Есть ли в помидорах крахмал. В каких продуктах содержится крахмал
Крахмал – это сложный углевод-полисахарид. Он необходим нашему организму для нормального функционирования. Именно углеводы являются источником энергии для организма. Они легко усваиваются и уступают по своей энергетической ценности только моносахаридам — простым углеводам.
Крахмалы есть двух видов – природные и рафинированные. Рафинированный крахмал – белый порошок без вкуса и запаха. Он применяется в кулинарии для приготовления фруктовых киселей, которые снабжают организм большим количеством килокалорий. Его делают из картофеля, кукурузы, риса, пшеницы и ячменя.
Есть также и модифицированный крахмал, который добавляют в продукты как консервант. С его помощью регулируется консистенция различных соусов и детского питания. Некоторые производители добавляют модифицированный крахмал в низкокачественные мясные продукты для удержания влаги в них.
В каких продуктах содержится крахмал
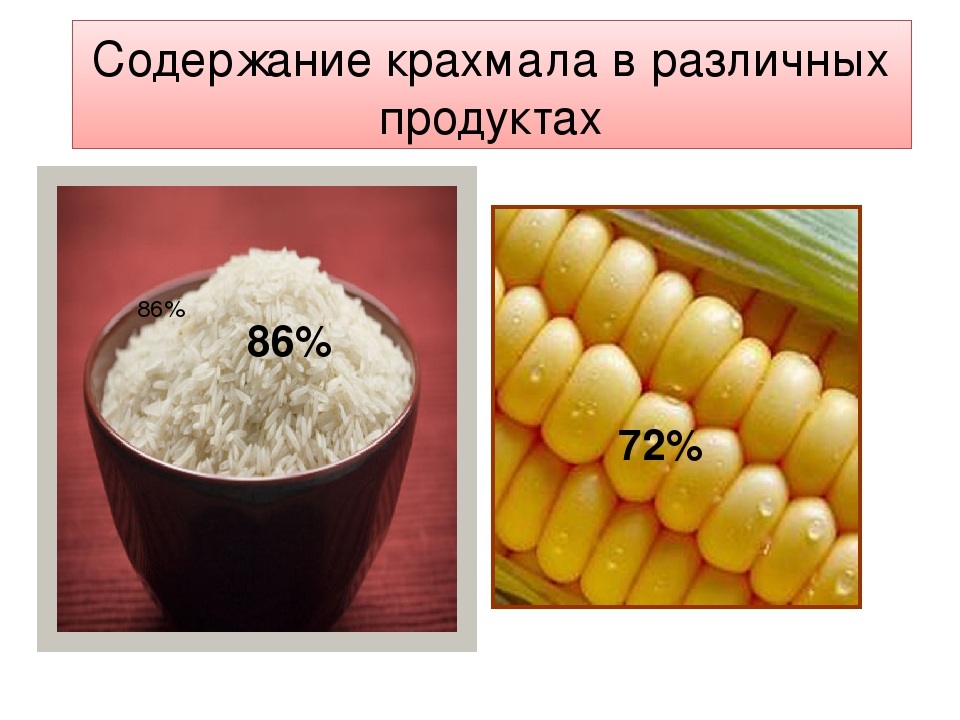
Почти все фрукты, овощи, бобовые и злаки содержат в своем составе крахмал в том или ином количестве.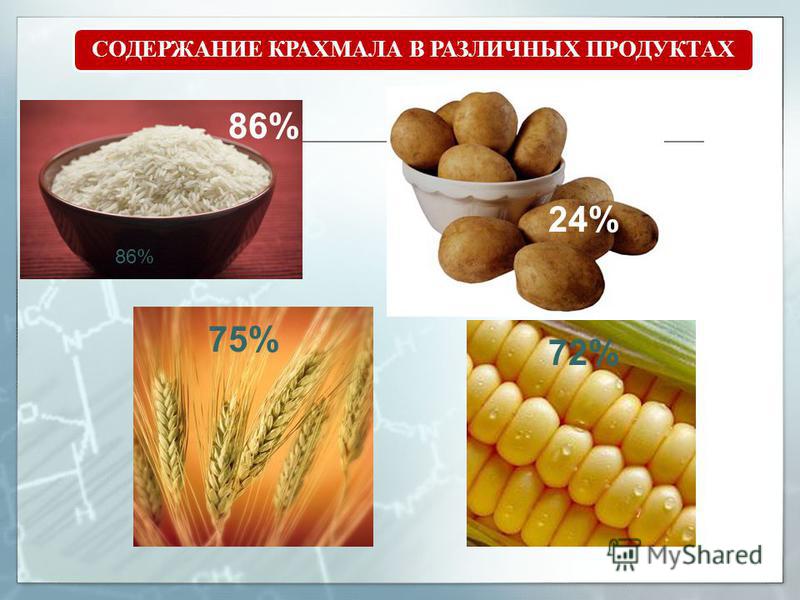
Продукты с высоким содержанием крахмала:
- бобовые: фасоль, чечевица, соя, горох;
- злаки: кукуруза, рис, гречка, нут;
- корнеплоды: картофель, батат;
Продукты с низким содержанием крахмала:
- морковь;
- свекла;
- репа;
- баклажан;
- кабачок.
Продукты, не содержащие крахмала:
- помидоры;
- огурцы;
- болгарский перец;
- капуста;
- листовые овощи и зелень (салат, щавель, шпинат).
Людям, которые вынуждены следить за уровнем сахара в крови, следует учитывать, что некоторые продукты, содержащие крахмал, имеют больший гликемический индекс, чем сахар. Именно свойство крахмала превращаться в глюкозу, делает его потенциально опасным для людей, страдающих сахарным диабетом.
Но крахмал – очень интересный углевод, который имеет свойство усваиваться с разной скоростью. А скорость усвоения крахмала зависит от способа обработки и последующего приготовления пищи из продуктов, содержащих крахмал.
Изделия из муки
Вся выпечка, даже та, в которую не было добавлено сахар, имеет очень высокий гликемический индекс. Степень усвоения крахмала и его превращение в глюкозу приводит к быстрому скачку уровня сахара в крови. Это нужно учитывать диабетикам.
Именно переработка зерновых, которые богаты крахмалом, в муку приводит к тому, что крахмал становится «быстрым углеводом» в данном случае.
Зато хлеб из цельных злаков усваивается значительно дольше, а часть крахмала вообще остается в первозданном виде. Ржаной хлеб или выпечка с отрубями имеет в своем составе резистентный крахмал, который способен даже понизить уровень сахара в крови, но при этом остается прекрасным источником энергии для организма.
Наличие клетчатки, которая есть в хлебе из муки грубого помола, способствует медленному усвоению крахмала, что гарантирует длительную выработку энергии из углеводов. Кроме того, клетчатка очищает кишечник, выводит из него шлаки и токсины.
Макаронные изделия из твердых сортов пшеницы, да еще и приготовленные по классическому итальянскому рецепту, то есть аль-денте, позволяют крахмалу долго растворяться в организме, переходя в глюкозу. Именно способ приготовления и самих макарон как сырья, и итальянской пасты способствует такому их свойству.
Именно способ приготовления и самих макарон как сырья, и итальянской пасты способствует такому их свойству.
Крахмал во фруктах
Фрукты цветные: вишня, красная черешня, смородина и им подобные, практически не имеют в составе крахмал, а лишь его следы – моносахарид глюкозу, которая практически не поднимает уровень сахара в крови
Яблоки и груши зеленых сортов содержат 0,5% крахмала. А если подвергнуть их термической обработке, например запеканию, то крахмал преобразуется в пектин, клетчатку и глюкозу.
Есть некоторое количество крахмала в бананах. Его уровень зависит от степени спелости фрукта. Чем зеленее банан – тем больше в нем крахмала.
Самый полезный крахмал
Полезный, или резистентный крахмал усваивается организмом человека достаточно долго. Углевод длительное время превращается в глюкозу с выделением большого количества энергии, которая способствует правильному функционированию организма. Благодаря этому проходит деление клеток в тканях и органах, происходят обменные процессы. Человек может делать работу, сопряженную с физическими нагрузками.
Человек может делать работу, сопряженную с физическими нагрузками.
Резистентный крахмал необходим организму в целях предотвращения онкологических заболеваний, так как он угнетающе действует на раковые клетки, не давая им делиться.
Полезный крахмал содержится в большом количестве в бобовых. Фасоль и чечевица считаются лидерами по содержанию «полезного» крахмала.
Следом за ними идут цельные злаки. Крупы, такие как гречка, рис, овес являются прекрасными поставщиками сложных углеводов. К тому же, каши очень вкусны в качестве гарнира к мясным и рыбным блюдам.
Картофель, топинамбур, батат, ямс – корнеплоды также богаты «полезным» крахмалом.
Резистентный крахмал в небольших количествах содержится в свежих овощах и фруктах, которые необходимо включать в ежедневный рацион.
Содержание углеводов в продуктах и их энергетическая ценность
| Наименование продукта | К-во крахмала (мг/100гр) | % от дневной нормы |
| Рис | 78 | 44 |
| Кукурузные хлопья | 74 | 42 |
| Мука пшеничная | 72 | 41 |
| Макароны | 70 | 40 |
| Пшено | 69 | 39 |
| Хлеб белый | 66 | 37 |
| Кукурузная мука | 65 | 37 |
| Гречка | 64 | 36 |
| Кукуруза свежая | 62 | 35 |
| Овес | 61 | 34 |
| Пшеница | 60 | 34 |
| Ячмень | 58 | 33 |
| Банан | 53 | 30 |
| Хлеб ржаной | 48 | 27 |
| Горох | 45 | 25 |
| Орехи кешью | 23 | 13 |
| Фисташки | 16 | 9 |
| Картофель коричневый | 15 | 8 |
| Тыквенные семечки | 14 | 8 |
| Кедровые орехи | 14 | 8 |
| Морковь | 14 | 8 |
| Картофель белый | 13 | 7 |
| Батат | 13 | 7 |
| Миндаль | 7 | 4 |
| Фундук | 4 | 2. 2 2 |
| Авокадо | 1.1 | 0.6 |
| Нектарин | 0.4 | |
| Грецкий орех | 0.6 | 0.3 |
| Яблоко | 0.5 | 0.2 |
| Земляника | 0.4 | 0.2 |
| Дыня | 0.3 | 0.1 |
Свойства крахмала
Когда мы употребляем в пищу крохмалсодержащие продукты, то в процессе гидрализации крахмал превращается в глюкозу, которая и является источником энергии. Глюкоза растворима и именно она способствует транспортировке полезных веществ внутрь клетки.
Если рафинированный крахмал прекрасно поддается гидрализации благодаря очищенной форме, то в природном виде он достаточно сложен для переваривания.
Именно поэтому все пищевое сырье, содержащее крахмал, должно проходить термическую обработку. Продукты варят, пекут, жарят, готовят на пару. Такой вид обработки способствует подготовке крахмала к процессу переваривания. В таком виде он легче усваивается организмом.
Чем мельче частицы продукта, тем легче усваивается крахмал.
То есть, цельные злаки, крупы, бобовые являются источником более «длительного» крахмала, который усваивается дольше, но при этом полезнее для организма.
Он не вызывает выброс глюкозы в кровь, а значит и поднятия уровня сахара.
Мука, особенно пшеничная, является источником «быстрого крахмала», который не очень-то полезен из-за быстроты всасывания глюкозы. Даже мелко порубленные крахмалистые овощи при приготовлении будут давать больше «быстрого крахмала», нежели сваренные крупными кусками, целиком или запеченные.
Человеку для полноценной жизни нужно все – белки, углеводы, жиры. Но во всем необходима мера. Каким бы не было полезным то или иное вещество, его переизбыток обязательно приведет к сбою в работе.
Каким бы не было полезным то или иное вещество, его переизбыток обязательно приведет к сбою в работе.
Все это в полной мере относится к крахмалу. Его недостаток вызовет истощение и упадок сил. Его переизбыток может повлечь за собой проблемы с работой сердечно-сосудистой и пищеварительной системы.
Будьте внимательны к своему здоровью и следите за питанием. Ведь от нашего образа жизни зависти и ее качество.
Крахмал, превращающийся в ходе переваривания в глюкозу, относится к наиболее распространенной в природе форме полисахаридов. Поэтому деление овощей на крахмалистые и некрахмалистые, изначально являвшееся частью теории , нашло место в универсальной . Некрахмалистые овощи — настоящие помощники в похудении, а вот с крахмалистыми стоит держать ухо востро! Но как не перепутать? Поможет наш справочный сервис с удобными таблицами.
Овощное меню — не синоним меню для похудения! Овощи бывают разные, и, соответственно, различаются правила их сочетания друг с другом и с другими продуктами
Наибольшим процентом содержания крахмала отличаются корнеплоды и крупные зерна, накапливающие питательные вещества для продолжения роста и обеспечения «продовольственным запасом» зародыша растения. Самым «выраженно крахмальным» овощем, бесспорно, является картофель — крахмал может представлять до 1/5 части объема клубня! Именно поэтому тем, кто хочет похудеть, в первую очередь отказаться от картошки.
Самым «выраженно крахмальным» овощем, бесспорно, является картофель — крахмал может представлять до 1/5 части объема клубня! Именно поэтому тем, кто хочет похудеть, в первую очередь отказаться от картошки.
Крахмалистые овощи: полный список
| Брюква |
| Кукуруза |
| Морковь |
| Свекла |
| Зрелые (сухие) бобы, кроме соевых |
| Зрелый (сухой) горох |
| Кабачки |
| Патиссоны |
| Картофель (все виды и сорта, включая сладкий) |
| Каштаны |
| Корни растений, пригодных к употреблению в пищу (хрен, петрушка, пастернак, сельдерей) |
| Тыква (круглая, осеннего созревания) |
| Топинамбур |
| Редька |
| Редис |
Умеренно крахмалистые овощи: два варианта списка
Некрахмалистые овощи: полный список
| Баклажан |
| Брокколи |
| Брюссельская капуста |
| Горчица |
| Зеленый горошек |
| Пекинская (китайская) капуста |
| Кольраби |
| Кочанная капуста (белокочанная, краснокочанная, савойская, огородная, кормовая) |
| Кресс-салат и водяной кресс |
| Спаржа |
| Летняя тыква (продолговатая желтая) |
| Латук и другие виды листового салата |
| Ботва репы и другие наземные зеленые части съедобных растений |
| Свекольные листья и листовая свекла (мангольд) |
| Лук (репчатый, шалот, порей, шнитт-лук, порей) |
| Огурец |
| Зелень одуванчика |
| Окра |
| Бамия |
| Петрушка (зелень) и другие столовые травы |
| Побеги бамбука |
| Сурепка (зелень) |
| Сельдерей (зелень) |
| Сладкий перец |
| Цикорий |
| Чеснок (зелень и зубчики) |
| Шпинат |
| Щавель |
Овощи, не содержащие крахмала
В рацион каждого здорового человека обязательно входят овощи. Список выращиваемых овощей чрезвычайно широк и включает в себя многие десятки наименований. Но не все овощи могут сочетаться в питании с другими группами продуктов.
Список выращиваемых овощей чрезвычайно широк и включает в себя многие десятки наименований. Но не все овощи могут сочетаться в питании с другими группами продуктов.
Овощи некрахмалистые
Список их включает такие овощи, как огурцы, корнишоны, капуста, (белокочанная, брюссельская, красная, брокколи), болгарский перец, спаржа, перец, турнепс, лук и другие.
В питании их можно сочетать с рыбой, мясом, фруктами, зеленью и жирами. С некрахмалистыми прекрасно сочетаются и крахмалистые овощи.
Список некрахмалистых овощей более широк по сравнению с крахмалистыми. Это вызвано тем, что сюда входит разнообразная зелень. Это петрушка, укроп, базилик, сельдерей, салат-латук, ревень, портулак, лук-порей, лист одуванчика и крапивы, щавель, спаржа, руккола, и так далее. Некрахмалистые овощи сочетаются со многими другими пищевыми продуктами и являются обязательной частью систем здорового питания.
При раздельном питании недопустимы только сочетания некрахмалистых овощей и молокопродуктов.
Овощи крахмалистые
Список крахмалистых овощей составляют: баклажаны, кабачки, морковь, зеленый горошек, тыква, свекла, редька, репа, патиссоны, кукуруза, брюква, топинамбур, сельдерея и хрена. В перечень продуктов, с которыми они превосходно сочетаются, входят и некрахмалистые овощи.
Список овощей, которые богаты крахмалами, включает и цветную капусту, что не совсем очевидно. При употреблении крахмалистых овощей необходимо дополнять пищу нетяжелыми жирами (сливками, сметаной, растительным маслом). При таком дополнении эти продукты полезны для организма человека и хорошо усваиваются.
Особые продукты
Особое место в отведено помидорам. Эти овощи особенно богаты кислотами. По сочетаемости они схожи с некоторыми фруктами, такими как гранаты или цитрусовые.
Мы рассмотрели два класса овощей. А в какой из них входит наша любимая картошка? По мнению диетологов, она не относится к овощам, но входит в перечень крахмалистых продуктов, как и злаки.
Промежуточным звеном между рассмотренными группами овощей при раздельном питании являются бобовые. Большинство бобовых, как и злаки, имеют большое содержание крахмала. Например, фасоль, сушеный горох и чечевица имеют до 45% крахмалов, но содержат и немало растительного белка. Поэтому для переваривания бобовые и злаки — тяжелая пища. Только соя среди бобовых содержит всего 3% крахмалов.
Любые овощи, список которых составляют обе рассмотренные группы овощей, лучше употреблять в пищу в сыром виде или после паровой обработки. Только при таком приготовлении в них остаются все необходимые для полноценного питания минеральные вещества, витамины и клетчатка.
Крахмал — сложный углевод, известный нам, как белый порошок, похожий на муку. Он не имеет запаха и выраженного вкуса, а вырабатывают его из различных растений, в которых он синтезируется. Из всех потребляемых человеком углеводов, на крахмал приходится около 80%.
Попадая в ЖКТ, этот углевод преобразуется в глюкозу, которая обеспечивает организм энергией. Поэтому при недостаточном употреблении продуктов, его содержащих, человек ощущает упадок сил, слабость и быстро утомляется.
Поэтому при недостаточном употреблении продуктов, его содержащих, человек ощущает упадок сил, слабость и быстро утомляется.
Сегодня на сайте «Популярно о здоровье», мы поговорим с вами про его пользу и вред. Вы узнаете, в каких продуктах содержится крахмал, список овощей и фруктов ходовых составим и обсудим далее…
Список продуктов, богатых крахмалом
Прежде всего, обратите внимание, что наиболее богаты этим веществом различные злаки: кукуруза, рис, пшеница и овес. Там его содержится, около 70-75%. Много этого вещества в зрелом горохе и бобах (за исключением соевых), манной крупе, пшеничной и ржаной муке.
Если говорить об овощах, то содержание крахмала наиболее высоко в корнеплодах, то есть в тех растениях, чьи съедобные части находятся под землей. Но и другие овощи также содержат некоторое количество этого углевода.
