| Международное наименование INN:
Group Однокомпонентные препараты торговые наименования препаратов, содержащих только активное вещество ГАММА-АМИНОБУТИРОВАЯ КИСЛОТА
Описания препаратов с недействующими рег. уд. или не поставляемые на рынок РФ |
 , покр. оболочкой, 250 мг: 20, 50 или 100 шт.
, покр. оболочкой, 250 мг: 20, 50 или 100 шт. , покр. пленочной оболочкой, 250 мг: 50 или 100 шт.
, покр. пленочной оболочкой, 250 мг: 50 или 100 шт.АМИНАЛОН таблетки — инструкция по применению, цена, дозировки, аналоги, противопоказания
Действующее вещество
— гамма-аминобутировая кислота (gamma-aminobutyric acid)
Состав и форма выпуска препарата
| Таблетки, покрытые пленочной оболочкой | 1 таб. |
| гамма-аминобутировая кислота | 250 мг |
10 шт. — упаковки ячейковые контурные (1) — пачки картонные.
— упаковки ячейковые контурные (1) — пачки картонные.
10 шт. — упаковки ячейковые контурные (2) — пачки картонные.
10 шт. — упаковки ячейковые контурные (4) — пачки картонные.
10 шт. — упаковки ячейковые контурные (5) — пачки картонные.
10 шт. — упаковки ячейковые контурные (10) — пачки картонные.
Фармакологическое действие
Гамма-аминобутировая кислота (GABA, гамма-аминомасляная кислота) является биогенным амином, восстанавливает процессы метаболизма в головном мозге, способствует утилизации глюкозы мозгом и удалению из него токсических продуктов обмена, обеспечивает нормализацию динамики нервных процессов в головном мозге. Повышает продуктивность мышления, улучшает память, оказывает мягкое психостимулирующее действие, благоприятно влияет на восстановление движений и речи после нарушения мозгового кровообращения. Способствует снижению и стабилизации повышенного АД и уменьшению субъективных симптомов артериальной гипертензии (головокружение, нарушение сна).
Оказывает умеренное антигипоксическое и противосудорожное действие. У больных сахарным диабетом снижает содержание глюкозы, при нормальном содержании глюкозы в крови оказывает обратный эффект (за счет гликогенолиза).
Фармакокинетика
Абсорбция быстрая, достаточно полная. Cmax в плазме крови достигается через 60 мин, затем концентрация быстро снижается. Через 24 ч GABA не определяется в плазме крови.Показания
Состояния после перенесенных нарушений мозгового кровообращения; после черепно-мозговых травм. Атеросклероз мозговых сосудов с явлениями размягчения головного мозга. Алкогольная энцефалопатия и полиневриты. Головная боль, головокружение, нарушения сна при артериальной гипертензии. Детский церебральный паралич; последствия черепно-мозговой родовой травмы у детей старше 3 лет; отставание умственного развития у детей. Кинетозы.Противопоказания
Повышенная чувствительность к GABA.
Дозировка
Суточная доза для взрослых составляет — 3-3.
Для профилактики и лечения кинетозов взрослым — по 500 мг 3 раза/сут в течение 3-4 сут, детям — по 250 мг 3 раза/сут в течение 3-4 сут.
Побочные действия
Возможны: тошнота, рвота, лабильность АД, диспепсия, повышение температуры тела, нарушение сна.Лекарственное взаимодействие
При одновременном применении возможно усиление действия производных бензодиазепина, многих снотворных и противоэпилептических средств.
Особые указания
В первые дни лечения возможны колебания АД.
Беременность и лактация
Противопокзано применение в I триместре беременности и в период грудного вскармливания.
Возможность применения во II и III триместрах беременности врач решает индивидуально.
Применение в детском возрасте
Применение возможно согласно режиму дозирования.
Применение в пожилом возрасте
Применение возможно согласно режиму дозирования.
Описание препарата АМИНАЛОН основано на официально утвержденной инструкции по применению и утверждено компанией–производителем.
Предоставленная информация о ценах на препараты не является предложением о продаже или покупке товара. Информация предназначена исключительно для сравнения цен в стационарных аптеках, осуществляющих деятельность в соответствии со статьей 55 ФЗ «Об обращении лекарственных средств».
Обнаружили ошибку? Выделите ее и нажмите Ctrl+Enter.
Амиодарон таблетки п/о 200мг №10х3 — Планета Здоровья
Наименование
Аминалон табл.по 250мг в конт.яч.уп №10х5
Описание
Таблетки покрытые оболочкой белого или почти белого цвета, двояковыпуклые.
Состав
Одна таблетка содержит: действующего вещества: гамма-аминомасляной кислоты – 250 мг; вспомогательные вещества: сахароза, повидон, магния стеарат, опадрай II (в т. ч. спирт поливиниловый частично гидролизованный, тальк, макрогол 3350 (полиэтиленгликоль), лецитин (соевый), титана диоксид E 171), мука пшеничная хлебопекарная, магния карбонат основной лёгкий, титана диоксид E 171, воск пчелиный жёлтый, парафин твердый, масло подсолнечное рафинированное.
ч. спирт поливиниловый частично гидролизованный, тальк, макрогол 3350 (полиэтиленгликоль), лецитин (соевый), титана диоксид E 171), мука пшеничная хлебопекарная, магния карбонат основной лёгкий, титана диоксид E 171, воск пчелиный жёлтый, парафин твердый, масло подсолнечное рафинированное.
Фармакотерапевтическая группа
Фармакологические свойства
Фармакодинамика
Ноотропное средство, улучшает процессы метаболизма тканей головного мозга, способствует утилизации глюкозы мозгом и удалению из него токсичных продуктов обмена. Повышает продуктивность мышления, улучшает память, оказывает умеренное психостимулирующее действие, благоприятно влияет на восстановление движений и речи после нарушения мозгового кровообращения. Обладает легким гипотензивным действием, снижает исходно повышенное артериальное давление и выраженность обусловленных гипертонией симптомов (головокружение, бессонница), незначительно урежает частоту сердечных сокращений.
Показания к применению
- состояния после перенесенных нарушений мозгового кровообращения и черепно-мозговых травм;
- психоорганический синдром;
- атеросклеротическая, гипертоническая и алкогольная энцефалопатия;
- детский церебральный паралич и последствия родовой травмы у детей в составе комплексной терапии;
- отставание умственного развития у детей;
- кинетозы (морская и воздушная болезнь).
Противопоказания
- повышенная чувствительность к гамма-аминомасляной кислоте и вспомогательным веществам, входящим в состав лекарственного средства;
- острая почечная недостаточность;
- беременность (I триместр), период лактации;
- детский возраст до 6 лет.


Способ применения и дозы
Применяют внутрь до еды. Взрослым и детям старше 7 лет назначают внутрь по 0,5–1,25 г 3 раза в день. Суточная доза – 1,5–3 г. Детям в возрасте 6 лет – 2,0-3,0 г/сут (8-12 таблеток) в 3 приема. Курс лечения не менее 2-3 недель (оптимально 2-4 мес). При необходимости курсы повторяют 1-2 раза в год. При лечении кинетозов назначают взрослым по 0,5 г (2 таблетки), детям по 0,25 г (1 таблетка) 3 раза в день в течение 3-4 сут, предшествующих поездке.
Побочное действие
- тошнота, рвота, диспепсические явления;
- повышение температуры тела, ощущение жара;
- бессонница;
- аллергические реакции;
- лабильность артериального давления.
Взаимодействие с другими лекарственными средствами
При одновременном применении возможно усиление угнетающего воздействия на ЦНС производных бензодиазепина, противосудорожных и седативно-гипнотических средств.
Усиливает эффект гипотензивных препаратов при одновременном с ними применении. При совместном применении с винпоцетином, нимодипином, ницерголином у пожилых лиц возможно развитие гипотонии и усугубление нарушений мозгового кровообращения в связи с развитием гипоперфузии мозга на фоне гипотонии.
При совместном применении с винпоцетином, нимодипином, ницерголином у пожилых лиц возможно развитие гипотонии и усугубление нарушений мозгового кровообращения в связи с развитием гипоперфузии мозга на фоне гипотонии.
Меры предосторожности
Лекарственное средство содержит сахарозу, данное средство не следует принимать пациентам с наследственной непереносимостью фруктозы, глюкозо-галактозной мальабсорбцией или дефицитом сахаразы-изомальтазы. Осторожность при использовании Следует соблюдать особую осторожность при использовании аминалона у пациентов с лабильным АД в первые дни лечения, в связи с возможностью значительного колебания АД.
Беременность и лактация
При беременности и в период лактации безопасность и эффективность применения аминалона не установлены.
Особенности влияния препарата на способность управлять автотранспортом или потенциально опасными механизмами
Во время применения аминалона необходимо соблюдать осторожность при управлении автотранспортом или потенциально опасными механизмами.
Передозировка
Аминалон обладает низкой токсичностью. Симптомы интоксикации возникают лишь при одномоментном применении чрезвычайно высоких доз аминалона (более 10-20 г) и характеризуются следующими симптомами: тошнотой, рвотой, болью в желудке, повышением температуры тела, головной болью, сонливостью и брадикардией. Лечение. Специфического антидота не существует. Меры помощи включают отмену препарата, промывание желудка с активированным углем, назначение обволакивающих средств (диосмектит, слизь крахмала), поддерживающую и симптоматическую терапию, направленную на устранение возникших нарушений.
Упаковка
10 таблеток в контурной ячейковой упаковке из пленки поливинилхлоридной и фольги алюминиевой. 5 или 10 контурных ячейковых упаковок вместе с листком-вкладышем помещают в пачку из картона.
Условия хранения
В защищенном от света и влаги месте, при температуре не выше 25 °С. Хранить в недоступном для детей месте.
Срок годности
3 года. Не использовать после окончания срока годности.
Не использовать после окончания срока годности.
Условия отпуска из аптек
По рецепту.
Каковы механизмы действия и фармакокинетика усилителей ГАМК?
Мольгаард-Нильсен Д., Хвиид А. Противоэпилептические препараты нового поколения и риск серьезных врожденных дефектов. JAMA . 2011 18 мая. 305 (19): 1996-2002. [Медлайн].
Costa J, Fareleira F, Ascencao R, et al. Клиническая сопоставимость новых противоэпилептических препаратов при рефрактерной парциальной эпилепсии: систематический обзор и метаанализ. Эпилепсия . 2011 июл.52 (7): 1280-91. [Медлайн].
Birbeck GL, French JA, Perucca E, Simpson DM, Fraimow H, George JM, et al. Основанное на фактах руководство: Выбор противоэпилептических препаратов для людей с ВИЧ / СПИДом: Отчет Подкомитета по стандартам качества Американской академии неврологии и Специальной целевой группы Комиссии по терапевтическим стратегиям Международной лиги против эпилепсии. Неврология . 2012, 10 января. 78 (2): 139-45. [Медлайн].
Неврология . 2012, 10 января. 78 (2): 139-45. [Медлайн].
Xu T, Bajjalieh SM.SV2 модулирует размер легко высвобождаемого пула секреторных везикул. Nat Cell Biol . 2001 3 августа (8): 691-8. [Медлайн].
Crowder KM, Gunther JM, Jones TA, Hale BD, Zhang HZ, Peterson MR, et al. Аномальная нейротрансмиссия у мышей, лишенных белка синаптических везикул 2A (SV2A). Proc Natl Acad Sci U S A . 1999, 21 декабря. 96 (26): 15268-73. [Медлайн]. [Полный текст].
Janz R, Goda Y, Geppert M, Missler M, Südhof TC.SV2A и SV2B действуют как избыточные регуляторы Ca2 + при высвобождении нейромедиаторов. Нейрон . 1999 24 декабря (4): 1003-16. [Медлайн].
Линч Б.А., Ламбенг Н., Нока К., Кенсель-Хаммес П., Баджалиех С.М., Матань А. и др. Белок синаптических везикул SV2A является местом связывания противоэпилептического препарата леветирацетама. Proc Natl Acad Sci U S A . 2004, 29 июня. 101 (26): 9861-6. [Медлайн]. [Полный текст].
и др. Белок синаптических везикул SV2A является местом связывания противоэпилептического препарата леветирацетама. Proc Natl Acad Sci U S A . 2004, 29 июня. 101 (26): 9861-6. [Медлайн]. [Полный текст].
Noyer M, Gillard M, Matagne A, Hénichart JP, Wülfert E.Новый противоэпилептический препарат леветирацетам (ucb L059), по-видимому, действует через специфический сайт связывания в мембранах ЦНС. евро J Pharmacol . 1995 14 ноября. 286 (2): 137-46. [Медлайн].
Beydoun A, Sachdeo RC, Rosenfeld WE, Krauss GL, Sessler N, Mesenbrink P, et al. Монотерапия окскарбазепином при приступах с частичным началом: многоцентровое двойное слепое клиническое испытание. Неврология . 2000, 27 июня. 54 (12): 2245-51. [Медлайн].
Gelisse P, Genton P, Kuate C, Pesenti A, Baldy-Moulinier M, Crespel A. Ухудшение судорог под действием окскарбазепина при ювенильной идиопатической генерализованной эпилепсии. Эпилепсия . 2004 Октябрь 45 (10): 1282-6. [Медлайн].
Ухудшение судорог под действием окскарбазепина при ювенильной идиопатической генерализованной эпилепсии. Эпилепсия . 2004 Октябрь 45 (10): 1282-6. [Медлайн].
Schachter SC, Vazquez B, Fisher RS, Laxer KD, Montouris GD, Combs-Cantrell DT, et al. Окскарбазепин: двойное слепое рандомизированное плацебо-контролируемое исследование монотерапии парциальных припадков. Неврология . 1999 10 марта. 52 (4): 732-7. [Медлайн].
Tecoma ES. Окскарбазепин. Эпилепсия .1999. 40 Приложение 5: S37-46. [Медлайн].
Каннингтон МС. Обновление Международного реестра беременностей ламотриджина для фонда эпилепсии. Эпилепсия . 2004 ноябрь 45 (11): 1468. [Медлайн].
Мацуо Ф. Ламотриджин. Эпилепсия . 1999. 40 Suppl 5: S30-6. [Медлайн].
[Медлайн].
Matsuo F, Bergen D, Faught E, Messenheimer JA, Dren AT, Rudd GD, et al. Плацебо-контролируемое исследование эффективности и безопасности ламотриджина у пациентов с парциальными припадками.Американский протокол ламотриджина 0,5 Группа клинических испытаний. Неврология . 1993, ноябрь 43 (11): 2284-91. [Медлайн].
Мидор К.Дж., Пенович П. Каков риск возникновения орофациальных расщелин в результате воздействия ламотриджина во время беременности ?. Неврология . 2008 сен 2. 71 (10): 706-7. [Медлайн].
Brodie MJ, Duncan R, Vespignani H, Solyom A, Bitenskyy V, Lucas C. Дозозависимая безопасность и эффективность зонизамида: рандомизированное двойное слепое плацебо-контролируемое исследование у пациентов с рефрактерными парциальными припадками. Эпилепсия . 2005 Январь 46 (1): 31-41. [Медлайн].
[Медлайн].
Чедвик Д. Зонисамид: химия, механизм действия и фармакокинетика . Нью-Джерси: Группа клиницистов; 2001.
Kothare SV, Valencia I, Khurana DS, Hardison H, Melvin JJ, Legido A. Эффективность и переносимость зонисамида при ювенильной миоклонической эпилепсии. Эпилептическое расстройство . 2004 Декабрь 6 (4): 267-70. [Медлайн].
Киллерман М., Бен-Менахем Э.Зонисамид при прогрессирующей миоклонической эпилепсии: длительные наблюдения у семи пациентов. Epilepsy Res . 1998 29 января (2): 109-14. [Медлайн].
Леппик И. Зонисамид Химия, механизм действия и фармакокинетика . Нью-Джерси: Группа клиницистов; 2001. 3-5.
Леппик ИП. Зонисамид. Эпилепсия . 1999. 40 Suppl 5: S23-9. [Медлайн].
1999. 40 Suppl 5: S23-9. [Медлайн].
Масакадзу С. Зонисамид Химия, механизм действия и фармакокинетика .Нью-Джерси: Группа клиницистов; 2001. 10-11.
Моррелл М. Проблемы женщин и использование зонисамида . Нью-Джерси: Группа клиницистов; 2001. 3-5.
Rogawski MA. Разнообразные механизмы противоэпилептических препаратов в стадии разработки. Epilepsy Res . 2006 июнь 69 (3): 273-94. [Медлайн].
Errington AC, Stöhr T, Heers C, Lees G. Исследуемое противосудорожное средство лакозамид избирательно усиливает медленную инактивацию потенциалзависимых натриевых каналов. Мол Фармакол . 2008, январь 73 (1): 157-69. [Медлайн].
Curia G, Biagini G, Perucca E, Avoli M. Лакозамид: новый подход к нацеливанию на потенциал-управляемые натриевые токи при эпилептических расстройствах. Препараты для ЦНС . 2009. 23 (7): 555-68. [Медлайн].
Лакозамид: новый подход к нацеливанию на потенциал-управляемые натриевые токи при эпилептических расстройствах. Препараты для ЦНС . 2009. 23 (7): 555-68. [Медлайн].
Errington AC, Coyne L, Stöhr T, Selve N, Lees G. Поиск механизма действия нового противосудорожного лакозамида. Нейрофармакология . 2006 июн. 50 (8): 1016-29.[Медлайн].
Доти П., Радд Г.Д., Стоер Т., Томас Д. Лакозамид. Нейротерапия . 2007 января, 4 (1): 145-8. [Медлайн].
Бен-Менахем Э, Битон В., Джатузис Д., Абу-Халил Б., Доти П., Радд Г.Д. Эффективность и безопасность перорального лакозамида в качестве дополнительной терапии у взрослых с частичными приступами. Эпилепсия . 2007 июл. 48 (7): 1308-17. [Медлайн].
Halász P, Kälviäinen R, Mazurkiewicz-Beldzinska M, Rosenow F, Doty P, Hebert D, et al. Дополнительный лакозамид при приступах с частичным началом: эффективность и безопасность — результаты рандомизированного контролируемого исследования. Эпилепсия . 2009 Март 50 (3): 443-53. [Медлайн].
Дополнительный лакозамид при приступах с частичным началом: эффективность и безопасность — результаты рандомизированного контролируемого исследования. Эпилепсия . 2009 Март 50 (3): 443-53. [Медлайн].
Чанг С., Сперлинг М.Р., Битон В., Краусс Г., Хеберт Д., Радд Г.Д. и др. Лакозамид в качестве дополнительной терапии при приступах с частичным началом: рандомизированное контролируемое исследование. Эпилепсия . 2010 июн. 51 (6): 958-67. [Медлайн].
Гильото Л. М., Лодденкемпер Т., Гути В. Д., Ротенберг А., Такеока М., Даффи Ф. Х. и др.Опыт применения лакозамида у детей с лекарственно-устойчивой фокальной эпилепсией. Педиатр Нейрол . 2011 июн. 44 (6): 414-9. [Медлайн].
Хейман Э., Лахат Э., Левин Н., Беркович М., Гандельман-Мартон Р. Предварительная эффективность и безопасность лакозамида у детей с рефрактерной эпилепсией. евро J Paediatr Neurol . 2012 16 января (1): 15-9. [Медлайн].
евро J Paediatr Neurol . 2012 16 января (1): 15-9. [Медлайн].
Афра П., Адамолекун Б. Лакозамидное лечение ювенильной миоклонической эпилепсии. Арест . 2012 г., 21 (3): 202-4. [Медлайн].
Мнацаканян Л, Чунг Дж.М., Цимеринов Э.И., Элиашив Д.С. Внутривенное введение лакозамида при рефрактерном бессудорожном эпилептическом статусе. Арест . 2012 г., 21 (3): 198-201. [Медлайн].
Cherry S, Judd L, Muniz JC, Elzawahry H, LaRoche S. Безопасность и эффективность лакозамида в отделении интенсивной терапии. Neurocrit Care . 2012 Апрель 16 (2): 294-8. [Медлайн].
Krauss GL, Klein P, Brandt C, Lee SK, Milanov I, Milovanovic M, et al.Безопасность и эффективность дополнительного ценобамата (YKP3089) у пациентов с неконтролируемыми фокальными припадками: многоцентровое, двойное слепое, рандомизированное, плацебо-контролируемое исследование зависимости реакции от дозы. Ланцет Нейрол . 2019 13 ноября. [Medline].
Ланцет Нейрол . 2019 13 ноября. [Medline].
Xcopri (ценобамат) [вкладыш в упаковке]. Парамус, Нью-Джерси: SK Life Science, Inc., ноябрь 2019 г. Доступно на [Полный текст].
