ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА — это… Что такое ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА?
- ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА
- ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА
Г А М К, HOOCh3Ch3Ch3Nh3, аминокислота, один из медиаторов нервной системы. Фермент, катализирующий образование Г.-а. к. путём декарбоксилирования L-глутаминовой к-ты (глутаматдекарбоксилаза), содержится в нервной ткани, где локализован в особых нейронах — источнике секреции медиаторной Г.-а. к. Активный синтез Г.-а. к. идёт и в нек-рых др. клетках, напр. в бета-клетках поджелудочной железы (и в развивающейся из них опухоли, инсулиноме), где функция Г.-а. к. неизвестна. У ракообразных и насекомых Г.-а. к. служит медиатором тормозных двигат. нейронов, а в мозге позвоночных — медиатором мн. тормозных интернейронов: поэтому её часто наз. медиатором синаптич. торможения. Однако было показано, что Г.-а.к., подобно др. медиаторам, оказывает как тормозящие, так и возбуждающие эффекты.

.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)

.
- ГАМИЯ
- ГАММАГЛОБУЛИНЫ
Смотреть что такое «ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА» в других словарях:
Гамма-аминомасляная кислота — Гамма аминомасляная кислота … Википедия
Гамма-аминомасляная кислота — Nh3Ch3Ch3 Ch3COOH, образуется путём декарбоксилирования (См. Декарбоксилирование) (под действием фермента декарбоксилазы) глутаминовой кислоты (См. Глутаминовая кислота). Обмен Г. а. к. в организме приводит к образованию янтарной кислоты … Большая советская энциклопедия
гамма-аминомасляная кислота — (ГАМК) естественный метаболит ткани мозга, представляющий собой масляную кислоту с аминогруппой в положении; предполагается участие ГАМК в процессе передачи нервных импульсов; препараты ГАМК используются в качестве сосудорасширяющих… … Большой медицинский словарь
Гамма-аминомасляная кислота (лекарственное средство) — Основная статья: Гамма аминомасляная кислота … Википедия
Гамма-аминомасляная кислота (ГАМК) — аминокислота, выполняющая функции тормозного нейромедиатора в сером веществе мозга, в клетках базального ганглия, которые проецируются на чёрную субстанцию, в клетках Пуркинье мозжечка и в дорсальном роге спинного мозга.
ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА (ГАМК) — Аминокислота, которая выполняет функции важного тормозного нейромедиатора во многих местах центральной нервной системы. Она находится в сером веществе, в клетках базального ганглия, которые проецируются на черную субстанцию, в клетках Пуркинье… … Толковый словарь по психологии
Гамма-аминомасляная кислота (ГАМК) — (gamma aminobiotic acid (GABA)) нейротрансмиттер, с низкой активностью которого связывают генерализованное тревожное расстройство … Общая психология: глоссарий
Альфа-аминомасляная кислота — Альфа аминомасляная кислота … Википедия
Глутаминовая кислота — Глутаминовая кислота … Википедия
Вальпроевая кислота — Химическое соединение … Википедия

Активное вещество ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА* (ACIDUM GAMMA-AMINOBUTYRICUM*)
USPDDN: γ-аминобутировая кислота.
MESH, RTECS:4-аминобутировая кислота.
RTECS: 4-аминобутаноевая кислота; γ-амино-n-бутировая кислота.
NLM: γ-аминобутаноевая кислота.
Mm = 103,12 Да. Белый кристаллический порошок со слабогорьким вкусом и специфическим запахом. Легкорастворим в воде, очень малорастворим — в спирте. Точка плавления — 203 °С. log P (октанол-вода) = –3,17.
Форма выпуска: таблетки п/о, капсулы твердые.
Лекарственные препараты содержащие активное вещество ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА*
психостимуляторы, средства применяемые при синдроме нарушения внимания и гиперактивности (adhd), и ноотропные средства
психостимуляторы, средства применяемые при синдроме нарушения внимания и гиперактивности (adhd), и ноотропные средства
ГАМК является основным медиатором, участвующим в процессах торможения в ЦНС. Под ее влиянием улучшаются энергетические процессы в головном мозгу, а также утилизация последним глюкозы, повышается потребление тканями кислорода, нормализуется кровоснабжение. ГАМК улучшает динамику нервных процессов в головном мозгу, мышление, память, внимание, способствует восстановлению движений и речи после нарушения мозгового кровообращения, оказывает мягкое психостимулирующее действие. По характеру действия приближается к ноотропным средствам.
ГАМК улучшает динамику нервных процессов в головном мозгу, мышление, память, внимание, способствует восстановлению движений и речи после нарушения мозгового кровообращения, оказывает мягкое психостимулирующее действие. По характеру действия приближается к ноотропным средствам.
заболевания сосудов головного мозга, хроническая недостаточность мозгового кровообращения, состояния после инсульта и травм головного мозга для восстановления памяти, внимания, речи, повышения двигательной и психической активности больных, устранения головокружения и головной боли, динамические нарушения мозгового кровообращения. ГАМК также применяют при алкогольных полиневритах, слабоумии, для лечения симптомокомплекса укачивания.
внутрь до еды, взрослым — в зависимости от характера и тяжести заболевания по 0,5–1,25 г 3 раза в сутки. Суточная доза обычно составляет 1,5–3 г. Курс лечения — от 2–3 нед до 2–6 мес. При необходимости проводят повторные курсы лечения. Для терапии симптомокомплекса укачивания назначают по 0,5 г 3 раза в сутки в течение 3 дней (для профилактики укачивания — по 0,5 г 3 раза в сутки в течение 3 дней, до возможного укачивания).
не установлены.
в отдельных случаях возможны диспептические явления, нарушения сна, ощущение жара, колебания АД в первые дни приема. При снижении дозы указанные явления проходят.
Препараты и лекарства с действующим веществом Гамма-аминомасляная кислота
{{/if}} {{each list}} ${this} {{if isGorzdrav}}Удалить
{{/if}} {{/each}} {{/if}} Поражение сосудов головного мозга (атеросклероз, гипертоническая болезнь и др.), цереброваскулярная недостаточность и дисциркуляторная энцефалопатия, нарушение памяти, внимания, речи, головокружение, головная боль, последствия инсульта и черепно-мозговой травмы, алкогольная энцефалопатия, алкогольный полиневрит, умственная отсталость у детей, слабоумие, детский церебральный паралич, эндогенная депрессия с преобладанием астеноипохондрических явлений и затруднением умственной деятельности, симптомокомплекс укачивания (морская и воздушная болезнь).
Фармакологическое действие
ноотропное, стимулирующее метаболизм в ЦНСЯвляется основным медиатором, участвующим в процессах центрального торможения. Улучшает кровоснабжение головного мозга, активирует энергетические процессы, повышает дыхательную активность тканей, улучшает утилизацию глюкозы и удаление токсических продуктов обмена. Взаимодействует со специфическими ГАМКергическими рецепторами А и Б типов. Улучшает динамику нервных процессов в головном мозге, повышает продуктивность мышления, улучшает память, оказывает умеренное психостимулирующее, антигипоксическое и противосудорожное действие.Способствует восстановлению речевых и двигательных функций после нарушения мозгового кровообращения. Оказывает умеренное гипотензивное действие, уменьшает исходно повышенное АД и выраженность обусловленных гипертонией симптомов (головокружение, бессонница), незначительно урежает ЧСС. У больных сахарным диабетом снижает уровень глюкозы в крови, при нормальной гликемии нередко вызывает гипергликемию, обусловленную гликогенолизом.