Список фруктов и овощей, богатых крахмалом :
Картофель;
— батат;
— бананы;
— топинамбур;
— хрен и имбирь;
— свекла;
— морковь;
— кабачки и патиссоны;
— помидоры;
— редька
Что касается фруктов, то они содержат, практически все, минимальное количество этого вещества..png) Исключение составляют бананы, особенно зеленые — там крахмала довольно много.
Исключение составляют бананы, особенно зеленые — там крахмала довольно много.
Продукты с низким содержанием крахмала
Минимальное содержание (около 1%) отмечено в таких растительных продуктах: Чесноке, кольраби, белокочанной капусте, грибах, болгарском перце, редисе, а также цикории, шпинате, пастернаке и других огородных растений — петрушке, укропе, листовом салате и т.д.
Чтобы проверить, есть ли в овощах крахмал и фруктах, нужно разрезать растительный продукт и капнуть на сторону разреза каплю йода. Если через минуту мякоть посинеет, этот углевод в продукте есть.
Полезные и вредные крахмалистые продукты
Легче всего переваривается крахмал из продуктов таких как рис и манная крупа. Труднее из гречки, ячневой и перловой крупы, а также из зернового хлеба и картофеля Труднее всего — из бобовых, в частности, фасоли и гороха.
Во многих продуктах питания содержится это вещество — мучных, хлебобулочных и макаронных изделиях, лепешках, сладкой выпечке, а также кашах, и киселях.
Этот углевод есть в кетчупах, майонезе, других приправах и соусах, которые мы покупаем в магазине. Очень много его в картофеле фри, чипсах, белом хлебе, булочках и выпечке.
Однако таких продуктов лучше избегать, особенно это касается тех, кто следит за весом. Дело в том, что они имеют повышенный гликемический индекс.
Крахмал, который они содержат очень быстро переваривается и усваивается, отчего происходит резкое повышение сахара. Однако его уровень также быстро спадает, от чего человек, после быстрого всплеска энергии, ощущает упадок сил.
Кроме того, частое потребление этих и подобных продуктов, негативно сказывается на общем здоровье и способствует ожирению.
А вот, цельнозерновой хлеб содержит резистентный крахмал. Он дольше и труднее переваривается, а его часть и вовсе не усваивается. Употребление такого продукта, наоборот, несколько снижает уровень сахара.
Также, «правильный» углевод содержится в фасоли и чечевице. Тут его содержание очень высоко. Несколько меньше его в цельных зернах гречки, коричневого риса, и овса.
Несколько меньше его в цельных зернах гречки, коричневого риса, и овса.
Кстати, качественные макароны, если их приготовить «аль денте», а не разваривать, также полезны и не влияют на массу тела.
Если уровень глюкозы в крови является для вас важным показателем, лучше полостью исключить из рациона продукты с повышенным гликемическим индексом. Употребляя крахмалистые овощи, знайте меру.
Зато любые фрукты (кроме бананов), овощи с умеренным и низким содержанием углеводов, а также листовую огородную зелень можно и нужно чаще включать в рацион.
Разнообразное питание, богатое витаминами, принесет только пользу вашему здоровью, уменьшит риск развития сердечно-сосудистых заболеваний. Будьте здоровы!
Мы очень часто слышим о том, что овощи обязательны для ежедневного употребления человеком. Но почему? Ведь есть многие другие продукты наделенные витаминами, но почему же нельзя упускать из внимания овощи? Все очень просто.
Именно с овощей начинается залог здорового питания:
- Овощи — не жирная пища;
- Они наделены множеством микроэлементов и витаминов;
- Некоторые овощи даже не нужно парить, жарить и варить.
 Их можно употребить свежими;
Их можно употребить свежими; - Они очень легкие и легко усваиваются организмом.
 Их можно употребить свежими;
Их можно употребить свежими;Но многие поспорят с тем, что не каждый овощ такой легкий и безобидный.
В связи с этим их делят на две категории:
- Овощи с содержанием крахмала;
- Овощи без содержания крахмала.
Крахмал – это один из видов углеводов, содержащихся в овощах. На вид это белое порошкообразное вещество, не имеющее запаха и вкуса. Данный порошок не растворим в холодной воде, поэтому, при ее слиянии образует вязкий клейстер.
Этот сложный углевод служит источником энергии, как растений, так и человека. У растений он в основном находится в клубнях и в стеблях, при распаде преобразуется в глюкозу, что питает растение энергией. В организме человека он тоже имеет свойство распадаться и превращается в сахара.
Польза и вред крахмала для организма
Крахмал в рационе человека выполняет функцию подпитки. Обогащая организм дополнительной энергией за счет расщепления и превращения в глюкозу. Данный процесс начинается с момента попадания в ротовую полость человека пищи содержащей его гранулы.
Данный процесс начинается с момента попадания в ротовую полость человека пищи содержащей его гранулы.
Не успев откусить кусок продукта, слюна начинает обволакивать каждую гранулу, на этом этапе образуется углевод под названием мальтоза.
После чего, данный углевод попадает в тонкую кишку и там под воздействием кислотной среды превращается в глюкозу, которая всасывается стенками кишечника и попадает в кровь, распространяясь по всему организму питая его энергией. Но нужно взять во внимание тот факт, что крахмал не всегда бывает полезен.
Положительные свойства крахмала:
- Благодаря содержанию большого количества вещества — амилозы, крахмал благоприятно влияет на желудок.
- Он является своего рода личным массажером стенок желудка и кишечника. При поглощении данного типа углевода человеком, он не распадается, а остается в виде комочка, который постепенно массирует стенки желудка, и налаживает систему пищеварения, уменьшая всасывания последним холестерина.
- Еще одним не менее полезным свойством рассматриваемого углевода является то, что он помогает человеку восстановить организм после резкого повышения, либо упадка сахара в организме. Это очень важно для людей, страдающих сахарным диабетом.
Отрицательные свойства крахмала:
- Люди, которые следят за своей фигурой и тщательно вычитывают уровень сахара в каждом продукте, скажут нам, что крахмалосодержащие продукты являются чуть ли не запретными. Именно он превращается в глюкозу (сахар), всасывается в стенки кишечника и в кровь и перенасыщает организм сахаром (если употребить такие продукты в избытке), что не может не сказаться на наборе веса.
- Крахмалистые соединения, обогащающие организм, иногда приводят к образованию жировых отложений. Поэтому когда вы слышите, что сбросить лишний вес можно благодаря добавлению в рацион овощей, то будь те осторожны, не все они окажутся полезными в этом деле.
Хорошим помощником в похудении будет данное средство. Компоненты конфет мягко воздействуют на организм, имеют на 100% натуральный состав, это определяет отсутствие побочных эффектов.
Компоненты конфет мягко воздействуют на организм, имеют на 100% натуральный состав, это определяет отсутствие побочных эффектов.
Конечно, особенно важно комбинировать прием таблеток с правильным питанием и физическими нагрузками. В таком случае эффективность будет максимальная, а результат заметен на лицо.
Продукты, не содержащие крахмал, либо с низким его содержанием
Необходимо отметить, что ни один продукт животного происхождения не относится к крахмалосодержащим, в то время как продукты растительного происхождения почти все являются таковыми.
Таблица продуктов с низким содержанием крахмала, либо вообще с его отсутствием, представленная ниже, включает в себя множество продуктов.
| Овощи без содержания крахмала | Овощи с низким содержанием крахмала |
| Лук, лук — порей, лук – шалот, шнитт — лук | Чеснок |
| Кервель | Тыква |
| Огурец | Горох |
| Корнишон | Артишок |
| Брюква | Кольраби |
| Портулак | Цикорий |
| Помидоры | Спаржа |
| Зелень: укроп, петрушка, щавель, шпинат, крапива, | Капуста |
| Баклажан | Зеленый и красный перец |
| Цветная капуста, красная капуста, брюссельская капуста | Редис |
| Брокколи | Пастернак |
| Одуванчик | Грибы |
| Ревень | Эндивий |
| Козлобородник | |
| Морковь |
Крахмалистые продукты очень быстро усваиваются организмом (картофель, бобовые и злаковые продукты), и в короткий срок насыщают его энергией, превращаясь в глюкозу.
Важно! Для полного усвоения крахмала требуется щелочная среда. Он тяжело усваивается, если употреблять его с продуктами, содержащими белок.
Овощи, имеющие данный углевод, хорошо усваиваются при сочетании его с пищей содержащей жиры. К ним относятся:
- Сметана;
- Растительное масло;
- Сливки и др.
Совмещая представленные жиры с овощами, содержащими низкий уровень крахмала, такими как, редиска, горох, капуста, грибы тыква и другими, организм человека насыщается различного типа микроэлементами и витаминами. Они все необходимы людям для стимулирования работы внутренних органов.
Вот как помидор и крахмал могут помочь в борьбе с морщинами
Маска с помидором прекрасно разглаживает возрастные глубокие морщины. Особенно эффективна в борьбе с проблемной кожей вокруг уголков рта и «гусиных лапок».
Крахмал – уже давно используется как домашний ботокс. Косметологи успешно применяют его в своей практике. Если постоянно применять маски на основе крахмала, то в течение короткого времени можно добиться упругости кожи, эластичности и здорового цвета кожи. Кроме этого, он абсолютно безвреден, риск нежелательных реакций почти нулевой.. А еще крахмал питает, отбеливает и насыщает кожу витаминами, нормализует работу сальных желез, вследствие чего кожа перестанет шелушиться, что исключит риск появления морщин.
Если постоянно применять маски на основе крахмала, то в течение короткого времени можно добиться упругости кожи, эластичности и здорового цвета кожи. Кроме этого, он абсолютно безвреден, риск нежелательных реакций почти нулевой.. А еще крахмал питает, отбеливает и насыщает кожу витаминами, нормализует работу сальных желез, вследствие чего кожа перестанет шелушиться, что исключит риск появления морщин.
Помидоры не менее полезны для кожи лица, их можно назвать витаминным коктейлем. В нем содержатся витамины группы В, которые повышают защитные свойства кожи, Витамин А успешно устраняет воспаления. Витамин К убирает пигментацию, а биотин регенерирует и восстанавливает кожу.
Нам понадобятся:
Помидор – 1 штука
Яичный желток – 1 штука
Картофельный крахмал – 1 чайная ложка
Оливковое масло – ½ чайной ложки
Способ приготовления маски на основе томатов и крахмала:
с помидора снимаем кожицу, разминаем мякоть с помощью вилки, добавляем яичный желток, картофельный крахмал и оливковое масло. Все ингредиенты тщательно перемешиваем. Полученную массу наносим на кожу лица, избегая попадания в глаза. Маску оставляем на 20 минут, далее смываем теплой водой и наносим свой любимый питательный крем.
Все ингредиенты тщательно перемешиваем. Полученную массу наносим на кожу лица, избегая попадания в глаза. Маску оставляем на 20 минут, далее смываем теплой водой и наносим свой любимый питательный крем.
Вся предоставленная информация носит ознакомительный характер и не может быть использована без обязательной консультации с врачом!
admin
автор статьи admin
Любое использование фото/видео/текстовых материалов без письменного разрешения редакции запрещено. Запрещено публиковать любые фрагменты материала, фотографии и видеоматериалы в пабликах ФБ, ВК, ОК Instagram.
Содержание крахмала в кабачках. Продукты богатые крахмалом
Несмотря на то, что крахмал, в небольших количествах входит в состав всех овощей, мы об этой субстанции знаем намного меньше чем должны. Поговорим о пользе и вреде крахмала, о том какие есть виды этого углевода и какие продукты питания самые крахмалистые.
Поговорим о пользе и вреде крахмала, о том какие есть виды этого углевода и какие продукты питания самые крахмалистые.
То, о чем немногие, наверное, знают, это то, что крахмал может принести пользу, а может и навредить. По умолчанию считается, что продукты содержащие крахмал имеют повышенный гликемический индекс, то есть усваиваются быстро и способствуют повышению уровня глюкозы в крови.
Однако не весь крахмал одинаков. Есть крахмал, который усваивается быстро, который усваивается медленно, и даже который не усваивается вовсе.
От чего это зависит и как определить какой из них лучше, постараюсь просто и понятно объяснить в этом материале.
Что собой представляет крахмал и его роль в питании человека
Крахмал это сложный углевод. В течение долгого времени считалось, что сложный углеводы способны меньше поднимать уровень глюкозы в крови, нежели разные виды сахаров. Однако практика показала, что некоторые виды крахмала имеют даже больший гликемический индекс чем сахар.
Людям, которые вынуждены следить за уровнем сахара в крови должны также отказаться и от продуктов содержащих крахмал, так как он очень быстро расщепляется и превращается в глюкозу.
Какой крахмал усваивается быстро, и какой медленно?
В первую очередь, давайте запомним, что крахмал содержится в том или ином количестве во всех фруктах и овощах. Просто где-то его много, а где-то очень мало.
А вот что действительно важно: Быстрота усвоения крахмала зависит от степени обработки продуктов.
Все зерновые и бобовые культуры очень богаты этим углеводом, но перемалывая, пшеницу, например, в муку, делая из нее хлеб и булочки, мы облегчаем усвоение крахмала. Таким образом, хлеб и выпечка способны поднять уровень сахара в крови за считанные минуты, даже если в рецепт не добавлялся традиционный сахар-песок.
Крахмал с самым высоким гликемическим индексом , именно тот, который содержится в белом хлебе, выпечке. И наоборот, цельнозерновой хлеб содержит меньше “быстрого крахмала”, его труднее переварить, и даже его часть не усваивается. Такой крахмал называют резистентным, и он способен снизить уровень глюкозы в крови после гипергликемии.
Такой крахмал называют резистентным, и он способен снизить уровень глюкозы в крови после гипергликемии.
Интересный факт! Один из продуктов питания который содержит крахмал, но усваивается долго, это макароны. Но не любые, а качественная паста, приготовленная «аль денте» то есть не совсем до конца. Оказывается, молекулы крахмала настолько плотно упакованы в пасте, что только половина из них усваивается моментально.
Какой крахмал самый полезный?
- Диетологи считают фасоль и чечевицу лучшими источниками «правильного» крахмала;
- На втором месте цельное зерно, такое как, например, греча, коричневый рис, киноа, амарант, овес.
- Если уровень глюкозы для вас является важным показателем, то старайтесь вообще исключить из своего рациона все продукты из белой муки.
- Также, готовые завтраки, это источник плохого крахмала.
В чем польза и вред крахмала?
Многие говорят о том, что крахмал очень полезное вещество, так как он питательный, это так и есть. Глюкоза тоже нужна организму. Однако нужно отдавать себе отчет, что каждый всплеск уровня глюкозы в крови это всегда нагрузка на сердечно-сосудистую систему.
Глюкоза тоже нужна организму. Однако нужно отдавать себе отчет, что каждый всплеск уровня глюкозы в крови это всегда нагрузка на сердечно-сосудистую систему.
Это всего лишь значит, что во всем нужно соблюдать норму. Крахмал нужен организму, так как он превращается в глюкозу, которая из крови попадает в клетки и питает их. Ненужная нагрузка на сердечно-сосудистую систему случается только когда клетки уже глюкозой перенасыщены, а уровень сахара в крови все еще высок.
Считается что резистентный крахмал очень важен для предотвращения онкологических заболеваний. Он содержится в картофеле и в свежих фруктах и овощах, в бобовых культурах.
С другой стороны, существенный вред здоровью может нанести легко-усваиваемый крахмал. Злоупотребление белым хлебом, сладкой выпечкой, может привести к очень серьезным заболеваниям, особенно это касается сердечно-сосудистой системы. Не исключены и проблемы поджелудочной железы. Ведь каждый раз, когда повышается уровень глюкозы резко, то и инсулина она вырабатывает больше. Если такие всплески постоянное явление, это не может не повысить риски для здоровья.
Если такие всплески постоянное явление, это не может не повысить риски для здоровья.
Продукты, содержащие крахмал
Как вы уже поняли, крахмал содержится во всех фруктах и овощах. Разница только в том, что где крахмала ничтожно мало, то просто считается, что его нет, так как объемы настолько незначительные, что даже сложно назвать это содержанием, правильнее будет «имеет следы крахмала».
Растительные продукты, которые содержат больше всего крахмала:
- Фасоль
- Чечевица
- Картофель
- Кукуруза
- Гречка
- Картофель батат
- Амарант
Продукты с небольшим содержанием крахмала:
- Морковь
- Свекла
- Баклажан
Некрахмалистые овощи:
- Огурцы
- Листовой салат
- Шпинат
- Болгарский перец
- Щавель
- Капуста
- Помидор
Есть ли крахмал в яблоках?
Есть. Только количество крахмала зависит от сорта и от спелости. В яблоке в котором меньше сладости, как правило, больше крахмала чем сахаров.
В яблоке в котором меньше сладости, как правило, больше крахмала чем сахаров.
Крахмал это нужный углевод, но только когда соблюдается мера в употреблении, как и во всем, впрочем. Крахмалистые продукты питания это верный способ получить незаурядную дозу энергии, именно этим крахмал и ценен. Тем не менее, рекомендую прислушаться к мнению специалистов, и ограничить, по возможности, потребление быстроусвояемого крахмала, чтобы снизить риск развития заболеваний сердечно-сосудистой системы.
Ешьте правильно и будьте здоровы!
Мы привыкли считать, что любые овощи полезны для похудения. Но отдельные источники делят всю продукцию на крахмалистые и некрахмалистые овощи. А иные диеты или советчики запрещают есть крахмалистые овощи совсем. Ну кто не слышал в наше время, что ради своей мечты придется отказаться от свеклы или морковки, забросить картошку и расстаться с бататом? Однако исключать крахмалистые овощи во всех случаях не стоит. Необходимо осмотрительно подбирать рацион для снижения веса, и учитывать собственные потребности и предпочтения.
Крахмалистые и некрахмалистые овощи и почему их делят
Традиционная диета номер восемь не содержала такого понятия, как крахмалистые овощи. Она рекомендовала лишь ограничить картофель и макароны в рационе худеющего. Сам термин появился несколько позднее. Исторически первыми его стали употреблять американские врачи-натуропаты начала 20 века. Именно они посчитали, что крахмалитстые овощи — в некоторых ситуациях чуть ли не равносильны яду. С тех пор много воды утекло, но картошку, свеклу, репу и батат обвиняют в том, что:
- они слишком калорийные, человек получает чересчур много энергии с ними;
- они содержат вредные соединения пасленовых, «закисляющие» кровь и отравляющие организм;
- некоторые из них слишком сладкие, повышают аппетит, так как организм реагирует на них слишком существенным выбросом инсулина.
В общем, крахмалистые овощи — враг худеющего человека. Постепенно этот постулат перетек и в популярные источники, и сегодня чуть ли не каждая школьница уверена, что толстеют — от картошки и свеклы, а худеют — от сельдерея и огурца.
