания (11-15 лет) начинается интенсивный прирост мышечной массы у мальчиков, который продолжается и после периода полового созревания. У девочек развитие мышечной массы в основном заканчивается с периодом полового созревания.
В опытах на животных установлено, что введение препаратов андрогенных гормонов (анаболиков) вызывает значительную интенсификацию синтеза мышечных белков, в результате чего увеличивается масса тренируемых мышц и как результат — их сила. Вместе с тем, гипертрофия скелетных мышц может происходить и без участия андрогенных и других гормонов (гормона роста, инсулина и тиреоидных гормонов).
Влияние тренировки на композицию и гипертрофию мышечных волокон различных типов[править | править код]
Доказано[7][8][9], что силовая тренировка и тренировка на выносливость не изменяют соотношения в мышцах медленных (I тип) и быстрых (II тип) мышечных волокон. Вместе с тем, эти виды тренировки способны изменять соотношение двух видов быстрых волокон, увеличивая процент мышечных волокон IIA типа и, соответственно, уменьшая процент мышечных волокон IIB типа.
В результате силовой тренировки степень гипертрофии быстрых мышечных волокон (II типа) значительно больше, чем медленных волокон (I типа), тогда как тренировка, направленная на выносливость, ведёт к гипертрофии в первую очередь медленных волокон (I типа). Эти различия показывают, что степень гипертрофии мышечного волокна зависит как от меры его использования в процессе тренировок, так и от его способности к гипертрофии.
Силовая тренировка связана с относительно небольшим числом повторных максимальных или близких к ним мышечных сокращений, в которых участвуют как быстрые, так и медленные мышечные волокна. Однако и небольшого числа повторений достаточно для развития гипертрофии быстрых волокон, что указывает на их большую предрасположенность к гипертрофии по сравнению с медленными волокнами. Высокий процент быстрых волокон (II типа) в мышцах служит важной предпосылкой для значительного роста мышечной силы при направленной силовой тренировке. Поэтому люди с высоким процентом быстрых волокон в мышцах имеют более высокие потенциальные возможности для развития силы и мощности.
Тренировка выносливости связана с большим числом повторных мышечных сокращений относительно небольшой силы, которые в основном обеспечиваются активностью медленных мышечных волокон. Поэтому при тренировке на выносливость более выражена гипертрофия медленных мышечных волокон (I типа) по сравнению с гипертрофией быстрых волокон (II типа).
- Коц Я.М. Спортивная физиология Учебник для институтов физической культуры. — М.: Физкультура и спорт, 1998. — 200 с.
- Самсонова А.В. Гипертрофия скелетных мышц человека: монография Национальный гос. ун-т физ. культуры, спорта и здоровья им. П.Ф. Лесгафта.- СПб.: [б.и.], 2011.- 203 с ил. ISBN 978-5-905064-25-8
- Самсонова А.В. Гипертрофия скелетных мышц человека: Учебное пособие.- 3-е изд. — СПб.: Политехника, 2015.- 159 с. ил.- (Серия «Силовая тренировка»). ISBN 978-5-7325-1063-8
- Солодков А.С., Сологуб Е.Б. Физиология человека. Общая. Спортивная. Возрастная: Учебник. М.: Терра-Спорт, Олимпия пресс. 2001. 520 с. ил. ISBN 5-94299-037-9
- Язвиков В.В. Влияние спортивной тренировки на состав мышечных волокон смешанных скелетных мышц человека // Теория и практика физической культуры. 1988. № 2. С. 48-50.
- Язвиков В.В., Морозов С.А., Некрасов А.Н. Корреляция между содержанием медленных волокон в наружной широкой мышце бедра и спортивными результатами // Физиология человека, 1990. – Т.16. – № 4. – С. 167-169.
- Alway S. E., MacDougall J. D., Sale D. G. Contractile adaptations in the human triceps surae after isometric exercise // Journal of Applied Physiology. 1989. V. 66. P. 2725–2732.
- Gibala, M.J., MacDougall J.D., Tarnopolsky M.A., Stauber W.T., Elorriaga A. Changes in human skeletal muscle ultrastructure and force production after acute resistance exercise // Journal of Applied Physiology, 1995. – V.78. – P. 702-708.
- MacDougall J.D., Elder G.C.B., Sale D.G., Moroz J.R. & Sutton, J.R. Effects of strength training and immobilization on human muscle fibers // European Journal of Applied Physiology. 1980. V. 43. P. 25–34.
- MacDougall J.D., Sale D.G., Alway S.E., Sutton J.R. Muscle fiber number in biceps brachii in bodybuilders and control subjects // Journal Applied Physiology, 1984. V 57. № 5. P. 1399-1403.