Противопоказания
Гиперчувствительность, детский возраст (до 1 года), острая почечная недостаточность, беременность (I триместр).
Применение при беременности и кормлении грудью
Противопоказан в I триместре беременности.
Гамма-аминомасляная кислота (ГАМК)
ГАММА-АМИНОМАСЛЯНАЯ КИСЛОТА (ГАМК, GABA)γ-Аминомасляная кислота
(сокр. ГАМК, GABA) — органическое соединение, непротеиногенная аминокислота, важнейший тормозной нейромедиатор центральной нервной системы (ЦНС) человека и других млекопитающих.
 Аминомасляная кислота является биогенным веществом. Содержится в ЦНС и принимает участие в нейромедиаторных и метаболических процессах в мозге. Гамма-аминомасляная кислота в организме образуется из другой аминокислоты — глутаминовой с помощью фермента глутаматдекарбоксилазы.
Аминомасляная кислота является биогенным веществом. Содержится в ЦНС и принимает участие в нейромедиаторных и метаболических процессах в мозге. Гамма-аминомасляная кислота в организме образуется из другой аминокислоты — глутаминовой с помощью фермента глутаматдекарбоксилазы.Гамма-аминомасляная кислота (ГАМК) является главным тормозящим нейротрансмиттером. Он мягок, уравновешен и не очень физически скоординирован. Его основная работа заключается в регулировании возбуждающих сигналов, посылаемых другими нейротрансмиттерами. Он позволяет мышцам и кровеносным сосудам расслабиться,а телу-нормально спать. Без его присутствия тело было бы под угрозой смерти от судорог !
Отношение к психоактивным молекулам: глутамат, «старшая сестра» ГАМК, является главным возбуждающим нейротрансмиттером. Большинство лекарств, которые мешают работе ГАМК, являются седативными, включая алкоголь, гамма-гидроксимасляная кислота (GHB), барбитураты и бензодиазепины.
ГАМК. Нейромедиатор — монополист «отрасли» торможения в нервной системе. Находится в состоянии вечной борьбы за влияние со своим бодрым отцом Глутаматом. Основная функция — гашение возбуждающих сигналов: ГАМК убеждает нейроны (и нас, их «хозяев») не реагировать на провокации агрессивных соседей и соблюдать спокойствие, чтобы не пасть жертвами глутаматных «козней» (например, инсульта). Вероятно, ГАМК участвует в поддержании нормального цикла сна и повышает усвоение глюкозы. Не исключено, что дирижирует она и какими-то сигнальными путями у растений — не зря же это основная аминокислота апопласта помидоров!
Итак, Гамма-аминомасляная кислота (ГАМК) — главный тормозной медиатор в нервной системе человека. Но только тех из нас, у кого она уже развита. А чтобы обеспечить нам поистине олимпийское спокойствие, ей иногда помогает пёстрая компания очень известных веществ. Мы познакомимся с ГАМК поближе и узнаем, что эта молекула не так проста, как кажется на первый взгляд.
Нейромедиатор покоя
Гамма-аминомасляная кислота (ГАМК; γ-aminobutyric acid, GABA) синтезируется в мозге из глутаминовой кислоты — еще одного нейромедитора — путем ее декарбоксилирования (удаления карбоксильной группы из основной цепи) (рис. 1). По химической классификации ГАМК — это аминокислота, но не привычная, то есть используемая для синтеза белковых молекул, α-аминокислота, где аминогруппа присоединена к первому атому углерода в цепочке. В ГАМК аминогруппа связана с третьим от карбоксильной группы атомом (в глутамате он был первым по счету до декарбоксилирования).
Рисунок 1. Синтез ГАМК. При помощи фермента глутаматдекарбоксилазы (GAD) из нейромедиатора глутамата получается другой нейромедиатор — ГАМК.
ГАМК синтезируется прямо в мозге и связывается с двумя типами рецепторов на поверхности нейронов — ГАМК-рецепторами типов А и В. Рецепторы типа А раньше подразделялись на рецепторы типов А и С (встречаются преимущественно в сетчатке глаза), но в последующем были объединены в связи с общностью действия. Этот тип рецепторов является ионотропным: при связывании с ними ГАМК в мембране нервной клетки открывается ионный канал, и ионы хлора устремляются в клетку, снижая ее реактивность. Мембрана нервной клетки обладает потенциалом покоя. Внутри клетки меньше заряженных ионов, чем снаружи, и это создает разницу зарядов. Снаружи превосходство создается хлором, кальцием и натрием, а внутри преобладают ионы калия и ряд отрицательно заряженных органических молекул. В теоретическом смысле у потенциала мембраны есть два пути: увеличение (называемое деполяризацией) и уменьшение (гиперполяризация) (рис. 2). В покое мембранный потенциал равен приблизительно −70…−90 мВ (милливольт), а при работе нервной системы начинается «перетягивание каната» между двумя силами — возбуждающими клетку (деполяризующими мембрану) и тормозящими ее (гиперполяризующими).
Этот тип рецепторов является ионотропным: при связывании с ними ГАМК в мембране нервной клетки открывается ионный канал, и ионы хлора устремляются в клетку, снижая ее реактивность. Мембрана нервной клетки обладает потенциалом покоя. Внутри клетки меньше заряженных ионов, чем снаружи, и это создает разницу зарядов. Снаружи превосходство создается хлором, кальцием и натрием, а внутри преобладают ионы калия и ряд отрицательно заряженных органических молекул. В теоретическом смысле у потенциала мембраны есть два пути: увеличение (называемое деполяризацией) и уменьшение (гиперполяризация) (рис. 2). В покое мембранный потенциал равен приблизительно −70…−90 мВ (милливольт), а при работе нервной системы начинается «перетягивание каната» между двумя силами — возбуждающими клетку (деполяризующими мембрану) и тормозящими ее (гиперполяризующими).
Рисунок 2. Схема возникновения потенциала действия на мембране клетки. Необходимоизменение содержания ионов внутри и снаружи клетки такой силы, чтобы значение заряда на мембране изменилось и достигло определенного порога. Если это происходит, то мембрана продолжает деполяризоваться дальше, нейрон возбуждается и передает сигнал другим клеткам. Овершут (инверсия) — период, когда потенциал мембраны положителен. Затем следует фаза реполяризации, и заряд мембраны возвращается к прежним значениям.
Необходимоизменение содержания ионов внутри и снаружи клетки такой силы, чтобы значение заряда на мембране изменилось и достигло определенного порога. Если это происходит, то мембрана продолжает деполяризоваться дальше, нейрон возбуждается и передает сигнал другим клеткам. Овершут (инверсия) — период, когда потенциал мембраны положителен. Затем следует фаза реполяризации, и заряд мембраны возвращается к прежним значениям.