Враг ли нам крахмал
В тех же околонатуропатских источниках указывается, что крахмал склеивает наши бедные кишечники, зашлаковывает их и вызывает у нас образование каловых камней. Что до врачей научной медицины, они так никогда и не увидели калового камня «во плоти». Зато американские натуропаты начала 20 века увидели множество исков от «залеченных» до чуть ли не полного «просветления ума» граждан. В общем-то, американцы не столько оздоровились от применения натуропатических методик, сколько приобрели стойкое неприятие к альтернативной медицине, которое и было с ними вплоть до 70 годов прошлого века.
Натуропаты, между тем, сделали большое дело — демонизировали крахмал. Но как же быть с тем фактом, что это — лишь один из видов углеводов, и он достаточно спокойно обрабатывается сначала слюной, потом — пищеварительными ферментами, а затем — попадает в кровь человека в виде глюкозы. Абсолютно тот же путь ждет углеводы из гречки и коричневого риса тоже. Вот только картошку мы обвиняем во всеобщей полноте, а рис считаем чуть ли не залогом стройности японской нации.
На самом деле, принципиальной разницы для здорового организма между картошкой и гречкой нет. Разве что, у картошки так называемый индекс сытости выше. Проблемы с крахмалом и его усвоением могут начаться у аллергиков. А вот диабетикам по общему правилу советуют только считать хлебные единицы, но не исключать обычный крахмал совсем. Таким образом, идея об исключении крахмала из здорового рациона основана на предположении.
Почему мы не должны есть только крахмалистые овощи
В список крахмалистых овощей попали картофель, свекла, топинамбур, батат, зеленый горошек, мини-кукуруза, морковка и репа. Мало того, что кукуруза — не овощи, да и зеленый горошек — скорее, бобовая культура, список достаточно полный. Проблема с крахмалом и ожирением состоит не в самом крахмале, а в переедании. Было научно доказано, что порции картошки в фаст-фуде за последнее столетие увеличились в разы. Даже если вы едите в обычном бистро, от салата со свеклой и майонезом вам не скрыться. Дело не в самом крахмале и овощах, а в огромных порциях и отсутствии знаний о сочетаемости продуктов.
Дело не в самом крахмале и овощах, а в огромных порциях и отсутствии знаний о сочетаемости продуктов.
Последние часто подменяют некоей теорией вроде раздельного питания или употребления какой-то еды отдельно от другой. Между тем, даже если человек чувствителен к инсулину, он может есть и крахмалистые овощи тоже с источниками белка и нормально себя чувствовать.
Однако если есть их отдельно велик риск переедания, так как это действительно «раскачивает» инсулиновые качели. А если жарить еще и на масле, то и риск ожирения не за горами. Наша цивилизация осуждает совсем не ту привычку. Мы можем очень спокойно есть обычные крахмалистые овощи, приготовив их на пару или сварив, и сдобрив куском мяса или рыбы. Но нам определенно не стоит заедать картошкой-фри булку с котлетой, так как первое сочетание сытное, а второе — нет.
Крахмал как углевод выполняет следующую роль в нашем питании:
- дает нам энергию, и силы;
- способствует нормальной переносимости физической нагрузки, позволяет тренироваться с полной отдачей;
- охраняет нашу нервную систему от перегрузок. Нашему мозгу требуется 140 г углеводов в сутки для нормальной работы, если мы не получаем их, вялость и усталость становятся нашими лучшими друзьями;
- помогает нам оставаться здоровыми в смысле репродуктивной системы. Гипоталамическая аменорея и нарушения цикла связываются современной медициной не только с туманными «нарушениями метаболизма», но и с вполне конкретными вещами вроде дефицита углеводов в рационе;
- способствует сжиганию жира в том смысле, что позволяет поддерживать здоровую секрецию гормонов щитовидной железы, а ведь именно они важны для скорости наших метаболических процессов;
- позволяет удешевить питание. Крахмалистые корнеплоды — недорогие источники энергии почти во всех странах.
 Нашему мозгу требуется 140 г углеводов в сутки для нормальной работы, если мы не получаем их, вялость и усталость становятся нашими лучшими друзьями;
Нашему мозгу требуется 140 г углеводов в сутки для нормальной работы, если мы не получаем их, вялость и усталость становятся нашими лучшими друзьями;В общем, мы не должны ограничиваться только зерновыми, когда дело доходит до пополнения энергетических запасов, и вполне можем поесть и корнеплоды тоже.
Польза некрахмалистых овощей для похудения
О пользе некрахмалистых овощей знают все. К некрахмалистым овощам относятся огурцы, кабачки, цуккини, вся зелень, тыква, также все виды капусты, помидоры и болгарский перец. Некрахмалистые овощи помогают нам:
К некрахмалистым овощам относятся огурцы, кабачки, цуккини, вся зелень, тыква, также все виды капусты, помидоры и болгарский перец. Некрахмалистые овощи помогают нам:
- получить нужное количество клетчатки. Для нормального пищеварения человеку необходимы 20-25 г клетчатки;
- получить необходимую организму воду;
- удовлетворить потребность в витаминах и минералах;
- насытиться быстрее;
- сохранять здоровое пищеварение даже на низкокалорийной диете;
- удерживать здоровый вес в любом возрасте.
Овощи помогают нам питаться вкусно и разнообразно, они полезны для здоровья и мы просто обязаны съедать порцию овощей с каждым основным приемом пищи. А еще вы можете не считать калории из некрахмалистых овощей, все равно их ничтожно мало там, настолько, что в диете, не ориентированной на подготовку бодибилдера к сцене ими можно пренебречь.
Итак, мы должны есть и крахмалистые и некрахмалистые овощи. Почти любая диета для снижения веса должна содержать 4-5 порций некрахмалистых и пару порций крахмалистых овощей в сутки. Стремитесь к разнообразию овощного стола и вы сохраните здоровье и красоту.
Стремитесь к разнообразию овощного стола и вы сохраните здоровье и красоту.
Современные принципы правильного питания предполагают, что каждый сознательный человек должен знать, чем он питается. Одним из основных компонентов нашего повседневного рациона является крахмал, а его избыток или недостаток способны создать настоящие проблемы со здоровьем, потому попробуем разобраться, что такое крахмал, зачем он нужен, где он есть, а где его нет.
Значение крахмала для организма человека
Питание человека должно быть сбалансированным в плане пополнения организма белками, углеводами и жирами. Именно углеводы считаются основным источником энергии в организме, особенно глюкоза, которая расщепляется довольно легко и с большим выделением тепла. Сама глюкоза, кстати, в чистом виде в продуктах встречается довольно редко, а проще всего организму ее получать именно из крахмала, тем более что тот содержится в огромном количестве продуктов питания.
Следовательно, первое свойство, ради которого крахмалосодержащие продукты стоит употреблять активнее – это подпитка организма энергией. Но на этом польза пищи, содержащей крахмал, не заканчивается. Ведь такое вещество полезно для полезных бактерий в кишечнике и повышения иммунитета, а также помогает наладить выработку желудочного сока и нормализовать уровень сахара в крови.
Но на этом польза пищи, содержащей крахмал, не заканчивается. Ведь такое вещество полезно для полезных бактерий в кишечнике и повышения иммунитета, а также помогает наладить выработку желудочного сока и нормализовать уровень сахара в крови.
Впрочем, регулировать количество крахмала в рационе иногда стоит и для того, чтобы ограничивать его количество. Так, избыток крахмала при малоподвижном способе жизни гарантированно приведет к набору веса, а в некоторых случаях этот компонент провоцирует такие побочные действия, как метеоризм или различные нарушения деятельности желудочно-кишечного тракта. По этой причине диетологи после постановки некоторых диагнозов советуют пациенту уменьшить количество крахмалистых овощей и фруктов в своем меню, для чего их необходимо знать.
Следует обратить внимание еще и на тот факт, что крахмал бывает натуральным и рафинированным. Первый, как это часто бывает с натуральными продуктами, не так уж и вреден – он присутствует преимущественно в корнеплодах, кашах и некоторых овощах. При таком питании набор веса вероятен лишь при гигантских порциях или совершенной неподвижности, так что ограничения обычно не накладываются. Другое дело – добавки на основе рафинированного крахмала, поскольку они очень калорийны и быстро насыщают, но согнать спровоцированный такой едой лишний вес очень проблематично. Ситуацию усугубляет еще и то, что подобные добавки (например, загустители) могут присутствовать в самых неожиданных продуктах, где крахмалу, казалось бы, не место.
Где этого вещества много?
Полный список крахмалистых продуктов питания составить весьма проблематично – как раз из-за тех добавок, которые могут присутствовать фактически где угодно. По этой причине просто рассмотрим только те виды еды, которые содержат много крахмала без всяких добавок.
- Злаки. Согласно народной поговорке, физически слабый человек «мало каши ел», а все потому, что именно в таком продукте процент содержания крахмала максимальный. В среднем содержание этого вещества здесь составляет около 70-75%, что очень много. Среди популярных разновидностей пищи из этой категории исключений особо не наблюдается. Утверждение о крахмалистости злаков справедливо для пшеницы и кукурузы, риса и овса, каш и муки из всех этих злаков, хлебобулочной и макаронной продукции, даже для гороха и бобов.
 В среднем содержание этого вещества здесь составляет около 70-75%, что очень много. Среди популярных разновидностей пищи из этой категории исключений особо не наблюдается. Утверждение о крахмалистости злаков справедливо для пшеницы и кукурузы, риса и овса, каш и муки из всех этих злаков, хлебобулочной и макаронной продукции, даже для гороха и бобов.
В среднем содержание этого вещества здесь составляет около 70-75%, что очень много. Среди популярных разновидностей пищи из этой категории исключений особо не наблюдается. Утверждение о крахмалистости злаков справедливо для пшеницы и кукурузы, риса и овса, каш и муки из всех этих злаков, хлебобулочной и макаронной продукции, даже для гороха и бобов.Единственным исключением являются соевые продукты.
- Корнеплоды и некоторые другие овощи. Плоды огородничества, особенно растущие под землей, тоже зачастую богаты крахмалом, хоть и не так радикально, как злаки. Особо выделяется здесь чеснок, где крахмала целых 26%, а из того, что люди едят массово и в больших количествах – картофель (15-18%). Даже растущие на поверхности помидоры могут стать источником крахмала, хоть его здесь сравнительно немного – около 5%.
- Фрукты. Большинство фруктов в свежем виде содержат крахмала совсем немного, а практически единственным исключением следует считать свежие бананы. Другое дело, что у такой пищи основную долю веса занимает вода, а потому, высушив фрукт, можно в несколько раз повысить концентрацию рассматриваемого вещества. По этой причине сухофрукты, особенно яблочные, грушевые и абрикосовые считаются очень калорийными и противопоказанными тем, кто имеет проблемы с лишним весом.
 Другое дело, что у такой пищи основную долю веса занимает вода, а потому, высушив фрукт, можно в несколько раз повысить концентрацию рассматриваемого вещества. По этой причине сухофрукты, особенно яблочные, грушевые и абрикосовые считаются очень калорийными и противопоказанными тем, кто имеет проблемы с лишним весом.
Другое дело, что у такой пищи основную долю веса занимает вода, а потому, высушив фрукт, можно в несколько раз повысить концентрацию рассматриваемого вещества. По этой причине сухофрукты, особенно яблочные, грушевые и абрикосовые считаются очень калорийными и противопоказанными тем, кто имеет проблемы с лишним весом.
Продукты без крахмала
Если диета требует сильно уменьшить количество потребляемого крахмала, то следует отказаться от большинства готовой магазинной продукции – там этот ингредиент наверняка присутствует в виде той или иной добавки. Наверняка придется отказаться от каш и выпечки, равно как и от макарон, а также от многих соусов. Впрочем, вряд ли хоть один диетолог посоветует отказаться от крахмала полностью – он ведь представляет определенную пользу для организма. Задача пациента – просто несколько снизить его поступление, так что при правильно составленной диете можно даже не отказывать себе в небольшом количестве выпечки.
Так, к некрахмалистым диетическим продуктам относятся, например, грибы, но основная потребность организма в пище будет восполняться за счет различных овощей. Список доступных вариантов не так уж ограничен: баклажаны и брокколи, обычная, брюссельская и пекинская капуста, зеленый горошек и тыква, огурцы и сладкий перец. Все эти компоненты позволят не только приготовить вкусный салат без лишних полисахаридов, но и побаловать себя более изысканными блюдами вроде овощного рагу или даже сладкой тыквенной каши.
На этом список доступных компонентов не заканчивается, дальше идут «приправы» к основной еде: шпинат и щавель, чеснок и цикорий, зелень сельдерея и петрушка.
Среди фруктов тоже есть варианты, как полакомиться десертом и не превысить нормальную дозировку крахмала. Из круглогодичных фруктов наиболее доступны яблоки, но не все. Диетологи советуют выбирать зеленые и твердые плоды, поскольку в них полисахаридов меньше. Остальные некрахмалистые фрукты скорее относятся к сезонным, но их сезоны не совпадают между собой, потому разнообразие в меню можно вносить круглый год благодаря клубнике, дыням и нектаринам. Из импортных, но популярных в нашей стране фруктов с низким содержанием крахмала можно отметить экзотический авокадо.
Остальные некрахмалистые фрукты скорее относятся к сезонным, но их сезоны не совпадают между собой, потому разнообразие в меню можно вносить круглый год благодаря клубнике, дыням и нектаринам. Из импортных, но популярных в нашей стране фруктов с низким содержанием крахмала можно отметить экзотический авокадо.
О том, что скажет диетолог про углеводы из некрахмалистых овощей, смотрите в следующем видео.
Для многих диет, которые включают в свой состав овощи или овощные блюди, необходимо знать, какие овощи максимально полезны при похудении , а какие овощи могут мешать избавлению от лишних килограммов. Много лет назад ученые пришли к выводу, что не все овощи одинаково полезны для похудения, несмотря на то, что все из них содержат витамины и полезные вещества. Чтобы похудение шло быстрее необходимо избегать крахмалистых овощей , то есть тех плодов, в которых слишком высоко содержание крахмала.
Быстрая навигация по статье:
Что такое крахмал
Совсем без крахмала нашему организму не обойтись. Попадание крахмала в организм человека способствует улучшению работы мозга, мышц , а также это вещество является основным источником питания для тела человека.
Крахмал – сложный углевод, который, попадая в организм, повышает уровень сахар в крови. К тому же, блюда богатые крахмалом способны усваиваться организмом очень быстро , вызывая через короткий промежуток времени новый приступ голода. А это особенно вредно для фигуры. Поэтому, если вы сидите на овощной диете, диете Кима Протасова и некоторых других видах овощных диет, обязательно изучите список крахмалистых и некрахмалистых овощей. Старайтесь максимально ограничить потребление крахмалистых овощей , чтобы ускорить процесс похудения.
В каких овощах содержится крахмал
В последнее время производилось много исследований продуктов на содержание крахмала. Изначально специалисты заговорили о крахмалистых и некрахмалистых овощах во время создания правил раздельного питания. В те времена, мало кто придавал этому значение. Более глубокие исследования подтвердили негативное влияние крахмала, содержащегося в овощах, на фигуру.
Более глубокие исследования подтвердили негативное влияние крахмала, содержащегося в овощах, на фигуру.
1 Разумеется, максимальное содержание крахмала содержится в картофеле. Даже в небольшом клубне может содержаться огромное количество крахмала. В некоторых корнеплодах пятую часть от объема всей картофелины составляет крахмал. Именно поэтому диетологи всего мира сошлись во мнении, что худеющим необходимо первым дело отказаться от картофеля. Особенно вреден картофель в жареном виде , а также картофель фри. Картофель, приготовленный таким способом, напитывается жиром, в котором он жарился. Помимо крахмала в нем также содержатся канцерогены, полученные путем жарки на подсолнечном масле. Такой вредный жир может вызывать не только ожирение, но и заболевания желудочно-кишечного тракта, раковые заболевания, и многое другое.
2
Еще одним крахмалистым овощем считается «царица полей» кукуруза . В ней содержится большое количество углеводов, и она имеет довольно большую калорийность в сравнении со многими другими овощами: 93 кКал на 100 г продукта. В вареной и консервированной кукурузе килокалорий еще больше. Поэтому прежде, чем баловать себя кукурузкой в составе салатов, или просто в отварном виде, необходимо взвесить все за и против для ваших форм.
В вареной и консервированной кукурузе килокалорий еще больше. Поэтому прежде, чем баловать себя кукурузкой в составе салатов, или просто в отварном виде, необходимо взвесить все за и против для ваших форм.
3 Бобовые культурные растения также считаются богатыми крахмалом продуктами. К таким бобовым растениям относятся фасоль, горох, нут, чечевица, соя и др. Бобовые продукты зеленого цвета содержат достаточное количество аскорбиновой кислоты, каротина, витамина В, минеральных веществ, однако излишнее употребление этих блюд может привести к набору веса.
4 Топинамбур считается отличным заменителем привычного всем картофеля. Причем по утверждению ученых, в этом корнеплоде содержится намного больше питательных веществ, чем в картофеле. Ему не страшны вредители, и он менее прихотлив в выращивании. Также по питательности топинамбур превосходит свеклу практически в два раза. Однако большое употребление этого крахмалосодержащего продукта может спровоцировать усиленное газообразование и дискомфорт в кишечнике.
5 Батат – еще один «заменитель» картофеля . Его также называют сладким картофелем, так как он очень поход по виду на картофель, а по вкусу напоминает сладкую тыкву. Его калорийность составляет 61 кКал на 100 г продукта. Помимо этого в корнеплоде содержится много воды, органических кислот, каротина.
6 Свекла – овощ, который весьма популярен на территории нашей страны. Его преимущество перед многим другими овощами в том, что при термической обработке он практически не теряет полезных веществ. Однако, для тех, кто следит за своим весом, употребление свеклы в пищу должно быть умеренным, так как она также является источником легкоусвояемого крахмала . Также свекла противопоказана людям, страдающим сахарным диабетом, мочекаменной болезнью, расстройством кишечника, гастритом.
7 Редька не слишком богата крахмалом, однако, все же содержит его. Редьку можно есть во время овощных диет, но в умеренных количествах. Хотя стоит отметить, что, скорее всего, съесть слишком много редьки вам не удастся, из-за ее специфического вкуса. Редьку рекомендуется есть в составе салатов , а не отдельно.
8 Морковь является очень полезным овощем, который многие ученые относят к крахмалистым овощам. Некоторые другие специалисты считают, что ее можно отнести к умеренно крахмалистым продуктам . В любом случае не стоит усердствовать с морковью. Морковь должна быть полностью исключена из рациона при наличии у человека аллергии. А также любому, даже абсолютно здоровому человеку, противопоказано есть сразу большое количество моркови. Это может спровоцировать «каротиновую желтуху» с появлением желтых пятен. Лечение такой желтухи простое – полное исключение моркови из рациона на длительный срок.
9 Кабачки , так же как и морковь, считаются многими специалистами по питанию крахмалистыми либо умеренно крахмалистыми. Во время овощной диеты необходимо обязательно включать к меню кабачок, так как его нежная мякоть способствует улучшению пищеварения, цвета лица и прочего. Однако, есть кабачок следует умеренно, чтобы избежать проблем с лишним весом.