- MacDougall J. D. Hypertrophy and Hyperplasia // In: The Encyclopedia of Sport Medicine. Strength and Power in Sport / Ed. P.V. Komi: Blackwell Publishing, Bodmin, Cornwall, 2003. V.3. P. 252–264.
- Zatsiorsky V.M., Kramer W.J. Science and Practice of Strength. Champaign, IL: Human Kinetics. 2006. 251 p. ISBN 978-0-7360-5628-1
- ↑ 1 2 Самсонова А.В. Гипертрофия скелетных мышц человека: Учебное пособие. — 3-е изд. — СПб.: Политехника, 215.
- ↑ MacDougall J. D. Hypertrophy and Hyperplasia // In: The Encyclopedia of Sport Medicine. Strength and Power in Sport. — Bodmin, Cornwall: Blackwell Publishing, 2003.
- ↑ Коц Я.М. Спортивная физиология Учебник для институтов физической культуры. — М.: Физкультура и спорт, 1998.
- ↑ Солодков А.С., Сологуб Е.Б. Физиология человека. Общая. Спортивная. Возрастная: Учебник. — М.: Терра-Спорт, Олимпия пресс, 2001.
- ↑ Zatsiorsky V.M., Kramer W.J. Science and Practice of Strength. — IL: Human Kinetics, 2006.
- ↑ Gibala, M.J., MacDougall J.D., Tarnopolsky M.A., Stauber W.T., Elorriaga A. Changes in human skeletal muscle ultrastructure and force production after acute resistance exercise // Journal of Applied Physiology. — 1995. — С. 702—708.
- ↑ MacDougall J.D., Elder G.C.B., Sale D.G., Moroz J.R. & Sutton, J.R. Effects of strength training and immobilization on human muscle fibers // European Journal of Applied Physiology. — 1980. — № 43. — С. 25–34.
- ↑ Язвиков В.В. Влияние спортивной тренировки на состав мышечных волокон смешанных скелетных мышц человека // Теория и практика физической культуры. — 1988. — № 2. — С. 48—50.
- ↑ Язвиков В.В., Морозов С.А., Некрасов А.Н. Корреляция между содержанием медленных волокон в наружной широкой мышце бедра и спортивными результатами // Физиология человека. — 1990. — Т. 16, № 4. — С. 167—169.
ru.wikipedia.org
Мышечная композиция — Википедия
Материал из Википедии — свободной энциклопедии
Мышечная композиция или композиция мышц — процентное соотношение мышечных волокон различного типа, которое для каждого конкретного человека предопределено генетически и закладывает его предрасположенность к определённым видам спортивных дисциплин[1][2].
Человеческая мышечная ткань составлена из двух основных типов мышечных волокон. Первый из них (тип I) условно называют «медленно сокращающимися», второй — «быстро сокращающимися» (тип II)[1][2].
Медленно сокращающиеся мышечные волокна оптимизированы эволюцией под задачи выполнения продолжительной работы аэробного характера, и поэтому характеризуются меньшей скоростью действий по сравнению с «быстрыми», но прекрасно развитой системой аэробного окисления. В свою очередь быстро сокращающиеся волокна обладают быстротой для выполнения кратковременных действий высокой интенсивности в анаэробном режиме и хорошо развитыми механизмами восполнения запасов АТФ. Стоит заметить, что внутри быстрых мышечных волокон выделяются два подтипа[1][2]:
- окислительно-гликолитические (тип II-A) с хорошо развитой способностью к аэробному окислению и энергообеспечению за счёт гликолиза,
- гликолитические (тип II-B) с преобладанием энергообеспечения за счёт гликолиза и креатинфосфатного пути.
Считается, что люди с преобладанием волокон первого типа предрасположены к длительным умеренным нагрузкам, а лица с преобладанием волокон второго типа — к силовым и скоростным видам спорта. Процентная доля волокон различного типа в мышечной массе отличается довольно сильно от человека к человеку, при этом спортивная тренировка может изменить соотношение разных типов (I и II) в очень ограниченном диапазоне значений (от 2 до 10 %), однако, соотношение двух подтипов (II-A и II-B) в некоторой степени поддаётся действию тренировок[1][2].
Известно также, что очень высокий процент медленных волокон типичен для людей никогда не занимавшихся спортом. Кроме этого, процент медленных и быстрых волокон в интенсивно и мало тренируемых мышцах у атлетов одинаковой спортивной специализации примерно одинаков и не зависит от тренировок даже многомесячной продолжительности. Однако такие тренировки однозначно влияют на факторы, определяющие выносливость (толщину медленных волокон, активность мышечных ферментов в окислительном метаболизме и т. п.)[2]
- ↑ 1 2 3 4 Киселёв В. А., Черемисинов В. Н. Факторы, определяющие скоростно-силовые способности // Физическая подготовка боксёра. — Москва: Дивизион, 2013. — С. 64. — 160 с. — ISBN 978-5-98724-114-1.