Чтобы понять, как это работает, надо учесть два момента. Первый — на один нейрон в то же самое время могут воздействовать несколько противоположно направленных сил: например, пять возбуждающих и три тормозящих нейрона сошлись на одной клетке в этом участке нервной системы. При этом они могут воздействовать на дендрит этого нейрона и на аксон в пресинаптической части. Второй момент — нервная клетка, испытывающая эти воздействия, будет работать по принципу «всё или ничего». Она не может одновременно послать сигнал и не посылать его. Все воздействия сигналов, пришедших на клетку, суммируются, и если итоговые изменения потенциала мембраны превысят определенное значение (называемое порогом возбуждения), то сигнал будет передан на другую клетку через синапс.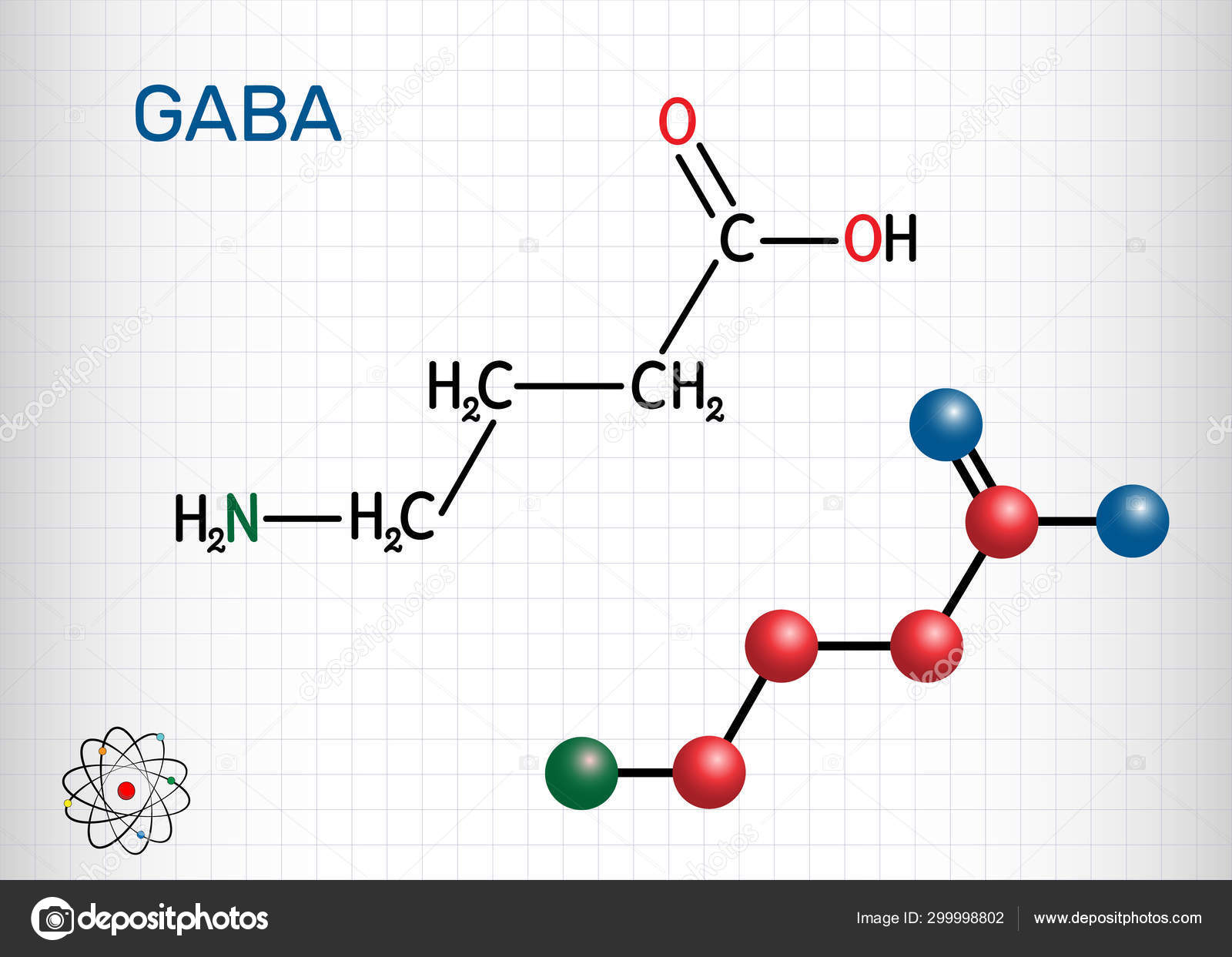 Если же пороговое значение не будет достигнуто, то извините — попробуйте еще раз, ребята. Всё это напоминает басню Крылова про лебедя, рака и щуку: каждый тянет в свою сторону, но не очень понятно, что из этого выйдет.
Если же пороговое значение не будет достигнуто, то извините — попробуйте еще раз, ребята. Всё это напоминает басню Крылова про лебедя, рака и щуку: каждый тянет в свою сторону, но не очень понятно, что из этого выйдет.
Итак, молекула ГАМК связалась с рецептором ионного канала. Ионный канал, обладающий довольно сложным строением (рис. 3), раскрывается и начинает пропускать внутрь клетки отрицательно заряженные ионы хлора. Под воздействием этих ионов происходит гиперполяризация мембраны, и клетка становится менее восприимчивой к возбуждающим сигналам других нейронов. Это первая и, пожалуй, главная функция ГАМК — торможение активности нервных клеток в нервной системе.
Рисунок 3. Ионотропный ГАМК-рецептор. Рецептор ГАМКА — гетеропентамер: состоит из 5 белковых субъединиц, которые в зависимости от гомологии аминокислотных последовательностей могут принадлежать к восьми разным семействам (чаще — к α, β, γ; члены ρ-семейства гомоолигомеризуются — получаются рецепторы ГАМКA-ρ, «бывшие» ГАМКC). Это определяет разнообразие ГАМКА-рецепторов. Схема строения рецептора. Слева: Каждая из субъединиц на длинном глобулярном N-конце, выходящем на поверхность нейрона, имеет характерную структуру «цистеиновая петля» и участки связывания ГАМК и других лигандов. Далее следуют 4 α-спиральных трансмембранных домена (между последними из них — большая цитоплазматическая петля, ответственная за связывание с цитоскелетом и «внутренними» модуляторами) и короткий C-конец. Справа: Пять субъединиц образуют ионный канал, ориентируясь вторым трансмембранным доменом (оранжевым цилиндром) друг к другу. Это четвертичная структура рецептора. При связывании с двумя молекулами ГАМК рецептор меняет конформацию, открывая пору для транспорта анионов.
Это определяет разнообразие ГАМКА-рецепторов. Схема строения рецептора. Слева: Каждая из субъединиц на длинном глобулярном N-конце, выходящем на поверхность нейрона, имеет характерную структуру «цистеиновая петля» и участки связывания ГАМК и других лигандов. Далее следуют 4 α-спиральных трансмембранных домена (между последними из них — большая цитоплазматическая петля, ответственная за связывание с цитоскелетом и «внутренними» модуляторами) и короткий C-конец. Справа: Пять субъединиц образуют ионный канал, ориентируясь вторым трансмембранным доменом (оранжевым цилиндром) друг к другу. Это четвертичная структура рецептора. При связывании с двумя молекулами ГАМК рецептор меняет конформацию, открывая пору для транспорта анионов.
Рецепторы типа В являются метаботропными, то есть влияют на обмен веществ в клетке.Они тоже снижают уровень возбуждения в клетке, но делают это более медленными способами, через систему G-белков. Рецепторы этого типа помогают клетке снизить чувствительность к возбуждающим воздействиям через влияние на кальциевые и калиевые каналы.