10 Тыква – сладкий и полезный овощ , который положительно влияет на пищеварение, работу кишечника, является великолепным поставщиком витаминов и важных микроэлементов в организм человека. Если вы сидите на овощной диете, то не стоит есть тыкву как самостоятельное блюдо. Можно позволять себе иногда немного тыквенного сока. Совершенно противопоказана худеющим рисовая каша с тыквой , поскольку и тыква и рис содержат огромное количество крахмала, что может негативно отразиться на фигуре.
В каких продуктах содержится крахмал. Таблица
В каких других продуктах содержится крахмал
Есть и другие продукты, в которых содержание крахмала намного выше, чем в самых крахмалистых овощах. Перед тем, как выбрать приемлемый для себя рацион питания, следует выяснить, какие продукты, помимо овощей богаты крахмалом и будут препятствовать наискорейшему похудению.
Среди других продуктов, которые содержат огромное количество крахмала, можно выделить рис (80-83%), ячмень(72%) и пшеницу (67%). Рожь (62%) и просо (56%) также богаты этим веществом. Поэтому худеющим не советуют увлекаться кашами и крупами, варить супы с добавлением риса, есть хлебобулочные изделия.
Все об углеводах или как правильно питаться:
Крахмал, превращающийся в ходе переваривания в глюкозу, относится к наиболее распространенной в природе форме полисахаридов. Поэтому деление овощей на крахмалистые и некрахмалистые, изначально являвшееся частью теории , нашло место в универсальной . Некрахмалистые овощи — настоящие помощники в похудении, а вот с крахмалистыми стоит держать ухо востро! Но как не перепутать? Поможет наш справочный сервис с удобными таблицами.
Овощное меню — не синоним меню для похудения! Овощи бывают разные, и, соответственно, различаются правила их сочетания друг с другом и с другими продуктами
Наибольшим процентом содержания крахмала отличаются корнеплоды и крупные зерна, накапливающие питательные вещества для продолжения роста и обеспечения «продовольственным запасом» зародыша растения. Самым «выраженно крахмальным» овощем, бесспорно, является картофель — крахмал может представлять до 1/5 части объема клубня! Именно поэтому тем, кто хочет похудеть, в первую очередь отказаться от картошки.
Крахмалистые овощи: полный список
| Брюква |
| Кукуруза |
| Морковь |
| Свекла |
| Зрелые (сухие) бобы, кроме соевых |
| Зрелый (сухой) горох |
| Кабачки |
| Патиссоны |
| Картофель (все виды и сорта, включая сладкий) |
| Каштаны |
| Корни растений, пригодных к употреблению в пищу (хрен, петрушка, пастернак, сельдерей) |
| Тыква (круглая, осеннего созревания) |
| Топинамбур |
| Редька |
| Редис |
Умеренно крахмалистые овощи: два варианта списка
Некрахмалистые овощи: полный список
| Баклажан |
| Брокколи |
| Брюссельская капуста |
| Горчица |
| Зеленый горошек |
| Пекинская (китайская) капуста |
| Кольраби |
| Кочанная капуста (белокочанная, краснокочанная, савойская, огородная, кормовая) |
| Кресс-салат и водяной кресс |
| Спаржа |
| Летняя тыква (продолговатая желтая) |
| Латук и другие виды листового салата |
| Ботва репы и другие наземные зеленые части съедобных растений |
| Свекольные листья и листовая свекла (мангольд) |
| Лук (репчатый, шалот, порей, шнитт-лук, порей) |
| Огурец |
| Зелень одуванчика |
| Окра |
| Бамия |
| Петрушка (зелень) и другие столовые травы |
| Побеги бамбука |
| Сурепка (зелень) |
| Сельдерей (зелень) |
| Сладкий перец |
| Цикорий |
| Чеснок (зелень и зубчики) |
| Шпинат |
| Щавель |
Овощи, не содержащие крахмала
Есть ли в помидорах крахмал. Значение крахмала для организма человека
Деление овощей по количеству крахмала
Различные овощные культуры значительно отличаются по содержанию крахмала. Например, 100 грамм картофеля (относится к группе овощей, богатых крахмалом) содержит до 19 грамм крахмала и 90 калорий, а брокколи (некрахмалистый овощ) всего до 6-7 грамм крахмала и 28 калорий (на 100 грамм продукта).
К списку овощей с высоким процентным содержанием крахмала относятся:
Овощами с низким уровнем крахмала являются:
- кабачки;
- спаржа;
- брюссельская капуста;
- брокколи;
- цветная капуста;
- капуста белокочанная;
- сельдерей;
- огурцы;
- баклажаны все сортов;
- лук;
- помидоры;
- шпинат;
- репка.
Учёные рекомендуют употреблять не менее 450 грамм овощей (любых) в день, чтобы полностью обеспечивает потребности организма в пищевых волокнах, витаминах и прочих ценных питательных веществах.
Все овощи богаты питательными веществами
Крахмалистые и некрахмалистые овощи содержат практически все группы питательных веществ, необходимых для полноценной работы организма человека.
Овощи богаты калием, витамином К, фолиевой кислотной и магнием. Данные биологически активные вещества и макроэлементы обладают внушительным набором полезных свойств:
- Фолиевая кислота, по данным американских учёных, необходима для поддержания метаболизма (синтеза аминокислот, регуляции высвобождения ферментов и усвоения прочих витаминов), а также для роста и развития плода во время беременности. Основной источник – бобовые культуры и листовые зелёные овощи.
- Витамин К нужен для поддержания минеральной плотности костей, улучшения чувствительности периферических тканей к инсулину. Витамин К также обладает противоопухолевой активностью. Получить вещество, как утверждают эксперты из Индии, можно из кинзы, листовой зелени, лука и капусты брокколи.
- Магний обеспечивает репликацию ДНК и РНК, регулирует высвобождение АТФ. Также он необходим для регуляции работы гладкой и поперечно-полосатой мускулатуры, проведения импульсов по нервным волокнам. При недостатке наблюдаются расстройства со стороны опорно-двигательной, периферической нервной и сердечно-сосудистой систем. Подобные сведения приводят специалисты Академии микронутриентной медицины (США). Наибольшее количество магния имеется в бобовых культурах и луке.
При употреблении рекомендуемой суточной дозы овощей, можно также полностью удовлетворить потребности организма в железе и цинке.
Важная особенность овощей – содержание большого количества антиоксидантных веществ (витамины С и Е), которые защищают клетки организма от негативного воздействия свободных радикалов и влияния окислительного стресса. При их отсутствии в рационе наблюдается изменение структуры ДНК, белков и липидов, что приводит к множеству заболеваний, в том числе смертельно опасных (рак). Антиоксиданты, по данным американских учёных, — один из главных способов профилактики злокачественной трансформации клеток организма.
Все овощи, при систематическом употреблении, замедляют старение за счёт воздействия на основные молекулярные механизмы данного процесса.
Овощные культуры содержат (в большинстве случаев) низкие объёмы глюкозы, жиров и ионов натрия, что делает их крайне полезным видом пищи.
Крахмалистые овощи содержат больше углеводов и калорий
Крахмалистые овощи отличаются высоким содержанием углеводов и большей калорийностью. Подобная особенность является предметом научных споров относительно безопасности приема овощей, богатых крахмалом, особенно при отягощённом эндокринном фоне.
Углеводы
Содержание углеводов в крахмалистых овощах примерно в 3-4 раза выше по сравнению с некрахмалистыми и составляет около 13-26 грамм на 100 грамм продукта.
Такие овощи немного быстрее повышают уровень глюкозы в крови после употребления, следовательно, их рекомендуются ограничивать при патологиях гликемического фона (сахарный диабет II типа, гипергликемия натощак, нарушение толерантности к глюкозе). Предпочтительная доза таких овощей для больных, страдающих сахарным диабетом – до 180 грамм в день.
Калории
Высокая энергетическая ценность крахмалистых овощей обусловлена повышенным содержанием углеводов. В среднем, их калорийность в 3-6 раз выше, чем у овощей с низким содержанием крахмала.
Крахмалистые овощи содержат от 70 до 175 калорий на 100 грамм, а некрахмалистые – до 35 калорий. Это важно для лиц, страдающих ожирением или находящихся на спортивной диете.
Овес
Употребление овса – один из самых удобных способов включить резистентный крахмал в свой рацион.
В 100 граммах приготовленных овсяных хлопьев может содержаться около 3,6 грамма резистентного крахмала. Цельнозерновой овес также богат антиоксидантами (11).
Дайте приготовленному овсу остыть в течение нескольких часов (или в течение ночи) – это позволит еще больше увеличить количество резистентного крахмала.
Резюме:
Овес является хорошим источником резистентного крахмала – около 3,6 грамма на 100 граммов приготовленных овсяных хлопьев.
Крахмалистые овощи богаты устойчивым крахмалом и белками
Ещё одна отличительная особенность крахмалистых овощей – высокий уровень устойчивого (резистентного) крахмала и ценных белков.
Устойчивый крахмал
Резистентный крахмал, как утверждают китайские учёные, содержится в больших количествах в картофеле и прочих корнеплодах, а также в большинстве бобов.
Основными достоинствами устойчивого крахмала являются:
- Улучшение здоровья пищеварительного аппарата.Доказано, что пищевые волокна улучшают состав микрофлоры, полезны при наличии инфекционных и аутоиммунных патологий кишечника, угнетают хроническое воспаление в стенке кишки и повышают эффектность ферментативного расщепления пищи.
- Помощь в похудении. Неперевариваемый крахмал активирует выработку гормонов сытости (пептид YY, глюканоподобный пептид, лептин), которые, воздействуя на головной мозг, приводят к появлению чувства сытости и насыщения организма.
- Улучшение всасывания питательных веществ. Данный вид пищевых волокон повышает биологическую доступность кальция, фосфора, железа, цинка и магния. Эффекты доказаны учёными из США.
- Снижение общего холестерина в крови.По информации испанских экспертов, устойчивая клетчатка ингибирует синтез холестерина, воздействуя на активность 9 генов, отвечающих за данный процесс.
- Снижение глюкозы в крови.Доказано, что бутират, который образуется в ходе взаимодействия кишечной микрофлоры с резистентным крахмалом, повышает чувствительность периферических тканей к инсулину, приводят к повышению утилизации глюкозы из крови.
- Укрепление иммунитета. Пищевые волокна нормализуют работу местных факторов иммунной системы в просвете пищеварительного аппарата, обеспечивают профилактику аутоиммунных и аллергических заболеваний.
- Улучшение зрения. Американские учёные доказали, что устойчивые пищевые волокна повышают плотность расположения фоторецепторов на сетчатке.
Белки
Большинство крахмалистых овощей (особенно бобовые культуры и чечевица) – важные источники белка. 100 грамм продукта обеспечивает организм 10 граммами белка, что составляет около 20% от суточной потребности.
Работники Университета МакМастер (Канада) утверждают, что белки способствуют быстрому развитию ощущения сытости, позволяют контролировать уровень аппетита и массу тела, позволяют поддерживать тонус и работоспособность скелетной мускулатуры.
Вареный и охлажденный рис
Рис – еще один недорогой продукт с высоким содержанием резистентного крахмала.
Один из популярных способов приготовления – приготовление большой кастрюли риса, которой хватит на несколько дней.
Это не только экономит время, но и увеличивает содержание резистентного крахмала, когда рис со временем остывает.
Коричневый рис может быть предпочтительнее белого риса из-за более высокого уровня содержания клетчатки. Коричневый рис также содержит больше микроэлементов, таких как фосфор и магний (12, 13).
Резюме:
Рис – хороший источник резистентного крахмала, особенно когда его оставляют остывать после приготовления.
Овощи, не содержащие крахмала.
Всеми любимые помидоры, из которых состоит хорошая томатная диета, не относятся ни к крахмалистым, ни к некрахмалистым овощам. Было выявлено, что основной пищевой характеристикой помидора является кислота, а не наличие крахмала. Ввиду большого содержания кислот (цитрусовая, яблочная и щавелевая) в составе помидор относят к кислым продуктам, и их не рекомендуется кушать с крахмалистыми овощами, однако разрешено с листовыми овощами и жирами. Калькулятор калорий онлайн.
Крахмалистые овощи содержат в своем составе крахмал, превращающийся в ходе переваривания в глюкозу, относится к наиболее распространенной в природе форме полисахаридов.
Поэтому деление овощей на крахмалистые и некрахмалистые, изначально являвшееся частью теории раздельного питания, нашло место в универсальной здоровом питании.
Некрахмалистые овощи — настоящие помощники в похудении, а вот с крахмалистыми стоит держать ухо востро!
Но как не перепутать? Вам поможет таблица со списком, выбрать для себя наиболее полезные овощи
Продукты, содержащие крахмал являются указанием, много ли в вашем рационе быстрых углеводов?
Как вы знаете, излишек в рационе быстрых углеводов способствуют набору массы тела, и если вы будете обращать пристальное внимание на их количество, тогда вы сможете регулировать калорийность своего рациона.
При похудении, всегда рекомендуется кушать овощи с низким содержанием крахмала, а как узнать крахмалистые овощи или не крахмалистые?
Некрахмалистые овощи помидор. Овощи зеленые и некрахмалистые — основа раздельного питания
В отличие от крахмалистых продуктов, сочетаемых с ограниченным перечнем, некрахмалистые продукты могут сочетаться почти со всеми продуктами. Специалисты рекомендуют всем есть эти продукты вместе с любыми другими, кроме молочных продуктов и молока.
Шелтоном выделяются промежуточные продукты. Такое название он дал бобовым (иначе говоря — зернобобовым): чечевице, гороху, фасоли, сое. В них содержится много растительных белков и крахмала. Нужно признать, что бобовые — тяжелая пища для желудка. В сое содержится крахмала меньше всего.
Зеленые листовые овощи относятся также к овощам без крахмала. Группа таких овощей весьма велика, вы можете из нее выбрать подходящие именно вам, на собственный вкус, и есть их с другими продуктами.
- Употребление в пищу большого количества овощей и фруктов — крахмалистых и некрахмалистых.
- Употребление в сутки не менее, чем 4 наименований овощей и фруктов.
- Употребление основной части углеводов только в сыром виде (норма в сутки — 400г).
Однако употребление в большом количестве сырых овощей иногда приводит к проблемам, связанным с кишечником и желудком. Они могут сопровождаться вздутием и даже болями в области живота. В таких случаях овощи нужно употреблять в пищу в тушеном, вареном виде, или готовить их на пару. Из некрахмалистых овощей хорошо готовить салаты для похудения.
Нужно помнить, что каждый выбирает сам, какие для него ближе принципы питания, однако не стоит забывать, что, в первую очередь, главное — не нанести вред своему организму. Поэтому специалисты-диетологи и рекомендуют иметь в своем рационе некрахмалистые и крахмалистые овощи. Так вы сможете сбалансировать питание и не навредите здоровью.
С чем употреблять крахмалистые овощи?
Крахмалистые овощи рекомендуется если и употреблять, то только в сочетании с зелеными некрахмалистыми овощами, жирами (как растительными, так и животными), избегая гастрономического тандема с белками, сахаром и кислотой.
Действует правило: один прием пищи — один вид крахмалистых овощей.
Некрахмалистые овощи — основа меню при снижении веса!
Продукты из этого списка прекрасно сочетаются со всеми видами крахмалистой и белковой пищи, отлично усваиваясь и не создавая никаких проблем ни желудочно-кишечному тракту, ни талии.
Молоко — единственный нежелательный спутник для некрахмалистых овощей, причем следует избегать не только обычного цельного, но и соусов на его основе (таких, как, например, бешамель).
Цветную капусту основоположник раздельного питания Герберт Шелтон относил к умеренно крахмалистым овощам, с одной стороны, по сочетаемости и диетическим свойствам относя ее к некрахмалистым собратьям, а с другой — отмечая, что, тем не менее, употреблять в пищу цветной капусты следует немного, и лучше всего вместе с жирами.
Последователи Шелтона расширили список умеренно крахмалистых овощей, добавив туда новых участников, которые хоть и содержат крахмал, но не в криминальных количествах.
Обратите внимание на то, что баклажан в разных трактовках также может относиться и к некрахмалистым, и к умеренно крахмалистым овощем. (см. таблицы ниже).
Обожаемые многими помидоры, на которых стоит популярная томатная диета, не нашли места ни в числе крахмалистых, ни среди некрахмалистых овощей, хотя в распространенном представлении это самый что ни на есть салатный овощ (правда, с ботанической точки зрения помидор – ягода).
Шелтон установил, что главное в пищевой характеристике помидора — его кислота, а не содержание крахмала.
Из-за высокого процента цитрусовой, яблочной и щавелевой кислот в составе краснощекие томаты относятся к кислой пище, и их нельзя есть ни с чем крахмалистым, зато можно — с листовыми овощами и жирами.
Пользуясь списками, вы сможете без труда ориентироваться в многообразии овощей, выбирая эффективные продукты для похудения и поддержания формы.
Но мне хочется сказать несколько слов в защиту крахмала
Спелые помидоры: состав, калорийность и витамины.
Предлагаю ознакомиться с полезными свойствами спелого помидора, на основании его химического состава. Рассматриваем спелый красный и сырой помидор. В консервированных, желтых и недозрелых все эти показатели совершенно иные.
Не смотря на то, что помидоры не обладают богатым витаминным и минеральным составом — их можно считать хорошим источником всяческих «полезностей», ведь зачастую мы кушаем их в салатах, то есть в сыром виде, а значит большинство витаминов них сохранены.
ПИТАТЕЛЬНЫЙ СОСТАВ ПОМИДОРА, 100 ГРАММ:
Углеводы, грамм3,9
В их составе: клетчатки 1,2 гр., крахмала 0 гр., сахары 2,6 гр.
Помидоры не содержат крахмала, но и клетчатки в них не много. По сему их нельзя рассматривать как хороший источник пищевых волокон. Сахара в томатах так же не много, примером даже в репчатом луке, сахара больше почти в два раза.
В их составе: жирные кислоты Omega-3 3 мг., Omega-6 80 мг.
Естественно жира в помидорах фактически нет. Поэтому приведенные показатели скорее статистические.
Белки, грамм0,9
Вода, грамм94,5
Традиционно водянистыми принято считать огурцы, но по факту помидоры не уступают им в водянистости.
Калорийность спелого помидора, ккал18
Помидоры очень низкокалорийные (один из самых низких показателей энергетической ценности среди овощей), поэтому они рекомендованны в диетах для похудения.
ВИТАМИНЫ И МИНЕРАЛЫ В СВЕЖИХ ПОМИДОРАХ, 100 ГРАММ:
Далее посмотрим витаминно-минеральный состав и оценим его в соотношении с суточной потребностью взрослого человека.
Наиболее значемые цифры в составе помидора: количество калия, меди, витаминов А и С.
КАЛИЙ В СВЕЖИХ ПОМИДОРАХ:
Конечно помидоры не могут похвастать рекордным содержанием калия в своём составе (лидеры здесь: картофель, банан и киви), но в отличии от вышеперечисленных они обладают крайне низкой калорийностью. Так что если вы заботитесь о своём «экстерьере», считайте помидоры, и огурцы кстати тоже, лучшим источником калия.