- ↑ 1 2 3 4 5 Коц Я. М. Композиция мышц // Спортивная физиология. Учебник для институтов физической культуры. — Москва: Физкультура и спорт, 1998. — 200 с.
ru.wikipedia.org
Мышцы — это… Что такое Мышцы?
 Старинный рисунок мышц человека
Старинный рисунок мышц человека 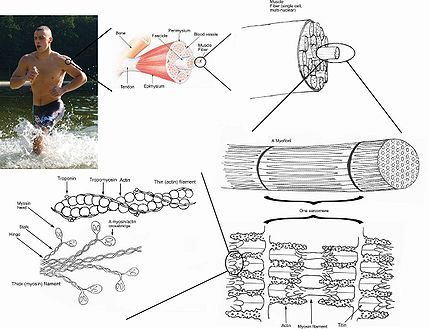 Строение скелетной мышцы
Строение скелетной мышцыМышцы или мускулы (от лат. musculus — мышка, маленькая мышь) — органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86,3 % из воды.
Мышцы позволяют двигать частями тела и выражать в действиях мысли и чувства. Человек выполняет любые движения — от таких простейших, как моргание или улыбка, до тонких и энергичных, какие мы наблюдаем у ювелиров или спортсменов — благодаря способности мышечных тканей сокращаться. От исправной работы мышц, состоящих из трёх основных групп, зависит не только подвижность организма, но и функционирование всех физиологических процессов. А работой всех мышечных тканей управляет нервная система, которая обеспечивает их связь с головным и спинным мозгом и регулирует преобразование химической энергии в механическую.
В теле человека 640 мышц (в зависимости от метода подсчёта дифференцированных групп мышц их общее число определяют от 639 до 850). Самые маленькие прикреплены к мельчайшим косточкам, расположенным в ухе. Самые крупные — большие ягодичные мышцы, они приводят в движение ноги. Самые сильные мышцы — икроножные(18,6), жевательные(10,2).
По форме мышцы очень разнообразны. Чаще всего встречаются веретенообразные мышцы, характерные для конечностей, и широкие мышцы — они образуют стенки туловища. Если у мышц общее сухожилие, а головок две или больше, то их называют двух-, трёх- или четырёхглавые мышцы.
Мышцы и скелет определяют форму человеческого тела. Активный образ жизни, сбалансированное питание и занятие спортом способствуют развитию мышц и уменьшению объёма жировой ткани.
Строение
Минимальный структурный элемент всех типов мышц — мышечное волокно, каждое из которых в отдельности является не только клеточной, но и физиологической единицей, способной сокращаться. Это связано со строением такого волокна, содержащего не только органеллы (ядро клетки, митохондрии, рибосомы, комплекс Гольджи), но и специфические элементы, связанные с механизмом сокращения — миофибриллы. В состав последних входят сократительные белки — актин и миозин.
Актин — сократительный белок, состоящий из 375 аминокислотных остатков с молекулярной массой 42300, который составляет около 15 % мышечного белка. Под световым микроскопом более тонкие молекулы актина выглядят светлой полоской (так называемые Ι-диски). В растворах с малым содержанием ионов актин содержится в виде единичных молекул с шарообразной структурой, однако в физиологических условиях, в присутствии АТФ и ионов магния, актин становится полимером и образует длинные волокна (актин фибриллярный), которые состоят из спирально закрученных двух цепочек молекул актина. Соединяясь с другими белками, волокна актина приобретают способность сокращаться, используя энергию, содержащуюся в АТФ.
Миозин — основной мышечный белок; содержание его в мышцах достигает 60 %. Молекулы состоят из двух полипептидных цепочек, в каждой из которых содержится более 2000 аминокислот. Белковая молекула очень велика (это самые длинные полипептидные цепочки, существующие в природе), а её молекулярная масса доходит до 470000. Каждая из полипептидных цепочек оканчивается так называемой головкой, в состав которой входят две небольшие цепочки, состоящие из 150—190 аминокислот. Эти белки проявляют энзиматическую активность АТФазы, необходимую для сокращения актомиозина. Под микроскопом молекулы миозина в мышцах выглядят темной полоской (так называемые А-диски).