Рецепторы этого типа помогают клетке снизить чувствительность к возбуждающим воздействиям через влияние на кальциевые и калиевые каналы.
Припадки и тревога
ГАМК-ергическая система головного мозга по своему строению напоминает все остальные (рис. 4). Есть ряд глубоко расположенных в мозге структур, откуда нервные волокна, выделяющие ГАМК, идут в другие части нервной системы. Поэтому ГАМК является тормозным нейромедиатором, регулирующим многие процессы — от мышечного тонуса до эмоциональных реакций.
Рисунок 4. ГАМК-ергические пути головного мозга человека. Скопления нервных клеток в глубине мозга рассылают свои отростки в разные отделы нервной системы, чтобы снижать излишний уровень возбуждения.
Однако тормозным медиатором ГАМК становится только в зрелом мозге. В развивающейся нервной системе ГАМК-ергические нейроны могут производить возбуждающее действие на клетки, также меняя проницаемость мембраны для ионов хлора [1]. В незрелых нервных клетках концентрация ионов хлора выше, чем в окружающей среде, и стимуляция рецепторов ГАМК приводит к выходу этих анионов из клетки и последующей деполяризации мембраны. Со временем созревает основная возбуждающая система мозга — глутаматная, — и ГАМК приобретает роль тормозного(гиперполяризующего мембрану) нейромедиатора.
Со временем созревает основная возбуждающая система мозга — глутаматная, — и ГАМК приобретает роль тормозного(гиперполяризующего мембрану) нейромедиатора.
Само созревание мозга — это сложный процесс, который на разных этапах онтогенеза регулируется множеством генов (рис. 5). Нарушение процессов созревания и миграции нейронов приводит к различным неврологическим заболеваниям, например, эпилепсии [2]. Эпилепсия — одно из самых распространенных неврологических заболеваний. При нём нейроны головного мозга генерируют нервные импульсы не так, как следуют — слишком часто и слишком сильно, что приводит к возникновению патологического очага возбуждения в мозге. Именно существование такого очага приводит к припадкам — самому главному и опасному симптому эпилепсии. Такая«разрядка» позволяет на время снизить возбуждение в нервной системе. Мутации в ряде генов приводят к тому, что ГАМК-ергические вставочные нейроны оказываются не на своем месте и не могут полноценно выполнять свои тормозящие функции.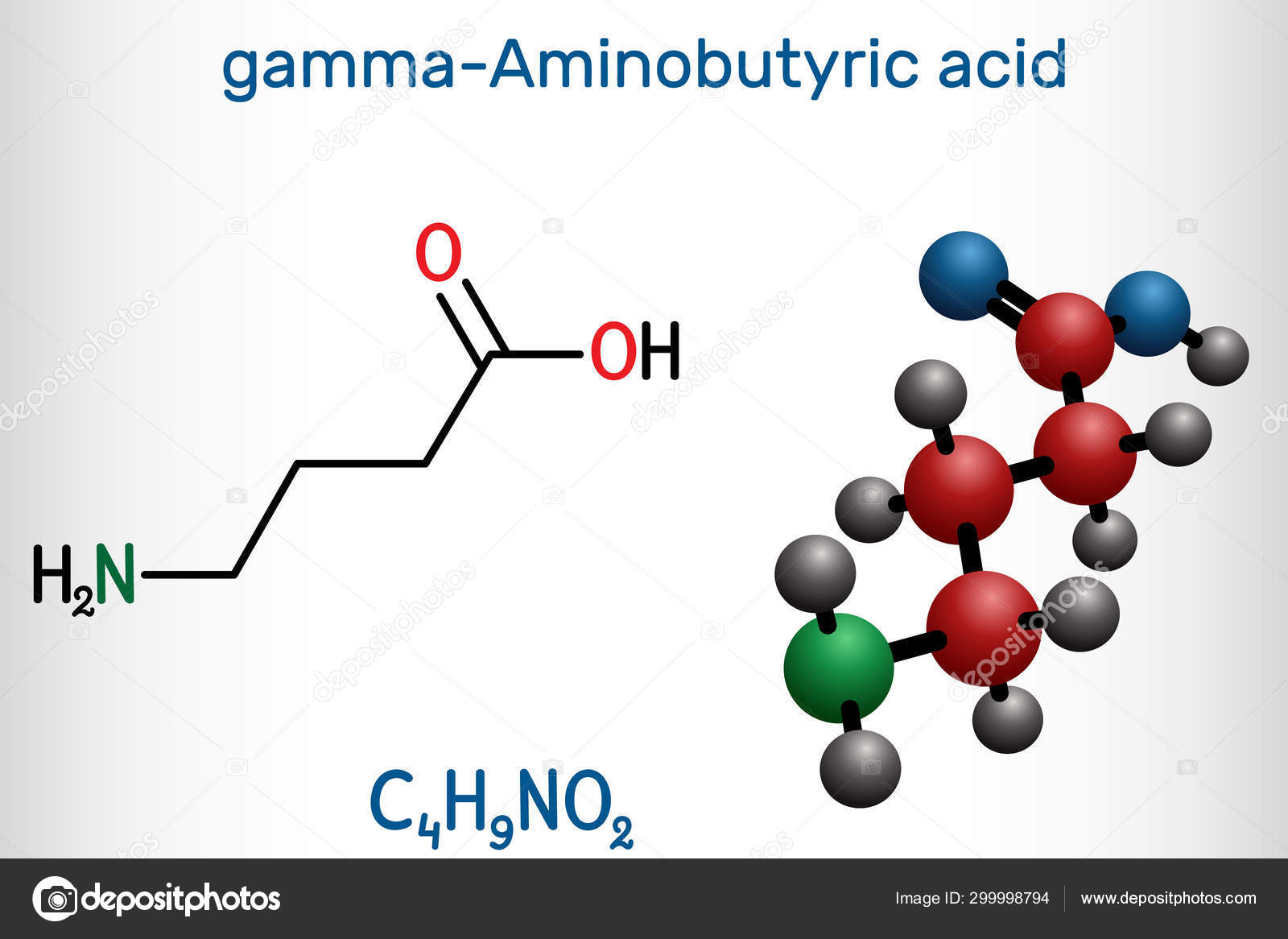 На мышиных моделях и при исследовании генотипа людей была установлена связь между мутациями, нарушением миграции и созревания ГАМК-ергических нейронов и развитием эпилепсии.
На мышиных моделях и при исследовании генотипа людей была установлена связь между мутациями, нарушением миграции и созревания ГАМК-ергических нейронов и развитием эпилепсии.