МЕДЬ В ПОМИДОРАХ:
Таким содержанием меди могут похвастать многие продукты: бананы, картофель, оливки, а примером в креветках, меди больше в три раза. Но! В отличии от помидоров все они содержат много больше калорий и в большинстве своём это продукты гораздо более дорогие.
ВИТАМИНЫ В ПОМИДОРАХ:
Как видете витаминный состав помидора довольно обширный: А, Е, С, В5, В6, В9, РР, К. И хотя ни в одной из категорий помидоры не лидируют, повторюсь еще раз: вы получаете этот витаминный комплекс не потребляя лишних калорий, что очень и очень хорошо.
Чем полезены свежие помидоры, вкратце: отличный низкокалорийный источник калия и меди, хороший источник витаминов А и С.
✎ Здесь продолжение этой рубрики: Состав и польза различных продуктов →
Домашняя диета | 2011 — 2018 | © Пожалуйста при использовании материалов этого сайта укажите источник гиперссылкой. | Карта сайта
12 продуктов, которые на самом деле нельзя хранить в холодильнике :: Здоровье :: РБК Стиль
© Rawpixel/Unsplash
Автор Ирина Рудевич
16 октября 2018
Чтобы сохранить продукты свежими как можно дольше, мы кладем их в холодильник. И часто совершаем ошибку — не все в холоде хранится лучше.
Помидоры
Из-за холодного воздуха в холодильнике помидоры теряют запах и вкусовые качества. Меняется структура овоща — помидор становится рыхлым в результате разрушения клеточных мембран. Чтобы томаты не потеряли вкус, цвет и запах, хранить их нужно при комнатной температуре.
Картофель
Для хранения картофеля лучше выбрать темное прохладное место на кухне. В холоде картофельный крахмал преобразуется в сахар. Если клубни лежат при температуре ниже +3 градусов, то появившаяся в них глюкоза добавит картошке излишне сладкий вкус и не позволит сохранить форму при термообработке. К тому же при варке в замерзших клубнях появятся канцерогены.
Авокадо
Особенность хранения авокадо зависит от степени зрелости плода. Спелый авокадо отличается коричневым оттенком, при этом основание черенка остается зеленым. Если оно уже темное, значит фрукт перезрелый, его лучше съесть сразу или ненадолго оставить на столе при комнатной температуре. Авокадо часто продаются недозрелыми, и холод только остановит созревание, оставив плод невкусным.
Лук
Репчатый лук впитывает запахи других продуктов и оставляет им свой запах. При повышенной влажности он размягчается и покрывается плесенью. Хранить лук нужно как картофель — в прохладном сухом месте, но ни в коем случае не вместе с картофельными клубнями: при совместном хранении продукты быстрее испортятся.
Мед
Этот ценный продукт можно хранить при температуре от -7 до +20 градусов. Если в холодильнике температура ниже, то лучше оставить мед в темном шкафу. Мед сохраняет полезные качества и при более низких температурах, но в таком случае он засахарится, изменит вкус и может стать непригодным для рецептов, в которых требуется вязкость жидкого меда. Кстати, если после хранения при минусовой температуре мед остается жидким, значит, производитель перегрел продукт, и в таком меде значительно меньше полезных веществ.
Консервированные овощи
Домашние соленья, консервированные и маринованные, отлично сохраняются при комнатной температуре благодаря добавлению в них соли, сахара и уксуса. Поэтому ставить соленья в холодильник просто нет смысла. Содержимое открытой банки лучше употребить в течение пары суток, при этом оптимально держать ее на балконе в прохладное время года, когда температура не опускается ниже нуля.
Соусы и приправы
Промышленные соусы и кетчупы также содержат много уксуса, а некоторые из них сделаны на основе растительных масел. Хранение в холодильнике не принесет им никакой пользы, достаточно переложить эти продукты в кухонный шкаф в плотно закрытых жестяных банках или стеклянной посуде.
Коньяк
В холодильнике коньяк может изменить вкус и запах. Оптимальной температурой хранения для этого напитка считается 12 градусов. Но коньяк может храниться в темном месте и при комнатной температуре порядка 20 градусов, если нет возможности поместить бутылки в прохладное место.
Цитрусовые
Апельсины, лимоны и грейпфруты относятся к одной группе продуктов, но хранить их вместе не стоит — так они гораздо быстрее портятся и плесневеют. В холодильнике цитрусовые увядают и размягчаются, а их кожура покрывается пятнами, поэтому хранить их лучше при комнатной температуре и съедать в течение нескольких дней после покупки. Лимоны при теплой температуре хранятся значительно дольше, чем апельсины. Последние можно нарезать и заморозить.
Хлеб
Любую выпечку лучше употреблять свежей, в течение двух-трех дней с момента приготовления или покупки. В холодильнике хлебобулочные изделия засыхают быстрее. При этом хлеб сохраняет влагу при заморозке, так что если вам необходимо уехать, а выбрасывать выпечку жалко, кладите ее в морозильную камеру.
Шоколад
Плитки шоколада многие кладут в холодильник, чтобы они не таяли и дольше хранились. Но когда шоколад достают из холода, вырабатывается конденсат. Он растворяет часть сахара и оставляет белый налет на поверхности батончиков и плиток. Это не значит что продукт испортился, но, к примеру, владелец шоколадного производства The Chocolate Bar Люк Оуэн Смит утверждает, что в холоде шоколад теряет насыщенный вкус.
Зелень
Петрушку, укроп, зеленый лук, кинзу, салат и базилик лучше замораживать. Так они сохранят влагу и вкусовые качества. При хранении в холодильнике растения вянут и витаминов в них становится значительно меньше. Если же оставить зелень в принесенном из магазина полиэтиленовом пакете или пленке, то это гарантирует рост плесени, с которой есть продукт уже нельзя. Единственный способ сохранить свежую зелень в холодильнике — завернуть ее в бумагу или хлопковую салфетку, которые впитают влагу, и зелень не будет гнить.
крахмал как удобрение для помидор
крахмал как удобрение для помидорПоисковые запросы: удобрение помидор во время цветения и плодоношения, где купить крахмал как удобрение для помидор, какое удобрение положить при посадке картофеля.
крахмал как удобрение для помидор
спец удобрения для томатов, куриные перья как удобрение для томатов отзывы, фосфорные удобрения для рассады перцев, таблица применения органического удобрения, удобрение для картофеля фаскокомплексное удобрение для томатов в открытом грунте
таблица применения органического удобрения Томаты можно выращивать как в открытом, так и закрытом грунте. А благодаря трем народным способам можно сделать рассаду помидор просто. Крахмал, который входит в состав отвара, помогает растениям накапливать питательные компоненты. Для того, чтобы сделать такое удобрение, нужно промыть картофель. Ведь для приготовления подкормки используется лишь крахмал и вода, и по сути мы получим просто кисель. Берем 300 гр крахмала, разводим в 3 литрах воды и кипятим на маленьком огне. Затем кисель переливаем в ведро и добавляем еще 10. Я приветствую Вас, друзья – огородники, на своем канале. И сегодня у нас на повестке дня: Подкормка крахмалом смородины. Любой смородины. Черной, красной, белой. Крахмал-как удобрение. Найти ответ или обсудить вопрос на Асиенда.ру. На телеканале Усадьба рассказывали, что смородина. Я интересуюсь, использовал ли кто-нибудь подобные удобрения (+ещё Бутон у меня есть) для томатов и огурцов. Я их выращиваю на балконе. И вообще стоит ли мне удобрять мои. Крахмал, как удобрение для смородины — можно ли подкармливать кусты крахмалом и как правильно это делать. Итак, ниже мы детально рассмотрим, как правильно использовать картофельные очистки в качестве удобрений, как подкормить смородиновую культуру при помощи этого компонента и какими. Если старые кроссовки попахивают, насыпьте в них крахмал и оставьте на ночь. Запах впитается. Перед тем как обуться, не забудьте высыпать крахмал! Накрахмалить одежду. Крахмал-спрей представляет собой аэрозоль. Это – подкормка крахмалом, смородина чрезвычайно отзывчива на крахмал. Причем обыкновенный порошковый крахмал из магазина тут не подойдет – данная форма практически неусвояема для смородины. С успехом можно использовать картофельные очистки, но это не лучший вариант, так как даже тонкие. удобрение для картофеля фаско какие удобрения под посадку картофеля технология внесения жидких органических удобрений диплом
минеральные удобрения для перца в теплице комплексное удобрение для томатов в открытом грунте удобрение для огурцов форум удобрение помидор во время цветения и плодоношения какое удобрение положить при посадке картофеля спец удобрения для томатов куриные перья как удобрение для томатов отзывы фосфорные удобрения для рассады перцев
Недавно занималась домашним цветоводством. Тоже приобрела Агроап после прочтения реальных отзывов. Так теперь моя коллекция напоминают зеленую оранжерею. Листва отрастает более насыщенного зеленого цвета. Растения стали более здоровыми, красивыми. Впечатления от удобрения только положительные. Во-вторых, состав комплекса позволяет растениям максимально усваивать микроэлементы и питательные вещества. Благодаря этому созревание и рост плодов происходит быстрее, чем обычно. Использование удобрения позволит вырастить здоровые растения не только в саду и на огороде, но и дома. Средство AgroUp обеспечивает высокую урожайность, плоды вырастают крупнее, более мясистые, в составе нет химических веществ. Очевидное преимущество — универсальность — в составе удобрения нет пестицидов, а действующие компоненты эффективны в отношении садовых, огородных и домашних культур. Также средство можно применять на любом грунте. 1 Способы внесения удобрений. 2 Правила подкормки перцев. 3 Как и чем подкармливать перец в открытом грунте и в теплице на разных этапах. 3.1 После высадки рассады в грунт. 3.2 В период цветения. А подкормки простые — важно соотношение основных элементов, которое отличается от подкормок после пересадки рассады. Чем подкормить перец в июне: удобрения и соотношения. Есть две точки зрения на подкормку перца — противоположные. Первая. Если после высадки в грунт важно внести в составе NPK. Вторая подкормка перца после высадки в грунт. Следующая подкормка необходима через 10-14 дней после начала цветения. В указанный раствор минеральных удобрений (смю выше) можно добавить немного коровяка. После подкормок, так же, как и после поливов обязательно проводить. При нехватке кальция, азота, железа и др. элементов перцы желтеют и плохо растут. Для подкормки кальцием можно использовать настой из измельченной яичной скорлупы. Травяной настой тоже придется по вкусу перцам, в нем много азота для роста растений. Для этого измельченную траву (можно. Удобрения для перца, подкормка. Какие удобрения и когда нужно вносить под перец. Минеральные удобрения для перца. Подкормка перца после высадки в грунт. Существует несколько различных видов удобрений, используемых для подкормки болгарского перца, которые, в общем, делятся на две большие группы: минеральные (химические) – выпускаются в форме питательных комплексов, специально подобранных под потребности овоща на разных этапах. Подкормка перца в открытом грунте: особенности процедуры. Преимущества минеральных и органических удобрений. Удобрения для перца в открытом грунте для богатого урожая. Болгарский перец опровергает расхожую фразу, что вкусные продукты не могут быть полезными. Этот овощ является. Первый раз удобрения вносят приблизительно через 10-15 дней после высадки рассады в теплицу. Минеральную подкормку можно составить. Третья подкормка перца в открытом грунте. Третья подкормка перцев после высадки в грунт проводится во время, когда завязь превратилась в полноценные. Подкормка перца в теплице и в открытом грунте. За 2-3 недели до сбора урожая минеральная подкормка под корень запрещена, чтобы вещества не попали в плоды. Подбор удобрений для болгарского и для острого перцев. 2 Правила внесения удобрений. 3 Стадия получения рассады. 4 Подкормка перца перед посадкой саженцев. К первой относятся химические (минеральные) удобрения, выпускаемые в виде питательных комплексов, разработанных с учетом особенностей каждого овоща. Но подбирать такие препараты нужно. Какие удобрения нужны для перца. Принято использовать минеральные питательные комплексы и органику. Чтобы культура чувствовала себя комфортно, чередуйте эти подкормки: неделю применяется органика, следующие 7 дней минеральные добавки. Смеси могут быть в сухом и жидком видах.
крахмал как удобрение для помидор
удобрение для огурцов форум
Так, магний больше всего необходим для супесчаных и песчаных почв. Он входит в состав хлорофилла и необходим для дыхания растений. Также он усиливает мобильность в почве фосфатов и позволяет им быстрее поступать в растения. Для чего подкармливать помидоры борной кислотой? Рецепт подкормки, как готовить, в какой пропорции. Если растения выказывают признаки дефицита бора, народное удобрение можно применять чаще. Томаты очень требовательны к питательности почвы. Поэтому для получения урожая приходится использовать различные удобрения. Подкормка томатов борной кислотой положительно сказывается на состоянии растений. Подкормка томатов борной кислотой – агрономы предупреждают: бор токсичен, это неоднозначное удобрение. Когда и как можно подкармливать, в каком количестве – нормы внесения и сроки, чем опасен избыток и как выглядит нехватка. Приготовить раствор борной кислоты для опрыскивания томатов очень легко. Нужно взять половину чайной ложки порошка на 10 литров воды. Для того, чтобы борная кислота легче растворилась, разведите порошок сначала в стакане с водой, тщательно размешайте, а затем влейте концентрат в ведро. Применение бора для томатов в теплице и открытом грунте. Симптомы недостатка с фото и советы как подкормить: провести полив. Как вовремя заметить недостаток и правильно подкормить помидоры. Томаты – достаточно неприхотливая овощная культура, выращивать которые просто и легко. Но и этим овощам для. Третья подкормка с бором проводится исключительно при необходимости ускорить созревание томатов, обитающих в открытом грунте, например. Борные подкормки полезны не только для помидоров, но и для большинства овощных культур. Когда подкормка необходима: признаки дефицита бора у растений. Дефицит бора у помидоров проявляется довольно выраженными. Видео-рецепт удобрения для томатов из борной кислоты, золы и йода. Обработка томатов на разных стадиях жизненного цикла. Все уличные работы выполняются только в сухую. Борная кислота — источник бора, важного для развития томатов микроэлемента. Регулярная подкормка бором увеличивает урожайность на 20-30%, защищает помидоры от болезней, холодов и засухи. Томат – одна из культур, особенно чувствительных к нехватке бора. Бор особенно нужен томатам во время цветения. Он участвует в формировании генеративных органов, способствует. Состав и полезные свойства удобрения для помидоров. 1. 23.12.2019. Как приготовить и когда применять подкормку. Как вносить удобрение. Польза борной кислоты для помидор и как ее разводить для опрыскивания. Бор – микроэлемент, в котором томаты нуждаются в период формирования корневой массы и цветочных кистей, образования бутонов, цветения и завязывания. крахмал как удобрение для помидор. какие удобрения под посадку картофеля. Отзывы, инструкция по применению, состав и свойства. Органические удобрения — удобрения, содержащие элементы питания растений преимущественно в форме органических соединений. К ним относят навоз, компосты, торф, солому, зелёное удобрение, ил (сапропель). В статье обсуждаем органические удобрения. Рассказываем о видах органических подкормок, составе и применении. Вы узнаете, что относится к органическим удобрениям, какие у них преимущества и недостатки. Применение органических удобрений, способы внесения. Почему важна правильная доза внесения органики? К какому виду относятся органические удобрения? Характеристика наиболее ценных удобрений. Органические удобрения состоят из веществ животного и растительного происхождения, которые, разлагаясь, образуют минеральные вещества, при этом в приземный слой выделяется диоксид углерода, необходимый для фотосинтеза растений. Кроме того, органические удобрения благотворно влияют. Органические удобрения — внесение, виды и их классификация. Органические удобрения являются источником питания растений и результатом повышения плодородия почвы. Органические удобрения – что это такое? Виды органических удобрений. Органические удобрения – это подкормки, которые вмещают питательные вещества для растений в форме соединений органического типа. Что такое органические удобрения? Это вещества природного происхождения, в составе которых содержатся все необходимые элементы для плодотворного и активного роста растений. Самое важное преимущество органики состоит в том, что они очень быстро усваиваются растениями, чего нельзя сказать об. Для успешного выращивания сельскохозяйственных и огородных культур применяют не только агрохимикаты. Для большего эффекта разные виды органических удобрений, характеристики которых могут сильно разниться. Виды органических удобрений. Органические удобрения – весьма разнообразная категория, в нее входят различные виды подкормки для растений. Рассмотрим наиболее популярные удобрения на наших участках, а также плюсы и минусы каждого из них. Перечисляя все органические удобрения, стоит. Органические и минеральные удобрения. Виды органических удобрений. Удобрения на основе растительного сырья. Фосфор в органических удобрениях. Роль фосфора в жизни растений. Какие органические удобрения содержат.
(PDF) Гранулы крахмала в плодах томатов демонстрируют сложную структуру разложения
Статья J. Agric. Food Chem., Vol. 57, No. 18, 2009 8487
ЦИТАТА ЛИТЕРАТУРЫ
(1) Служба экономических исследований, U. Commodity Highlights 2004, 24 июня, 11.
(2) Обиадалла-Али, Х. Понимание разделения углерода в помидорах
Фрукты; Институт молекулярной физиологии растений им. Макса Планка: Golm,
Germany, 2003.
(3) Obiadalla-Ali, H .; Ферни, А.Р.; Kossmann, J .; Ллойд, Дж. Р.
Анализ развития углеводного обмена в плодах томата
(Lycopersicon esculentum cv. Micro-Tom). Physiol. Растение.
2004,120 (2), 196–204.
(4) Schaffer, A. A .; Петрейков, М. Метаболизм сахарозы в крахмал в плодах томата
, испытывающих временное накопление крахмала. Завод Phy-
сиол. 1997, 113 (3), 739–746.
(5) daSilva, P. M. F. R .; Eastmond, P.J .; Hill, L.M .; Smith, A.M .;
Rawsthorne, S.Метаболизм крахмала в развивающихся зародышах масличных семян
рапса. Planta 1997, 203 (4), 480–487.
(6) Smith, A.M .; Zeeman, S.C .; Смит, С. М. Распад крахмала. Анну.
Ред. Биол. Растений. 2005,56, 73–98.
(7) Beckles, D.M .; Craig, J .; Smith, A.M. АДФ-глюкозопирофос-
форилаза находится в пластиде развивающихся плодов томата. Завод
Physiol. 2001, 126 (1), 261–266.
(8) Robinson, N.L .; Hewitt, J.D .; Беннетт, А.Поглощает метаболизм в плодах томата
1. Изменения в развитии углеводного обмена —
ферментов. Plant Physiol. 1988, 87 (3), 727–730.
(9) Beckles, D. M. Субклеточное расположение АДФ-глюкозо-пирофо-
сфорилазы в накапливающих крахмал клетках; Кембриджский университет:
Кембридж, Великобритания, 1998.
(10) N’tchobo, H .; Дали, Н .; Nguyen-Quoc, B .; Фойе, C. H .; Yelle, S.
Синтез крахмала в томате остается постоянным на протяжении всего развития плода
и зависит от поставки сахарозы и активности сахарозы
синтазы.J. Exp. Бот. 1999,50 (338), 1457–1463.