Актомиозин — белковый комплекс, состоящий из актина и миозина, характеризующийся энзиматической активностью АТФазы. Это значит, что благодаря энергии, освобожденной в процессе гидролиза АТФ, актомиозин может сокращаться. В физиологических условиях актомиозин создает волокна, находящиеся в определенном порядке. Фибриллярные части молекул миозина, собранные в пучок, образуют так называемую толстую нить, из которой перпендикулярно выглядывают миозиновые головки. Молекулы актина соединяются в длинные цепочки; две таких цепочки, спирально закрученные друг вокруг друга, составляют тонкую нить. Тонкая и толстая нити расположены параллельно таким образом, что каждая тонкая нить окружена тремя толстыми, а каждая толстая нить — шестью тонкими; миозиновые головки цепляются за тонкие нити.
Типы мышц
В зависимости от особенностей строения мышцы человека делят на 3 типа или группы.
Первая группа мышц — скелетные, или поперечнополосатые мышцы. Скелетных мышц у каждого из нас более 600. Мышцы этого типа способны произвольно, по желанию человека, сокращаться и вместе со скелетом образуют опорно-двигательную систему. Общая масса этих мышц составляет около 40 % веса тела, а у людей, активно развивающих свои мышцы, может быть ещё больше. С помощью специальных упражнений размер мышечных клеток можно увеличивать до тех пор, пока они не вырастут в массе и объёме и не станут рельефными. Сокращаясь, мышца укорачивается, утолщается и движется относительно соседних мышц. Укорочение мышцы сопровождается сближением её концов и костей, к которым она прикрепляется. В каждом движении участвуют мышцы как совершающие его, так и противодействующие ему, что придаёт движению точность и плавность.
Второй тип мышц, который входит в состав клеток внутренних органов, кровеносных сосудов и кожи, — гладкая мышечная ткань, состоящая из характерных мышечных клеток (миоцитов). Короткие веретеновидные клетки гладких мышц образуют пластины. Сокращаются они медленно и ритмично, подчиняясь сигналам вегетативной нервной системы. Медленные и длительные их сокращения происходят непроизвольно, то есть независимо от желания человека.
Гладкие мышцы, или мышцы непроизвольных движений, находятся главным образом в стенках полых внутренних органов, например пищевода или мочевого пузыря. Они играют важную роль в процессах, не зависящих от нашего сознания, например в перемещении пищи по пищеварительному тракту.
Отдельную (третью) группу мышц составляет сердечная поперечнополосатая (исчерченная) мышечная ткань (миокард). Она состоит из кардиомиоцитов. Сокращения сердечной мышцы не подконтрольны сознанию человека, она иннервируется вегетативной нервной системой.
Классификация
Мышечная ткань живых организмов представлена многочисленными мышцами различной формы, строения, процесса развития, выполняющими разнообразные функции. Различают:
по функции
- сгибатели (лат. flexores)
- разгибатели (лат. extensores)
- отводящие (лат. abductores)
- приводящие (лат. adductores)
- вращатели (лат. rotatores) кнутри (лат. pronatores) и кнаружи (лат. supinatores)
- сфинктеры и делятаторы
- синергисты и антагонисты
по направлению волокон
- прямая мышца — с прямыми параллельными волокнами
- поперечная мышца — с поперечными волокнами
- круговая мышца — с круговыми волокнами
- косая мышца — с косыми волокнами
- одноперистая — косые волокна прикрепляются к сухожилию с одной стороны
- двуперистая — косые волокна прикрепляются к сухожилию с двух сторон
- многоперистая — косые волокна прикрепляются к сухожилию с нескольких сторон
- полусухожильная
- полуперепончатая
по отношению к суставам
Учитывается число суставов, через которые перекидывается мышца:
- односуставные
- двусуставные
- многосуставные
По форме
- простые
- веретенообразные
- прямые
- длинные (на конечностях)
- короткие
- широкие
- сложные
- многоглавые
- двуглавые
- трехглавые
- четырехглавые
- многосухожильные
- двубрюшные
- с определенной геометрической формой
- квадратные
- дельтовидные
- камбаловидные
- пирамидальные
- круглые
- зубчатые
- треугольные
- ромбовидные
- трапециевидные
- многоглавые
Сокращения мышц
В процессе сокращения нити актина проникают глубоко в промежутки между нитями миозина, причем длина обеих структур не меняется, а лишь сокращается общая длина актомиозинового комплекса — такой способ сокращения мышц называется скользящим. Скольжение актиновых нитей вдоль миозиновых нуждается в энергии, энергия, необходимая для сокращения мышц, освобождается в результате взаимодействия актомиозина с АТФ с расщеплением последнего на АДФ и H3PO4.’ Кроме АТФ важную роль в сокращении мышц играет вода, а также ионы кальция и магния. Скелетная мышца состоит из большого количества мышечных волокон — чем их больше, тем сильнее мышца.