Рисунок 5. Гены, отвечающие за созревание мозга, включаются в работу на разных этапах онтогенеза. Эмбриональный и постнатальный периоды разделены точкой P0 (рождение). За рост, созревание и функцию тормозящих клеток отвечают гены DLX, ARX, DCX, RELN. Семейство генов DLX (distal-less homeobox) кодирует гомеодомен-содержащие транскрипционные факторы. Большинство экспрессируется при формировании органов чувств и миграции клеток гребня и вставочных нейронов; регулируют экспрессию гена ARX. ARX (aristaless-related homeobox) кодирует гомеодомен-содержащий транскрипционный фактор, контролирующий дифференцировку клеток различных органов. В развивающемся мозге он необходим для миграции вставочных нейронов. DCX (doublecortin) кодирует даблкортин (lissencephalin-X) — ассоциированный с микротрубочками белок, синтезируемый в незрелых нейронах при их делении (маркер нейрогенеза, в том числе у взрослых). Он необходим для правильной миграции и дифференцировки нейробластов, поскольку влияет на динамику микротрубочек цитоскелета (стабилизирует их и группирует). RELN (reelin) — ген секретируемого сигнального гликопротеина рилина. При развитии нервной системы волокна радиальной глии ориентируются в направлении большей концентрации рилина, выстраивая «пути» для миграции нейронов. Необходим этот белок и для правильного построения слоев коры. Активен RELN и в других тканях, даже у взрослых. В развитом мозге рилин секретируется ГАМК-ергическими вставочными нейронами гиппокампа и коры. Вероятно, он стимулирует удлинение нейронных отростков, влияет на синаптическую пластичность и память.
Он необходим для правильной миграции и дифференцировки нейробластов, поскольку влияет на динамику микротрубочек цитоскелета (стабилизирует их и группирует). RELN (reelin) — ген секретируемого сигнального гликопротеина рилина. При развитии нервной системы волокна радиальной глии ориентируются в направлении большей концентрации рилина, выстраивая «пути» для миграции нейронов. Необходим этот белок и для правильного построения слоев коры. Активен RELN и в других тканях, даже у взрослых. В развитом мозге рилин секретируется ГАМК-ергическими вставочными нейронами гиппокампа и коры. Вероятно, он стимулирует удлинение нейронных отростков, влияет на синаптическую пластичность и память.
Другим аспектом тормозящего действия ГАМК является влияние на эмоциональные процессы — в частности на тревогу. Тревога — это очень обширное понятие. В нём заключены как и совершенно здоровые реакции человека на стрессовые воздействия(экзамен, темная подворотня, признание в любви), так и патологические состояния (тревожные расстройства в медицинском смысле этого слова). Исходя из положений современной психиатрической науки, можно сказать, что есть нормальная тревога и тревога как болезнь. Тревога становится болезнью, когда она мешает вашей повседневной или профессиональной жизни, блокируя принятие любых решений — даже самых необходимых.
Исходя из положений современной психиатрической науки, можно сказать, что есть нормальная тревога и тревога как болезнь. Тревога становится болезнью, когда она мешает вашей повседневной или профессиональной жизни, блокируя принятие любых решений — даже самых необходимых.
Отделом мозга, который отвечает за эмоциональные реакции, является миндалевидное тело — скопление нервных клеток в глубине нашей головы. Это одна из самых древних и важных частей нервной системы у животных. Особой специальностью миндалевидного тела являются отрицательные эмоции — мы гневаемся, злимся, боимся и тревожимся через миндалину. ГАМК позволяет мозгу снижать интенсивность этих переживаний.
Таблетка от нервов
Лекарства, которые эффективны в борьбе с тревогой и припадками, должны связываться с рецептором ГАМК. Они не являются прямыми стимуляторами рецептора, т.е. не связываются с той же частью молекулы, что и ГАМК. Их роль заключается в том, что они повышают чувствительность ионного канала к ГАМК, немного меняя его пространственную организацию. Такие химические вещества называются аллостерическими модуляторами. К аллостерическим модуляторам ГАМК-рецепторов относятся этанол, бензодиазепины и барбитураты.
Такие химические вещества называются аллостерическими модуляторами. К аллостерическим модуляторам ГАМК-рецепторов относятся этанол, бензодиазепины и барбитураты.
Алкоголь известен своим расслабляющим и противотревожным эффектом. Растворы этилового спирта в различных концентрациях с давних пор широко используются населением Земли для успокоения нервов. Этанол дарит людям расслабление, связываясь с рецептором ГАМК и упрощая его дальнейшее взаимодействие с медиатором. Бывает такое, что люди переоценивают свои возможности в употреблении спиртного, и это приводит к постепенной потере контроля над своими действиями и нарастанием заторможенности. Наступает алкогольное гиперраслабление, которое при продолжении употребления может дойти до алкогольной комы — настолько сильным оказывается угнетающее действие спирта на центральную нервную систему. Потенциально алкоголь мог бы использоваться во время хирургических операций как наркозное средство (раньше в критических ситуациях — например, на фронте — так и поступали — Ред. ), но спектр концентраций, где он выключает болевую чувствительность и еще не«выключает» человека полностью, слишком мал.
), но спектр концентраций, где он выключает болевую чувствительность и еще не«выключает» человека полностью, слишком мал.
Другой класс веществ — барбитураты — сейчас используется в неврологии для лечения эпилептических судорог. Все лекарства этого класса — аллостерические модуляторы, производные барбитуровой кислоты — барбитала (рис. 6). Сам барбитал продавался известной фирмой Bayer под торговым названием «Веронал». В дальнейшем были синтезированы другие производные барбитуровой кислоты: фенобарбитал («Люминал») и бензобарбитал. Эти препараты, появившиеся в начале ХХ века, стали первым эффективным и относительно безопасным лекарством для борьбы с эпилепсией. Производные барбитуровой кислоты использовались и для борьбы с нарушениями сна, но в меньших дозах.
Рисунок 6. Молекула барбитуровой кислоты.
Еще одной группой лекарств, усиливающих действие ГАМК на клетки, являются бензодиазепины. Как и предыдущие вещества, бензодиазепины связываются с рецептором ГАМК типа А (рис. 7). На одной из субъединиц ионного канала есть специальное место, куда присоединяется бензодиазепин. Все препараты этого класса обладают седативным (успокоительным), противотревожным и противосудорожным действием. Сейчас психиатры и неврологи считают плохим тоном лечить тревогу и бессонницу у пациентов длительными курсами бензодиазепинов, а уж тем более назначать их постоянный прием. К этим препаратам довольно быстро вырабатывается зависимость, и отмена приводит к стойким нарушениям сна и возобновлению тревоги. По этим причинам рекомендуется назначать бензодиазепины короткими курсами — на несколько дней.
7). На одной из субъединиц ионного канала есть специальное место, куда присоединяется бензодиазепин. Все препараты этого класса обладают седативным (успокоительным), противотревожным и противосудорожным действием. Сейчас психиатры и неврологи считают плохим тоном лечить тревогу и бессонницу у пациентов длительными курсами бензодиазепинов, а уж тем более назначать их постоянный прием. К этим препаратам довольно быстро вырабатывается зависимость, и отмена приводит к стойким нарушениям сна и возобновлению тревоги. По этим причинам рекомендуется назначать бензодиазепины короткими курсами — на несколько дней.
Рисунок 7. Схематическое представление сайтов связывания (с лекарственными препаратами) на наиболее распространенной изоформе ГАМКА-рецептора. Примечание: α1, β2 и γ2 представляют собой субъединицы наиболее широко распространенной изоформы ГАМКА-рецептора в центральной нервной системе. Сокращения: Cl— pore, хлоридная пора; BDZ, бензодиазепин; ETF, этифоксин; NS, нейростероид; GABA, гамма-аминомасляная кислота (ГАМК).