Секейра Д.Дж., Ван Эсс П.Дж., Браун Т.Л. и др. Эффективность найзилама (USL261; назальный спрей с мидазоламом) у пациентов с острыми повторяющимися приступами: результаты рандомизированного клинического исследования фазы 3 ARTEMIS-1 (Abst 3.269). Представлено на ежегодном собрании Американского общества эпилепсии в 2017 г. Вашингтон, округ Колумбия, 1-5 декабря 2017 г.
Валтоко (диазепам для интраназального введения) [вкладыш в упаковке]. Сан-Диего, Калифорния: Neurelis, Inc., январь 2020 г. Доступно на [Полный текст].
Sperling M, Hogan R, Biton V, Tarquinio D, Carrazana E. 12-месячное открытое исследование безопасности повторных доз Valtoco (NRL-1, назальный спрей диазепам) у пациентов с эпилепсией: промежуточный отчет ( П1.5-028). Неврология.2019 Apr 09; 92 (15 доп.). [Полный текст].
12-месячное открытое исследование безопасности повторных доз Valtoco (NRL-1, назальный спрей диазепам) у пациентов с эпилепсией: промежуточный отчет ( П1.5-028). Неврология.2019 Apr 09; 92 (15 доп.). [Полный текст].
Aicardi J, Mumford JP, Dumas C, Wood S. Вигабатрин в качестве начальной терапии инфантильных спазмов: европейский ретроспективный обзор. Сабрил И.С. Группы исследователей и коллег. Эпилепсия . 1996 июл.37 (7): 638-42. [Медлайн].
Мальмгрен К., Бен-Менахем Э., Фризен Л. Визуальная токсичность вигабатрина: эволюция и дозовая зависимость. Эпилепсия . 2001 Май. 42 (5): 609-15. [Медлайн].
Спенс С.Дж., Санкар Р.Дефекты поля зрения и другие офтальмологические нарушения, связанные с вигабатрином. Лекарственная безопасность . 2001. 24 (5): 385-404. [Медлайн].
 webmd.com»> Томас Л., Тримбл М., Шмитц Б., Ринг Х. Вигабатрин и расстройства поведения: ретроспективный обзор. Epilepsy Res . 1996 Сентябрь 25 (1): 21-7. [Медлайн].
webmd.com»> Томас Л., Тримбл М., Шмитц Б., Ринг Х. Вигабатрин и расстройства поведения: ретроспективный обзор. Epilepsy Res . 1996 Сентябрь 25 (1): 21-7. [Медлайн].Backonja M, Beydoun A, Edwards KR, Schwartz SL, Fonseca V, Hes M и др. Габапентин для симптоматического лечения болезненной нейропатии у пациентов с сахарным диабетом: рандомизированное контролируемое исследование. JAMA . 1998, 2 декабря. 280 (21): 1831-6. [Медлайн].
Кондо Т., Фромм Г. Х., Шмидт Б. Сравнение габапентина с другими противоэпилептическими и ГАМКергическими препаратами. Epilepsy Res . 1991, апрель, 8 (3): 226-31. [Медлайн].
Петров О.А., Хайдер Ф., Ротман Д.Л., Мэтсон Р.Х. Влияние габапентина на ГАМК, гомокарнозин и пирролидинон головного мозга у пациентов с эпилепсией. Эпилепсия . 2000 июн. 41 (6): 675-80. [Медлайн].
41 (6): 675-80. [Медлайн].
Бен-Менахем Э.Фармакология прегабалина и ее значение в клинической практике. Эпилепсия . 2004. 45 Suppl 6: 13-8. [Медлайн].
Броди MJ. Прегабалин в качестве дополнительной терапии парциальных припадков. Эпилепсия . 2004. 45 Suppl 6: 19-27. [Медлайн].
Тейлор С.П., Анджелотти Т., Фауман Э. Фармакология и механизм действия прегабалина: субъединица альфа2-дельта (альфа2-дельта) кальциевых каналов как мишень для открытия противоэпилептических препаратов. Epilepsy Res . 2007 Февраль 73 (2): 137-50. [Медлайн].
Эрранте Л.Д., Петров О.А. Острые эффекты габапентина и прегабалина на клеточную ГАМК, глутамат и глутамин в переднем мозге крыс. Арест . 2003 июл.12 (5): 300-6. [Медлайн].
 webmd.com»> Qin N, Yagel S, Momplaisir ML, Codd EE, D’Andrea MR. Молекулярное клонирование и характеристика субъединицы альфа (2) дельта-4 человеческого потенциалзависимого кальциевого канала. Мол Фармакол .2002 Сентябрь 62 (3): 485-96. [Медлайн].
webmd.com»> Qin N, Yagel S, Momplaisir ML, Codd EE, D’Andrea MR. Молекулярное клонирование и характеристика субъединицы альфа (2) дельта-4 человеческого потенциалзависимого кальциевого канала. Мол Фармакол .2002 Сентябрь 62 (3): 485-96. [Медлайн].Дули DJ, Донован С.М., Пагсли Т.А. Стимул-зависимая модуляция высвобождения [(3) H] норэпинефрина из срезов неокортекса крысы габапентином и прегабалином. Дж. Pharmacol Exp Ther . 2000 Декабрь 295 (3): 1086-93. [Медлайн].
Bryans JS, Davies N, Gee NS, Dissanayake VU, Ratcliffe GS, Horwell DC, et al. Идентификация новых лигандов для сайта связывания габапентина на альфа-2-дельта-субъединице кальциевого канала и их оценка в качестве противосудорожных средств. J Med Chem . 1998 21 мая. 41 (11): 1838-45. [Медлайн].
Вартанян М.Г., Радулович Л. Л., Кинсора Дж.Дж., Серпа К.А., Вергнес М., Бертрам Э. и др. Профиль активности прегабалина на моделях эпилепсии и атаксии на грызунах. Epilepsy Res . 2006 Март 68 (3): 189-205. [Медлайн].
Л., Кинсора Дж.Дж., Серпа К.А., Вергнес М., Бертрам Э. и др. Профиль активности прегабалина на моделях эпилепсии и атаксии на грызунах. Epilepsy Res . 2006 Март 68 (3): 189-205. [Медлайн].
French JA, Kugler AR, Robbins JL, Knapp LE, Garofalo EA. Исследование доза-эффект дополнительной терапии прегабалином у пациентов с парциальными припадками. Неврология .2003 27 мая. 60 (10): 1631-7. [Медлайн].
Arroyo S, Anhut H, Kugler AR, Lee CM, Knapp LE, Garofalo EA, et al. Дополнительное лечение прегабалином: рандомизированное двойное слепое плацебо-контролируемое исследование зависимости реакции от дозы у взрослых с парциальными припадками. Эпилепсия . 2004, январь, 45 (1): 20-7. [Медлайн].
Бейдун А., Усман Б.М., Куглер А.Р., Грейнер М.Дж., Кнапп Л.Е., Гарофало Е.А. Безопасность и эффективность двух схем прегабалина для дополнительного лечения парциальной эпилепсии. Неврология . 2005 8 февраля. 64 (3): 475-80. [Медлайн].
Неврология . 2005 8 февраля. 64 (3): 475-80. [Медлайн].
Элджер К.Э., Броди М.Дж., Анхут Х., Ли С.М., Барретт Дж.А. Дополнительное лечение прегабалином у пациентов с парциальными припадками: новая оценка лечения с гибкими и фиксированными дозами в двойном слепом плацебо-контролируемом исследовании. Эпилепсия . 2005 Декабрь 46 (12): 1926-36. [Медлайн].
Лосади Д., Хемминг К., Марсон АГ. Добавка прегабалина при лекарственно-устойчивой парциальной эпилепсии. Кокрановская база данных Syst Rev . 2008 23 января. CD005612. [Медлайн].
Cramer JA, Mattson RH, Bennett DM, Swick CT. Различные концентрации свободной и общей вальпроевой кислоты при монолекарственной и комбинированной терапии. Ther Drug Monit . 1986. 8 (4): 411-5. [Медлайн].
 webmd.com»> Taverna S, Mantegazza M, Franceschetti S, Avanzini G. Вальпроат избирательно снижает постоянную фракцию тока Na + в нейронах неокортекса. Epilepsy Res .1998 Сентябрь 32 (1-2): 304-8. [Медлайн].
webmd.com»> Taverna S, Mantegazza M, Franceschetti S, Avanzini G. Вальпроат избирательно снижает постоянную фракцию тока Na + в нейронах неокортекса. Epilepsy Res .1998 Сентябрь 32 (1-2): 304-8. [Медлайн].Vreugdenhil M, Wadman WJ. Модуляция натриевых токов в нейронах СА1 крысы карбамазепином и вальпроатом после киндлинг-эпилептогенеза. Эпилепсия . 1999 ноябрь 40 (11): 1512-22. [Медлайн].
Мидор К.Дж., Бейкер Г.А., Браунинг Н., Клейтон-Смит Дж., Комбс-Кантрелл Д.Т., Коэн М. и др. Когнитивные функции в возрасте 3 лет после воздействия на плод противоэпилептических препаратов. N Engl J Med .2009 16 апреля. 360 (16): 1597-605. [Медлайн]. [Полный текст].
Мидор К.Дж., Бейкер Г.А., Браунинг Н., Коэн М.Дж., Бромли Р.Л., Клейтон-Смит Дж. И др. Воздействие противоэпилептических препаратов на плод и когнитивные исходы в возрасте 6 лет (исследование NEAD): проспективное обсервационное исследование. Ланцет Нейрол . 2013 марта 12 (3): 244-52. [Медлайн].
Ланцет Нейрол . 2013 марта 12 (3): 244-52. [Медлайн].
Пеллок Дж. М., Броди МД. Фелбамате: обновление 1997 года. Эпилепсия . 1997 Декабрь 38 (12): 1261-4. [Медлайн].
Rho JM, Donevan SD, Rogawski MA.Механизм действия противосудорожного фелбамата: противоположное действие на рецепторы N-метил-D-аспартата и гамма-аминомасляной кислоты A. Энн Нейрол . 1994 Февраль 35 (2): 229-34. [Медлайн].
Wasterlain CG, Adams LM, Wichmann JK, Sofia RD. Фелбамат защищает нейроны CA1 от апоптоза в модели глобальной ишемии песчанок. Инсульт . 1996, 27 июля (7): 1236-40. [Медлайн].
Wallis RA, Panizzon KL. Глициновая реверсия защиты от гипоксии фелбамата. Нейроотчет . 1993. 4 (7): 951-4.
 webmd.com»> Wasterlain CG, Adams LM, Schwartz PH, Hattori H, Sofia RD, Wichmann JK. Постгипоксическое лечение фелбаматом оказывает нейрозащитное действие на крысиной модели гипоксии-ишемии. Неврология . 1993, ноябрь 43 (11): 2303-10. [Медлайн].
webmd.com»> Wasterlain CG, Adams LM, Schwartz PH, Hattori H, Sofia RD, Wichmann JK. Постгипоксическое лечение фелбаматом оказывает нейрозащитное действие на крысиной модели гипоксии-ишемии. Неврология . 1993, ноябрь 43 (11): 2303-10. [Медлайн].Banta JT, Hoffman K, Budenz DL, Ceballos E, Greenfield DS. Предполагаемая двусторонняя острая закрытоугольная глаукома, вызванная топираматом. Ам Дж. Офтальмол .2001 июл.132 (1): 112-4. [Медлайн].
Глаузер Т.А., Левисон П.М., Риттер Ф., Сакдео Р.С. Топирамат при синдроме Леннокса-Гасто: открытое лечение пациентов, завершивших рандомизированное контролируемое исследование. Группа изучения топирамата YL. Эпилепсия . 2000. 41 Приложение 1: S86-90. [Медлайн].
Очоа JG. Зуд — редкий, но неприятный побочный эффект топирамата. Эпилепсия . 2000. 41 (Supp 7):
2000. 41 (Supp 7):
Privitera M, Fincham R, Penry J, Reife R, Kramer L, Pledger G и др.Плацебо-контролируемое исследование диапазона доз топирамата при рефрактерной парциальной эпилепсии с использованием суточных доз 600, 800 и 1000 мг. Topiramate YE Study Group. Неврология . 1996 июн. 46 (6): 1678-83. [Медлайн].
Санкар PS, Паскуале LR, Grosskreutz CL. Увеальный выпот и вторичная закрытоугольная глаукома, связанные с применением топирамата. Arch Ophthalmol . 2001 август 119 (8): 1210-1. [Медлайн].
Shank RP, Gardocki JF, Vaught JL, Davis CB, Schupsky JJ, Raffa RB, et al.Топирамат: доклиническая оценка нового структурно нового противосудорожного средства. Эпилепсия . 1994 март-апрель. 35 (2): 450-60. [Медлайн].
Zona C, Ciotti MT, Avoli M.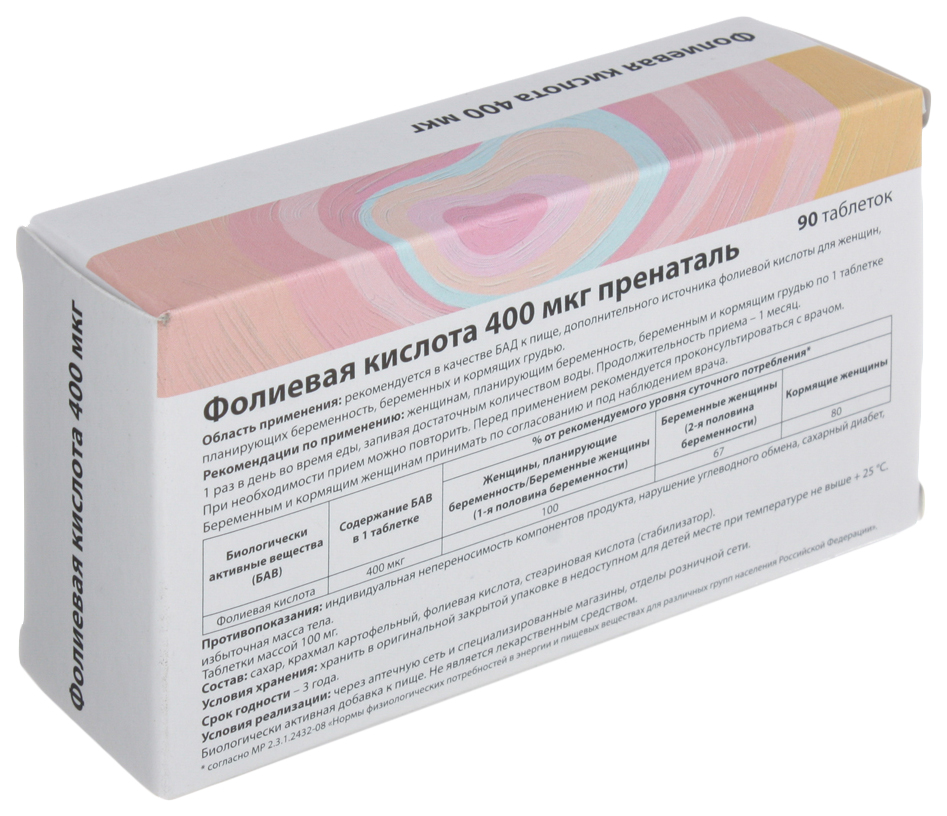 Топирамат ослабляет потенциал-зависимые натриевые токи в гранулярных клетках мозжечка крысы. Neurosci Lett . 1997 15 августа. 231 (3): 123-6. [Медлайн].
Топирамат ослабляет потенциал-зависимые натриевые токи в гранулярных клетках мозжечка крысы. Neurosci Lett . 1997 15 августа. 231 (3): 123-6. [Медлайн].
Krauss GL, Serratosa JM, Villanueva V, Endziniene M, Hong Z, French J, et al. Рандомизированное исследование III фазы 306: дополнительный перампанел при рефрактерных приступах с частичным началом. Неврология . 1 мая 2012 г. 78: 1408-15. [Медлайн].
French JA, Krauss GL, Biton V, Squillacote D, Yang H, Laurenza A, et al. Дополнительный перампанель при рефрактерных приступах с частичным началом: рандомизированное исследование III фазы 304. Неврология . 7 августа. 79: 589-96. [Медлайн].
French JA, Krauss GL, Biton V, Squillacote D, Yang H, Laurenza A, et al. Оценка дополнительного перампанела у пациентов с рефрактерными приступами с частичным началом: результаты рандомизированного глобального исследования III фазы 305. Эпилепсия . 2012 20 августа. 53: [Medline].
Эпилепсия . 2012 20 августа. 53: [Medline].
Fycompa (перампанель) [листок-вкладыш]. Озеро Вудклифф, Нью-Джерси: Eisai Inc., июнь 2015 г. Доступно на [Полный текст].
Fycompa (перампанель) [листок-вкладыш]. Озеро Вудклифф, Нью-Джерси: Eisai Inc., октябрь 2012 г. Доступно на [Полный текст].
Cataldi M, Lariccia V, Secondo A, di Renzo G, Annunziato L. Противоэпилептический препарат леветирацетам снижает инозитол-1,4,5-трифосфат-зависимое увеличение [Ca2 +] I, вызванное АТФ и брадикинином в клетках PC12. Дж. Pharmacol Exp Ther . 2005 г., май. 313 (2): 720-30. [Медлайн].
Cereghino JJ, Biton V, Abou-Khalil B, Dreifuss F, Gauer LJ, Leppik I. Леветирацетам при парциальных припадках: результаты двойного слепого рандомизированного клинического исследования. Неврология . 2000 25 июля. 55 (2): 236-42. [Медлайн].
Неврология . 2000 25 июля. 55 (2): 236-42. [Медлайн].
Gower AJ, Hirsch E, Boehrer A, Noyer M, Marescaux C. Влияние леветирацетама, нового противоэпилептического препарата, на судорожную активность в двух генетических моделях эпилепсии на крысах. Epilepsy Res . 1995 22 ноября (3): 207-13. [Медлайн].
Loscher W, Hönack D, Rundfeldt C. Антиэпилептогенные эффекты нового противосудорожного препарата леветирацетама (ucb L059) в киндлинг-модели височной эпилепсии. Дж. Pharmacol Exp Ther . 1998 Февраль 284 (2): 474-9. [Медлайн].
Патсалос PN. Фармакокинетический профиль леветирацетама: к идеальным характеристикам. Pharmacol Ther . 2000 Февраль 85 (2): 77-85.[Медлайн].
Privitera M. Эффективность леветирацетама: обзор трех основных клинических испытаний. Эпилепсия . 2001. 42 Дополнение 4: 31-5. [Медлайн].
Эпилепсия . 2001. 42 Дополнение 4: 31-5. [Медлайн].
Shorvon SD, Löwenthal A, Janz D, Bielen E, Loiseau P. Многоцентровое двойное слепое рандомизированное плацебо-контролируемое исследование леветирацетама в качестве дополнительной терапии у пациентов с рефрактерными парциальными припадками. Европейская группа изучения леветирацетама. Эпилепсия . 2000 Сентябрь 41 (9): 1179-86.[Медлайн].
Кляйн П., Шиманн Дж., Сперлинг М.Р., Уайтсайдс Дж., Лян В., Сталвей Т. и др. Рандомизированное двойное слепое плацебо-контролируемое многоцентровое исследование в параллельных группах для оценки эффективности и безопасности дополнительного бриварацетама у взрослых пациентов с неконтролируемыми приступами с частичным началом. Эпилепсия . 2015 Декабрь 56 (12): 1890-1898. [Медлайн].
Девинский О. , Патель А.Д., Кросс Дж. Х., Вильянуэва В., Виррелл Е.С., Привитера М. и др.Влияние каннабидиола на припадки при синдроме Леннокса-Гасто. N Engl J Med . 2018 17 мая. 378 (20): 1888-1897. [Медлайн].
, Патель А.Д., Кросс Дж. Х., Вильянуэва В., Виррелл Е.С., Привитера М. и др.Влияние каннабидиола на припадки при синдроме Леннокса-Гасто. N Engl J Med . 2018 17 мая. 378 (20): 1888-1897. [Медлайн].
Thiele EA, Marsh ED, French JA, Mazurkiewicz-Beldzinska M, Benbadis SR, Joshi C, et al. Каннабидиол у пациентов с судорогами, связанными с синдромом Леннокса-Гасто (GWPCARE4): рандомизированное двойное слепое плацебо-контролируемое исследование фазы 3. Ланцет . 2018 17 марта. 391 (10125): 1085-1096. [Медлайн].
Девинский О., Кросс Дж. Х., Локс Л., Марш Е., Миллер И., Наббут Р. и др.Испытание каннабидиола для лекарственно-устойчивых приступов при синдроме Драве. N Engl J Med . 2017 25 мая. 376 (21): 2011-2020. [Медлайн].