(11) Carrari, F .; Baxter, C .; Усадель, Б .; Urbanczyk-Wochniak, E .; Занор,
М. И .; Нунес-Неси, А .; Никифорова, В .; Центеро, Д .; Ratzka, A .; Pauly,
М .; Sweetlove, L.J .; Fernie, A.R. Комплексный анализ метаболитов и уровней транскриптов
выявляет метаболические сдвиги, лежащие в основе развития плодов томатов
, и подчеркивает регуляторные аспекты поведения метаболической сети
. Plant Physiol. 2006, 142 (4), 1380–1396.
(12) Nguyen-Quoc, B .; Фойе, К. Х. Роль «бесполезных циклов» с участием инвертазы
и сахарозосинтазы в метаболизме сахарозы плодов томатов
. J. Exp. Бот 2001,52 (358), 881–889.
(13) Luengwilai, K .; Беклс, Д. М. Структурные исследования и морфо-
логия крахмала плодов томата. J. Agric. Food Chem. 2009,57, 282–291.
(14) Huber, K. C .; БеМиллер, Дж. Н. Визуализация каналов и полостей
гранул кукурузы и крахмала сорго.Cereal Chem. 1997, 74 (5), 537–541.
(15) Apinan, S .; Yujiro, I .; Hidefumi, Y .; Такеши, Ф .; Myllarinen, P .;
Forssell, P .; Поутанен, К. Визуальное наблюдение за гранулами гидролизованного картофеля
крахмала с помощью альфа-амилазы с конфокальной лазерной сканирующей микроскопией
. Starch-Staerke 2007,59 (11), 543–548.
(16) Alonso, A. P .; Vigeolas, H .; Raymond, P .; Rolin, D .; Dieuaide-
Noubhani, М. Новый цикл субстрата в растениях. доказательства высокого оборота глюкозо-фосфат-глюкоза
в стационарных экспериментах in vivo и экспериментах по импульсной маркировке
с [C-13] глюкозой и [C-14] глюкозой.
Plant Physiol. 2005, 138 (4), 2220–2232.
(17) Geigenberger, P .; Ститт, М. Бесполезный цикл синтеза сахарозы и деградации
участвует в регулировании распределения между сахарозой, крахмалом
и дыханием в семядолях прорастающих проростков Ricinus com-
munis L, когда транспорт флоэмы ингибируется. Planta 1991,
185 (1), 81–90.
(18) Wilson, J. D .; Bechtel, D. B .; Тодд, Т. С .; Сейб, П. А. Измерение
распределения гранул пшеничного крахмала по размерам с использованием анализа изображения
и технологии лазерной дифракции.Cereal Chem. 2006,83
(3), 259–268.
(19) Puncha-Arnon, S .; Puttanlek, C .; Rungsardthong, V .; Патипанават,
Вт .; Уттапап, Д. Изменения физико-химических свойств и морфо-
логии крахмала канны во время развития корневища. Углеводы. Polym.
2007,70 (2), 206–217.
(20) Hargreaves, J. A .; Априс, Т. Оборот крахмала и сахарозы в корнях
Pisum sativum. Фитохимия, 1988, 27 (6), 1627–1629.
(21) Хилл, С.А .; Априс, Т. Потоки углеводного обмена в созревающих бананах. Планта 1994,192 (1), 52–60.
(22) Fannon, J. E .; Hauber, R.J .; Бемиллер, Дж. Н. Поверхностные поры гранул крахмала
. Cereal Chem. 1992,69 (3), 284–288.
(23) Wang, F .; Sanz, A .; Brenner, M. L .; Смит, А. Синтаза сахарозы, накопление крахмала
и сила поглощения плодов томата. Plant Physiol.
1993,101 (1), 321–327.
(24) Alonso, A. P .; Раймонд, П.; Rolin, D .; Dieuaide-Noubhani, M.
Циклы субстрата в центральном метаболизме кончиков корней кукурузы при гипоксии
. Фитохимия 2007,68 (16-18), 2222–2231.
(25) Schwender, J .; Ohlrogge, J .; Шахар-Хилл, Ю. Понимание потока в метаболических сетях
растений. Curr. Opin. Plant Biol. 2004,7 (3), 309–
317.
Получено 13 мая 2009 г. Отредактированная рукопись получена 7 августа 2009 г.
Принята 10 августа 2009 г. Фонд Анандамахидола поддержал
K.L. Эта работа финансировалась грантом
MCB-0620001 Национального научного фонда, выданным D.M.B. и Калифорнийский университет экспериментального фонда
Закона о выкупе станций CA-D * -PLS-7821-H.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
NTRC играет решающую роль в метаболизме крахмала, окислительно-восстановительном балансе и росте плодов помидоров
Abstract
НАДФН-тиоредоксинредуктаза С (NTRC) образует отдельный каскад восстановления тиола в пластидах, сочетая активность НАДФН-тиоредоксинредуктазы и тиоредоксинредуктазы единственный полипептид.Хотя NTRC является важным регулятором фотосинтетических процессов в листьях, его функция в гетеротрофных тканях остается неясной. Здесь мы сосредотачиваемся на роли NTRC в развитии плодов томата ( Solanum lycopersicum ), представляющих гетеротрофные органы хранения, важные для сельского хозяйства и питания человека. Мы использовали специфический для плода промотор для снижения экспрессии NTRC посредством РНК-интерференции в развивающихся плодах томатов на 60-80% по сравнению с диким типом. Это привело к замедлению роста плодов, в результате чего полностью созревшие плоды стали меньше и легче, содержали меньше сухого вещества и больше воды.В незрелых плодах подавление NTRC уменьшало временное накопление крахмала, что приводило к последующему снижению растворимых сахаров в спелых плодах. Ингибирование синтеза крахмала было связано с уменьшением состояния окислительно-восстановительной активации АДФ-Glc пирофосфорилазы и растворимой синтазы крахмала, которые катализируют соответственно первую и конечную стадии полимеризации биосинтеза крахмала. Это сопровождалось снижением уровня ADP-Glc. Подавление NTRC также привело к сильному увеличению восстановительных состояний окислительно-восстановительных систем NAD (H) и NADP (H).Профилирование метаболитов интерференционных линий NTRC-РНК выявило повышенные уровни органических и аминокислот, но пониженные уровни сахара, что означает, что NTRC регулирует осмотический баланс развивающихся плодов. Эти результаты показывают, что NTRC действует как центральный узел в регулировании углеродного метаболизма и окислительно-восстановительного баланса в гетеротрофных плодах томатов, влияя на развитие плодов, а также на конечный размер и качество плодов.
Редукционно-окислительная (окислительно-восстановительная) регуляция, по-видимому, является фундаментальным интегратором метаболических путей в различных субклеточных компартментах (Geigenberger and Fernie, 2014).В хлоропластах растений существуют две разные окислительно-восстановительные системы тиолов: система ферредоксин (Fdx) -тиоредоксин (Trx), которая зависит от восстановления Fdx посредством фотосинтетического транспорта электронов в ответ на свет, и НАДФН-зависимая Trx-редуктаза C (NTRC). система, которая зависит от NADPH и, таким образом, может быть связана с Fdx-NADPH редуктазой на свету или метаболизмом сахара в темноте (Buchanan and Balmer, 2005; Zaffagnini et al., 2018). NTRC является необычным белком, поскольку он содержит домены НАДФН-Trx-редуктазы и Trx на одном и том же полипептиде (Serrato et al., 2004). Эта особенность позволяет NTRC использовать NADPH в качестве источника электронов для регулирования различных белков-мишеней хлоропластов посредством тиол-дисульфидной модуляции (Spínola et al., 2008; Geigenberger et al., 2017).
При анализе мутантов, переносящих ДНК Arabidopsis ( Arabidopsis thaliana ), было установлено, что NTRC участвует в различных функциях хлоропластов. Есть in planta доказательства того, что NTRC важен для восстановления 2-Cys пероксиредоксинов (2-Cys Prxs) для удаления перекиси водорода (Pérez-Ruiz et al., 2006; Кирхштайгер и др., 2009; Pérez-Ruiz and Cejudo, 2009) и для облегчения синтеза крахмала посредством окислительно-восстановительной регуляции ADP-Glc пирофосфорилазы (AGPase; Michalska et al., 2009; Lepistö et al., 2013). Кроме того, NTRC имеет перекрывающиеся функции с Fdx-зависимыми системами Trx для оптимизации фотосинтетических процессов, таких как нефотохимическое тушение световой энергии (Thormählen et al., 2015, 2017; Naranjo et al., 2016; Yoshida and Hisabori, 2016), Calvin- Активность цикла Бенсона (Thormählen et al., 2015; Никканен и др., 2016; Yoshida and Hisabori, 2016), биосинтез хлорофилла (Richter et al., 2013; Pérez-Ruiz et al., 2014; Da et al., 2017) и синтез АТФ (Carrillo et al., 2016; Nikkanen et al., 2016). В этом контексте NTRC косвенно поддерживает восстанавливающую способность пула Fdx-TRX, ограничивая их повторное окисление через цикл окисления, включающий пероксид водорода и окисленные 2-Cys Prxs (Pérez-Ruiz et al., 2017). Следовательно, NTRC имеет решающее значение в балансировании путей окисления и восстановления в хлоропласте для оптимизации фотосинтеза, общего роста растений и развития в ответ на свет (Thormählen et al., 2015, 2017; Carrillo et al., 2016; Ёсида и Хисабори, 2016).
Помимо своей роли в хлоропластах, NTRC также присутствует в нефотосинтетических пластидах вместе с Fdx-зависимыми Trxs (Kirchsteiger et al., 2012). Однако пока мало что известно о роли NTRC в гетеротрофных тканях растений, которые лишены фотосинтетических функций. Имеются данные о том, что NTRC влияет на синтез крахмала в корнях Arabidopsis (Michalska et al., 2009), но это не влияет на фенотип роста мутанта Arabidopsis ntrc (Kirchsteiger et al., 2012). Комплементация мутанта ntrc сверхэкспрессией NTRC под контролем лист-специфичного промотора привела к фенотипам дикого типа, но мутантный фенотип остался неизменным при использовании корневого-специфичного промотора (Kirchsteiger et al., 2012) . Таким образом, эти данные указывают на роль NTRC в фотосинтетических листьях, а не в нефотосинтетических корнях.
Помидор ( Solanum lycopersicum ) — важная садовая культура и основной компонент рациона человека, служащий основной модельной системой для изучения физиологии и развития плодов (Kanayama, 2017).На протяжении всего процесса развития плодов происходит множество изменений цвета, твердости, вкуса и аромата, которые связаны с метаболическими сдвигами, регулируемыми развитием (например, изменения уровней углеводов, кислот и летучих соединений; Carrari et al., 2006) . Примечательно, что обилие углеводов является ключевым фактором, определяющим урожай и качество фруктов (Канаяма, 2017). Во время развития незрелые плоды томатов действуют как сильные поглощающие ткани, в которые огромное количество углерода импортируется в виде Suc из тканей листа (Osorio and Fernie, 2014).Большая часть Suc мобилизуется в Glc и Fru, а затем превращается в крахмал в качестве промежуточного хранилища углерода, способствующего поглощающей активности незрелых плодов (Osorio and Fernie, 2014). На более поздних стадиях созревания крахмал разлагается до растворимых сахаров, которые являются ключевыми определяющими факторами вкуса и качества фруктов (Davies et al., 1981; Gierson and Kader, 1986).
Было высказано предположение, что изменения временного пула крахмала незрелых плодов важны для накопления растворимого сахара в созревающих фруктах, что определяет конечный размер и качество плодов (Schaffer et al., 2000; Петрейков и др., 2009). В этом контексте AGPase служит ферментом, ограничивающим скорость синтеза крахмала в развивающихся плодах томатов (Petreikov et al., 2009). AGPase — это гетеротетрамерный фермент, расположенный в пластиде и состоящий из двух больших и двух малых субъединиц, которые подвергаются аллостерической активации глицерат-3-фосфатом (3-PGA) и ингибируются неорганическим фосфатом (Pi; Preiss, 1973). AGPase также подвержена окислительно-восстановительной регуляции, включающей обратимое восстановление регуляторного дисульфида, связывающего малые субъединицы, что влияет на аллостерические свойства фермента (Fu et al., 1998; Tiessen et al., 2002). Ранее было продемонстрировано, что манипулирование клеточным окислительно-восстановительным балансом во время развития плодов томата приводит к изменениям в соотношении НАДФН-НАДФ + , которые сопровождаются изменениями в состояниях окислительно-восстановительной активации НАДФ-малатдегидрогеназы и AGPазы развивающиеся плоды (Centeno et al., 2011). Изменения окислительно-восстановительного состояния AGPase были дополнительно связаны с изменениями синтеза крахмала в незрелых плодах и последующими изменениями содержания растворимого сахара в зрелых плодах (Centeno et al., 2011). Хотя эти исследования предоставляют доказательства того, что окислительно-восстановительное регулирование посредством тиол-дисульфидной модуляции участвует в развитии плодов томатов, лежащие в основе специфические для фруктов тиол-окислительно-восстановительные системы остаются нерешенными.
В этом исследовании мы подавили экспрессию гена NTRC , специфически в тканях плодов, путем создания конструкции РНК-интерференции (РНКи) под контролем промотора пататина B33, специфичного для плода. Линии NTRC-RNAi характеризовались снижением уровня транскриптов NTRC и уровня белка в развивающихся плодах от 60% до 80%.В незрелых плодах подавление NTRC уменьшало временное накопление крахмала за счет снижения состояния окислительно-восстановительной активации AGPазы и активности растворимой синтазы крахмала, что впоследствии привело к снижению накопления растворимых сахаров во время созревания и снижению урожайности и качества полностью созревших плодов. . Это сопровождалось повышенным состоянием восстановления окислительно-восстановительных пар НАД (Н) и НАДФ (Н). Эти результаты подтверждают ранее неизвестную функцию NTRC как центрального узла в регулировании углеродного метаболизма и окислительно-восстановительного баланса в развивающихся фруктах.
РЕЗУЛЬТАТЫ
Получение трансгенных растений томатов со сниженной экспрессией
NTRC под контролем специфичного для плода промотораЧтобы подавить экспрессию гена NTRC , специфически в тканях плодов, мы создали конструкцию NTRC-RNAi под контролем промотор пататина B33, который, как было ранее установлено, обеспечивает специфическую экспрессию плодов в растениях томатов (Rocha-Sosa et al., 1989; Frommer et al., 1994; Obiadalla-Ali et al., 2004), при этом канамицин служит маркер для выбора ().Полученную конструкцию трансформировали с помощью процесса, опосредованного Agrobacterium tumefaciens , в растения томатов. Для идентификации функциональных линий NTRC-RNAi для генотипирования были отобраны образцы плодов, собранных через 35 и 65 дней после цветения (DAF). Как показано на фиг. 2, 26 и 33 линии NTRC-RNAi, все содержат ген неомицинфосфотрансферазы II ( npt II), что указывает на присутствие кассеты NTRC-RNAi в их геноме. Уровень транскрипта гена NTRC (LOC101254347) снизился на 60-80% по сравнению с диким типом в трех линиях РНКи в образцах плодов 35- () и 65-DAF (дополнительный рис.S1A). Кроме того, уровни белка NTRC RNAi-2, RNAi-26 и RNAi-33 снизились на 50-80% по сравнению с диким типом, как в образцах плодов 35- (), так и 65-DAF (дополнительный рисунок S1, Б и В). Экспрессия второго гена NTRC (LOC101266017; Nájera et al., 2017) также была проанализирована, но его экспрессия была слишком низкой, чтобы ее можно было обнаружить в тканях плодов. Таким образом, мы пришли к выводу, что линии NTRC-RNAi 2, 26 и 33 подходят для изучения функции NTRC в плодах томата.
Молекулярная характеристика линий 2, 26 и 33 NTRC-RNAi по сравнению с диким типом (WT) в 35 плодах томатов DAF.A. Конструкция РНКи, нацеленная на ген NTRC . B. Генотипирование с помощью ПЦР-анализа для обнаружения гена npt II в линиях NTRC-RNAi. C. Определение уровня транскрипта NTRC с использованием RT-qPCR. D, Обнаружение белков NTRC с помощью иммуноблоттинга. E, Количественная оценка содержания белка NTRC с использованием программного обеспечения Image-J. Уровень адаптерных комплексов клатрина ( CAC ) служит эталонным геном для генотипирования и RT-qPCR. Уровень белка большой субъединицы рибулозо-1,5-бисфосфаткарбоксилазы / оксигеназы (RbcL) служил в качестве контроля нагрузки для иммуноблоттинга.Результаты представляют собой среднее значение ± стандартное отклонение от шести биологических повторов. Для статистического анализа использовали ANOVA с тестом Даннета (* P <0,05 по сравнению с диким типом).
Специфическое для фруктов замалчивание NTRC не оказывает существенного влияния на процесс развития плодов
Различные стадии развития плодов томатов характеризуются изменениями пигментации и созревания. Чтобы проанализировать, влияет ли снижение экспрессии NTRC на процесс развития плодов томата, мы проанализировали уровни хлорофилла и белка светособирающего комплекса II (Lhcb2), а также экспрессию генов, участвующих в синтезе этилена и созревании в 35-DAF и 65-DAF. фрукты.В плодах 35-DAF содержание хлорофилла и уровень белка Lhcb2 существенно не изменились в линиях NTRC-RNAi по сравнению с диким типом, в то время как ни один параметр не был обнаружен в плодах 65-DAF во всех генотипах (дополнительный рисунок S2, A и B). ). При сравнении линий NTRC-RNAi с линиями дикого типа не было видимых изменений в цвете плодов при 35 DAF () или 65 DAF (дополнительный рис. S3A). Чтобы дополнительно проанализировать, влияет ли подавление NTRC на процессы созревания, мы проанализировали экспрессию генов, участвующих в производстве этилена (S-аденозилгомо-цис-гидролаза [ SAHh3 ]; Yang et al., 2017) и созревание (НЕСЗРЕВАЮЩИЕ [ NOR ]; Ma et al., 2019) как в 35-DAF, так и в 65-DAF плодах. Оба гена были активированы, когда плоды стали зрелыми, но уровни их экспрессии существенно не изменились между линиями дикого типа и NTRC-RNAi (дополнительный рис. S2, C и D). В совокупности подавление NTRC не повлияло на процесс развития плодов. В то время как плоды 35-DAF представляют зрелую зеленую стадию, а плоды 65-DAF представляют стадию полного созревания, между линиями NTRC-RNAi и диким типом не было различий в зависимости от стадии.
Фенотипический анализ полностью созревших плодов томата (65 DAF) линий 2, 26 и 33 NTRC-RNAi по сравнению с диким типом (WT). A, видимый фенотип спелых плодов. Масштабная линейка = 2 см. B, длина и ширина плода. C, плоды в свежем весе. D, отношение сухого веса плода к весу в сыром виде. Результаты представляют собой среднее значение ± стандартное отклонение от шести биологических повторностей (каждая повторность содержит данные от одного до трех плодов). Для статистического анализа использовали ANOVA с тестом Даннета (* P <0,05 по сравнению с диким типом).