Наиболее распространенная в ЦНС комбинация субъединиц (около 40 % ГАМКА-рецепторов) — двух α1, двух β2 и одной γ2, располагающихся вокруг хлоридной поры. GABA site (на поверхности, стык α и β) — место, где ГАМК присоединяется к рецептору; BDZ site (на поверхности, стык α и γ) — сайт связывания бензодиазепинов, ETF site (на β) — этифоксина, NS site (в канале) — нейростероидов. Сайты связывания барбитуратов и этанола предположительно находятся в глубине канала (на трансмембранных доменах). В первом случае, вероятно, главную роль играет β-субъединица, с этанолом же взаимодействуют разные субъединицы, включая ρ и δ, но их чувствительность различается.
Причина нелюбви к бензодиазепинам кроется в их побочных эффектах, которых довольно много, и не все они учитываются официальными структурами [4]. Во-первых, бензодиазепины, как и все ГАМК-ергические препараты, вызывают стойкую зависимость. Во-вторых, бензодиазепины ухудшают память человека. Применение препаратов этой группы усиливает тормозящее влияние ГАМК на клетки гиппокампа — центра памяти. Это может приводить к затруднениям в запоминании новой информации, что и наблюдается на фоне приема бензодиазепинов, особенно у пожилых людей.
Это может приводить к затруднениям в запоминании новой информации, что и наблюдается на фоне приема бензодиазепинов, особенно у пожилых людей.
Для лечения тревоги врачи в настоящее время используют антидепрессанты и другие препараты, например, этифоксин [3]. Об этой и других группах препаратов, применяемых в комплексном лечении уже не тревожности, а депрессии следует искать информацию в истории антидепрессантов. Дадим лишь краткую информацию по антидепрессантам в контексте ГАМК и серотонина, главного фигуранта аннотаций антидепрессантных препаратов. Так, на серотониновых рецепторах «висит» огромное количество функций. Через них реализует свой эффект огромное количество лекарств и наркотиков. И все это еще как-то можно было бы игнорировать, если бы не тот факт, что серотонин вообще не особенно-то и участвует в формировании настроения. Основной возбуждающий нейромедиатор в головном мозге человека — это аминокислота глутамат. Основной тормозящий — γ-аминомасляная кислота (ГАМК), которая получается из того же глутамата. Серотонин, дофамин, норадреналин и прочие гормоны выполняют вспомогательную модулирующую функцию.
Серотонин, дофамин, норадреналин и прочие гормоны выполняют вспомогательную модулирующую функцию.
К середине 2000-х годов стали проясняться некоторые механизмы формирования эмоций. В то же время, из анализа биологических теорий развития депрессий, единого взгляда на проблему до сих пор нет (Табл.1).
Таблица 1. Существующие биологические теории патофизиологии депрессии. | ||
Теория | Аргументы «за» | Аргументы «против» |
Нарушение глутаматной передачи | Уровень глутамата и глутамина в префронтальной коре понижен Внутривенное введение кетамина (антагониста NMDA-рецепторов) вызывает антидепрессантный эффект | Уровень глутамата в затылочной коре увеличен |
Снижение ГАМК-передачи | Уровень ГАМК в плазме, спинномозговой жидкости, префронтальной и затылочной коре снижен | ГАМК работает в >30% синапсов в мозгу, что подразумевает неспецифичность действия |
Нарушение циркадных ритмов | Депривация сна и световая терапия оказывают антидепрессантный эффект | Связь между «clock-генами» и депрессией не выявляется |
Нарушение функций эндогенных опиоидов | Агонисты δ-опиоидного рецепторы оказывают на приматов антидепрессантное действие и повышают уровень нейротрофина в мозгу | Отсутствуют крупномасштабные исследования, подтверждающие такую связь |
а также: дисбаланс моноамины / ацетилхолин, цитокиновый обмен между иммунной и нервной системами, нарушение функций тироксина, нарушение работы некоторых «контуров» мозга и др. | ||
Таким образом, ГАМК, несмотря на свою узкую «специальность», — удивительный нейромедиатор. В развивающемся мозге γ-аминомасляная кислота возбуждает нервные клетки, а в развившемся, наоборот, снижает их активность. Она отвечает за чувство спокойствия, а препараты, активирующие ее рецепторы, приносят врачам массу поводов для тревоги. Такой предстала перед нами гамма-аминомасляная кислота — простая молекула, отвечающая за то, чтобы наши мозги не «перегорели».
К разделу: Гамма-аминомасляная кислота (ГАМК) и кишечный микробиом
Литература
- Y. Ben-Ari, J.-L. Gaiarsa, R. Tyzio, R. Khazipov. (2007). GABA: A Pioneer Transmitter That Excites Immature Neurons and Generates Primitive Oscillations. Physiological Reviews. 87, 1215-1284;
- Bozzi Y., Casarosa S., Caleo M. (2012). Epilepsy as a neurodevelopmental disorder. Front. Psychiatry. 3, 19;
- Nuss Ph. (2015). Anxiety disorders and GABA neurotransmission: a disturbance of modulation. Neuropsychiatr. Dis. Treat. 11, 165–175;
- Lader M. (2011). Benzodiazepines revisited—will we ever learn? Addiction. 106, 2086–2109;
 (2015). Anxiety disorders and GABA neurotransmission: a disturbance of modulation. Neuropsychiatr. Dis. Treat. 11, 165–175;
(2015). Anxiety disorders and GABA neurotransmission: a disturbance of modulation. Neuropsychiatr. Dis. Treat. 11, 165–175;Будьте здоровы!
ССЫЛКИ К РАЗДЕЛУ О ПРЕПАРАТАХ ПРОБИОТИКАХ
- ПРОБИОТИКИ
- ДОМАШНИЕ ЗАКВАСКИ
- БИФИКАРДИО
- КОНЦЕНТРАТ БИФИДОБАКТЕРИЙ ЖИДКИЙ
- ПРОПИОНИКС
- ЙОДПРОПИОНИКС
- СЕЛЕНПРОПИОНИКС
- МИКРОЭЛЕМЕНТНЫЙ СОСТАВ
- ПРОБИОТИКИ С ПНЖК
- БИФИДОБАКТЕРИИ
- ПРОПИОНОВОКИСЛЫЕ БАКТЕРИИ
- ПРОБИОТИКИ И ПРЕБИОТИКИ
- СИНБИОТИКИ
- РОЛЬ МИКРОБИОМА В ТЕРАПИИ РАКА
- АНТИОКСИДАНТНЫЕ СВОЙСТВА
- АНТИОКСИДАНТНЫЕ ФЕРМЕНТЫ
- АНТИМУТАГЕННАЯ АКТИВНОСТЬ
- МИКРОФЛОРА КИШЕЧНОГО ТРАКТА
- МИКРОБИОМ ЧЕЛОВЕКА
- МИКРОФЛОРА И ФУНКЦИИ МОЗГА
- ПРОБИОТИКИ И ХОЛЕСТЕРИН
- ПРОБИОТИКИ ПРОТИВ ОЖИРЕНИЯ
- МИКРОФЛОРА И САХАРНЫЙ ДИАБЕТ
- ПРОБИОТИКИ и ИММУНИТЕТ
- МИКРОБИОМ И АУТОИММУННЫЕ БОЛЕЗНИ
- ПРОБИОТИКИ и ГРУДНЫЕ ДЕТИ
- ПРОБИОТИКИ, БЕРЕМЕННОСТЬ, РОДЫ
- ДИСБАКТЕРИОЗ
- ВИТАМИННЫЙ СИНТЕЗ
- АМИНОКИСЛОТНЫЙ СИНТЕЗ
- АНТИМИКРОБНЫЕ СВОЙСТВА
- СИНТЕЗ ЛЕТУЧИХ ЖИРНЫХ КИСЛОТ
- СИНТЕЗ БАКТЕРИОЦИНОВ
- ФУНКЦИОНАЛЬНОЕ ПИТАНИЕ
- АЛИМЕНТАРНЫЕ ЗАБОЛЕВАНИЯ
- ПРОБИОТИКИ ДЛЯ СПОРТСМЕНОВ
- ПРОИЗВОДСТВО ПРОБИОТИКОВ
- ЗАКВАСКИ ДЛЯ ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ
- НОВОСТИ
GABA (ГАМК) что это, как работает и в чем польза
Благодаря многочисленным исследованиям сегодня ГАМК официально признана эффективным цитопротекторным антигипоксантом и антиоксидантом для разных органов и систем.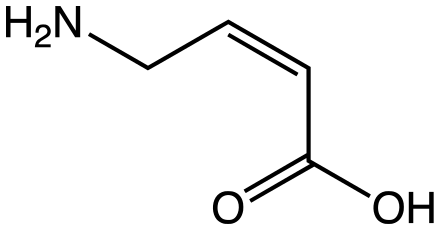 Ей приписывают противоопухолевые и противовоспалительные свойства, а также способность положительно влиять на работу иммунной системы.