Gaston TE, Bebin EM, Cutter GR, Liu Y, Szaflarski JP, UAB CBD Program. Взаимодействие каннабидиола и широко используемых противоэпилептических препаратов. Эпилепсия . 2017 6 августа [Medline].
Взаимодействие каннабидиола и широко используемых противоэпилептических препаратов. Эпилепсия . 2017 6 августа [Medline].
Андерсон П. Каннабидиол уменьшает судороги при различных расстройствах эпилепсии. Новости и перспективы Medscape.Доступно по адресу http://www.medscape.com/viewarticle/872763#vp_3. 05 декабря 2016 г .; Доступ: 25 августа 2017 г.
Диакомит (стирипентол) [вставка в упаковку]. Франция: Биокодекс. Август 2018 г. Доступно по ссылке [Полный текст].
Brodie MJ, Lerche H, Gil-Nagel A, Elger C, Hall S, Shin P, et al. Эффективность и безопасность дополнительного эзогабина (ретигабина) при рефрактерной парциальной эпилепсии. Неврология . 2010 16 ноября. 75 (20): 1817-24. [Медлайн].
Luszczki JJ.Противоэпилептические препараты третьего поколения: механизмы действия, фармакокинетика и взаимодействия. Pharmacol Rep . 2009 март-апрель. 61 (2): 197-216. [Медлайн].
Pharmacol Rep . 2009 март-апрель. 61 (2): 197-216. [Медлайн].
Биохимия, Гамма-аминомасляная кислота, артикул
[1]
Bowery NG, Smart TG, ГАМК и глицин как нейротрансмиттеры: краткая история. Британский журнал фармакологии. 2006, янв [PubMed PMID: 16402094]
[2]
Петров О.А., ГАМК и глутамат в мозге человека.The Neuroscientist: обзорный журнал, посвященный нейробиологии, неврологии и психиатрии. 2002 Dec; [PubMed PMID: 12467378]
[3]
Caputo F, Bernardi M, Лекарства, действующие на систему ГАМК, при лечении больных алкоголизмом. Современный фармацевтический дизайн. 2010 [PubMed PMID: 20482512]
[4]
Брохан Дж. , Гоудра Б.Г., Роль агонистов рецепторов ГАМК в анестезии и седации.Препараты ЦНС. Октябрь 2017 г. [PubMed PMID: 2
, Гоудра Б.Г., Роль агонистов рецепторов ГАМК в анестезии и седации.Препараты ЦНС. Октябрь 2017 г. [PubMed PMID: 2
38]
[6]
Meldrum BS, Глутамат как нейромедиатор в головном мозге: обзор физиологии и патологии. Журнал питания. 2000 Apr; [PubMed PMID: 10736372]
[7]
Löscher W, Основная фармакология вальпроата: обзор после 35 лет клинического использования для лечения эпилепсии.Препараты ЦНС. 2002 [PubMed PMID: 12269861]
[10]
Griffin CE 3rd, Kaye AM, Bueno FR, Kaye AD, Фармакология бензодиазепинов и эффекты, опосредованные центральной нервной системой. Журнал Охзнера. Лето 2013 г. [PubMed PMID: 23789008]
Лето 2013 г. [PubMed PMID: 23789008]
[11]
Pinard A, Seddik R, Bettler B, рецепторы GABAB: физиологические функции и механизмы разнообразия.Достижения в фармакологии (Сан-Диего, Калифорния). 2010; [PubMed PMID: 20655485]
[12]
Ладера С., дель Кармен Годино М., Хосе Кабанеро М., Торрес М., Ватанабе М., Лухан Р., Санчес-Прието Дж. Пресинаптические рецепторы ГАМК подавляют высвобождение глутамата через каналы GIRK в коре головного мозга крыс. Журнал нейрохимии. 2008 декабрь [PubMed PMID: 155]
[13]
Бак LK, Schousboe A, Waagepetersen HS, Цикл глутамат / ГАМК-глутамин: аспекты транспорта, гомеостаз нейротрансмиттеров и перенос аммиака.Журнал нейрохимии. 2006 г., август [PubMed PMID: 16787421]
[14]
Ngo DH, Vo TS, Обновленный обзор фармацевтических свойств гамма-аминомасляной кислоты.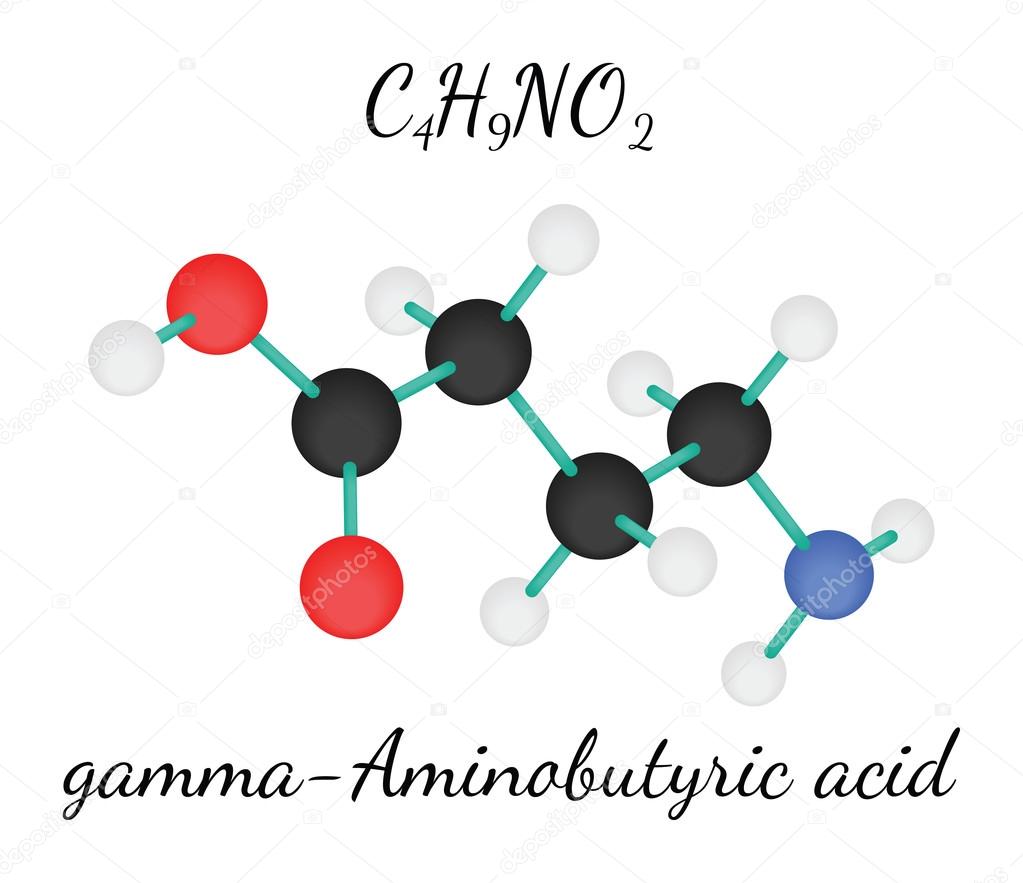 Молекулы (Базель, Швейцария). 24 июля 2019 г. [PubMed PMID: 31344785]
Молекулы (Базель, Швейцария). 24 июля 2019 г. [PubMed PMID: 31344785]
[15]
Ramírez-Jarquín UN, Lazo-Gómez R, Tovar-Y-Romo LB, Tapia R, Спинальные тормозные цепи и их роль в дегенерации двигательных нейронов.Нейрофармакология. 2014 июл [PubMed PMID: 24157492]
[16]
Цзе Ф., Инь Г, Ян В., Ян М., Гао С., Лв Дж., Ли Б. Стресс в регуляции системы миндалины ГАМК и отношение к нейропсихиатрическим заболеваниям. Границы нейробиологии. 2018; [PubMed PMID: 30154693]
[17]
Wong CG, Bottiglieri T, Snead OC 3rd, ГАМК, гамма-гидроксимасляная кислота и неврологические заболевания.Анналы неврологии. 2003 [PubMed PMID: 128]
[18]
Chen ZW, Olsen RW, Белки, ассоциированные с рецептором GABAA: ключевой фактор, регулирующий функцию рецептора GABAA. Журнал нейрохимии. 2007 Янв; [PubMed PMID: 17083446]
[19]
Симеон Т.А., Доневан С.Д., Ро Дж. М., Молекулярная биология и онтогенез рецепторов гамма-аминомасляной кислоты (ГАМК) в центральной нервной системе млекопитающих.Журнал детской неврологии. 2003 Jan; [PubMed PMID: 12661937]
[20]
Хиллмер А.Т., Мейсон Г.Ф., Фусито Л.М., О’Мэлли С.С., Косгроув К.П., Как визуализация глутамата, гамма-аминомасляной кислоты и дофамина может помочь в клиническом лечении алкогольной зависимости и абстиненции. Алкоголизм, клинические и экспериментальные исследования. 2015 декабрь [PubMed PMID: 26510169]
[21]
Chiapponi C, Piras F, Piras F, Caltagirone C, Spalletta G, Система ГАМК при шизофрении и расстройствах настроения: мини-обзор исследований визуализации третьего поколения.Границы психиатрии. 2016 [PubMed PMID: 27148090]
[22]
Батллори М., Молеро-Луис М., Касадо М., Сьерра С., Артуч Р., Ормазабал А. Биохимические анализы спинномозговой жидкости для диагностики нейрометаболических состояний. Чего нам ожидать? Семинары по детской неврологии. 2016 ноя; [PubMed PMID: 28284389]
[23]
Петти F, Концентрации гамма-аминомасляной кислоты (ГАМК) в плазме и расстройства настроения: анализ крови на маниакально-депрессивную болезнь? Клиническая химия.1994, февраль [PubMed PMID: 8313610]
[24]
Дэвис М. Роль рецепторов ГАМКА в опосредовании воздействия алкоголя на центральную нервную систему. Журнал психиатрии [PubMed PMID: 121]
[25]
Bandelow B, Baldwin D, Abelli M, Bolea-Alamanac B, Bourin M, Chamberlain SR, Cinosi E, Davies S, Domschke K, Fineberg N, Grünblatt E, Jarema M, Kim YK, Maron E, Masdrakis V, Mikova O , Nutt D, Pallanti S, Pini S, Ströhle A, Thibaut F, Vaghi MM, Won E, Wedekind D, Wichniak A, Woolley J, Zwanzger P, Riederer P, Биологические маркеры тревожных расстройств, ОКР и ПТСР: согласованное заявление .Часть II: Нейрохимия, нейрофизиология и нейропознание. Всемирный журнал биологической психиатрии: официальный журнал Всемирной федерации обществ биологической психиатрии. 2017 Apr; [PubMed PMID: 27419272]
[27]
Möhler H, Система ГАМК при тревоге и депрессии и ее терапевтический потенциал. Нейрофармакология. 2012, янв [PubMed PMID: 21889518]
[28]
Жиронелл А. Гипотеза ГАМК при эссенциальном треморе: свет и тени.Тремор и другие гиперкинетические движения (Нью-Йорк, Нью-Йорк). 2014 [PubMed PMID: 25120944]
[29]
Gong T, Xiang Y, Saleh MG, Gao F, Chen W., Edden RAE, Wang G, Тормозная моторная дисфункция при подтипах болезни паркинсона. Журнал магнитно-резонансной томографии: JMRI. 2018 июн; [PubMed PMID: 28960581]
[30]
Wu C, Sun D, рецепторы ГАМК в развитии, функции и травмах мозга.Метаболическое заболевание головного мозга. 2015 г., апрель [PubMed PMID: 24820774]
[31]
Hsu YT, Chang YG, Chern Y, Insights of GABA {sub} A {/ sub} ergic system alteration при болезни Хантингтона. Открытая биология. 5 декабря 2018 г. [PubMed PMID: 30518638]
[33]
Leo S, Capo C, Ciminelli BM, Iacovelli F, Menduti G, Funghini S, Donati MA, Falconi M, Rossi L, Malaspina P, дефицит SSADH в итальянской семье: новая мутация гена ALDH5A1, влияющая на сайт связывания субстрата янтарного полуальдегида .Метаболическое заболевание головного мозга. 2017 Октябрь; [PubMed PMID: 28664505]
[34]
Medina-Kauwe LK, Tobin AJ, De Meirleir L, Jaeken J, Jakobs C, Nyhan WL, Gibson KM, дефицит 4-аминобутират аминотрансферазы (ГАМК-трансаминазы). Журнал наследственных нарушений обмена веществ. 1999 июнь [PubMed PMID: 10407778]
[35]
Эрцгаард П., Кампо С., Калабрезе А., Эффективность и безопасность перорального баклофена при лечении спастичности: обоснование интратекального баклофена.Журнал восстановительной медицины. 6 марта 2017 г. [PubMed PMID: 28233010]
[37]
Силлс GJ, Механизмы действия габапентина и прегабалина. Современное мнение в фармакологии. 2006 г., февраль [PubMed PMID: 16376147]
[38]
An H, Godwin J, Флумазенил при передозировке бензодиазепинов.CMAJ: журнал Канадской медицинской ассоциации = journal de l’Association medicale canadienne. 2016 Dec 6; [PubMed PMID: 273]
[39]
Сахинович М.М., Струйс MMRF, Абсалом А.Р., Клиническая фармакокинетика и фармакодинамика пропофола. Клиническая фармакокинетика. 2018 декабрь [PubMed PMID: 30019172]
[40]
Chua HC, Chebib M, GABA {sub} A {/ sub} рецепторы и разнообразие в их структуре и фармакологии.Достижения в фармакологии (Сан-Диего, Калифорния). 2017; [PubMed PMID: 28528665]
Влияние препаратов, усиливающих ГАМК, на людей, зависимых от кокаина — 3 — Полный текст
ГАМК — это тормозящий нейротрансмиттер, который находится в основном в головном мозге. Высокий уровень ГАМК приводит к низкому уровню дофамина, другого нейротрансмиттера, который является химическим веществом «хорошего самочувствия» мозга и играет главную роль в наркомании кокаина. Тягу к кокаину можно уменьшить, снизив уровень дофамина.Это делает препараты, изменяющие ГАМК, потенциальным средством лечения кокаиновой зависимости. В этом исследовании будут задействованы пять препаратов, повышающих уровень ГАМК: тиагабин, топирамат, вальпроевая кислота, баклофен и прогестерон. Цель исследования — оценить взаимодействие между препаратами ГАМК и кокаином с точки зрения безопасности и реакции тяги. В свою очередь, эти результаты могут служить ориентиром для будущих исследований фармакотерапии кокаином.
Это 6-дневное стационарное исследование начнется с ознакомительной сессии для ознакомления участников с процедурами исследования.Вечером первого дня все участники получат плацебо. В дни 2–4 участники получат одно из пяти препаратов ГАМК. Препараты ГАМК будут вводиться в постепенно увеличивающихся дозах для достижения терапевтического уровня при сохранении безопасности и минимизации побочных эффектов. Состоятся три экспериментальных кокаиновых сеанса; один, когда участники получают препарат плацебо, и два, когда участники получают препараты ГАМК. Во время этих сеансов кокаин будет вводиться внутривенно тремя возрастающими дозами, каждая из которых разделена 30-минутными интервалами.Это позволит субъективным и физиологическим реакциям участников во время приема кокаина вернуться к исходным уровням до следующей дозы. Кровь будет отбираться после каждой дозы; ЧСС, артериальное давление и ЭКГ будут контролироваться во время сеансов. В конце каждого сеанса будут разосланы анкеты для оценки эффектов кокаина и связанных с ним состояний настроения, а также для того, чтобы участники могли сообщить о любых побочных эффектах, депрессии, влечении или симптомах отмены.
Это исследование закрыто и опубликовано.
Границы | Препараты рецепторов ГАМК и нейрональная пластичность в поощрении и отвращении: фокус на вентральной тегментальной области
Введение
ГАМК Агонисты рецептора обычно подавляют активность мозга. Бензодиазепины (BZ) и другие ГАМК-миметики подавляют экспрессию генов в головном мозге, включая гены, связанные с нейропластичностью, такие как нейротрофический фактор мозга (BDNF), фактор роста нервов (NGF) и Fos-гены (Zafra et al., 1991; Huopaniemi et al., 2004). Также сообщалось, что BZ и летучий анестетик изофлуран снижают долгосрочную потенциацию (LTP) в нескольких областях мозга (Higashima et al., 1998; Kulisch et al., 2011; Piao et al., 2013). Более того, БЖ и общие анестетики способны нарушать нейрогенез как у молодых, так и у взрослых животных (Wu and Castren, 2009; Erasso et al., 2013; Thal et al., 2014). Все эти факты предполагают, что агонисты рецептора GABA A имеют небольшой потенциал для нейропластичности за пределами критических периодов развития мозга, когда роль ГАМКергических интернейронов является обязательной (Hensch and Stryker, 2004).С другой стороны, подавляя локальную ингибирующую регуляцию, рецепторы ГАМК и на ГАМКергических интернейронах могут косвенно вызывать активацию основных популяций нейронов, сетевой эффект, называемый растормаживанием. Например, ГАМКергические нейроны в латеральной части центрального ядра миндалины возбуждаются низкими острыми дозами BZ посредством растормаживания, и это связано с анксиолитическими эффектами (Beck and Fibiger, 1995; Salminen et al., 1996; Panhelainen and Korpi, 2012). Кроме того, агонисты рецептора GABA A могут активировать дофаминергические (DAergic) нейроны вентральной тегментальной области (VTA) путем растормаживания (Heikkinen et al., 2009; Tan et al., 2010; Ващинкина и др., 2012).
В этом обзоре мы обсудим недавние достижения в понимании того, как модуляторы рецепторов GABA A влияют как на ГАМКергические, так и на глутаматергические синапсы, а также как их однократное или повторное лечение может модулировать пластичность системы вознаграждения. Учитывая широкое применение препаратов рецептора GABA A , более полное понимание того, как они модулируют систему вознаграждения мозга, может помочь в разработке новых стратегий создания новых соединений для преодоления их терапевтических ограничений.
GABA
A Рецепторы и их модуляторыБензодиазепины, ингаляционные и внутривенные анестетики, барбитураты, нейростероиды и другие ГАМК-миметики — все они взаимодействуют с рецептором GABA A и способствуют функции рецептора, вызывая сильные фармакологические и поведенческие эффекты (Sieghart, 1995; Korpi et al., 2002). Они действуют на отдельные участки рецептора GABA A и увеличивают проводимость мембранного аниона (Cl — и бикарбонат), тем самым в большинстве случаев вызывая гиперполяризацию, которая оказывает тормозящее действие на возбуждение постсинаптических нейронов.
Рецепторы GABA A принадлежат к суперсемейству «cys-loop» лиганд-управляемых ионных каналов (Alexander et al., 2013). Они существуют как гетеропентамерные структуры, обычно состоящие из двух субъединиц α, двух субъединиц β и одной субъединицы γ или δ (McKernan and Whiting, 1996). BZ-чувствительные рецепторы GABA A содержат субъединицы α1, α2, α3 и / или α5, и в результате содержания субъединицы γ2 они преимущественно расположены в синаптических сайтах (за исключением α5), тогда как α4 или содержащие субъединицу α6 рецепторы GABA A очень чувствительны к нейростероидам, GABA-миметикам, таким как мусцимол и THIP (габоксадол; 4,5,6,7-тетрагидроизоксазол [4,5-c] пиридин-3-ол), в целом анестетики, такие как изофлуран и этомидат.Поскольку они часто обладают субъединицей δ вместо γ2, они располагаются в пери- или внесинаптических сайтах (Olsen and Sieghart, 2009).
Модуляторы рецепторовGABA A эффективны при широком спектре показаний (тревожные расстройства, паника, бессонница, мышечный спам, контроль припадков при эпилепсии и алкогольной абстиненции, седация агрессивных пациентов, успокоение тревожных пациентов перед операциями и введение анестезии. ), воздействуя на различные подтипы рецепторов GABA A , которые демонстрируют уникальную гетерогенность с точки зрения функции, кинетики, фармакологического профиля и распределения в головном мозге (Uusi-Oukari and Korpi, 2010).Комбинация фармакологических и генетических подходов показала, что 1) рецепторы GABA A , содержащие субъединицу α1, в основном опосредуют седативный и вызывающий привыкание эффекты BZ, 2) рецепторы, содержащие субъединицу α2 или α3, опосредуют анксиолитический и миорелаксантный эффекты и 3 Рецепторы, содержащие субъединицу α5, опосредуют нарушающие память эффекты BZ (Crestani et al., 1999; Rudolph and Mohler, 2004). Кроме того, появляется все больше доказательств того, что внесинаптические рецепторы, содержащие субъединицу α5 гиппокампа, способствуют амнестическому действию общих анестетиков (Grasshoff et al., 2005), тогда как стимуляция рецепторов, содержащих δ-субъединицу, опосредует анксиолитический, противосудорожный, анестезирующий и аверсивный эффекты нейростероидов у мышей (Mihalek et al., 1999; Ващинкина и др., 2014).