Специфическое для фруктов подавление NTRC приводит к получению более мелких и легких фруктов с пониженным соотношением сухого веса к свежему весу
Чтобы исследовать, влияет ли подавление NTRC на общий рост плодов, полностью созревшие плоды (65 DAF) были собраны для измерения окончательный размер и вес плодов. Как показано на фиг. 7, хотя и линии дикого типа, и линии РНКи были полностью спелыми, конечный размер плодов линий NTRC-РНКи был явно меньше, чем у линий дикого типа. Кроме того, линии NTRC-RNAi показали уменьшение высоты плода на 20% и ширины плода на 15% по сравнению с диким типом ().Конечный вес свежих плодов томатов в линиях NTRC-RNAi был на 40% ниже, чем у плодов дикого типа (), а плоды томатов NTRC-RNAi также содержали меньше сухого вещества и больше воды, чем плоды дикого типа (). Эффекты подавления NTRC на рост плодов и содержание сухого вещества также наблюдались при 35 DAF, указывая на то, что они не были вызваны эффектами развития (дополнительный рисунок S3). Взятые вместе, специфическое для фруктов подавление NTRC привело к образованию более мелких и легких плодов, состоящих из меньшего количества сухого вещества в растениях томатов.
Специфическое для фруктов подавление NTRC приводит к снижению накопления крахмала и растворимых сахаров во время развития плодов
Развитие плодов томата характеризуется временным накоплением крахмала в незрелых плодах, который впоследствии мобилизуется, чтобы способствовать накоплению растворимых сахаров, особенно Glc, в зрелых плодах (Schaffer et al., 2000). Чтобы выяснить, связано ли подавление роста плодов в линиях NTRC-RNAi с уменьшением временного пула крахмала, мы проанализировали уровни крахмала и растворимых сахаров во время развития плодов.В незрелых плодах (35 DAF) подавление NTRC привело к сильному снижению накопления крахмала до 25-50% от уровня дикого типа (). В зрелых плодах (65 DAF), где крахмал был почти полностью мобилизован во всех генотипах, линии NTRC-RNAi все еще демонстрировали тенденцию к снижению уровней остаточного крахмала по сравнению с диким типом (). В противоположность этому, подавление NTRC не влияет на содержание белка в плодах ни на одной из стадий развития (Supplemental Fig. S4).
Накопление крахмала и растворимых сахаров в плодах томатов дикого типа (WT) и NTRC-RNAi линий 2, 26 и 33.Уровни крахмала (A и B) и растворимых сахаров (C и D) измеряли в плодах, собранных при 35 (A и C) и 65 DAF (B и D). Результаты представляют собой среднее значение ± стандартное отклонение от шести биологических повторов. Для статистического анализа использовали ANOVA с тестом Даннета (* P <0,05 по сравнению с диким типом). FW, свежий вес
Чтобы исследовать, сопровождается ли ингибирование накопления крахмала увеличением растворимых сахаров, мы проанализировали уровни Glc, Fru, Suc и мальтозы в линиях NTRC-RNAi по сравнению с диким типом.На обеих стадиях развития уровень Glc, основного растворимого сахара в плодах томатов, был сильно снижен в линиях NTRC-RNAi по сравнению с диким типом, в то время как уровни Fru, Suc и мальтозы показали лишь незначительное снижение (, и; Дополнительная таблица S1). Таким образом, вмешательство в экспрессию NTRC в плодах томата значительно ухудшило временное накопление крахмала и окончательное накопление растворимого сахара во время их развития, в то время как не было никакого воздействия на содержание белка в плодах.
Профиль метаболитов плодов томата NTRC-RNAi линий 2, 26 и 33 NTRC-RNAi по сравнению с диким типом (WT).Профиль метаболитов измеряли во фруктах, собранных при 35 (слева) и 65 DAF (справа), с помощью GC-TOF-MS. A, Сахар и сахарные спирты. B, Органические кислоты. C, аминокислоты и промежуточные соединения цикла трикарбоновых кислот. D, Разное. Результаты нормализуются к дикому типу с преобразованием log 2 и визуализируются в виде тепловой карты с иерархической кластеризацией, выполняемой программным обеспечением R. Данные взяты из дополнительной таблицы S1.
Вмешательство в экспрессию NTRC в томатных плодах оказывает незначительное влияние на уровни фосфорилированных промежуточных продуктов и адениновых нуклеотидов
Чтобы выяснить, связано ли уменьшение накопления крахмала и сахара с изменениями сахара и энергетического метаболизма, мы дополнительно проанализировали уровень фосфорилированных метаболиты и нуклеотиды аденина в развивающихся плодах томата.В линиях NTRC-RNAi уровни Glc-6-фосфата (G6P), Fru-6-фосфата и Glc-1-фосфата (G1P) не показали устойчивых изменений при 35 или 65 DAF по сравнению с диким типом (дополнительная информация Таблицы S1 и S2). Также не было обнаружено существенных изменений в уровне гликолитического промежуточного звена 3-PGA между линиями дикого типа и NTRC-RNAi на любой стадии развития. Также были проанализированы триозофосфаты, но их уровень был ниже предела обнаружения, что характерно для гликолитического пути, действующего в гетеротрофных тканях (Hatzfeld and Stitt, 1990).Кроме того, уровни АТФ и АДФ и отношение АТФ к АДФ в линиях NTRC-RNAi существенно не изменились во время развития плода по сравнению с диким типом (дополнительная таблица S2). Данные также показывают, что уровни субстратов (G1P и ATP) и аллостерических регуляторов (3-PGA и Pi) AGPase существенно не изменились в трансгенных плодах. Взятые вместе, эти результаты демонстрируют, что вмешательство в экспрессию NTRC не нарушает гликолитический метаболизм или энергетическое состояние аденилата в развивающихся фруктах.
Вмешательство в экспрессию NTRC в плодах томатов приводит к глобальным изменениям в их профиле метаболитов
Чтобы получить более глубокое представление о более глобальных изменениях метаболитов в линиях томатов NTRC-RNAi, мы затем выполнили профилирование метаболитов с помощью газовой хроматографии по времени пролета. масс-спектрометрия (GC-TOF-MS). Сначала мы подтвердили, что линии NTRC-RNAi показали в целом более низкие уровни сахара по сравнению с диким типом (слева), что имело место для Glc (от 55% до 80% от уровня дикого типа), Fru (от 65% до 85% от уровня дикого типа). % от уровня дикого типа) и Suc (от 40% до 55% от уровня дикого типа), а также других сахаров.Более того, при подавлении NTRC уровни некоторых сахаров снижались сильнее при 65 DAF, чем при 35 DAF (справа), включая гентиобиозу (в 3,6 раза), мальтозу (в 2 раза) и рибозу (в 2,3 раза). За исключением myo, -инозита, не было значительных изменений в сахарных спиртах в линиях NTRC-RNAi ().
Напротив, уровни нескольких органических кислот показали возрастающую картину в линиях NTRC-RNAi по сравнению с уровнями дикого типа, включая уровни 2-иминдазолидон-4-карбоновой кислоты, трансаконитовой кислоты, трансафеиновой кислоты и хинной кислоты, которые являются участвует в синтезе фенольных соединений.Уровни других, таких как 1-пирролин-2-карбоксилат и гулоновая кислота, снизились (). Снижение уровня гулоновой кислоты может быть связано с более низким содержанием Glc, поскольку эта кислота образуется в результате окисления Glc. Промежуточные продукты цикла трикарбоновых кислот также показали возрастающую картину по сравнению с диким типом. Уровни лимонной кислоты и 2-оксоглутаровой кислоты были увеличены в 2-4 раза в линиях NTRC-RNAi на обеих стадиях развития плода, в то время как яблочная кислота, фумаровая кислота и янтарная кислота показали лишь незначительные изменения по сравнению с диким типом () .Интересно, что предшественник цикла трикарбоновых кислот, пировиноградная кислота, увеличивался только в незрелых плодах линий NTRC-RNAi по сравнению с диким типом (слева).
В дополнение к органическим кислотам уровни аминокислот также увеличивались, в то время как экспрессия NTRC снижалась. Asn, 3-циано-Ala и homo-Ser были в 1,5–2 раза выше в линиях NTRC-RNAi в незрелых плодах (слева), и это изменение становилось более серьезным по мере созревания плодов (справа). Другие аминокислоты, такие как Trp, His, Gln и Met, показали умеренное увеличение в линиях NTRC-RNAi по сравнению с диким типом.Метаболиты, такие как мочевина, амины, нуклеозиды и их производные, также были проанализированы, но результаты выявили лишь незначительные различия между линиями NTRC-RNAi и диким типом на обеих стадиях (). В целом, эти результаты показывают, что вмешательство в экспрессию NTRC влияет на баланс между сахарами, органическими кислотами и аминокислотами во время развития плода.
Вмешательство в экспрессию NTRC в плодах томатов приводит к повышению клеточного редокс-состояния пиридиновых нуклеотидов
Редокс-пары NAD (H) и NADP (H) участвуют в различных метаболических процессах в различных субклеточных компартментах (Berger et al., 2004; Geigenberger and Fernie, 2014), а также служат кофакторами для NTRC, чтобы уменьшать и регулировать последующие пластидные белки (Pérez-Ruiz and Cejudo, 2009). Чтобы выяснить, влияет ли подавление NTRC на уровни этих пиридиновых нуклеотидов, мы затем проанализировали уровни NAD (P) H и NAD (P) + в развивающихся плодах томатов. В незрелых плодах (35 DAF) нарушение экспрессии NTRC привело к сильному увеличению уровней восстановленных форм, NADPH и NADH, в то время как уровни окисленных форм, NADP + и NAD + , снизились ().Это привело к 2-5-кратному увеличению отношения НАДФН-НАДФ + и 1,5-2-кратному увеличению отношения НАДН-НАД + в плодах линий NTRC-RNAi по сравнению с плодами линий NTRC-RNAi. дикого типа ().
Изменение уровня пиридиновых нуклеотидов в плодах томатов линий NTRC-RNAi по сравнению с диким типом (WT). Уровни НАДФ + , НАДФН, НАД + и НАДН (А и В), а также отношения НАДФ к НАДФ + и НАДН к НАД + (C и D) были измерены во фруктах, собранных в 35 (A и C) и 65 DAF (B и D).Результаты представляют собой среднее значение ± стандартное отклонение от шести биологических повторов. Для статистического анализа использовали ANOVA с тестом Даннета (* P <0,05 по сравнению с диким типом). FW, свежий вес
В полностью спелых плодах (65 DAF) подавление NTRC также привело к увеличению NADPH и NADH, но уровни окисленных форм NADP + и NAD + существенно не изменились (), что привело к незначительному увеличению соотношений NADPH к NADP + и NADH к NAD + в линиях NTRC-RNAi ().Вместе эти результаты показывают, что снижение экспрессии NTRC вызывает повышенное состояние восстановления окислительно-восстановительных пар NAD (H) и NADP (H) в развивающихся плодах томатов, причем эффект смягчается, когда плоды созревают.
Вмешательство в экспрессию NTRC в плодах томатов приводит к снижению состояния окислительно-восстановительной активации AGPase и снижению уровней ADP-Glc
Для исследования возможных механизмов, участвующих в ингибировании накопления фруктового крахмала в ответ на снижение экспрессии NTRC, мы измеряли активность ферментов, участвующих в метаболизме крахмала.Сначала мы проанализировали AGPase, которая катализирует первую коммитируемую стадию биосинтеза крахмала, поскольку ее активность регулируется посредством тиол-дисульфидной модуляции с участием систем Trx и NTRC в пластиде (Geigenberger and Fernie, 2014; Geigenberger et al., 2017). Поскольку AGPase активируется за счет восстановления межмолекулярной дисульфидной связи между двумя небольшими субъединицами (малая субъединица 1 AGPase [APS1]) гетеротетрамерного холофермента (Ballicora et al., 2000; Tiessen et al., 2002; Hendriks et al., 2003), мы проанализировали изменения окислительно-восстановительного состояния APS1, проанализировав характер его мономеризации, когда экспрессия NTRC была снижена.В развивающихся плодах (35 DAF) нарушение экспрессии NTRC незначительно снижает коэффициенты мономеризации APS1 () на ~ 20–30%, что указывает на повышенное окисление белка. Также были протестированы образцы спелых плодов (65 DAF), но не было устойчивых различий между линиями дикого типа и РНКи в мономеризации APS1 (дополнительный рисунок S5A).
Редокс и аллостерические свойства AGPазы и содержание эндогенного ADP-Glc в 35 плодах томатов DAF линий NTRC-RNAi 2, 26 и 33 по сравнению с диким типом (WT).A, окислительно-восстановительное состояние межмолекулярной дисульфидной связи в APS1. B, активность AGPазы, определенная с активацией 10 мМ 3-PGA или без нее. C, кинетика активности AGPase в присутствии различной концентрации 3-PGA, как показано на рисунке. D, содержание эндогенного ADP-Glc. Отношение мономеров к димерам соответствующих белков количественно определяли с помощью программного обеспечения Image-J. Результаты представляют собой среднее значение ± стандартная ошибка ( n = 5–6). Для статистического анализа использовали ANOVA с тестом Даннета (* P <0,05 по сравнению с диким типом).FW, сырая масса
AGPase также подвержена аллостерической регуляции, поскольку активируется 3-PGA и ингибируется Pi (Kleczkowski, 1999; Hwang et al., 2005). Редокс-регуляция AGPase изменяет кинетические свойства холофермента, при этом окисление APS1 приводит к ферменту, менее чувствительному к аллостерической активации (Tiessen et al., 2002; Hendriks et al., 2003). Чтобы выяснить, приводит ли нарушение экспрессии NTRC в плодах томата к изменению кинетических свойств AGPase, мы проанализировали активность AGPase в отсутствие или в присутствии ее аллостерического активатора, 3-PGA, в плодах 35-DAF.Как показано на фиг.3, линии NTRC-RNAi показали активность AGPase, сравнимую с активностью AGPase дикого типа, когда 3-PGA не включали в ферментный анализ, но наблюдалось снижение активности AGPase на 30% по сравнению с активностью AGPase дикого типа в присутствии 10 мм 3-PGA. Чтобы исследовать кинетику активации 3-PGA более подробно, мы выполнили анализ фермента AGPase в присутствии различных концентраций 3-PGA. показывает, что у дикого типа увеличение концентрации 3-PGA от 0,025 до 10 мМ приводило к прогрессивному увеличению активности AGPase от 0.От 2 до 6,0 нмоль мин -1 мг -1 белка. AGPase из линий NTRC-RNAi была менее чувствительна к активации 3-PGA, что приводило к снижению активности на 30-50% при инкубации в присутствии 0,25-10 мМ 3-PGA. Этот диапазон концентраций соответствует концентрации 3-PGA в пластиде in vivo, которая, по оценкам, составляет от 0,3 до 0,5 мм с использованием неводной фракции растущих клубней картофеля ( Solanum tuberosum ) (Farré et al., 2001; Tiessen et al. ., 2002) и развивающихся семян ячменя ( Hordeum vulgare ) (Tiessen et al., 2012) и от 0,7 до 1 мм с использованием уровней 3-PGA из дополнительной таблицы S1 (70–100 нмоль г -1 свежей массы), учитывая, что вакуоль не содержит 3-PGA и составляет 90% объема клетки. . Общая активность AGPase в полностью спелых плодах (65 DAF), которые характеризуются общим подавлением синтеза крахмала (), была в 10 раз ниже, чем в незрелых плодах (35 DAF) как в присутствии, так и в отсутствие 3-PGA в ферментный анализ, и не было значительных различий между линиями дикого типа и NTRC-RNAi (дополнительный рис.S5B).
ADP-Glc является прямым продуктом реакции, катализируемой AGPase, и представляет собой конечный предшественник конечных реакций полимеризации при синтезе крахмала. В плодах с 35-DAF подавление NTRC снижало концентрацию ADP-Glc до 50-75% от уровня дикого типа (). Это обеспечивает независимое подтверждение того, что молчание NTRC приводит к ингибированию активности AGPase in vivo.
Подавление NTRC приводит к снижению активности синтазы растворимого крахмала
Помимо AGPase, мы также исследовали другие ферменты, участвующие в метаболизме крахмала в 35-DAF плодах.Растворимые и связанные с гранулами синтазы крахмала катализируют конечные реакции полимеризации биосинтеза крахмала. Как показано на фиг.4, нарушение экспрессии NTRC снижает активность синтазы растворимого крахмала до 25-40% от уровня дикого типа, в то время как активность связанной с гранулами синтазы крахмала снижается лишь незначительно. Растворимая синтаза крахмала подвергается посттрансляционной окислительно-восстановительной регуляции и является потенциальной мишенью для NTRC in vitro (Skryhan et al., 2015, 2018). Напротив, подавление NTRC не оказало значительного влияния на активность фосфорилаз крахмала и гидролаз крахмала, которые участвуют в деградации крахмала ().Вместе эти результаты означают, что подавление экспрессии NTRC приводит к снижению накопления крахмала за счет совместного ингибирования как AGPазы, так и синтазы растворимого крахмала.
Таблица 1.