Ей приписывают противоопухолевые и противовоспалительные свойства, а также способность положительно влиять на работу иммунной системы.
Особенно заметно влияние ГАМК на сон. Под ее влиянием снижается психическое возбуждение, активизируются энергетические процессы мозга, улучшается дыхательная активность тканей и утилизация глюкозы, что приводит к качественному глубокому сну.
Действие GABA на организм:
- способствует успокоению и расслаблению
- контролирует «панические» состояния, связанные с тревогой
- снижает мышечный тонус
- уменьшает судороги при эпилепсии
- устраняет мышечные спазмы
- снижает частоту сердечных сокращений
- нормализует кровяное давление
- помогает контролировать беспокойство
- содействует качественному сну
- уменьшает содержания сахара в крови
- повышает общий иммунитет
- ускоряет процесс восстановления после перенесенных травм
- улучшает сексуальную активность
С помощью препаратов GABA лечат умственную отсталость, ДЦП, постинсультные состояния, а также восстанавливают функции мозга после травм. Прием добавок с ГАМК целесообразен при высоких умственных, физических и психологических нагрузках. Они помогают нормализировать работу мозга, справится со стрессом.
Прием добавок с ГАМК целесообразен при высоких умственных, физических и психологических нагрузках. Они помогают нормализировать работу мозга, справится со стрессом.
ГАБА для спортсменов
Сегодня многие профессиональные спортсмены признают пользу гамма-аминомасляной кислоты для спортивных достижений. Преимуществом спортпита на ее основе является высокая эффективность при отсутствии гормональной составляющей. Воздействуя на переднюю долю гипофиза головного мозга, ГАМК стимулирует выработку гормона роста, благодаря чему усиливается рост мышечной ткани и снижается процент жира во всем организме. Это является особенно важным для бодибилдеров и атлетов.
Преимущества ГАБА для спортсменов:
➦ повышает содержание гормона роста в 4-6 раз, являясь безопасной альтернативой стероидам
➦ стимулирует выработку соматотропина даже после тренировки
➦ участвует в процессе жиросжигания, способствуя более быстрой сушке
➦ помогает быстрее добиться красивого рельефного тела
➦ нормализирует работу нервной системы
➦ устраняет бессонницу, улучшает сон
➦ ускоряет восстановление после травм: вывихов и растяжений.
Добавки с GABA стабильно пользуются высоким спросом у профессиональных атлетов и любителей, что является лучшим подтверждением их высокой эффективности. Единичные негативные отзывы связаны с неправильным применением препаратов с ГАБа или нарушением тренировочного процесса.
Важно помнить, что все диетические и спортивные добавки принимают в определенной дозировке, курсами, после консультации с врачом.
Гамма-аминомасляная кислота (ГАМК), Аминалон [LifeBio.wiki]
γ-аминомасляная кислота (ГАМК) является главным тормозным нейромедиатором в центральной нервной системе млекопитающих. Он играет роль в регуляции возбудимости нейронов по всей нервной системе. В организме человека ГАМК также непосредственно отвечает за регулирование мышечного тонуса.
Хотя с химической точки зрения вещество является аминокислотой, в научных или медицинских статьях ГАМК редко упоминается в таком качестве, поскольку термин «аминокислота», используемый без уточнения, относится к альфа-аминокислотам, каковой ГАМК не является. ГАМК также не включена в состав белков.
При спастической диплегии у людей, поглощение ГАМК нарушается в результате повреждения нервов при поражении верхнего двигательного нейрона, что приводит к гипертонии мышц.
ГАМК также не включена в состав белков.
При спастической диплегии у людей, поглощение ГАМК нарушается в результате повреждения нервов при поражении верхнего двигательного нейрона, что приводит к гипертонии мышц.
ИЮПАК название: 4-аминобутановая кислота
Молекулярная формула: C4H9NO2
Молярная масса: 103,120 г / моль
Внешний вид: белый микрокристаллический порошок
Плотность: 1,11 г / мл
Температура плавления: 203,7 ° C (398,7 ° F ; 476,8 К)
Температура кипения: 247,9 ° C (478,2 ° F ; 521,0 К)
Растворимость в воде: 130 г/100 мл
Кислотность (рКа): 4,23 (карбоксил), 10,43 (амино)
Краткий обзор
ГАМК является наиболее активным тормозным нейроамином человеческого головного мозга. Она регулирует действие множества тормозных и седативных процессов, происходящих в ткани головного мозга, и поэтому чрезвычайно важна для релаксации.
Концентрации ГАМК постоянно контролируются организмом, в результате чего количество ГАМК в тканях человеческого тела является сбалансированным. Благодаря этим регуляционным факторам, пищевая добавка ГАМК не способна оказать чрезмерно подавляющее действие на организм. Человеческое тело слишком привычно к регуляции ГАМК, и поэтому её пероральный приём не может оказать значительного воздействия на человеческую физиологию.
Тем не менее, другие соединения способны (различными путями) косвенно увеличить уровень ГАМК в организме, что, в свою очередь оказывает тормозное действие.
ГАМК также известна как гамма-аминомасляная кислота.
Благодаря этим регуляционным факторам, пищевая добавка ГАМК не способна оказать чрезмерно подавляющее действие на организм. Человеческое тело слишком привычно к регуляции ГАМК, и поэтому её пероральный приём не может оказать значительного воздействия на человеческую физиологию.
Тем не менее, другие соединения способны (различными путями) косвенно увеличить уровень ГАМК в организме, что, в свою очередь оказывает тормозное действие.
ГАМК также известна как гамма-аминомасляная кислота.
ГАМК является тормозным нейромедиатором, но пищевая добавка ГАМК выраженного тормозящего действия не оказывает.
Часто принимается в паре с препаратами, увеличивающими содержание окиси азота.
Внимание! ГАМК является одним из главных нейротрансмиттеров головного мозга. Важно помнить, что совместный ее прием с нейроактивными препаратами или антидепрессантами может спровоцировать отрицательные побочные эффекты.