Несмотря на полезность модуляторов рецепторов GABA A , их использование может приводить к побочным эффектам, ограничивающим их эффективность. Общие анестетики могут вызывать послеоперационную когнитивную дисфункцию в течение недель или месяцев у пожилых пациентов (Canet et al., 2003; Newman et al., 2007), и их использование при длительных операциях в неонатальном периоде, хотя и необходимо, вызвало серьезные мысли о возможных нейрональных нарушениях. повреждение и долгосрочные когнитивные эффекты, как видно на доклинических моделях (Jevtovic-Todorovic et al., 2013). Помимо краткосрочных нежелательных эффектов BZ, таких как головокружение и нарушение моторики, их долгосрочные эффекты включают нарушение архитектуры сна, спутанность сознания и ухудшение памяти, толерантность и зависимость (Licata and Rowlett, 2008). Кроме того, хроническое повышенное воздействие нейростероидов, по-видимому, ускоряет развитие симптомов болезни Альцгеймера на различных моделях мышей (Bengtsson et al., 2012, 2013). Таким образом, использование этих препаратов вызывает стойкую нейроадаптацию в головном мозге, что также может способствовать возникновению побочных эффектов.
GABA
A Наркотики в моделях зависимости на животныхЗависимость все чаще рассматривается как болезнь аберрантной нейроадаптации в системе вознаграждения мозга (Volkow and Baler, 2014). Дофаминовые нейроны VTA считаются важным центром, по крайней мере, на ранних этапах зависимости. VTA имеет реципрокные связи со многими центрами переднего мозга, такими как прилежащее ядро (NAc, брюшное полосатое тело), дорсальное полосатое тело, медиальная префронтальная кора (mPFC), расширенная миндалевидное тело (особенно включая базолатеральное (BLA) и центральное ядра миндалины и ложа). ядра stria terminalis) и боковые габенулы, цепи которых составляют основную часть путей поощрения и отвращения.
Риск развития зависимости — одна из основных проблем, ограничивающих клиническое использование BZs и других модуляторов GABA A (O’Brien, 2005). Поведение, связанное с зависимостью, также можно изучить на экспериментальных моделях животных. Наркотики, вызывающие злоупотребление, активируют систему вознаграждения и оказывают подкрепляющее действие при самостоятельном введении лекарственного средства, в потенцировании вознаграждения за внутричерепную электрическую самостимуляцию, а также в индуцированном лекарством обучении и кондиционировании. Различные неселективные агонисты BZ, такие как диазепам и мидазолам, вводятся самостоятельно (Tan et al., 2010). Они вознаграждаются в парадигме условного предпочтения места (Spyraki et al., 1985) и способны увеличивать скорость реакции и снижать порог вознаграждения внутричерепной самостимуляции (Straub et al., 2010). Предпочитающий субъединицу α1 агонист золпидем вводится павианом самостоятельно (Ator, 2002), но не вызывает предпочтения места у крыс (Meririnne et al., 1999), демонстрируя, таким образом, несколько неясные эффекты. Это может быть связано с сильным дозозависимым седативным действием золпидема.Содержащие α2-субъединицу рецепторы GABA A в NAc были необходимы для предпочтения мидазолама в тесте, дающем мышам выбор между раствором сахарозы и раствором сахарозы / мидазолама (Engin et al., 2014). Эндогенный нейростероид GABA A , агонист аллопрегнанолон, проявляет различные эффекты в моделях зависимости на животных. Он может поддерживать пероральное самостоятельное введение, хотя и не в действующей обстановке (Sinnott et al., 2002). Это снижает порог вознаграждения за внутричерепную самостимуляцию (Fish et al., 2014), а при внутрибрюшинном введении (в / б) было показано, что он вызывает либо поощрение (Finn et al., 1997), либо отвращение (Beauchamp et al., 2000). Пропофол полезен в условном предпочтении места (Pain et al., 1996), в то время как изофлуран и этотидат, селективный к субъединицам GABA A β2 / 3, до сих пор не тестировался.
В попытках дальнейшей локализации связанных с вознаграждением эффектов модуляторов GABA A в цепи мозга использовались внутримозговые инфузии лекарств.ГАМК Агонист мусцимол и антагонист бикукуллин демонстрируют интересные эффекты, поскольку они оба усиливают и вознаграждают животных, не принимавших лекарства, при введении в VTA (Ikemoto et al., 1997, 1998; Laviolette and van der Kooy, 2001). Эти результаты были объяснены как эффекты, возникающие из-за двух систем вознаграждения, одна из которых является DAergic, а другая не-DAergic. Считается, что мускимол ингибирует нейроны VTA-ГАМК и, таким образом, подавляет нейроны VTA-DA, что приводит к чувствительному к антагонисту DA вознаграждению, в то время как бикукуллин также действует на нейроны VTA-ГАМК, но активирует их для воздействия на плохо определенный не-DAergic путь вознаграждения, нечувствительный к антагонистам DA-рецепторов (Laviolette и ван дер Кой, 2001).Остается изучить, например, действует ли бикукуллин на нейроны VTA GABA, проецируясь на холинергические интернейроны NAc, тем самым усиливая ассоциативное обучение аппетита (Brown et al., 2012). Эти исследования интрацеребральной инфузии должны быть воспроизведены более специфическими в региональном и нейронном отношении методами, такими как хемогенетика или оптогенетика, чтобы избежать возможного затруднения диффузии из места инфузии в другие регионы и в различные популяции нейронов. Кроме того, мозговая схема вознаграждения и отвращения выходит далеко за рамки VTA, что подчеркивает важность системных экспериментов с наркотиками для трансляции.
Пластичность глутаматергических и ГАМКергических синапсов
Все больше данных указывает на то, что нейроадаптация (пластичность) в головном мозге проявляется как в глутаматергических, так и в ГАМКергических синапсах (для обзора см. Castillo et al., 2011; Luscher and Malenka, 2011; Kullmann et al., 2012). Синаптические связи очень пластичны и постоянно модифицируются факторами окружающей среды и учебными задачами. Следует отметить, что генерация новых глутаматергических и ГАМКергических синапсов происходит по разным механизмам с разными факторами, регулирующими процессы (описанные ниже).
Основными местами контакта глутаматергических пресинаптических окончаний являются дендритные шипы (Gray, 1959; Dailey and Smith, 1996; Hering and Sheng, 2001), а динамические изменения в морфологии дендритных шипов были связаны с изменениями в синаптической силе (Segal , 2005). Морфология позвоночника подвержена быстрому изменению паттернами нейрональной активности и активацией постсинаптических рецепторов глутамата (Lang et al., 2004; Matsuzaki et al., 2004). Таким образом, синаптическая эффективность может регулироваться множеством механизмов.
Напротив, преобладающие места контакта для ГАМКергических пресинаптических окончаний расположены не на шипах, а непосредственно на дендритных стержнях (Freund and Buzsaki, 1996; Somogyi et al., 1998). Однако ГАМКергическая система также взаимодействует с синапсами на шипах. Недавние исследования показали, что активность рецепторов GABA A влияет на созревание позвоночника (Heinen et al., 2003; Jacob et al., 2009). В культурах гиппокампа крыс снижение эндоцитоза рецептора GABA A (и повышение активности) связано со снижением созревания позвоночника и снижением уровня белка постсинаптической плотности-95 (PSD-95; Jacob et al., 2009), а в зрительной коре взрослых мышей с нокаутом субъединицы α1 рецептора GABA A (что приводит к усиленному компенсаторному ингибированию) плотность шипов снижается с соответствующим снижением экспрессии PSD-95 (Heinen et al., 2003) , результаты обоих исследований согласуются с активацией рецептора GABA A , снижающей LTP (Higashima et al., 1998). Другое исследование (Shen et al., 2010) показало, что зависимая от развития экспрессия рецепторов GABA A , содержащих субъединицу α4, в дендритных шипах нейронов гиппокампа CA1 играет роль в индукции LTP.Примечательно, что активация этих рецепторов снижает деполяризацию, которая необходима для снятия блокады рецепторов NMDA Mg 2+ , что приводит к снижению LTP. Более того, небольшая популяция ГАМКергических интернейронов, которые экспрессируют соматостатин, заставляет синапсы нацеливаться непосредственно на дендритные головки шипов у мышей mPFC (Chiu et al., 2013), будучи способными контролировать локальные постсинаптические потоки Ca 2+ внутри шипов. Все эти различные механизмы показывают, что активность рецептора GABA A может регулировать быструю динамику и долгосрочные структурные события в глутаматных синапсах.
Сила ГАМКергических синапсов определяется размером выделяемого пула пресинаптических ГАМК и количеством и / или разнообразием рецепторов ГАМК A , сидящих на постсинаптической мембране, что, в свою очередь, в значительной степени определяется переносом рецепторов в плазму и из нее. мембраны, включая введение рецептора, латеральную диффузию внутри мембраны, удаление, рециркуляцию и деградацию (Vithlani et al., 2011). Интересно, что двухфотонная визуализация с высоким разрешением в органотипических культурах гиппокампа показала, что новые ГАМКергические синапсы образуются в результате появления новых бутонов на ранее существовавших пересечениях аксон-дендрит (Wierenga et al., 2008). Следовательно, в этой модели пластичность в ГАМКергических синапсах зависит от количества доступных пересечений аксонов и дендритов, что делает их более ограниченными, чем сайты пластичности в глутаматергических связях. Однако пока неизвестно, могут ли эти результаты in vitro быть обобщены на все ГАМКергические синапсы, и, следовательно, необходимы дальнейшие исследования in vivo .
Медикаментозная пластичность ГАМКергических синапсов
Помимо быстрой модуляции стробирования каналов, как BZ, так и нейростероиды обладают более длительным действием, контролируя количество и подтипы рецепторов GABA A на плазматической мембране (Vithlani et al., 2011; Deeb et al., 2012; Абрамян и др., 2014). Примечательно, что обработка культур гиппокампа флуразепамом, агонистом BZ, модулирует перенос рецептора GABA A , способствуя селективной деградации рецепторов GABA A , содержащих субъединицу α2, после их удаления с постсинаптической мембраны, что приводит к уменьшению размера и количества синапсов, и наконец, к подавлению синаптического торможения (Jacob et al., 2012). Развитие толерантности к седативным эффектам диазепама связано со снижением связывания [ 3 H] L-655,708 с рецепторами, содержащими субъединицу α5 зубчатой извилины гиппокампа (в зависимости от рецепторов, содержащих субъединицу α1), эффект, который был расширяется отсутствием толерантности у мышей с точечными мутантами по α5 субъединице (van Rijnsoever et al., 2004). Обработка C. elegans агонистом ГАМК-сайта мусцимолом также приводит к селективному удалению рецепторов ГАМК А из синапсов (Davis et al., 2010). Напротив, нейростероиды могут избирательно увеличивать трафик внесинаптических рецепторов GABA A путем встраивания новых рецепторов GABA A , содержащих субъединицу α4, в мембрану, что приводит к усилению тонического ингибирования у мышей (Abramian et al., 2014) . Таким образом, адаптация нейронов к препаратам GABA A посредством модуляции переноса рецепторов вызывает долговременные изменения в эффективности ингибирования GABAergic.Однако следует отметить, что эффекты лигандов GABA A на субъединицы рецептора очень сильно зависят от экспериментальной модели, и эффекты, наблюдаемые в моделях клеточных культур, часто бывает трудно воспроизвести in vivo (Uusi-Oukari and Корпи, 2010).
Наиболее очевидным препятствием для понимания нейроадаптации, индуцированной модуляторами рецептора GABA A , является разнообразие их нейронов-мишеней. Фактически, каждый нейрон, контролируемый тормозными терминалами, экспрессирует свою уникальную комбинацию рецепторов GABA A (Luddens et al., 1995; Olsen and Sieghart, 2009). Еще одним препятствием для изучения нейрональной и структурной пластичности, индуцированной лекарственными средствами, является тот факт, что интернейроны, являющиеся основной мишенью для модуляторов рецептора GABA A , сами иннервируются как глутаматергическими, так и GABA-ергическими синапсами. Кроме того, эндогенные модуляторы рецепторов GABA A , такие как нейростероиды, уровни которых постоянно колеблются, например, во время стресса, менструального цикла и развития, могут напрямую влиять на память и процесс обучения (Maguire et al., 2005; Shen et al., 2010), а также влияют на действие других модуляторов рецептора GABA A .
KCC2-опосредованный морфогенез позвоночника
Котранспортер 2 хлорида калия (KCC2) экспрессируется в нейронах, создавая движущую силу для перемещения ионов хлора в клетку через анионный канал рецептора GABA A , что затем приводит к гиперполяризующему действию GABA (Rivera et al. , 1999). Независимо от его транспортной функции Cl —, KCC2 также привлек внимание благодаря своей структурной роли как в глутаматергических, так и в ГАМКергических синапсах (Li et al., 2007; Хорн и др., 2010; Sun et al., 2013). В глутаматергических синапсах KCC2, расположенный в шейке и головке дендритных шипов, связывается с актиновым цитоскелетом посредством линкерного белка 4.1 N (Li et al., 2007). Хотя точные молекулярные механизмы все еще остаются неуловимыми, взаимодействие KCC2 с цитоскелетом является критическим для созревания шипов и для стабильности кластеров рецепторов AMPA (Li et al., 2007; Gauvain et al., 2011). В отличие от глутаматергических синапсов, ГАМКергические синапсы обычно располагаются непосредственно на дендритном стволе (Freund, Buzsaki, 1996; Somogyi et al., 1998). Экспрессия KCC2 регулируется молекулой клеточной адгезии нейролигин-2, которая в основном локализована в ГАМКергических синапсах. Нокдаун нейролигина-2 подавляет экспрессию KCC2 и снижает ГАМКергический синаптогенез и, что интересно, влияя на уровни KCC2, он также подавляет количество глутаматергических синапсов (Sun et al., 2013).
Хотя пока нет прямых доказательств структурных изменений, вызванных лекарствами через KCC2-зависимые механизмы, несколько исследований продемонстрировали изменения в уровнях экспрессии KCC2.Хроническое лечение BZ агонистом золпидемом усиливает экспрессию KCC2 в лимбическом переднем мозге мышей (Shibasaki et al., 2013). Также нейростероид аллопрегнанолон временно изменяет экспрессию KCC2 и уровни белка во время созревания мозга у самцов крыс (Modol et al., 2014). Однако лечение общими анестетиками, мидазоламом, пропофолом и кетамином не изменяет экспрессию KCC2 у крыс в течение первых двух постнатальных недель, когда происходит созревание экспрессии KCC2 (Lacoh et al., 2013). Дальнейшие исследования должны быть направлены на выяснение того, играют ли опосредованные KCC2 механизмы роль в нейрональной пластичности, вызванной ГАМКергическими препаратами (Kang et al., 2006), и какие конкретные изоформы KCC2 играют роль в структурных изменениях, поскольку недавнее исследование предполагает, что KCC2a и изоформы KCC2b имеют разное региональное распределение мозга и, вероятно, разные роли в функциях нейронов (Markkanen et al., 2014).
Медикаментозная синаптическая пластичность в VTA
VTA широко изучена, учитывая ее фундаментальную роль в мотивации и вознаграждении (см. Обзор Luscher and Malenka, 2011).Нейроны VTA DA проецируются в основном в NAc и mPFC и менее широко в гиппокамп и миндалину. Они получают глутаматергические входы от многих областей мозга, включая mPFC, латеральный гипоталамус, боковую габенулу и гиппокамп, и в дополнение к локальному ингибирующему контролю от VTA ГАМКергических интернейронов, ГАМКергические входы в DA нейроны исходят от NAc, вентрального паллидума, ядер расширенная миндалина и ростромедиальное тегментальное ядро (Jhou et al., 2009; Omelchenko, Sesack, 2009; Watabe-Uchida et al., 2012).
Глутаматергическая передача в VTA имеет решающее значение для усиливающих эффектов наркотиков, вызываемых злоупотреблением: подавление глутаматергической передачи в VTA снижает вознаграждение кокаина и героина (Xi and Stein, 2002; You et al., 2007) и предотвращает восстановление кокаина- или поиск героина (Bossert et al., 2004; Sun et al., 2005). Глутаматергические синапсы на нейронах VTA DA могут подвергаться как NMDAR-зависимой LTP, так и NMDAR-независимой длительной депрессии (LTD; Jones et al., 2000; Thomas and Malenka, 2003).Ранее предполагалось, что синаптическая пластичность в мезолимбической DA-системе играет роль в процессе усиления лекарственного средства и зависимости. В течение последнего десятилетия накопились данные электрофизиологических исследований, показывающие, что помимо острой активации нейронов VTA DA, злоупотребляющие наркотики также вызывают длительную пластичность в синапсах этих нейронов. Некоторые классические наркотики, вызывающие злоупотребление, такие как кокаин, амфетамин, морфин, никотин и этанол, обладают общей способностью индуцировать NMDAR-зависимый LTP в глутаматергических синапсах нейронов VTA DA посредством вставки новых AMPAR, лишенных субъединицы GluA2 (Ungless et al., 2001; Saal et al., 2003; Люшер, Маленка, 2011).
Было обнаружено, что помимо усиления глутаматергической передачи, различные вызывающие привыкание препараты, такие как морфин, никотин, кокаин и этанол, нарушают ГАМКергическую передачу в нейронах VTA DA, предотвращая LTP ГАМКергических синапсов (LTP GABA ; Nugent et al. др., 2007; Гуань, Е, 2010). В VTA LTP GABA запускается активацией рецептора NMDA в глутаматных синапсах и требует передачи сигналов оксид азота-цГМФ (Nugent et al., 2007; Ньюджент и Кауэр, 2008). Блокада LTP GABA может дополнительно увеличивать высвобождение DA путем подавления локальных нейронов ГАМК (Liu et al., 2000; Nugent and Kauer, 2008; Niehaus et al., 2010). Какие именно ГАМКергические входы участвуют в LTP GABA и индуцируют ли модуляторы рецептора GABA A LTP GABA , до сих пор не исследовано.
GABA
A Бензодиазепиновые рецепторные препараты Лекарства, вызывающие нейрональную пластичность в VTAПоскольку положительные модуляторы бензодиазепинового сайта рецептора ГАМК A обладают хорошо известным потенциалом злоупотребления и действуют как положительные и / или отрицательные (Panlilio et al., 2005) усилителей на различных животных моделях зависимости, их влияние на синаптическую пластичность в нейронах VTA DA недавно было изучено. Действительно, диазепам и золпидем, как и другие наркотики, вызывают пластичность глутаматергических синапсов, контактирующих с нейронами VTA DA (Heikkinen et al., 2009). В частности, BZ индуцировали LTP, что предотвращалось совместным введением антагониста BZ флумазенила и антагониста рецептора NMDA дизоцилпина (MK-801; Heikkinen et al., 2009). BZ-индуцированный LTP в нейронах VTA DA был связан со вставкой новых GluA2-дефицитных рецепторов AMPA, и локальная сеть внутри VTA была достаточной для индукции LTP посредством ингибирования GABAergic интернейронов VTA (Tan et al., 2010). Кроме того, Tan et al. (2010) исследовали мутантную мышь с BZ-нечувствительными α1-субъединицами и обнаружили, что у этих мышей мидазолам не способен ингибировать активацию ГАМКергических интернейронов VTA, подавлять DA нейроны, вызывать пластичность в глутаматергических синапсах или поддерживать лекарственное средство. подкрепляющее поведение.Это предполагает, что BZ-индуцированное растормаживание DA нейронов является ключевым механизмом, участвующим в BZ-индуцированной пластичности в VTA DA нейронах, а также в усилении BZ.
Лекарства, нацеленные на внесинаптическую ГАМК
Рецепторы A вызывают пластичность нейронов VTA DA, но вызывают отвращениеСуществует потребность в новых анксиолитических / седативных препаратах, не имеющих возможности злоупотребления. Один из подходов заключался в нацеливании на ГАМКергическую систему, отдельную от бензодиазепин-чувствительных рецепторов ГАМК А , т.е.е., внесинаптические рецепторы GABA A , содержащие δ-субъединицу (Olsen and Sieghart, 2009). Однако δ-субъединица экспрессируется по пути вознаграждения в VTA, NAc, mPFC и гиппокампе (Pirker et al., 2000; Hortnagl et al., 2013). Было показано, что THIP и мусцимол, которые действуют на сайты агонистов рецептора GABA A с преимущественной активацией высокоаффинных внесинаптических рецепторов (Chandra et al., 2010), увеличивают частоту возбуждения DA нейронов (Waszczak and Walters, 1980). . Таким образом, было необходимо изучить эффекты модуляторов внесинаптической системы GABA A на пластичность пути вознаграждения и возможный усиливающий потенциал этих препаратов.