Изменения активности ферментов метаболизма крахмала в плодах томатов 35-DAF дикого типа и линиях NTRC-RNAiРезультаты представляют собой среднее значение с SE из шести биологических повторов. Для статистического анализа использовался ANOVA с тестом Даннета. (* P <0,05 по сравнению с диким типом).FW, свежий вес.
| Ферменты | Дикий тип | RNAi-2 | RNAi-26 | RNAi-33 |
|---|---|---|---|---|
| Растворимая синтаза крахмала (нмоль мин. 44,8 ± 4,2 | 16,8 ± 2,6 * | 11,4 ± 3,3 * | 18,6 ± 6,2 * | |
| Гранулированная синтаза крахмала (пмоль мин -1 мг -1 FW) | 2,9 ± 0,2 | 2.0 ± 0,1 * | 2,0 ± 0,1 * | 2,4 ± 0,2 |
| Фосфорилаза крахмала (нмоль мин −1 мг −1 белок) | 49,6 ± 10,9 | 76,5 ± 18,3 | 4,230,7 ± | 40,0 ± 11,3 |
| Гидролазы крахмала (мкмоль мин −1 мг −1 белка) | 2,4 ± 0,4 | 2,3 ± 0,7 | 1,1 ± 0,2 | 1,0 ± 0,4 |
Крахмал и пищевая ценность — Top 221 FoodsОвощилук-порей, зелень репы, швейцарский мангольд, зелень горчицы, ямс, фенхель, артишоки, брокколи, цветная капуста, капуста, стручковая фасоль, брюссельская капуста, капуста, сладкий картофель, картофель, морковь, салат, салат айсберг, шпинат, шиитак грибы, сельдерей, салат, грибы, картофель, печеный картофель, лук, спаржа, тыква, редис, таро, репа, окра, ревень, сладкая кукуруза, вигна, водоросли, бобыФруктыбанан, яблоко, апельсин, персик, абрикосы, мускатная тыква, лимон, виноград, арбуз, клубника, зеленый виноград, киви, черника, кабачки, инжир, медвяная роса, баклажаны, финики, оливки, груша, кокос, клюква, гранат, манго, ананас, вишня, дыня, грейпфрут, мандарин, нектарины, изюм, сливы, помидоры, перец чили, перец чили, перец, авокадо, огурец, кабачки летом, кабачки зимой, папайя, чернослив, фруктовый салат, хурмаМорепродуктылосось, треска, сардины, креветки, тунец, рыба, салат из тунца, палтус, морской окунь, гребешок, кр ab, лобстер, устрицы, сельдь, пикша, палтус, скумбрия, щука, минтай, форельНапиткикофе, яблочный сок, клюквенный сок, красное вино, обезжиренное молоко, белое вино, вино, молоко, пиво, алкоголь, кокс, соя молоко, цельное молоко, сливовый сок, чай, коктейль, кокосовое молоко, апельсиновый сок, энергетические напитки, безалкогольные напитки, грейпфрутовый сок, лимонный сок, лимонадОрехи и семенасемена чиа, семена подсолнечника, орехи пекан, тыквенные семечки, фундук, орехи, миндаль, семена кунжута, грецкие орехи, кешью, арахис, каштаны, кокосовое мясо, орехи гинкго, смешанные орехи, орехи макадамияФасоль и чечевицасоевые бобы, пегая фасоль, фасоль, чечевица, черная фасоль, фасоль лима, морская фасоль, свекла, фасоль, фасоль гарбанзо, запеченная фасольМясоговядина, стейк, оленина, бекон, курица, жареный цыпленок, индейка, грудка индейки, куриная грудка, мясной рулет, ветчина, баранина, сосиски, легкое куриное мясо, куриное темное мясо, курица ножка, жареный цыпленок, жареный цыпленок, куриное бедро, куриные крылышки, утка, гусь, фазан, перепел, голубь n, светлое мясо индейки, темное мясо индейки, грудка индейки, окорочка индейки, крыло индейки, эму, страус, сосиски, паштет, свиная колбаса, салями, колбаса, корейка свиная, фарш, рулька ягненка, телятина, бизон, говяжья печень, говяжьи ребрышкиFast Foodsкартофель фри, пицца, кусок пиццы, хот-дог, гамбургер, чизбургер, большой макинтошМолочные продукты и сыррикотта, йогурт, творог, яйцо, яичные белки, козий сыр, чеддер, сыр чеддер, сыр, яйцо вкрутую, сыр фета, моцарелла, обезжиренный йогуртХлеб, рис, крупы и макаронные изделиярожь, просо, льняное семя, ячмень, гречка, цельнозерновой хлеб, бублик, белый хлеб, хлеб, киноа, кукуруза, спагетти, рис, макароны, коричневый рис, белый рис, вареное яйцо, мексиканский сыр, фруктовый йогуртСпреды, соусы и травыхумус, майонез, масло, арахисовое масло, соевый соус, тимьян, базилик, розмарин, перец, кориандр , укроп, петрушка, специи, соус чилиСладости, десерты и закускипопкорн, чипсы, начос, батончик из мюсли, салти крекеры, чизкейк, желе, мороженое, пончики, шоколад, пудинги, замороженный йогурт, батончик сникерс, круассаны, датское тесто, английские кексы, черничные кексы, пирог, поп-пироги, кофейный кекс, крекеры, картофельные чипсы, рисовые лепешки, тортилья чипсы, помадка, пудинги, сиропы, темный шоколад, мороженое с фруктами, крендели, пирожныеСухие завтракимюсли, чирио, овсяные хлопья, хлопья келлоггс, хлопья General MillsРазноеспирулина, мед, сахар, коричневый сахар, тофу, оливковое масло, чеснок, темпе, овес, заправка для салатов, томатный суп, грибной суп, куриный суп, овощной суп, детское питание, растительное масло, шортенинг, рыбий жир, соевое масло, соевый белок, подливка, изюм, смородина, грушевый бальзам, суккоташ, макароны , яичная лапша, японская лапша, макароны с сыром, буррито |
Наука о томатном кетчупе (и как это делается)
Наверное, то, что я собираюсь сказать, очень спорно, но это всего лишь мое скромное мнение: кетчуп портит гамбургеры и картофель фри.Вроде по-настоящему их разоряет. Такое разочарование — откусить отличный сочный гамбургер и обнаружить, что он буквально покрыт кетчупом. И я обнаружил, что трачу хорошую картошку фри на кетчуп. Тем не менее, наука о кетчупе очень интересна!
В настоящее время кетчуп является основным продуктом питания во многих ресторанах и домах. Большинство из нас знает кетчуп как сладкий томатный соус, идеально подходящий для макания или заправки. Однако кетчуп начинался как более общая приправа, которая даже не обязательно содержала помидоры.Похоже, что англичане обнаружили в Китае ферментированную ароматную приправу столетия назад. Вернувшись домой, они попытались воспроизвести его, и со временем он превратился в (томатный) кетчуп, который мы знаем сейчас.
Ketjap
Доказательство того, что кетчуп появился в азиатских соусах, все еще можно найти в наши дни. В Нидерландах довольно распространен соус кетчап, индонезийский соевый соус. Название при произношении очень похоже на кетчуп!
Свежие помидоры — основной ингредиент кетчупа!Состав кетчупа
Основным ингредиентом современного кетчупа, или томатного кетчупа, являются помидоры.Сумма зависит от марки и типа. Помидоры моют и мелко измельчают до получения однородной консистенции. Попутно с помидоров удаляется часть воды, чтобы соус загустел. Поскольку помидоры содержат в основном воду, около 95% помидоров составляет вода, поэтому вес исходных помидоров может быть выше, чем фактический вес помидоров в готовом кетчупе. Например, из 148 г помидоров изготавливается 100 г определенного томатного кетчупа Heinz (февраль 2018 г., веб-сайт магазина Albert Heijn).
Вы можете подумать, что кетчуп — хороший способ израсходовать старые некачественные помидоры. Однако часто бывает наоборот. Помидоры для кетчупа можно собирать в расцвете сил, так как время, необходимое для того, чтобы добраться от урожая до производственного объекта, зачастую очень короткое. Им не нужно учитывать поездки томатов в разные магазины и т. Д. Крупные бренды томатного кетчупа часто имеют собственные контракты с производителями томатов на выращивание помидоров специально для их кетчупа.
Кроме помидоров, в кетчупе еще больше, иначе это была бы просто томатная паста или соус.Два других основных ингредиента, как правило, — это уксус и сахар. И то, и другое необходимо для того, чтобы помидоры не испортились и кетчуп можно было хранить достаточно долго. Вы также часто найдете в кетчупе соль и специи / травы.
Только один из многих способов насладиться кетчупом: с французскими тостами!Кетчуп — способ сохранить помидоры
Свежие помидоры, в отличие, например, от яблок, нельзя хранить очень долго. Если вы когда-либо хранили помидоры дома, вы знаете, что они склонны к размягчению.После того, как кожица станет мягкой, эти области более склонны к росту плесени, из-за чего помидор портится. Так как помидоры не растут круглый год, в сезон бывает много помидоров, а в сезоны — очень мало. Консервирование помидоров — это способ сохранить их круглый год, как и приготовление кетчупа!
Более низкая активность воды помогает сохранить кетчуп
Как мы обсуждали ранее, продукты с низкой активностью воды менее подвержены порче микроорганизмами. Как только активность воды станет достаточно низкой, бактерии и плесень больше не смогут расти в пище.
Активность воды в пище — это мера количества доступной воды в пище. Низкая активность воды (близкая к 0) означает, что воды мало. Высокое значение (близкое к 1) означает, что доступной воды много. Патогенным микроорганизмам (вызывающим заболевание) для нормального роста обычно требуется высокая активность воды, часто> 0,9.
Что вызывает более низкую водную активность кетчупа?
Активность воды в свежем помидоре очень близка к единице, ведь в нем столько воды.Кетчуп имеет активность воды, близкую к 0,93, благодаря добавлению в помидоры как сахара (около 20 мас.%), Так и соли (всего около 1 мас.%). Это все еще достаточно много для роста большинства микроорганизмов. Чтобы остановить рост, потребовалось бы гораздо больше сахара или соли, что сделало бы томатный кетчуп довольно непривлекательным. Тем не менее, безопасность пищевых продуктов часто зависит от теории препятствий, которая создает несколько препятствий, каждое из которых замедляет порчу и вместе предотвращает порчу.
Помидоры сами по себе слегка кислые.Понижение значения pH
Следующим и основным препятствием является низкий уровень pH кетчупа. Низкое значение pH означает, что пища кислая. Помидоры сами по себе уже кислые. Однако у уксуса значение pH несколько ниже, в результате чего конечное значение pH кетчупа составляет примерно 3,9, хотя точное значение будет зависеть от марки и типа. При таком значении pH многие микроорганизмы больше не растут, что является серьезным препятствием для сохранения кетчупа.
Так почему же кетчуп не такой кислый? Это из-за того сахара.Сладость сахара скрывает кислотность уксуса.
Как делают кетчуп
Кетчуп начинается с сортировки и мытья помидоров. С этого момента процесс немного отличается в зависимости от производителя, но состоит из нескольких основных этапов.
Прежде всего, помидор нужно разделить на более мелкие кусочки. Это можно сделать путем измельчения или с помощью гомогенизатора, который измельчает все кусочки помидора. Помидор также можно очистить от кожуры. Пилинг можно проводить с помощью пара.Короткая обработка помидора горячим паром ослабит кожицу и позволит легко удалить ее.
Помидоры должны пройти хотя бы один этап термической обработки. Достаточно теплая термообработка также может быть названа «горячим перерывом», и она дезактивирует любые ферменты в помидоре. В противном случае эти ферменты, которые присутствуют в природе, могли бы расщепить пектин. Пектин — это большая коллоидная молекула, которая делает томатный кетчуп более густым, как и тыквенное пюре. Если все это разложить во время обработки, кетчуп будет намного более водянистым.
Другой термообработкой может быть процесс приготовления. Во время этого процесса томатная паста смешивается с уксусом, сахаром и другими ингредиентами и готовится вместе до получения желаемой консистенции. В процессе приготовления вода может испариться, чтобы сконцентрировать кетчуп, и возможные микроорганизмы могут погибнуть. Процесс горячего перерыва и процесс приготовления не обязательно должны быть одним и тем же процессом.
Вместо того, чтобы начинать со свежих помидоров, производители кетчупа покупают уже приготовленную томатную пасту / пюре и превращают их в кетчуп.В этом случае производителю нужно только смешать пасту с собственными ингредиентами, чтобы приготовить рецепт кетчупа, приготовить его и разлить в упаковку! На видео ниже показан пример такого процесса. Обратите особое внимание на то, насколько густа томатная паста вначале, для этого требуется очень специальное оборудование, которое даже можно закачивать в процесс.
Цвет кетчупа
Кетчуп не требует дополнительных красителей, чтобы стать красным. Использование ярко-красных помидоров даст ярко-красный кетчуп.Этот красный цвет возникает из-за молекулы ликопина, которая входит в состав томатов и делает их красными.
При переработке кетчупа не рекомендуется использовать оборудование для обработки железа из-за риска потери красного цвета. Кроме того, следует ограничить воздействие кислорода, чтобы предотвратить потерю цвета и превращение помидора в коричневый цвет.
Кетчуп выталкивают из тюбика.Поток кетчупа специальный
Если вы когда-нибудь ели кетчуп, вы просто знаете, что он не выходит из бутылки сам по себе, даже если вы перевернете его вверх дном.Довольно странно, правда? Нарезанные кубиками помидоры и томатный сок немедленно вытекут сами по себе. Мы считаем это настолько нормальным, хотя это особое поведение, что вы, возможно, даже не задумывались над этим.
Кетчуп — это так называемая неньютоновская жидкость. Ньютоновская жидкость, такая как вода, имеет постоянную вязкость или «толщину» независимо от того, встряхивается ли она, перемешивается, сжимается и т. Д. Она изменяется только с температурой. Для неньютоновской жидкости это не так, поэтому ее вязкость (поведение потока) изменяется в зависимости от того, как вы прикладываете напряжение, такое как встряхивание или перемешивание.Кетчуп не потечет, когда вы перевернете бутылку, однако, если вы встряхнете ее, она потечет! Кетчуп из-за этого буквально становится тоньше, это то, что мы называем истончением при сдвиге.
Область, изучающая этот тип поведения в пищевых продуктах, называется реология пищевых продуктов и является прекрасным примером области физики пищевых продуктов.
Кетчуп для загустения
Как мы уже говорили в разделе обработки, пектин играет важную роль в загущении кетчупа. Если весь пектин разрушится, кетчуп станет водянистым.Использование помидоров низкого качества или помидоров, которые уже начали портиться, также приведет к получению более водянистого кетчупа. Этого можно избежать, снова добавив загустители, такие как крахмал (например, из кукурузы) или жевательные резинки (аналогичные тем, что в мороженом).
Источники
Химико-генетическая дорожная карта для улучшения томатного вкуса, Denise Tieman et. al, S cience 27 января 2017: Vol. 355, выпуск 6323, стр. 391-394, DOI: 10.1126 / science.aal1556
pH кетчупа, ссылка
Как был изобретен кетчуп, National Geographic, 2014, ссылка
Семь видов кетчупа (кетчупа), которые вы встретите в кулинарной книге XIX века «Кусочки голубого неба», ссылка; отличная статья, копающаяся в разных кэтсупах, большинство из них не томатное
Значения активности воды некоторых пищевых ингредиентов и продуктов, Schmidt, S.J., ссылка, с.412
Совет консультантов и инженеров NPCS, Полная книга по производству томатов и томатных продуктов (выращивание и переработка) (2-е пересмотренное издание), 2017 г., глава 10, ссылка
Фезерстоун, С., Полный курс консервирования и родственных процессов: Том 3 Процедуры обработки консервированных пищевых продуктов, 2015 год, глава 5, ссылка
амилопластов — обзор | ScienceDirect Topics
4 Формирование и увеличение немутантных гранул
Растительные клетки имеют несколько типов пластид, таких как пропластиды, хлоропласты, хлорамилопласты, хромопласты и амилопласты, в зависимости от видов и тканей.Badenhuizen 17 утверждал, что, хотя некоторые пластиды не образуют крахмал в естественных условиях, все они могут быть индуцированы к образованию крахмала путем плавания кусочков ткани в растворе сахара. Хотя крахмал может вырабатываться в различных пластидах при добавлении сахара, хлоропласты и амилопласты являются основными местами накопления крахмала в природе. Переходный крахмал вырабатывается в хлоропластах днем и мобилизуется ночью. В течение продолжительных световых периодов количество мелких гранул в хлоропласте увеличивается, но размер гранул остается относительно небольшим. 17 Контроль синтеза и разложения крахмала в хлоропластах обсуждается в разделе 3.6.
Резервный или запасной крахмал накапливается в специализированных лейкопластах, называемых амилопластами, а иногда и в хлорамилопластах. Амилопласты — это органеллы, ограниченные двойной мембраной, которые развиваются из пропластидов. Duvick 160 изучал раннее развитие пластидов и крахмала в клетках эндосперма кукурузы. Он описал небольшие нити, которые образовывали выступы в клетках эндосперма кукурузы.Затем, по его словам, внутри выступов филаментов образовались гранулы крахмала. Основываясь на наблюдениях под электронным микроскопом, 17,151,161 филаменты, наблюдаемые в живых клетках Duvick 160 , по-видимому, являются пропластидами, развивающимися в амилопласты (нити с выступами). Пропластиды и молодые амилопласты на фиксированных участках имеют очень неправильную форму и, вероятно, принимают различные формы (амебоидные) в живой клетке. 161
Было показано, что внутренняя мембрана молодых амилопластов ячменя 151 и кукурузы 161,162 широко инвагинирована с образованием канальцев, пластинок стромы или пузырьков.Badenhuizen 162 заметил, что гранулы крахмала образуются в «карманах», образованных пластинчатой структурой. Он предположил, что эти карманы необходимы для инициации образования гранул крахмала, возможно, путем стимулирования локально повышенных концентраций ферментов и субстратов. Внутренняя мембрана хлоропластов содержит специфические транслокаторы, необходимые для переноса метаболитов между стромой хлоропласта и цитозолем. 163,164 На основании сходства хлоропластов и амилопластов предполагается, что внутренняя мембрана амилопластов также участвует в регуляции переноса метаболитов.Таким образом, инвагинации внутренней мембраны, указанные выше, могут эффективно увеличивать площадь поверхности мембраны и, возможно, обеспечивать более эффективный перенос субстрата в амилопласты. 151
Tandecarz et al. 165 рассмотрел данные за несколько лет, подтверждающие вывод о том, что биосинтез крахмала включает специфическое инициирующее событие, опосредованное UDP-Glc: протеинтрансглюкозилазой (UPTG) (EC 2.4.1.112), полипептидом с молекулярной массой 38000. UPTG, которому почти абсолютно необходим Mn 2+ , является активным ферментом и в то же время акцептором глюкозила.Полученный глюкозилированный полипептид служит глюкозильным праймером, необходимым для удлинения полимера за счет действия синтазы крахмала.
Предполагается, что инициация и удлинение полисахарида крахмала происходят в строме амилопласта. Строма (основное вещество) амилопластов при исследовании под электронным микроскопом кажется однородным. 17,161 Однако Badenhuizen 17,18,162 обнаружил гранулярные частицы в строме амилопласта ткани, зафиксированные в перманганате калия.Хотя наблюдаемая гранулярная структура могла быть артефактом, вызванным процедурой фиксации, она действительно показывала присутствие материала в строме, который накапливался в амилопластах, а затем уменьшался с образованием гранул крахмала. 18 Накопление и уменьшение этих частиц также происходило во время роста гранул крахмала. 18 Badenhuizen 18 назвал эти частицы коацерватными каплями и предположил, что они прикрепляются к периферии гранулы крахмала.Он пришел к выводу 17,18 , что молекулы крахмала продуцируются в строме амилопласта, а затем завершенные молекулы становятся частью растущей гранулы крахмала. Шеннон и др. 166 подвергали растения кукурузы воздействию 14 CO 2 и определяли распределение 14 C в амилозном и амилопектиновом компонентах крахмала через 1-6 часов. Они обнаружили, что удельная активность ( 14 C / мг полисахарида) амилозы и амилопектина увеличивалась с одинаковой скоростью, и что радиоактивность распределялась по молекулам полисахарида.Они пришли к выводу, что эти данные подтверждают предположение Баденхейзена 17,18 о том, что молекулы крахмала полностью синтезируются в строме амилопласта, а затем откладываются на поверхности гранул. После того, как полисахариды стали частью гранулы, не было доказательств последующего превращения амилозы в амилопектин. 166 Это противоречит выводам, сделанным на основе долгосрочных исследований 14 C-мечения пшеничного крахмала. 167,168 В результате этих исследований оказалось, что сначала синтезируется амилоза, а затем превращается в амилопектин.