Различают два типа мышечных сокращений. Если оба конца мышцы неподвижно закреплены, происходит изометрическое сокращение, и при неизменной длине напряжение увеличивается. Если один конец мышцы свободен, то в процессе сокращения длина мышцы уменьшится, а напряжение не изменяется — такое сокращение называют изотоническим; в организме такие сокращения имеют большее значение для выполнения любых движений.
Из гладких мышц (гладкой мышечной ткани) состоят внутренние органы, в частности, стенки пищевода, кровеносные сосуды, дыхательные пути и половые органы. Гладкие мышцы отличаются так называемым автоматизмом, то есть способностью приходить в состояние возбуждения при отсутствии внешних раздражителей. И если сокращение скелетных мышц продолжается около 0,1 сек, то более медленные сокращения гладких мышц продолжается от 3 до 180 сек. В пищеводе, половых органах и мочевом канале возбуждение передаётся от одной мышечной клетки к следующей. Что касается сокращения гладких мышц, находящихся в стенках кровеносных сосудов и в радужной оболочке глаза, то оно не переносится с клетки на клетку; к гладким мышцам подходят симпатические и парасимпатические нервы автономной нервной системы.
Говоря о сердечной мышце (миокарде), следует отметить, что при нормальной работе она затрачивает на сокращение около 1 сек, а при увеличении нагрузки скорость сокращений увеличивается. Уникальная особенность сердечной мышцы — в ее способности ритмично сокращаться даже при извлечении ее из организма.
Мышцы
См. также
Примечания
Литература
dic.academic.ru
Гладкие мышцы — Википедия
Материал из Википедии — свободной энциклопедии
 Гладко-мышечная ткань, гематоксилин-эозин.
Гладко-мышечная ткань, гематоксилин-эозин.Гладкие мышцы — сократимая ткань, в отличие от поперечнополосатых мышц не имеющая поперечной исчерченности.
Гладкие мышцы у беспозвоночных и позвоночных[править | править код]
У некоторых беспозвоночных гладкие мышцы образуют всю мускулатуру тела. У позвоночных они входят в состав оболочек внутренних органов: кишечника, кровеносных сосудов, дыхательных путей, выделительных и половых органов, а также многих желёз. Клетки гладких мышц у беспозвоночных разнообразны по форме и строению; у позвоночных в большинстве случаев веретенообразные, сильно вытянутые, с палочковидным ядром, длиной 50—250 мкм, в матке беременных животных — до 500 мкм; окружены волокнами соединительной ткани, образующими плотный футляр.
Сократимый материал — протофибриллы — обычно располагается в цитоплазме изолированно; только у некоторых животных они собраны в пучки — миофибриллы. В гладких мышцах найдены все три вида сократимого белка — актин, миозин и тропомиозин. Преимущественно встречаются протофибриллы одного типа (диаметром около 100 мкм).
Клеточных органоидов (митохондрии, комплекс Гольджи, элементы эндоплазматического ретикулума) в гладких мышцах меньше, чем в поперечнополосатой мускулатуре. Они располагаются преимущественно на полюсах ядра в цитоплазме, лишённой сократимых элементов. Клеточная мембрана часто образует карманы в виде пиноцитозных пузырьков, что указывает на резорбцию и всасывание веществ поверхностью клетки.
Установлено, что гладкие мышцы — группа различных по происхождению тканей, объединяемых единым функциональным признаком — способностью к сокращению. Так, у беспозвоночных гладкие мышцы развиваются из мезодермальных листков и целомического эпителия. У позвоночных гладкие мышцы слюнных, потовых и молочных желёз происходят из эктодермы, гладкие мышцы внутренних органов — из мезенхимы и т. д. Соседние клетки гладких мышц контактируют друг с другом отростками так, что мембраны двух клеток соприкасаются. В мышцах кишечника мыши зоны контакта занимают 5 % поверхности клеточной мембраны. Здесь, вероятно, происходит передача возбуждения от одной клетки к другой (см. Синапсы).
В отличие от поперечнополосатых мышц, для гладких мышц характерно медленное сокращение, способность долго находиться в состоянии сокращения, затрачивая сравнительно мало энергии и не подвергаясь утомлению. Двигательная иннервация гладких мышц осуществляется отростками клеток вегетативной нервной системы, чувствительная — отростками клеток спинальных ганглиев. Не каждая клетка гладких мышц имеет специализированное нервное окончание.
ru.wikipedia.org