Однократная доза THIP и другого внесинаптического GABA A модулятор рецептора нейростероид ганаксолон дозозависимо индуцировала аналогичный AMPA-рецептор-опосредованный LTP в нейронах VTA DA посредством основного действия усиленного тонического ингибирования VTA GABAergic интернейронов (Vashchinkina et al., 2012 , 2014). Важно отметить, что пластичность, индуцированная как THIP, так и ганаксолоном, сохранялась по крайней мере в течение шести дней, в то время как BZ вызывали этот эффект только в течение трех дней (Heikkinen et al., 2009). Эффекты THIP и ганаксолона отсутствовали у мышей с нокаутом рецептора δ-GABA A , и оба лечения усиливали выпрямление тока AMPA, указывая на снижение нацеливания на субъединицы GluA2.
Неожиданно, несмотря на вышеупомянутые аналогичные эффекты на нейроадаптации в нейронах VTA DA, модуляторы синаптических и внесинаптических рецепторов GABA A вызывают различное поведение в парадигмах самостоятельного введения лекарства и кондиционирования места, которые ранее были связаны с активностью нейронов DA VTA (обзор см. в Luscher, Malenka, 2011). В то время как активация синаптических рецепторов GABA A , содержащих субъединицу α1 в VTA пероральным мидазоламом, усиливается при самостоятельном введении, активация внесинаптических рецепторов ip THIP или ганаксолоном приводит к поведению избегания, как это наблюдается в условном отвращении к месту, и THIP не вводят самостоятельно ни мыши, ни бабуины (Tan et al., 2010; Ващинкина и др., 2012, 2014). Эти аверсивные эффекты были устранены у мышей с дефицитом δ-субъединицы рецептора GABA A (Ващинкина и др., 2014), что свидетельствует об особой роли этой популяции рецепторов в VTA. Эти поведенческие данные подтверждают гипотезу о том, что активация синаптических и внесинаптических рецепторов GABA A является положительной и отрицательной, соответственно. Это может зависеть от основных целевых областей мозга, как показано внутримозговыми инъекциями мусцимола (см. Ниже), который также преимущественно нацелен на внесинаптические рецепторы (Chandra et al., 2010). При использовании системы кондиционирования места введение мусцимола в оболочку NAc вызывало условное предпочтение места при введении спереди и условное отвращение при введении сзади (Reynolds and Berridge, 2002). Инфузии мусцимола и бикукуллина внутри БЛА не влияли на вознаграждение (Zarrindast et al., 2004; Macedo et al., 2006).
Более того, теперь становится ясно, что отвращающие лекарства или переживания могут быстро активировать определенные DA нейроны в VTA, а также вызвать длительное усиление их глутаматергических синапсов.Фактически об этом уже сообщалось в ранней статье Saal et al. (2003), где авторы показали, что у мышей 5-минутный стресс плавания при 6 ° C в водяной бане вызывает такую же пластичность нейронов VTA DA, как и классические наркотики.
Нацелены ли препараты рецептора GABA
A на одни и те же популяции нейронов VTA DA?DA нейроны во многих отношениях расходятся, например, по своим электрофизиологическим характеристикам, уязвимости к нейродегенерации и регуляции нейропептидами (Короткова и др., 2004; Lammel et al., 2011). В частности, Lammel et al. (2011) показали, что DA нейроны в VTA мыши организованы в анатомические и электрофизиологические субпопуляции внутри DAergic ядер в зависимости от их концевых полей проекции. Однако у крыс нейроны VTA могут быть более гетерогенными (Margolis et al., 2006, 2012). Ингибирующий контроль интернейронов VTA GABA является важным регулятором активности нейронов VTA DA и следующего поведенческого результата. Было показано, что отвращающие стимулы увеличивают возбуждение нейронов VTA GABA (Creed et al., 2014). Однако, в то время как нейроны ГАМК в VTA, по-видимому, довольно точно реагируют на отвращение посредством возбуждения, ответы в нейронах VTA DA более разнородны: активация, отсутствие ответа или торможение не наблюдаются у обезьян (Matsumoto and Hikosaka, 2009). Вознаграждающие или отталкивающие стимулы могут по-разному модулировать активность DA-нейронов в зависимости от области мозга, на которую эти нейроны проецируются. У мышей опыт употребления кокаина избирательно воздействовал на DA-клетки, проецируемые на медиальную оболочку NAc, в то время как отталкивающий стимул влиял на DA-клетки, проецирующиеся на PFC, а DA-нейроны, проецирующиеся на боковую оболочку NAc, были модифицированы как поощрительными, так и отталкивающими стимулами, что позволяет предположить, что мезокортиколимбическая DA-система состоит из анатомически различных цепей, модифицированных различной мотивационной релевантностью (Lammel et al., 2011). У анестезированных крыс шок стопы ингибировал DA нейроны в дорсальной VTA, тогда как DA нейроны в вентральной VTA становились фазически возбужденными (Brischoux et al., 2009). У мышей большинство DA нейронов VTA снижало возбуждение при пугающих событиях, но небольшая группа DA нейронов была активирована (Wang and Tsien, 2011). В другом исследовании сообщалось, что такое же количество DA-нейронов было активировано, ингибировано или не изменено защемлением хвоста, а также было показано, что у мышей с нарушенным NMDA-рецептором-опосредованным контролем DA-нейронов DAergic активация в ответ на аверсивный стимул ослаблялась. приводящее к нарушению аверсивной обусловленности (Zweifel et al., 2011). Эти находки предполагают, что усиление передачи сигналов DA может быть вызвано стимулами, имеющими мотивационное отношение либо к положительному, либо к отталкивающему направлению, и они указывают на предполагаемые множественные популяции нейронов DA DA с различными афферентными и эфферентными связями.
DA-нейроны мыши с выраженным катионным током, активируемым гиперполяризацией. I ч. -ток обнаруживается в латеральном VTA и проецируется на латеральную оболочку NAc, в то время как DA-нейроны медиального заднего VTA проецируются в mPFC и медиальный NAc. раковины, и не имеют или имеют очень маленькие I h -токи (Lammel et al., 2011). Следует отметить, что исследование, показывающее BZ-индуцированную глутаматергическую пластичность в VTA DA нейронах, использовало большой I h -ток в качестве маркера DA нейронов (Heikkinen et al., 2009). Таким образом, в основном изучалась субпопуляция нейронов VTA DA, которые проецировались на латеральную оболочку NAc. В исследованиях с THIP и ганаксолоном использовали генетически модифицированную линию мышей с флуоресцентным белковым маркером, экспрессируемым в тирозингидроксилаза-положительных нейронах (Ващинкина и др., 2012, 2014). Это позволяло также регистрировать DA нейроны в более медиальных областях VTA с небольшими или отсутствующими I h -токами и проецироваться на mPFC. Возможно, что BZ являются положительно подкрепляющими из-за в основном активации и модификации DA нейронов, обнаруживающих вознаграждение, тогда как препараты, активирующие внесинаптические рецепторы GABA A , вызывают условное отвращение, потому что они активируют и индуцируют LTP в DA нейронах, участвующих в отрицательной мотивации. Эту гипотезу еще предстоит тщательно проверить, хотя последующее исследование участков регистрации пластичности DA нейронов с помощью THIP не выявило какой-либо анатомической локализации в пределах VTA (Ващинкина и др., 2012).
И BZ, и внесинаптические модуляторы GABA A вызывают сильное ингибирование интернейронов VTA GABA (Heikkinen et al., 2009; Tan et al., 2010; Vashchinkina et al., 2012, 2014). Таким образом, подобный растормаживающий механизм, как полагают, вызывает активацию и постоянную модуляцию нейронов VTA DA обоими классами препаратов GABA A , что, в свою очередь, предполагает, что они могут быть нацелены на разные популяции интернейронов VTA GABA, обогащенных синаптическими и / или внесинаптические рецепторы GABA A .Интересно, что недавнее исследование показывает, что летучий растворитель толуол, который также является положительным модулятором рецепторов GABA A , индуцирует LTP в глутаматергических синапсах нейронов VTA DA, которые проецируются на ядро и оболочку NAc, тогда как он не может повлиять на синапсы. нейронов, проецирующих mPFC (Beckley et al., 2013). Важно отметить, что люди злоупотребляют толуолом, и в моделях зависимости на грызунах он увеличивает активацию нейронов VTA DA и высвобождение DA в NAc и демонстрирует положительное подкрепление (Lee et al., 2006; Ригель и др., 2007; Lubman et al., 2008). Действие толуола на интернейроны ГАМК до сих пор не изучено.
Таким образом, возможная гетерогенность VTA как основных нейронов DA, так и интернейронов и проекционных нейронов ГАМК в ответах и опосредовании стимулов и влиянии на поведение является важным предметом для будущих исследований. Эта информация потребуется для дальнейшего развития идей рационального лечения зависимости.
Медикаментозная структурная пластичность в ВТА и ее эфферентах
Компоненты базальных ганглиев и мезолимбического пути среднего мозга (VTA и черная субстанция) и полосатого тела (NAc и дорсомедиальные и дорсолатеральные полосы) связаны по спирали, так что вентральные полосатые области выступают в медиальные части DAergic области среднего мозга, которые впоследствии посылает проекции в более дорсальные области полосатого тела, которые, в свою очередь, проецируются на более латеральные области среднего мозга и так далее (Haber et al., 2000). Возникающая привлекательная гипотеза состоит в том, что первоначальная лекарственная пластичность в DAergic среднем мозге, а затем и в вентральном полосатом теле будет задействовать все больше и больше дорсальных полосатого тела во время хронического употребления наркотиков и усиливать связность в этих спиральных проекциях, что приводит к компульсивному поиску наркотиков. проявляется на поздних стадиях зависимости (Grueter et al., 2012). Дорсомедиальная и дорсолатеральная части полосатого тела регулируют, соответственно, целенаправленные и стимульно-ответные привычные движения (Yin and Knowlton, 2006; Redgrave et al., 2010). Большинство нейронов вентрального и дорсального полосатого тела являются проектирующими нейронами со средними шипами (MSN), активность которых в значительной степени зависит от возбуждающих сигналов от кортикальных и лимбических областей (Sesack and Grace, 2010). Таким образом, пластичность возбуждающих синапсов полосатого тела изменила бы работу цепей полосатого тела, то есть как мотивированное, так и компульсивное поведение. Зависимость, по-видимому, связана с исключительно интенсивной синаптической и структурной пластичностью, обусловленной наркотическим опытом, на разных уровнях мезолимбической системы DA (Robinson and Kolb, 1999).
GABA A модуляторами рецепторов злоупотребляют, но на сегодняшний день нет данных о том, способны ли они вызывать изменения плотности нейронального шипа в системе вознаграждения. Более ранняя работа Sarti et al. (2007) продемонстрировали корреляцию индуцированной кокаином пластичности глутаматергических синапсов с увеличением плотности шипов VTA нейронов крыс. В этой работе традиционная пропитка Гольджи-Кокса была использована для изучения дендритных шипов, а морфология клеток была использована для идентификации и подтипа нейронов VTA DA.Поскольку модуляторы рецептора GABA A также вызывают подобную нейропластичность рецептора глутамата в нейронах VTA DA, мы были озадачены вопросом, изменяют ли диазепам и / или THIP морфологию и плотность шипов. Через 24 часа после однократного введения доз диазепама и THIP, которые индуцируют LTP в VTA (Heikkinen et al., 2009; Vashchinkina et al., 2012), мозг мышей подвергали окрашиванию по Гольджи и подсчету позвоночника (рис. 1). В дополнение к VTA, мы проанализировали области CA1 и CA3 гиппокампа, поскольку известно, что BZ вызывают стойкую синаптическую пластичность, особенно в пирамидных нейронах областей CA1 и CA3 (Таблица 1).
РИСУНОК 1. Пирамидные клетки области СА1 гиппокампа проявляют различные морфологические ответы на лечение диазепамом и THIP. THIP увеличивал плотность шипов на базальных вторичных дендритах пирамидных клеток CA1 ( F 2,15 = 9,69, p <0,01), но не в CA3 или VTA ( p > 0,05). Напротив, диазепам (DZ) не обнаружил каких-либо изменений плотности шипов базальных и апикальных дендритных шипов в этих областях ( p > 0.05). Слева схематических нейрона изображают шипы вторичных или базальных дендритных ветвей, которые оценивались здесь. В середине репрезентативные изображения нейронов, окрашенных по Гольджи, на горизонтальных срезах. Вставки — дендриты с шипами. Масштабная линейка 10 мкм. Справа графики, показывающие плотность позвоночника через 24 часа после однократной инъекции DZ или THIP. Среднее значение ± SEM для n = 10, 5 и 5 мышей в группах носителя (Veh), DZ и THIP, соответственно.** p <0,01, (однофакторный дисперсионный анализ с последующим апостериорным тестом Бонферрони ). Мышей C57BL / 6J (возраст 21–26 дней) декапитировали через 24 часа после обработки DZ (5 мг / кг, внутрибрюшинно), THIP (6 мг / кг, внутрибрюшинно) или носителем (Heikkinen et al., 2009; Vashchinkina et al. al., 2012; Все процедуры с животными были одобрены правительством провинции Южная Финляндия). Мозг был быстро препарирован и немедленно помещен в фиксатор Гольджи-Кокса и обработан с использованием набора FD Rapid GolgiStain Kit (FD Neuro Technologies).Затем импрегнированный мозг разрезали на серийные горизонтальные срезы толщиной 80 мкм с использованием криостата (Leica Microsystems, Nussloch, Германия) и окрашивали с использованием протокола набора. Для каждой области мозга плотность шипов на 10 мкм оценивалась в пяти клетках. Примечательно, что в клетках VTA шипы определяли на вторичных дендритах, в пирамидных клетках CA1 и CA3 на базальных дендритных ветвях с использованием 100-кратного объектива микроскопа Zeiss Axioplan (Carl Zeiss MicroImaging GmbH, NA = 1,3, Ph4, DIC, масло) и позднее проанализировано программным обеспечением ImageJ Национальных институтов здравоохранения (http: // rsb.info.nih.gov/ij). Подсчет позвоночника проводился вслепую относительно лечения. Только выступы, которые показывают непрерывность с дендритным стержнем, считались шипами.
ТАБЛИЦА 1. Резюме нейропластичности, индуцированной лекарственными средствами рецептора GABA A .
Мы обнаружили, что количество базальной плотности шипов в нейронах гиппокампа и VTA согласуется с результатами (Sarti et al., 2007). Обработка THIP увеличивала плотность шипов на базальных вторичных дендритах пирамидных клеток CA1 (рис. 1), но не в CA3 или VTA.Напротив, лечение диазепамом не показало никаких изменений плотности шипов базальных дендритных шипов ни в этих областях, ни в апикальных (данные не показаны). Это в целом согласуется с предыдущим исследованием, в котором острое и хроническое лечение BZ не было связано с повышением уровня белков BDNF или c-Fos в гиппокампе (Licata et al., 2013). Поскольку Сарти и др. (2007) обнаружили явное увеличение VTA шипов после дозы кокаина, вызывающей нейропластичность, наши результаты предполагают, что различные механизмы или разные популяции VTA DA нейронов могли быть вовлечены в эффекты различных лигандов GABA A .Дополнительные исследования с использованием ретроградных нейротрейсеров для уточнения мишеней и новых иммунофлуоресцентных методов совместного окрашивания для точного фенотипирования нейронов (Spiga et al., 2011) необходимы для разрешения пораженных нейронов VTA. Несмотря на сравнимые использованные дозы, вызывающие нейропластичность VTA, диазепам и THIP оказывали различное влияние на ремоделирование позвоночника. Эти результаты показывают, что острые эффекты ГАМК-препаратов не всегда сопровождаются изменениями плотности шипованных дендритов в нейронах гиппокампа или ВТА.
Было показано, что повторное воздействие кокаина увеличивает количество молчащих синапсов и плотность дендритных шипов в оболочке NAc (Robinson and Kolb, 1999; Huang et al., 2009; Dobi et al., 2011; Kim et al., 2011). Также MSN спинного стриатума демонстрируют повышенную плотность дендритных шипов после хронического воздействия кокаина (Ren et al., 2010). Через несколько месяцев после многократного воздействия метамфетамина плотность позвоночника увеличилась в MSN дорсолатерального полосатого тела, структура, которая поддерживает привычное поведение, но уменьшилась в дорсомедиальном полосатом теле, что важно для целенаправленных движений (Jedynak et al., 2007). В исследовании на приматах хронического употребления этанола сообщалось об увеличении плотности позвоночника скорлупы (аналог приматов дорсолатерального полосатого тела у грызунов), а также об усилении глутаматергической передачи и повышенной внутренней возбудимости МСН в этой области (Cuzon Carlson et al., 2011). Этот образец структурной пластичности спинного полосатого тела поддерживает концепцию, согласно которой во время прогрессирования зависимости поведение направляется в сторону привычного приема и поиска наркотиков.
Прекращение хронического лечения препаратами, вызывающими привыкание, приводит к гипофункции нейронов VTA DA.Например, отказ от каннабиноидов или морфина глубоко влияет на морфологические характеристики нейронов VTA DA и плотность шипов MSN оболочки NAc (Spiga et al., 2003, 2010). Еще предстоит изучить, изменяет ли хроническое лечение модуляторами рецептора GABA A морфологию нейронов VTA DA или NAc и дорсальных стриатальных MSN в отношении зависимости и / или абстиненции.
Широкое распространение рецепторов GABA A по всему мозгу предполагает более широкое распространение модуляторов рецепторов GABA A на нейропластичность в других областях мозга, таких как NAc, mPFC, гиппокамп и миндалевидное тело (Licata and Rowlett, 2008).Фактически, в нескольких сообщениях показана блокада гиппокампа LTP после острых in vitro эффектов BZs и изофлурана (суммированные в таблице 1). Это согласуется с известным эффектом когнитивной дисфункции, вызванной общими анестетиками, и нарушения памяти из-за BZ у людей (Canet et al., 2003; Newman et al., 2007). С другой стороны, симптомы отмены после повторного приема этих препаратов связаны с синтезом новых глутаматергических рецепторов и потенцированием LTP в гиппокампе и коре головного мозга (Таблица 1).Таким образом, кажется, что глутаматергические рецепторы регулируются по-разному в зависимости от конкретной фазы действия лекарства.
Заключение
ГАМКергическая нейротрансмиссия, как известно, участвует в процессах нейрональной пластичности в критические периоды развития, и препараты, действующие на рецепторы ГАМК A , такие как BZ, считаются скорее как притупляющие нейропластичность, чем вызывающие ее. Однако недавние эксперименты с дофаминовыми системами среднего мозга показали, что препараты GABA A , действующие на различные подтипы рецепторов, вызывают стойкую нейропластичность глутаматных рецепторов нейронов VTA DA, но пока мало доказательств в поддержку широко распространенных структурных изменений, вызванных этими рецепторами. наркотики.Основной механизм того, как препараты GABA A вызывают пластичность, включает растормаживание посредством первичного ингибирования ГАМКергических интернейронов. Вероятно, что потребуются более динамичные методы, такие как микроскопия in vivo на дендритных шипах, чтобы лучше понять эффекты структурной нейропластичности / нейротоксичности общих анестетиков, БЖ и других типов анксиолитиков и седативных средств. Исходя из имеющихся данных, выбор анестетика будет важен для этих будущих экспериментов!
Авторские взносы
Елена Ващинкина, Анна Панхелайнен, Теему Айтта-ахо и Эса Р.Корпи задумал и разработал эксперименты, Елена Ващинкина провела эксперименты и анализ данных, а Елена Ващинкина, Анне Панхелайнен, Теему Айтта-ахо и Эса Р. Корпи написали статью.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Работа поддержана Академией Финляндии (Esa R.Корпи), Фонд Сигрид Джуселиус (Эса Р. Корпи, Теему Аитта-ахо), Финский фонд изучения алкоголя (Елена Ващинкина, Анне Панхелайнен, Теему Айтта-ахо), Исследовательский фонд Орион-Фармос (Эса Р. Корпи, Теему Айтта-ахо), Фонд Джейн и Аатос Эркко (Анне Панхелайнен, Эса Р. Корпи) и Финский культурный фонд (Анне Панхелайнен).
Список литературы
Абрамян А. М., Комененсия-Ортис Э., Моджил А., Вьен Т. Н., Накамура Ю., Мур Ю. Е. и др.(2014). Нейростероиды способствуют фосфорилированию и встраиванию в мембрану внесинаптических рецепторов ГАМК. Proc. Natl. Акад. Sci. США 111, 7132–7137. DOI: 10.1073 / pnas.1403285111
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Александер, С. П., Бенсон, Х. Э., Факценда, Э., Поусон, А. Дж., Шарман, Дж. Л., Спеддинг, М. и др. (2013). Краткое руководство по фармакологии 2013/14: ионные каналы, управляемые лигандами. Br. J. Pharmacol. 170, 1582–1606.DOI: 10.1111 / bph.12446
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Beauchamp, M.H., Ormerod, B.K., Jhamandas, K., Boegman, R.J., и Beninger, R.J. (2000). Нейростероиды и вознаграждение: аллопрегнанолон вызывает у крыс условное отвращение к месту. Pharmacol. Biochem. Behav. 67, 29–35. DOI: 10.1016 / S0091-3057 (00) 00299-9
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бек, К.Х. и Фибигер Х.С. (1995). Условные вызванные страхом изменения в поведении и экспрессии гена c-fos непосредственно на ранней стадии: с предварительной обработкой диазепамом и без нее. J. Neurosci. 15, 709–720.
Pubmed Аннотация | Pubmed Полный текст | Google Scholar
Бекли, Дж. Т., Эвинс, К. Э., Федарович, Х., Гилстрап, М. Дж., И Вудворд, Дж. Дж. (2013). Медиальная префронтальная кора обратно пропорционально регулирует индуцированные толуолом изменения маркеров синаптической пластичности мезолимбических дофаминовых нейронов. J. Neurosci. 33, 804–813. DOI: 10.1523 / JNEUROSCI.3729-12.2013
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бенгтссон, С. К., Йоханссон, М., Бэкстрем, Т., Нич, Р. М., и Ван, М. (2013). Кратковременное, но хроническое повышение уровня аллопрегнанолона вызывает ускоренную патологию БА на двух моделях мышей по-разному. Curr. Alzheimer Res. 10, 38–47.
Pubmed Аннотация | Pubmed Полный текст | Google Scholar
Бенгтссон, С.К., Йоханссон, М., Бэкстрем, Т., и Ван, М. (2012). Хроническое лечение аллопрегнанолоном ускоряет развитие болезни Альцгеймера у мышей AbetaPP (Swe) PSEN1 (DeltaE9). J. Alzheimers Dis. 31, 71–84.
Pubmed Аннотация | Pubmed Полный текст | Google Scholar
Браун, М. Т., Тан, К. Р., О’Коннор, Э. К., Никоненко, И., Мюллер, Д., и Люшер, К. (2012). Вентральная тегментальная область GABA-проекции приостанавливают аккумбальные холинергические интернейроны для улучшения ассоциативного обучения. Природа 492, 452–456. DOI: 10.1038 / природа11657
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кане, Дж., Редер, Дж., Расмуссен, Л. С., Энлунд, М., Койперс, Х. М., Хэннинг, К. Д. и др. (2003). Когнитивная дисфункция после небольших операций у пожилых людей. Acta Anaesthesiol. Сканд. 47, 1204–1210. DOI: 10.1046 / j.1399-6576.2003.00238.x
CrossRef Полный текст | Google Scholar
Чандра, Д., Халонен, Л.M., Linden, A.M, Procaccini, C., Hellsten, K., Homanics, G.E., et al. (2010). Прототипический агонист рецепторов ГАМК (А) мусцимол действует преимущественно через сайты связывания с высоким сродством переднего мозга. Нейропсихофармакология 35, 999–1007. DOI: 10.1038 / npp.2009.203
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Чиу, К. К., Лур, Г., Морс, Т. М., Карневейл, Н. Т., Эллис-Дэвис, Г. К., и Хигли, М. Дж. (2013). Компартментализация ГАМКергического ингибирования дендритными шипами. Наука 340, 759–762. DOI: 10.1126 / science.1234274
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Crestani, F., Lorez, M., Baer, K., Essrich, C., Benke, D., Laurent, J.P., et al. (1999). Снижение кластеризации рецепторов ГАМК приводит к усилению беспокойства и предвзятости в отношении сигналов угрозы. Нат. Neurosci. 2, 833–839. DOI: 10.1038 / 12207
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кузон Карлсон, В.К., Сиболд, Г. К., Хелмс, К. М., Гарг, Н., Одагири, М., Рау, А. Р. и др. (2011). Синаптические и морфологические нейроадаптации скорлупы, связанные с длительным рецидивирующим употреблением алкоголя у приматов. Нейропсихофармакология 36, 2513–2528. DOI: 10.1038 / npp.2011.140
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Дейли М. Э. и Смит С. Дж. (1996). Динамика дендритной структуры в развивающихся срезах гиппокампа. J. Neurosci. 16, 2983–2994.
Google Scholar
Дэвис, К. М., Стерт, Б. Л., Фридман, А. Дж., Ричмонд, Дж. Э., Бессеро, Дж. Л., Грант, Б. Д. и др. (2010). Регулируемый лизосомный трафик как механизм регуляции изобилия рецепторов GABAA в синапсах у Caenorhabditis elegans . Мол. Cell Neurosci. 44, 307–317. DOI: 10.1016 / j.mcn.2010.04.002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Доби, А., Сиболд, Г. К., Кристенсен, К. Х., Бок, Р., и Альварес, В. А. (2011). Вызванная кокаином пластичность прилежащего ядра зависит от клетки и развивается без длительного изъятия. J. Neurosci. 31, 1895–1904. DOI: 10.1523 / JNEUROSCI.5375-10.2011
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Энгин, Э., Бахурин, К. И., Смит, К. С., Хайнс, Р. М., Рейнольдс, Л. М., Танг, В. и др. (2014). Нейронная основа бензодиазепинового вознаграждения: потребность в альфа2-содержащих рецепторах ГАМКА в прилежащем ядре. Нейропсихофармакология 39, 1805–1815. DOI: 10.1038 / npp.2014.41
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Финн Д. А., Филлипс Т. Дж., Окорн Д. М., Честер Дж. А. и Каннингем К. Л. (1997). Благоприятный эффект нейроактивного стероида 3 альфа-гидрокси-5 альфа-прегнан-20-она у мышей. Pharmacol. Biochem. Behav. 56, 261–264. DOI: 10.1016 / S0091-3057 (96) 00218-3
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Рыба, Э.У., Уитмен, Б. Дж., Диберто, Дж. Ф., Робинсон, Дж. Э., Морроу, А. Л., и Маланга, К. Дж. (2014). Эффекты нейроактивного стероида аллопрегнанолона на внутричерепную самостимуляцию у мышей C57BL / 6J. Психофармакология 231, 3415–3423. DOI: 10.1007 / s00213-014-3600-8
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Фройнд, Т. Ф., и Бузаки, Г. (1996). Интернейроны гиппокампа. Гиппокамп 6, 347–470. DOI: 10.1002 / (SICI) 1098-1063 (1996) 6: 4 <347 :: AID-HIPO1> 3.0.CO; 2-я
CrossRef Полный текст
Gauvain, G., Chamma, I., Chevy, Q., Cabezas, C., Irinopoulou, T., Bodrug, N., et al. (2011). Котранспортер K-Cl нейронов KCC2 влияет на содержание постсинаптических рецепторов AMPA и латеральную диффузию в дендритных шипах. Proc. Natl. Акад. Sci. США 108, 15474–15479. DOI: 10.1073 / pnas.1107893108
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Грассхофф К., Рудольф У. и Антковяк Б.(2005). Молекулярные и системные механизмы общей анестезии: концепция «множественных и множественных механизмов». Curr. Opin. Анестезиол. 18, 386–391. DOI: 10.1097 / 01.aco.0000174961.
.dc
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Грей, Э. Г. (1959). Электронная микроскопия синаптических контактов на дендритных шипах коры головного мозга. Nature 183, 1592–1593. DOI: 10.1038 / 1831592a0
CrossRef Полный текст | Google Scholar
Габер, С.Н., Фадж, Дж. Л., и МакФарланд, Н. Р. (2000). Стриатонигростриатальные пути у приматов образуют восходящую спираль от панциря к дорсолатеральному стриатуму. J. Neurosci. 20, 2369–2382.
Pubmed Аннотация | Pubmed Полный текст | Google Scholar
Хазенедер, Р., Крацер, С., фон Мейер, Л., Эдер, М., Кохс, Э., и Раммес, Г. (2009). Изофлуран и севофлуран дозозависимо ухудшают долгосрочную потенциацию гиппокампа. Eur. J. Pharmacol. 623, 47–51.DOI: 10.1016 / j.ejphar.2009.09.022
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хейккинен, А. Э., Мойккинен, Т. П., и Корпи, Э. Р. (2009). Длительная модуляция глутаматергической передачи в дофаминовых нейронах VTA после однократного приема агонистов бензодиазепина. Нейропсихофармакология 34, 290–298. DOI: 10.1038 / npp.2008.89
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хайнен, К., Бейкер, Р. Э., Спайкер, С., Розаль, Т., ван Пелт, Дж., И Брюссаард, А. Б. (2003). Нарушение созревания дендритных шипов в субъединице альфа1 рецептора ГАМКА нокаутирует мышей. Неврология 122, 699–705. DOI: 10.1016 / S0306-4522 (03) 00477-9
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хигасима М., Киношита Х. и Кошино Ю. (1998). Различия в эффектах золпидема и диазепама на повторяющееся ингибирование и долгосрочное потенцирование в срезах гиппокампа крыс. Neurosci. Lett. 3, 77–80. DOI: 10.1016 / S0304-3940 (98) 00178-5
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хорн, З., Рингстедт, Т., Блесс, П., Кайла, К., и Херлениус, Э. (2010). Преждевременная экспрессия KCC2 у эмбриональных мышей нарушает нервное развитие с помощью независимого от транспорта ионов механизма. Eur. J. Neurosci. 31, 2142–2155. DOI: 10.1111 / j.1460-9568.2010.07258.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Hortnagl, H., Тасан, Р. О., Визельталер, А., Кирчмайр, Э., Зигарт, В., и Сперк, Г. (2013). Паттерны экспрессии мРНК и белка для 12 субъединиц рецептора ГАМКА в головном мозге мыши. Неврология 236, 345–372. DOI: 10.1016 / j.neuroscience.2013.01.008
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Huang, Y.H., Lin, Y., Mu, P., Lee, B.R., Brown, T.E., Wayman, G., et al. (2009). При употреблении кокаина in vivo возникают молчащие синапсы. Neuron 63, 40–47.DOI: 10.1016 / j.neuron.2009.06.007
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Huopaniemi, L., Keist, R., Randolph, A., Certa, U., and Rudolph, U. (2004). Диазепам-индуцированная адаптивная пластичность, выявленная с помощью профилирования экспрессии, специфичной для рецептора альфа1-ГАМК. J. Neurochem. 88, 1059–1067. DOI: 10.1046 / j.1471-4159.2003.02216.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Икемото, С., Мерфи, Дж. М., и Макбрайд, У. Дж. (1997). Самостоятельная инфузия антагонистов GABAA непосредственно в вентральную область покрышки и прилегающие области. Behav. Neurosci. 111, 369–380. DOI: 10.1037 / 0735-7044.111.2.369
CrossRef Полный текст | Google Scholar
Иззо, Э., Аута, Дж., Импаньатиелло, Ф., Песолд, К., Гвидотти, А., и Коста, Э. (2001). Изменения декарбоксилазы глутаминовой кислоты и рецептора глутамата во время толерантности и зависимости к бензодиазепинам. Proc. Natl.Акад. Sci. США 98, 3483–3488. DOI: 10.1073 / pnas.051628698
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Джейкоб, Т. К., Михельс, Г., Силаева, Л., Хейдон, Дж., Суккол, Ф., и Мосс, С. Дж. (2012). Лечение бензодиазепином вызывает подтип-специфические изменения в транспортировке рецепторов ГАМК и снижает синаптическое торможение. Proc. Natl. Акад. Sci. США 109, 18595–18600. DOI: 10.1073 / pnas.1204994109
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Джейкоб, Т.К., Ван, К., Витлани, М., Салиба, Р. С., Суккол, Ф., Пангалос, М. Н. и др. (2009). Перенос мембран рецепторов ГАМК регулирует зрелость позвоночника. Proc. Natl. Акад. Sci. США 106, 12500–12505. DOI: 10.1073 / pnas.0
3106Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Евтович-Тодорович В., Абсалом А. Р., Бломгрен К., Брамбринк А., Кросби Г., Калли Д. Дж. И др. (2013). Анестезиологическая нейротоксичность и нейропластичность: отчет и заявление экспертной группы на основе Зальцбургского семинара BJA. Br. J. Anaesth. 111, 143–151. DOI: 10.1093 / bja / aet177
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Джоу, Т. К., Гейслер, С., Маринелли, М., Дегармо, Б. А., и Зам, Д. С. (2009). Мезопонтинное ростромедиальное тегментальное ядро: структура, нацеленная на боковые габенулы, которые проецируются на вентральную тегментальную область Tsai и компактную черную субстанцию. J. Comp. Neurol. 513, 566–596. DOI: 10.1002 / cne.21891
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Джонс, С., Корнблюм, Дж. Л., и Кауэр, Дж. А. (2000). Амфетамин блокирует длительную депрессию синапсов в вентральной тегментальной области. J. Neurosci. 20, 5575–5580.
Google Scholar
Канг, Т.С., Ким, Д.С., Ким, Дж. Э., Квак, С. Е., Ю, К. Ю., Хван, И. К. и др. (2006). Измененная экспрессия котранспортеров K + -Cl- влияет на быстрое подавление парных импульсов во время активации рецепторов ГАМК в гиппокампе песчанок. Brain Res. 1072, 8–14. DOI: 10.1016 / j.brainres.2005.12.002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ким, С., Шин, Дж. К., Юн, Х. С. и Ким, Дж. Х. (2011). Блокада фосфорилирования ERK в прилежащем ядре подавляет экспрессию индуцированной кокаином поведенческой сенсибилизации у крыс. Korean J. Physiol. Pharmacol. 15, 389–395. DOI: 10.4196 / kjpp.2011.15.6.389
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Корпи, Э.Р., Грюндер Г. и Ладденс Х. (2002). Лекарственные взаимодействия на рецепторах ГАМК. Прог. Neurobiol. 67, 113–159. DOI: 10.1016 / S0301-0082 (02) 00013-8
CrossRef Полный текст | Google Scholar
Лако, К. М., Бодоган, Т., Кайла, К., Фьюмелли, Х., и Вуцкитс, Л. (2013). Общие анестетики не нарушают экспрессию котранспортера хлорида калия KCC2 у новорожденных крыс во время всплеска роста мозга. Br. J. Anaesth. 110 (Приложение 1), i10 – i18.DOI: 10.1093 / bja / aet063
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ланг, К., Барко, А., Заблоу, Л., Кандел, Э. Р., Сигельбаум, С. А., и Захаренко, С. С. (2004). Временное расширение синаптически связанных дендритных шипов при индукции долговременной потенциации гиппокампа. Proc. Natl. Акад. Sci. США 101, 16665–16670. DOI: 10.1073 / pnas.0407581101
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лавиолетт, С.Р., и ван дер Кой, Д. (2001). Рецепторы GABAA в вентральной тегментальной области контролируют двунаправленную передачу сигналов вознаграждения между дофаминергической и недофаминергической нервными мотивационными системами. Eur. J. Neurosci. 13, 1009–1015. DOI: 10.1046 / j.1460-9568.2001.01458.x
CrossRef Полный текст | Google Scholar
Ли, Х., Хируг, С., Кай, К., Людвиг, А., Блесс, П., Коликова, Дж. И др. (2007). KCC2 взаимодействует с дендритным цитоскелетом, способствуя развитию позвоночника. Нейрон 56, 1019–1033.DOI: 10.1016 / j.neuron.2007.10.039
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Licata, S.C., Shinday, N.M., Huizenga, M.N., Darnell, S.B., Sangrey, G.R., Rudolph, U., et al. (2013). Изменения нейротрофического фактора головного мозга в гиппокампе мышей после острого, но не повторного лечения бензодиазепинами. PLoS ONE 8: e84806. DOI: 10.1371 / journal.pone.0084806
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лю Ф., Ван, К., Приступа, З. Б., Ю, X. М., Ван, Ю. Т., и Низник, Х. Б. (2000). Прямое соединение белок-белок обеспечивает перекрестную связь между рецепторами дофамина D5 и γ-аминомасляной кислоты. Природа 403, 274–280. DOI: 10.1038 / 35001232
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Маседо К. Э., Мартинес Р. К. и Брандао М. Л. (2006). Обусловленный и безусловный страх, организованный в нижних бугорках, по-разному чувствителен к инъекциям мусцимола в базолатеральное ядро миндалины. Behav. Neurosci. 120, 625–631. DOI: 10.1037 / 0735-7044.120.3.625
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Markkanen, M., Karhunen, T., Llano, O., Ludwig, A., Rivera, C., Uvarov, P., et al. (2014). Распределение изоформ KCC2a и KCC2b нейронов в ЦНС мышей. J. Comp. Neurol. 522, 1897–1914. DOI: 10.1002 / cne.23510
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
МакКернан, Р.М. и Уайтинг П. Дж. (1996). Какие подтипы рецепторов GABAA действительно встречаются в головном мозге? Trends Neurosci. 19, 139–143. DOI: 10.1016 / S0166-2236 (96) 80023-3
CrossRef Полный текст | Google Scholar
Михалек, Р. М., Банерджи, П. К., Корпи, Э. Р., Куинлан, Дж. Дж., Файерстоун, Л. Л., Ми, З. П. и др. (1999). Ослабленная чувствительность к нейроактивным стероидам у мышей с нокаутом δ-субъединицы γ-аминобутирата типа a. Proc. Natl. Акад. Sci. США 96, 12905–12910.DOI: 10.1073 / pnas.96.22.12905
CrossRef Полный текст | Google Scholar
Модол, Л., Касас, К., Ллидо, А., Наварро, X., Палларес, М., и Дарбра, С. (2014). Неонатальное введение аллопрегнанолона или финастерида изменяет экспрессию котранспортера K Cl в гиппокампе на раннем этапе развития у самцов крыс. J. Steroid Biochem. Мол. Биол. 143C, 343–347. DOI: 10.1016 / j.jsbmb.2014.05.002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Мори, К., Тогаши, Х., Кодзима, Т., Мацумото, М., Охаши, С., Уэно, К. и др. (2001). Различные эффекты анксиолитических агентов, диазепама и тандоспирона, агониста 5-HT (1A), на долгосрочную потенциацию гиппокампа in vivo. Pharmacol. Biochem. Behav. 69, 367–372. DOI: 10.1016 / S0091-3057 (01) 00546-9
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ньюман С., Стигалл Дж., Хирани С., Шаэфи С. и Мейз М. (2007). Послеоперационная когнитивная дисфункция после внесердечных операций: систематический обзор. Анестезиология 106, 572–590. DOI: 10.1097 / 00000542-200703000-00023
CrossRef Полный текст | Google Scholar
О’Брайен, К. П. (2005). Использование бензодиазепинов, злоупотребление и зависимость. J. Clin. Психиатрия 66 (Приложение 2), 28–33.
Google Scholar
Панхелайнен, А. Э., Корпи, Э. Р. (2012). Доказательства роли ингибирования орексинергических нейронов в анксиолитических и седативных эффектах диазепама: исследование c-Fos. Pharmacol.Biochem. Behav. 101, 115–124. DOI: 10.1016 / j.pbb.2011.12.011
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Пяо, М. Х., Лю, Ю., Ван, Ю. С., Цю, Дж. П., и Фэн, С. С. (2013). Летучий анестетик изофлуран ингибирует индукцию LTP нейронов CA1 гиппокампа посредством механизмов, опосредованных подтипом α4β2 nAChR. Ann. Пт. Анест. Реаним. 32, e135 – e141. DOI: 10.1016 / j.annfar.2013.05.012
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Пиркер, С., Шварцер, К., Визельталер, А., Зигарт, В., и Сперк, Г. (2000). Рецепторы ГАМК: иммуноцитохимическое распределение 13 субъединиц в головном мозге взрослой крысы. Неврология 101, 815–850. DOI: 10.1016 / S0306-4522 (00) 00442-5
CrossRef Полный текст | Google Scholar
Rammes, G., Starker, L.K., Haseneder, R., Berkmann, J., Plack, A., Zieglgansberger, W., et al. (2009). Анестезия изофлураном обратимо улучшает когнитивные функции и долгосрочную потенциацию (ДП) за счет усиления экспрессии субъединицы рецептора 2B NMDA. Нейрофармакология 56, 626–636. DOI: 10.1016 / j.neuropharm.2008.11.002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Редгрейв П., Койзет В., Комоли Э., МакХаффи Дж. Г., Лериш М., Вотрелль Н. и др. (2010). Взаимодействие между верхним бугром среднего мозга и базальными ганглиями. Фронт. Нейроанат. 4: 132. DOI: 10.3389 / fnana.2010.00132
CrossRef Полный текст | Google Scholar
Рен, З., Sun, W. L., Jiao, H., Zhang, D., Kong, H., Wang, X., et al. (2010). Рецепторы допамина D1 и N-метил-D-аспартата и регулируемая внеклеточными сигналами киназа опосредуют морфологические изменения нейронов, вызванные повторным введением кокаина. Неврология 168, 48–60. DOI: 10.1016 / j.neuroscience.2010.03.034
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Рейнольдс, С. М., и Берридж, К. С. (2002). Положительная и отрицательная мотивация в оболочке прилежащего ядра: двухвалентные рострокаудальные градиенты для GABA-вызванного приема пищи, вкусовые реакции «симпатия» / «антипатия», предпочтение / избегание места и страх. J. Neurosci. 22, 7308–7320.
Pubmed Аннотация | Pubmed Полный текст | Google Scholar
Ригель А.С., Сапата А., Шиппенберг Т.С. и Френч Э.Д. (2007). Злоупотребление толуолом при вдыхании увеличивает высвобождение дофамина в прилежащем ядре за счет прямой стимуляции нейронов вентральной тегментальной области. Нейропсихофармакология 32, 1558–1569. DOI: 10.1038 / sj.npp.1301273
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ривера, К., Voipio, J., Payne, J. A., Ruusuvuori, E., Lahtinen, H., Lamsa, K., et al. (1999). Котранспортер K + / Cl — KCC2 вызывает гиперполяризацию ГАМК во время созревания нейронов. Природа 397, 251–255. DOI: 10.1038 / 16697
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Робинсон, Т. Э., и Колб, Б. (1999). Изменения морфологии дендритов и дендритных шипов в прилежащем ядре и префронтальной коре после многократного лечения амфетамином или кокаином. Eur. J. Neurosci. 11, 1598–1604. DOI: 10.1046 / j.1460-9568.1999.00576.x
CrossRef Полный текст | Google Scholar
Рудольф, У., и Молер, Х. (2004). Анализ функции рецепторов ГАМК и анализ фармакологии бензодиазепинов и общих анестетиков с помощью генетики мышей. Annu. Rev. Pharmacol. Toxicol. 44, 475–498. DOI: 10.1146 / annurev.pharmtox.44.101802.121429
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Шен, Х., Сабаляускас, Н., Шерпа, А., Фентон, А. А., Стельцер, А., Аоки, К. и др. (2010). Решающая роль рецепторов α4βδ GABAA в формировании дефицита обучения в период полового созревания у мышей. Наука 327, 1515–1518. DOI: 10.1126 / science.1184245
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Шибасаки, М., Масукава, Д., Исии, К., Ямагиши, Ю., Мори, Т., и Судзуки, Т. (2013). Участие ко-переносчика K + -Cl- KCC2 в сенсибилизации к морфин-индуцированной гиперлокомотии при хроническом лечении золпидемом в мезолимбической системе. J. Neurochem. 125, 747–755. DOI: 10.1111 / jnc.12258
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Sieghart, W. (1995). Структура и фармакология подтипов рецепторов γ-аминомасляной кислоты. Pharmacol. Ред. 47, 181–234.
Google Scholar
Саймон В., Хапфельмайер Г., Кохс Э., Циглгансбергер В. и Раммес Г. (2001). Изофлуран блокирует синаптическую пластичность в гиппокампе мыши. Анестезиология 94, 1058–1065.DOI: 10.1097 / 00000542-200106000-00021
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Синнотт, Р. С., Марк, Г. П., и Финн, Д. А. (2002). Усиливающие эффекты нейростероида аллопрегнанолона у крыс. Pharmacol. Biochem. Behav. 72, 923–929. DOI: 10.1016 / S0091-3057 (02) 00776-1
CrossRef Полный текст | Google Scholar
Somogyi, P., Tamas, G., Lujan, R., and Buhl, E.H. (1998). Характерные особенности синаптической организации в коре головного мозга. Brain Res. Brain Res. Ред. 26, 113–135. DOI: 10.1016 / S0165-0173 (97) 00061-1
CrossRef Полный текст | Google Scholar
Сонг, Дж., Шен, Г., Гринфилд, Л. Дж. Младший, и Тиц, Э. И. (2007). Глутаматергическая пластичность, вызванная бензодиазепиновой абстиненцией, включает повышающую регуляцию рецепторов α-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты, содержащих GluR1, в нейронах CA1 гиппокампа. J. Pharmacol. Exp. Ther. 322, 569–581. DOI: 10.1124 / jpet.107.121798
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Спига, С., Acquas, E., Puddu, M. C., Mulas, G., Lintas, A., and Diana, M. (2011). Одновременная Гольджи-Кокса и иммунофлуоресценция с использованием конфокальной микроскопии. Brain Struct. Функц. 216, 171–182. DOI: 10.1007 / s00429-011-0312-2
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Спига, С., Серра, Г. П., Пудду, М. К., Фоддай, М., и Дайана, М. (2003). Нарушения в ВТА, вызванные отменой морфина: конфокальная лазерная сканирующая микроскопия. Eur.J. Neurosci. 17, 605–612. DOI: 10.1046 / j.1460-9568.2003.02435.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Tan, K. R., Brown, M., Labouebe, G., Yvon, C., Creton, C., Fritschy, J. M., et al. (2010). Нейронные основы аддиктивных свойств бензодиазепинов. Природа 463, 769–774. DOI: 10.1038 / nature08758
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Thal, S.C., Timaru-Kast, R., Wilde, F., Merk, P., Johnson, F., Frauenknecht, K., et al. (2014). Пропофол ухудшает нейрогенез и неврологическое восстановление и увеличивает уровень смертности взрослых крыс после черепно-мозговой травмы. Крит. Care Med. 42, 129–141. DOI: 10.1097 / CCM.0b013e3182a639fd
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
van Rijnsoever, C., Tauber, M., Choulli, M.K., Keist, R., Rudolph, U., Mohler, H., et al. (2004). Потребность в рецепторах α5-ГАМКА для развития толерантности к седативному действию диазепама у мышей. J. Neurosci. 24, 6785–6790. DOI: 10.1523 / JNEUROSCI.1067-04.2004
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ващинкина Э., Маннер А. К., Вековищева О., ден Холландер Б., Ууси-Оукари М., Аитта-Ахо Т. и др. (2014). Агонист нейростероидов рецептора GABAA вызывает стойкую нейропластичность в дофаминовых нейронах VTA. Нейропсихофармакология 39, 727–737. DOI: 10.1038 / npp.2013.258
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ващинкина, Е., Panhelainen, A., Vekovischeva, O.Y., Aitta-aho, T., Ebert, B., Ator, N.A., et al. (2012). Габоксадол, агонист сайта ГАМК, вызывает устойчивые изменения в дофаминовых нейронах вентральной тегментальной области, но не оказывает положительного воздействия на мышей или павианов. J. Neurosci. 32, 5310–5320. DOI: 10.1523 / JNEUROSCI.4697-11.2012
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ватабэ-Учида, М., Чжу, Л., Огава, С. К., Ваманрао, А., и Учида, Н.(2012). Картирование всего мозга прямых входов в дофаминовые нейроны среднего мозга. Нейрон 74, 858–873. DOI: 10.1016 / j.neuron.2012.03.017
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Сян К. и Тиц Э. И. (2007). Бензодиазепин-индуцированная пластичность рецептора CA1 гиппокампа α-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты (AMPA), связанная с тяжестью синдрома отмены: дифференциальная роль потенциалзависимых кальциевых каналов и N-метил-D-аспарагиновой кислоты кислотные рецепторы. Behav. Pharmacol. 18, 447–460. DOI: 10.1097 / FBP.0b013e3282d28f2b
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ю, З. Б., Ван, Б., Зицман, Д., Азари, С., и Уайз, Р. А. (2007). Роль условного высвобождения глутамата вентральной тегментальной части при поиске кокаина. J. Neurosci. 27, 10546–10555. DOI: 10.1523 / JNEUROSCI.2967-07.2007
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Зафра, Ф., Castren, E., Thoenen, H., and Lindholm, D. (1991). Взаимодействие между системами передатчиков глутамата и γ-аминомасляной кислоты в физиологической регуляции нейротрофического фактора мозга и синтеза фактора роста нервов в нейронах гиппокампа. Proc. Natl. Акад. Sci. США 88, 10037–10041. DOI: 10.1073 / pnas.88.22.10037
CrossRef Полный текст | Google Scholar
Зарриндаст, М. Р., Ахмади, С., Хаери-Рохани, А., Резайоф, А., Джафари, М. Р., и Джафари-Сабет, М.(2004). Рецепторы GABAA в базолатеральной миндалине участвуют в обеспечении морфинового вознаграждения. Brain Res. 1006, 49–58. DOI: 10.1016 / j.brainres.2003.12.048
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Цвайфель, Л.С., Фадок, Дж. П., Аргилли, Э., Гарелик, М. Г., Джонс, Г. Л., Дикерсон, Т. М. и др. (2011). Активация дофаминовых нейронов имеет решающее значение для аверсивного кондиционирования и предотвращения генерализованной тревоги. Нат. Neurosci. 14, 620–626. DOI: 10.1038 / nn.2808
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Что такое ГАМК?
Перейти к:
Гамма- Аминомасляная кислота или гамма-аминомасляная кислота (ГАМК) — нейромедиатор, обнаруженный в головном мозге. Его активность в мозге была выяснена в 1957 году, когда ученые сообщили, что ГАМК оказывает ингибирующее действие на нейроны раков.
Сейчас он широко известен как главный ингибирующий рецептор в коре головного мозга человека, но в последние годы он стал широко доступной пищевой добавкой как в Европе, так и в Соединенных Штатах.ГАМК считается «пищевой добавкой»; как таковая, она может продаваться без доказательств ее эффективности. ГАМК широко распространена в природе.
ГАМК является тормозным нейромедиатором нервной системы. Кредит изображения: Молекулярное искусство / Shutterstock
Какова роль и распределение ГАМК?
ГАМК распределяется в головном мозге неравномерно. Его ингибирующие эффекты являются результатом подавления нейрональной активности и синаптической пластичности. Хотя основная функция ГАМК — нейротрансмиттер, она имеет структуру аминокислоты и поэтому называется аминокислотным нейромедиатором.ГАМК вырабатывается, когда глутамин в пище сначала превращается в глутамат, а затем в ГАМК декарбоксилазой глутаминовой кислоты. В мозге взрослого человека ГАМК является ингибитором, тогда как в эмбриональном мозге это хорошо установленный возбуждающий передатчик, который выполняет регионально-специфические роли в соответствии со стадией развития эмбриона.
Изучение ГАМК — Часть 1 Играть
ГАМК-рецепторы, агонисты и модуляторы
Почти половина всех синапсов мозга экспрессирует какой-либо рецептор ГАМК.Рецепторы ГАМК широко, но по-разному распределены в ЦНС, и они могут активироваться несколькими изостерами ГАМК, которые представляют собой соединения с одинаковым числом атомов и электронов. Существует 2 типа рецепторов, которые может стимулировать ГАМК: ГАМК и ГАМК. ГАМКА-рецепторы ионотропны; при связывании ГАМК связанный ионный канал, проницаемый для отрицательно заряженных ионов хлорида, открывается для гиперполяризации мембранного потенциала. Это снижает вероятность того, что нейрон запустит потенциал действия.
РецепторыGABAB представляют собой метаботропные рецепторы или рецепторы, связанные с G-белком. Преобладающий эффект стимуляции GABAB — открытие калиевых каналов. Как и в случае с хлоридом, приток калия приводит к гиперполяризации, что снижает вероятность того, что нейрон активирует потенциал действия.
GABAA также имеет сайты аллостерического связывания для других веществ, называемых модуляторами GABA. Они способны увеличивать или уменьшать действие ГАМК; при его отсутствии они не действуют.В зависимости от их эффекта они классифицируются как положительные или отрицательные аллостерические модуляторы (PAM и NAM соответственно).
Ярким примером класса положительных аллостерических модуляторов рецептора ГАМКА являются бензодиазепины. Они облегчают взаимодействие между ГАМК и ГАМК, тем самым усиливая ингибирующий эффект ГАМК. Эти виды препаратов обычно используются в седативных целях.
Другие положительные модуляторы включают барбитураты и алкоголь. Отрицательные модуляторы, напротив, конкурируют со связыванием положительного модулятора в том же месте.Флумазенил является одним из таких примеров конкурентного антагониста; он используется для предотвращения передозировки бензодиазепинов.
Некоторые препараты проявляют нецелевые отрицательные эффекты аллостерических модуляторов. Например, фторхинолоновые антибиотики, антидепрессанты и бета-лактамные антибиотики могут отрицательно модулировать ГАМК. Агонисты ГАМК также производят тот же эффект, что и положительные аллостерические модуляторы; они приводят к снижению нейронных сигналов в мозгу пользователя. Напротив, антагонисты ГАМК увеличивают мозговую активность — например, габазин или бикукуллин.
ГАМК и гематоэнцефалический барьер
Исторически считалось, что ГАМК не может преодолевать гематоэнцефалический барьер (ГЭБ), что ставит под сомнение эффективность добавок ГАМК для облегчения беспокойства или улучшения сна среди других положительных эффектов. Однако доказательства, подтверждающие проницаемость ГАМК, противоречивы. несколько исследований не смогли показать, что ГАМК проникает через ГЭБ, в то время как другие демонстрируют ее способность делать это.
Согласно статье, опубликованной в Frontiers in Psychology (2015), механизм, лежащий в основе добавок ГАМК, неясен, и необходимы дальнейшие исследования, чтобы установить поведенческие эффекты ГАМК.Авторы предполагают, что нет достаточных доказательств, чтобы исключить возможный эффект. Более того, исследования, в которых использовались миметики ГАМК, которые структурно не отражают молекулу ГАМК, и доставляли ГАМК через механизмы, которые не отражают введение пользователем. В частности, проницаемость ГАМК для ГЭБ ограничена грызунами.
Несмотря на противоречивые данные о его проникновении в ЦНС, ГАМК по-прежнему может влиять на мозг через кишечную нервную систему, которая содержит множество рецепторов ГАМК.Таким образом, эффективность добавок ГАМК ограничена неофициальными данными.
Список литературы
- Moss, F.J., Imoukhuede, P.I, Just, H. & Lester, H.A. ТРАНСПОРТЕРЫ | Транспортеры ГАМК: структура, олигомеризация, торговля и фармакология определяют возбудимость нейронов, Энциклопедия фундаментальных исследований эпилепсии. (2009) DOI: 10.1016 / B978-012373961-2.00116-8
- Гетц, Т., Вульф, П. и Висден, В. ГАМКА-рецепторы: молекулярная биология, клеточная биология и фармакология. Энциклопедия неврологии. (2009) DOI: 10.1016 / B978-008045046-9.01236-5
- Sigel, E. et al. Структура, функция и модуляция рецепторов GABAA. JBC. (2012) DOI: 10.1074 / jbc.R112.386664
- Киннерсли, A.M.7 Турано, Ф.Дж. Гамма-аминомасляная кислота (ГАМК) и реакция растений на стресс. Критических обзоров в науках о растениях. (2000) DOI: 10.1080 / 073526800277
- Owens, D.f. И Кригштейн, А.Р. Есть ли в габа что-то большее, чем синаптическое торможение? Природа (2002) doi: 10.1038 / nrn919
Дополнительная литература
Ассоциация гена α2 рецептора гамма-аминомасляной кислоты (GABRA2) с расстройством, связанным с употреблением алкоголя
Addolorato G, Leggio L, Hopf FW, Diana M, Bonci A (2012). Новые терапевтические стратегии для лечения алкогольной и наркотической зависимости: акцент на ГАМК, ионных каналах и транскраниальной магнитной стимуляции. Нейропсихофармакология 37 : 163–177.
CAS Статья Google ученый
Agrawal A, Edenberg HJ, Foroud T., Bierut LJ, Dunne G, Hinrichs AL et al (2006). Связь GABRA2 с наркотической зависимостью в совместном исследовании генетики алкоголизма. Behav Genet 36 : 640–650.
Артикул Google ученый
Американская психиатрическая ассоциация (1994) Диагностическое и статистическое руководство по психическим расстройствам (DSM) Четвертое издание American Psychiatric Press: Вашингтон, округ Колумбия.
Барретт Дж. К., Фрай Б., Маллер Дж., Дейли М. Дж. (2005). Haploview: анализ и визуализация карт LD и гаплотипов. Биоинформатика 21 : 263–265.
CAS Статья Google ученый
Bauer LO, Covault J, Harel O, Das S, Gelernter J, Anton R et al (2007). Вариация в GABRA2 предсказывает алкогольное поведение участников проекта MATCH. Alcohol Clin Exp Res 31 : 1780–1787.
CAS Статья Google ученый
Берут ЖЖ (2007). Генетическая изменчивость, которая способствует никотиновой зависимости. Фармакогеномика 8 : 881–883.
CAS Статья Google ученый
Берут Л. Дж., Стрикленд Дж. Р., Томпсон Дж. Р., Afful SE, Коттлер Л. Б. (2008). Употребление наркотиков и зависимость у лиц с кокаиновой зависимостью, лиц из местных сообществ и их братьев и сестер. Зависимость от наркотиков и алкоголя 95 : 14–22.
Артикул Google ученый
Boyle AP, Hong EL, Hariharan M, Cheng Y, Schaub MA, Kasowski M et al (2012). Аннотация функциональных вариаций в личных геномах с помощью RegulomeDB. Genome Res 22 : 1790–1797.
CAS Статья Google ученый
Калифано JA (2007).Высшее общество: как злоупотребление психоактивными веществами разрушает Америку и что с этим делать. PublicAffairs: Нью-Йорк, Нью-Йорк, США.
Чанг Ю.Т., Сан Х.С., Фанн С.С., Чанг С.Дж., Ляо Чж., Хуанг Дж.Л. и др. (2002). Связь кластера генов рецептора гамма-аминомасляной кислоты А с алкогольной зависимостью у тайваньской хань. Mol Psychiatry 7 : 828–829.
CAS Статья Google ученый
Cossart R, Bernard C, Ben-Ari Y (2005).Множественные аспекты ГАМКергических нейронов и синапсов: множественные судьбы передачи сигналов ГАМК при эпилепсии. Trends Neurosci 28 : 108–115.
CAS Статья Google ученый
Cossette P, Liu L, Brisebois K, Dong H, Lortie A, Vanasse M et al (2002). Мутация GABRA1 при аутосомно-доминантной форме юношеской миоклонической эпилепсии. Нат Генет 31 : 184–189.
CAS Статья Google ученый
Ково Дж., Гелернтер Дж., Хессельброк В., Неллиссери М., Кранцлер Х.Р. (2004).Аллельная и гаплотипическая связь GABRA2 с алкогольной зависимостью. Am J Med Genet B Neuropsychiatr Genet 129B : 104–109.
Артикул Google ученый
Ково Дж., Гелернтер Дж., Дженсен К., Антон Р., Кранцлер Х.Р. (2008). Маркеры в 5′-области GABRG1 связаны с алкогольной зависимостью и находятся в неравновесном сцеплении с маркерами в соседнем гене GABRA2. Нейропсихофармакология 33 : 837–848.
CAS Статья Google ученый
Дэвис М. (2003). Роль рецепторов GABAA в опосредовании воздействия алкоголя на центральную нервную систему. J Psychiatry Neurosci 28 : 263–274.
PubMed PubMed Central Google ученый
Дэвис К.М., Ву Дж.Й. (2001). Роль глутаматергической и ГАМКергической систем при алкоголизме. J Biomed Sci 8 : 7–19.
CAS Статья Google ученый
DerSimonian R, Laird N (1986). Мета-анализ в клинических исследованиях. Контрольные клинические испытания 7 : 177–188.
CAS Статья Google ученый
Dick DM, Edenberg HJ, Xuei X, Goate A, Hesselbrock V, Schuckit M et al (2005). В совместном исследовании генетики алкоголизма не обнаружено ассоциации генов рецепторов GABAA на хромосоме 5 с алкоголизмом. Am J Med Genet B Neuropsychiatr Genet 132B : 24–28.
Артикул Google ученый
Дик Д.М., Планкетт Дж., Уэтерилл Л.Ф., Сюэй Х, Гоут А., Хессельброк В. и др. (2006). Связь между GABRA1 и алкогольным поведением в совместном исследовании генетики алкоголизма. Alcohol Clin Exp Res 30 : 1101–1110.
CAS Статья Google ученый
Duval S, Tweedie R (2000).Обрезать и заполнить: простой метод тестирования и корректировки с учетом предвзятости публикации в метаанализе, основанный на воронкообразных диаграммах. Биометрия 56 : 455–463.
CAS Статья Google ученый
Эденберг Х.Дж. (2002). Совместное исследование генетики алкоголизма: обновление. Alcohol Res Health 26 : 214–218.
PubMed Google ученый
Edenberg HJ, Dick DM, Xuei X, Tian H, Almasy L, Bauer LO et al (2004).Вариации GABRA2, кодирующей альфа-2-субъединицу рецептора GABA (A), связаны с алкогольной зависимостью и колебаниями мозга. Am J Hum Genet 74 : 705–714.
CAS Статья Google ученый
Эггер М., Дэйви Смит Г., Шнайдер М., Миндер С. (1997). Предвзятость в метаанализе обнаруживается с помощью простого графического теста. BMJ 315 : 629–634.
CAS Статья Google ученый
Енох М.А., Ходжкинсон, Калифорния, Юань К., Олбо Б., Вирккунен М., Гольдман Д. (2009).GABRG1 и GABRA2 как независимые предикторы алкоголизма в двух популяциях. Нейропсихофармакология 34 : 1245–1254.
CAS Статья Google ученый
Фер С., Сандер Т., Тадич А., Лензен К.П., Ангелеску И., Клаве С. и др. (2006). Подтверждение ассоциации гена GABRA2 с алкогольной зависимостью анализом подтипа. Psychiatr Genet 16 : 9–17.
Артикул Google ученый
Фоли П.Ф., Ло Э. У., Иннес Д. Д., Уильямс С. М., Танненберг А. Э., Харпер К. Г. и др. (2004). Ассоциативные исследования полиморфизма генов нейротрансмиттеров у алкоголиков европеоидной расы. Ann N Y Acad Sci 1025 : 39–46.
CAS Статья Google ученый
Франк Дж., Сишон С., Трейтлейн Дж., Райдингер М., Маттейзен М., Хоффманн П. и др. (2012).Общегеномная значимая связь между алкогольной зависимостью и вариантом в кластере генов ADH. Биол наркоманов 17 : 171–180.
CAS Статья Google ученый
Fu Q, Heath AC, Bucholz KK, Nelson E, Goldberg J, Lyons MJ et al (2002). Общий генетический риск большой депрессии, алкогольной зависимости и зависимости от марихуаны: вклад антисоциального расстройства личности у мужчин. Arch Gen Psychiatry 59 : 1125–1132.
Артикул Google ученый
Гелернтер Дж., Кранцлер Х.Р. (2009). Генетика алкогольной зависимости. Hum Genet 126 : 91–99.
CAS PubMed PubMed Central Google ученый
Gelernter J, Panhuysen C, Weiss R, Brady K, Hesselbrock V, Rounsaville B et al (2005).Сканирование геномных связей для выявления кокаиновой зависимости и связанных с ней признаков: существенные связи кокаиновой зависимости и паранойи, вызванной кокаином. Am J Med Genet B Neuropsychiatr Genet 136B : 45–52.
Артикул Google ученый
Гелернтер Дж., Шерва Р., Кестерер Р., Алмаси Л., Чжао Х., Кранцлер Х. и др. (2013). Полногеномное исследование зависимости от кокаина и связанных с ней признаков: FAM53B идентифицирован как ген риска. Mol Psychiatry (предварительная онлайн-публикация 20 августа 2013 г .; doi: 10.1038 / mp.2013.99.
Goldman D, Oroszi G, Ducci F (2005). Генетика зависимостей: раскрытие генов. Nat Rev Genet 6 : 521–532.
CAS PubMed PubMed Central Google ученый
Хан Д.Х., Боло Н., Дэниэлс М.А., Лю И.К., Мин К.Дж., Ким СН и др. (2008). Тяга к алкоголю и пище во время лечения алкогольной зависимости: модуляция аллелем Т 1519T> C GABAAalpha6. Alcohol Clin Exp Res 32 : 1593–1599.
CAS Статья Google ученый
Heath AC, Whitfield JB, Martin NG, Pergadia ML, Goate AM, Lind PA et al (2011). Исследование ассоциации риска алкоголизма в обществе с количественными признаками генома: результаты и последствия. Биологическая психиатрия 70 : 513–518.
Артикул Google ученый
Hsu YP, Seow SV, Loh EW, Wang YC, Chen CC, Yu JM и др. (1998).Поиск мутаций рядом с альтернативно сплайсированным 8-аминокислотным экзоном в гене субъединицы гамма 2 рецептора ГАМКА и отсутствие аллельной ассоциации с алкоголизмом среди четырех групп аборигенов и китайцев хань на Тайване. Brain Res. Mol. Brain Res. 56 : 284–286.
CAS Статья Google ученый
Иттивут К., Листман Дж., Мутирангура А., Малисон Р., Ково Дж., Кранцлер Х. Р. и др. (2008). Неравновесные паттерны межпопуляционного сцепления генов GABRA2 и GABRG1 в кластерном локусе GABA на хромосоме 4 человека. Геномика 91 : 61–69.
CAS Статья Google ученый
Иттивут Ч., Ян Б.З., Кранцлер Х.Р., Антон РФ, Хирунсатит Р., Вайс РД и др. (2012). Вариации GABRG1 и GABRA2, связанные с алкогольной зависимостью у афроамериканцев. Alcohol Clin Exp Res 36 : 588–593.
CAS Статья Google ученый
Кан Дж.К., Шен В., Макдональд Р.Л. (2009).Мутация GABRG2, Q351X, связанная с генерализованной эпилепсией с фебрильными припадками плюс, имеет как потерю функции, так и доминантно-отрицательное подавление. J Neurosci 29 : 2845–2856.
CAS Статья Google ученый
Кендлер К.С., Майерс Дж., Прескотт, Калифорния (2007). Специфика генетических и экологических факторов риска появления симптомов зависимости от каннабиса, кокаина, алкоголя, кофеина и никотина. Arch Gen Psychiatry 64 : 1313–1320.
Артикул Google ученый
Хан А.Дж., Рували М., Чоудхури Г., Матур Н., Хусейн К., Пармар Д. (2009). Полиморфизм цитохрома P450 2E1 и взаимодействие с другими генетическими факторами риска и предрасположенность к алкогольному циррозу печени. Mutat Res 664 : 55–63.
CAS Статья Google ученый
Лаппалайнен Дж., Крупицкий Э., Ремизов М., Пчелина С., Тараскина А., Звартау Э. и др. (2005).Связь алкоголизма с подтипом альфа2 рецепторов гамма-аминомасляной кислоты в российской популяции. Alcohol Clin Exp Res 29 : 493–498.
CAS Статья Google ученый
Li D, Collier DA, He L (2006). Мета-анализ показывает сильную положительную связь гена нейрегулина 1 (NRG1) с шизофренией. Hum Mol Genet 15 : 1995–2002.
CAS Статья Google ученый
Ли Д., Чжао Х., Гелернтер Дж. (2011).Сильная связь гена алкогольдегидрогеназы 1B (ADh2B) с алкогольной зависимостью и заболеваниями, вызванными алкоголем. Биологическая психиатрия 70 : 504–512.
CAS Статья Google ученый
Ли Т, Лю Х, Чжао Дж, Ху Х, Болл Д.М., Ло эль В. и др. (2002). Анализ аллельной ассоциации дофаминовых рецепторов D2, D3, 5-HT2A и GABA (A) gamma2 и генов переносчиков серотонина со злоупотреблением героином у китайских субъектов. Am J Med Genet 114 : 329–335.
Артикул Google ученый
Линь С.К., Чен С.К., Болл Д., Лю Х.С., Ло Э.В. (2003). Гендерно-специфический вклад генов субъединицы ГАМК (A) в 5q33 при расстройстве, связанном с употреблением метамфетамина. Pharmacogenomics J 3 : 349–355.
CAS Статья Google ученый
Линд П.А., Макгрегор С., Агравал А., Монтгомери Г.В., Хит А.С., Мартин Н.Г. и др. (2008).Роль GABRA2 в алкогольной зависимости, курении и употреблении запрещенных наркотиков в выборке населения Австралии. Alcohol Clin Exp Res 32 : 1721–1731.
CAS Статья Google ученый
Ло Э. У., Хигучи С., Мацусита С., Мюррей Р., Чен К. К., Болл Д. (2000). Анализ ассоциации кластера генов субъединицы рецептора ГАМК (A) на 5q33-34 и алкогольной зависимости в популяции Японии. Mol Psychiatry 5 : 301–307.
CAS Статья Google ученый
Ло Э. У., Смит И., Мюррей Р., Маклафлин М., МакНалти С., Болл Д. (1999). Связь между вариантами в кластере генов GABAAbeta2, GABAAalpha6 и GABAAgamma2 и алкогольная зависимость в шотландской популяции. Mol Psychiatry 4 : 539–544.
CAS Статья Google ученый
Ло Э. У., Тан Н. Л., Ли Д. Т., Лю С. И., Стадлин А. (2007).Анализ ассоциации генов субъединиц рецептора ГАМК на 5q33 с героиновой зависимостью у китайского мужского населения. Am J Med Genet B Neuropsychiatr Genet 144B : 439–443.
CAS Статья Google ученый
Люшер К., Маленка RC (2011). Синаптическая пластичность, вызванная лекарствами, при зависимости: от молекулярных изменений до ремоделирования цепей. Нейрон 69 : 650–663.
Артикул Google ученый
Lydall GJ, Saini J, Ruparelia K, Montagnese S, McQuillin A, Guerrini I et al (2011).Изучение генетической ассоциации однонуклеотидных полиморфизмов GABRA2 и электроэнцефалография при алкогольной зависимости. Neurosci Lett 500 : 162–166.
CAS Статья Google ученый
Маклин П.Дж., Фарб Д.Х., Рассек С.Дж. (1995). Картирование гена субъединицы альфа 4 (GABRA4) на хромосоме 4 человека определяет кластер генов альфа 2-альфа 4-бета 1-гамма 1: еще одно доказательство того, что современные кластеры генов рецепторов ГАМК происходят из наследственного кластера. Геномика 26 : 580–586.
CAS Статья Google ученый
Нестлер Э.Дж. (2005). Есть ли общий молекулярный путь к зависимости? Nat Neurosci 8 : 1445–1449.
CAS Статья Google ученый
Нишияма Т., Икеда М., Ивата Н., Сузуки Т., Китадзима Т., Яманучи Ю. и др. (2005).Связь гаплотипов между геном субъединицы гамма2 рецептора ГАМКА (GABRG2) и расстройством, связанным с употреблением метамфетамина. Pharmacogenomics J 5 : 89–95.
CAS Статья Google ученый
Олфсон Э., Берут Л.Дж. (2012). Конвергенция полногеномных ассоциаций и исследований генов-кандидатов на алкоголизм. Alcohol Clin Exp Res 36 : 2086–2094.
CAS Статья Google ученый
Онори Н., Турчи С., Солито Дж., Джесуита Р., Бушеми Л., Тальабраччи А. (2010).GABRA2 и расстройства, связанные с употреблением алкоголя: нет доказательств связи в итальянском исследовании случай-контроль. Alcohol Clin Exp Res 34 : 659–668.
CAS Статья Google ученый
Park CS, Park SY, Lee CS, Sohn JW, Hahn GH, Kim BJ (2006). Связь между алкоголизмом и генетическим полиморфизмом генов рецепторов ГАМК на хромосоме 5q33-34 в корейской популяции. J Korean Med Sci 21 : 533–538.
CAS Статья Google ученый
Пиеруччи-Лага А., Гелернтер Дж., Чан Дж., Ариас А., Кубеллс Дж. Ф., Фаррер Л. и др. (2007). Надежность диагностических критериев DSM-IV с использованием полуструктурированной оценки наркотической зависимости и алкоголизма (SSADDA). Зависимость от наркотиков и алкоголя 91 : 85–90.
Артикул Google ученый
Причард Дж. К., Стивенс М., Розенберг Н. А., Доннелли П. (2000).Картирование ассоциаций в структурированных популяциях. Am J Hum Genet 67 : 170–181.
CAS Статья Google ученый
Радел М., Вальехо Р.Л., Ивата Н., Арагон Р., Лонг Дж. К., Вирккунен М. и др. (2005). Локализация гена алкогольной зависимости на основе гаплотипа в кластере генов 5q34 {гамма} -аминомасляной кислоты типа А. Arch Gen Psychiatry 62 : 47–55.
CAS Статья Google ученый
Райх Т., Эденберг Х. Дж., Гоут А., Уильямс Дж. Т., Райс Дж. П., Ван Эрдевег П. и др. (1998).Полногеномный поиск генов, влияющих на риск алкогольной зависимости. Am J Med Genet 81 : 207–215.
CAS Статья Google ученый
Сандер Т., Болл Д., Мюррей Р., Патель Дж., Самоховец Дж., Винтерер Г. и др. (1999). Анализ ассоциации вариантов последовательностей кластера генов GABA (A) alpha6, beta2 и gamma2 и алкогольной зависимости. Alcohol Clin Exp Res 23 : 427–431.
CAS Статья Google ученый
Soyka M, Preuss UW, Hesselbrock V, Zill P, Koller G, Bondy B (2008). Полиморфизм гена субъединицы рецептора GABA-A2 (GABRA2) и риск алкогольной зависимости. J Psychiatr Res 42 : 184–191.
CAS Статья Google ученый
Сюняев С., Раменский В., Кох И., Токарный W 3-й, Кондрашов А.С., Борк П. (2001).Прогнозирование вредоносных аллелей человека. Hum Mol Genet 10 : 591–597.
CAS Статья Google ученый
True WR, Xian H, Scherrer JF, Madden PA, Bucholz KK, Heath AC и др. (1999). Общая генетическая уязвимость мужчин к никотиновой и алкогольной зависимости. Arch Gen Psychiatry 56 : 655–661.
CAS Статья Google ученый
Ван Дж.К., Фоуд Т., Хинрихс А.Л., Ле NX, Бертельсен С., Бадд JP и др. (2013).Полногеномное ассоциативное исследование количества симптомов алкогольной зависимости в расширенных родословных идентифицирует C15orf53. Mol Psychiatry 18 : 1218–1224.
CAS Статья Google ученый
Вайнер Дж. Л., Валенсуэла, CF (2006). Этаноловая модуляция ГАМКергической передачи: вид из среза. Pharmacol Ther 111 : 533–554.
CAS Статья Google ученый
Вульф Б. (1955).Об оценке связи между группой крови и заболеванием. Энн Хам Генет 19 : 251–253.
CAS Статья Google ученый
Xian H, Scherrer JF, Grant JD, Eisen SA, True WR, Jacob T et al (2008). Вклад генетики и окружающей среды в зависимость от никотина, алкоголя и каннабиса у близнецов мужского пола. Зависимость 103 : 1391–1398.
Артикул Google ученый
Сюй З., Тейлор Дж. А. (2009).SNPinfo: интеграция информации о GWAS и генах-кандидатах в функциональный отбор SNP для исследований генетических ассоциаций. Nucleic Acids Res 37 (выпуск веб-сервера): W600 – W605.
CAS Статья Google ученый
Зинцарас Э. (2012). Варианты рецептора гамма-аминомасляной кислоты А, альфа-2 (GABRA2) как отдельные маркеры алкоголизма: метаанализ. Psychiatr Genet 22 : 189–196.
CAS Статья Google ученый
Что такое добавки ГАМК? | DreamLife Recovery
Гамма-аминомасляная кислота (ГАМК) — это аминокислота и природное химическое вещество в вашем мозгу, а также нейротрансмиттер или химический посредник. Однако ГАМК считается тормозящим нейротрансмиттером, потому что он блокирует определенные сигналы мозга, тем самым снижая активность нервной системы.
ГАМК оказывает успокаивающее действие, когда присоединяется к рецептору ГАМК в головном мозге.Благодаря успокаивающему эффекту в последние годы стали популярны добавки с ГАМК. В то время как аминокислоты часто можно найти в продуктах питания, ГАМК содержится только в определенных продуктах, включая чай, помидоры, соевые бобы и ферментированные продукты, такие как кимчи, мисо и темпе.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) одобрило ГАМК для нас в качестве пищевых добавок. Его можно найти в конфетах, жевательной резинке, спортивных напитках и закусочных. Добавки бывают в форме таблеток, капсул или иногда порошка, которые можно смешивать с коктейлями и напитками.
Были проведены исследования, чтобы понять эффективность, но только с натуральной ГАМК, которая присутствует в пище, или биосинтетической ГАМК, производимой путем ферментации. Также еще не существует рекомендуемой суточной дозы ГАМК, поэтому перед началом приема любых добавок ГАМК следует проконсультироваться с врачом.
Для чего используются добавки ГАМК?Стресс — распространенный фактор в повседневной жизни, но он неуклонно растет из года в год. По данным Американского института стресса, примерно 33 процента людей сообщают о сильном стрессе.Кроме того, 77 процентов людей говорят, что стресс влияет на их физическое здоровье, 73 процента говорят, что стресс влияет на их психическое здоровье, а 48 процентов говорят, что они испытывают трудности со сном из-за стресса. Добавки
ГАМК часто назначают при стрессе, который может вызвать снижение концентрации и расстановки приоритетов. ДобавкиGABA могут помочь уменьшить стресс, беспокойство и бессонницу благодаря своему успокаивающему эффекту. У людей с такими заболеваниями, как тревожность, паническое расстройство, депрессия, синдром дефицита внимания и гиперактивности, судорожное расстройство и болезнь Паркинсона, могут быть более низкие уровни ГАМК.
Эффективность добавок ГАМК до сих пор неясна, однако в настоящее время проводятся исследования некоторых видов использования ГАМК. Популярные способы применения включают беспокойство, высокое кровяное давление, бессонницу, стресс и усталость.
Исследование аминомасляных кислот, проведенное в 2012 году с участием 63 участников, показало, что прием 100 миллиграммов (мг) ГАМК значительно снижает стресс.
В 2015 году было проведено еще одно исследование аминомасляной кислоты на группе из 30 человек. Исследование показало, что прием 800 мг ГАМК улучшает мышление и способности к выполнению задач, например определение приоритетов.
Возможные побочные эффекты ГАМКНеобходимо провести дополнительные исследования, чтобы точно сказать, каковы потенциальные побочные эффекты добавок ГАМК.
Некоторые люди сообщают о следующих побочных эффектах:
- Сонливость
- Головная боль
- Мышечная слабость
- Расстройство желудка
Добавки влияют на всех по-разному, поэтому, если вы и ваш поставщик медицинских услуг решите, что добавки ГАМК могут принести вам пользу , важно обращать внимание на любые побочные эффекты, которые вы испытываете.Ваш врач также сможет помочь определить, взаимодействуют ли добавки ГАМК с любыми другими добавками или лекарствами, которые вы, возможно, принимаете.
Какими лекарствами по рецепту чаще всего злоупотребляют?
Узнать большеПодходят ли вам добавки ГАМК?
Перед тем, как начать прием добавок ГАМК, вам нужно проконсультироваться с врачом. Вместе вы можете определить, принесет ли вам пользу добавление ГАМК, а затем составить план действий.
Для получения дополнительной информации о добавках и других лекарствах, потенциально связанных с зависимостью, свяжитесь с приемной комиссией DreamLife Recovery сегодня.
Ресурсы:
- Steenbergen, L., Sellaro, R., Stock, AK. et al. Введение γ-аминомасляной кислоты (ГАМК) улучшает процессы выбора действий: рандомизированное контролируемое исследование. Sci Rep 5, 12770 (2015)
- Yoto, A., Murao, S., Motoki, M. et al. Пероральный прием γ-аминомасляной кислоты влияет на настроение и деятельность центральной нервной системы во время стрессового состояния, вызванного умственными задачами. Аминокислоты 43, 1331–1337 (2012)
- «Нейротрансмиттеры в качестве пищевых добавок: влияние ГАМК на мозг и поведение» — Эверт Бунстра, Рой де Клейн, Лоренца С. Кользато, Аннеке Алькемад, Бирте У. Forstmann, Sander Nieuwenhuis Front Psychol . 2015; 6: 1520. Опубликовано онлайн, 2015 6 октября
- Hepsomali P, Groeger JA, Nishihira J and Scholey A (2020) Влияние перорального приема гамма-аминомасляной кислоты (ГАМК) на стресс и сон у людей: систематический обзор.