ГАМК инструкция по применению
Чаще всего добавка ГАМК применяется в дозах 3000-5000 мг (для повышения метаболизма гормона роста). Является ли это оптимальной дозировкой, точно не известно.
Является ли это оптимальной дозировкой, точно не известно.
Краткий обзор
ГАМК (гамма-аминомасляная кислота) является одним из наиболее выраженных нейроактивных пептидов головного мозга. Она задействована во множестве подавляющих и тормозных процессов, связанных с парасимпатической нервной системой. ГАМК образуется из возбуждающего нейромедиатора глутамата с помощью фермента глутаматдекарбокилазы и может быть преобразована обратно в глутамат в цикле трикарбоновых кислот.1)
Концентрация ГАМК
Установлено, что изменения концентрации ГАМК головного мозга и концентрации общей ГАМК находятся в прямой зависимости друг от друга. Изменение содержания ГАМК в головном мозге обязательно приводит к изменению концентрации общей ГАМК и наоборот.
Накопление ГАМК в мозге ускоряется, когда содержание ГАМК падает ниже физиологического уровня, и замедляется, когда содержание ГАМК превышает физиологический уровень. Такое поведение кислоты обусловлено тем, что она является ингибитором собственной транспортировки в мозг, и прекращает своё накопление при концентрациях выше нормы. Благодаря этому механизму, неврологический уровень ГАМК остаётся сбалансированным.2) И всё же, ГАМК не может снизить своё накопление до нуля. Установлено, что наивысший уровень собственного ингибирования ГАМК составляет 80%. Из чего следует, что чрезмерный прием ГАМК может пересилить её собственное ингибирование путём пассивной диффузии.
Когда уровень ГАМК в головном мозге превышает физиологический, мозг начинает вытеснять избыток кислоты. Скорость вытеснения ГАМК через гемаэнцефалический барьер приблизительно в 16 раз превышает скорость её накопления. Удаление избытка ГАМК из нервных тканей активизируется как защитная реакция организма от завышенного тормозного воздействия.
Благодаря этому механизму, неврологический уровень ГАМК остаётся сбалансированным.2) И всё же, ГАМК не может снизить своё накопление до нуля. Установлено, что наивысший уровень собственного ингибирования ГАМК составляет 80%. Из чего следует, что чрезмерный прием ГАМК может пересилить её собственное ингибирование путём пассивной диффузии.
Когда уровень ГАМК в головном мозге превышает физиологический, мозг начинает вытеснять избыток кислоты. Скорость вытеснения ГАМК через гемаэнцефалический барьер приблизительно в 16 раз превышает скорость её накопления. Удаление избытка ГАМК из нервных тканей активизируется как защитная реакция организма от завышенного тормозного воздействия.
ГАМК и гемаэнцефалический барьер
У взрослых наблюдается минимальное проникновение ГАМК из системного круга кровообращения в ткани головного мозга. Также отмечено, что гемаэнцефалический барьер молодых людей обладает наивысшей пропускающей способностью. При избытке ГАМК в организме, ГАМК ингибирует собственное поступление через гемаэнцефалический барьер, в чем прослеживается ее сходство с бета-аланином, хотя в этом механизме ГАМК проявляет себя более ярко. Установлено, что окись азота может увеличить пропускающую способность гемаэнцефалического барьера.
Установлено, что окись азота может увеличить пропускающую способность гемаэнцефалического барьера.
ГАМК и гормон роста
Долгое время полагалось, что приём ГАМК усиливает секрецию гормона роста, и в этом есть доля правды, только «гормон роста» в этом случае включает лишь определённый подкласс аналогов. Имуннореактивный гормон роста (irGH) и имуннофункциональный гормон роста (ifGH) – два аналога, уровень которых увеличивается после принятия добавки ГАМК.3) Несмотря на то, что ГАМК неэффективно проникает через гемаэнцефалический барьер, она оказывает вышеупомянутое действие неврологически, а точнее, через выработку дофамина в гипофизе.
Интересное изменение в действии ГАМК на секрецию ГР наблюдается при упражнениях с отягощением, а именно увеличение площади под кривой и более высокие пиковые значения. Действие ГАМК достигает максимума через 30 минут упражнений после принятия ГАМК и через 75 минут при отсутствии физической нагрузки (в состоянии покоя).
Несмотря на то, что на данный момент прямое воздействие ГАМК на гормон роста не доказано (так же как и биотрансформация ГАМК в другие амины в печени), многие учёные считают, что вероятность этой взаимосвязи высока.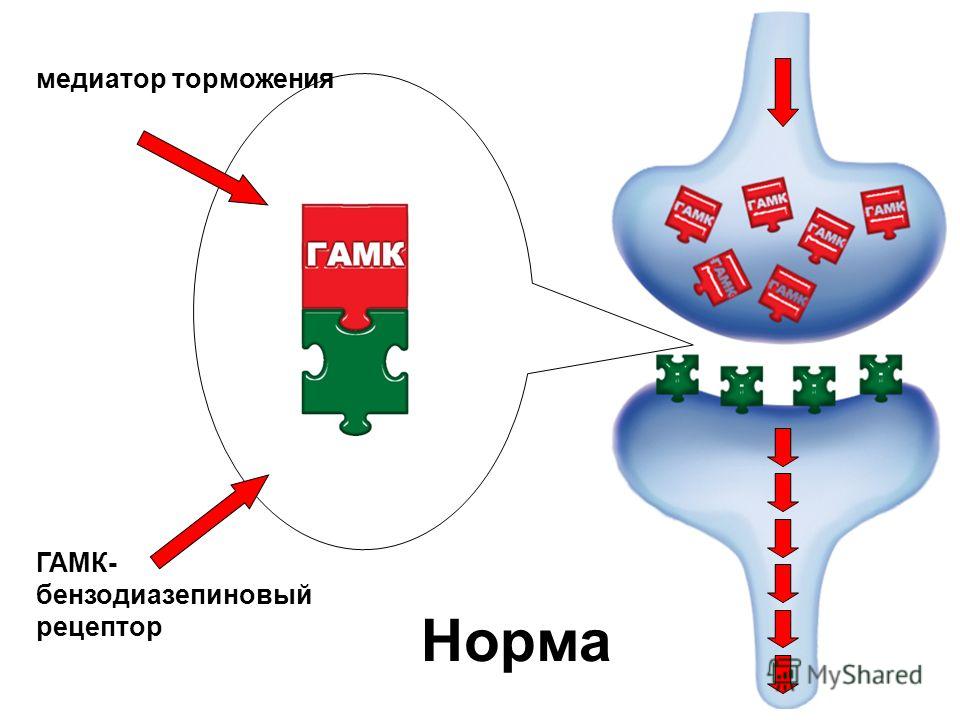 Следует отметить, что гормон роста встречается в 100 различных изоформах и что действие изоформ irGH и ifGH может отличаться от действия наиболее распространённой изоформы 22kDa.4)
Следует отметить, что гормон роста встречается в 100 различных изоформах и что действие изоформ irGH и ifGH может отличаться от действия наиболее распространённой изоформы 22kDa.4)
Функция
Медиатор
У позвоночных, ГАМК действует на тормозные синапсы в мозге путем связывания со специфическими трансмембранными рецепторами в плазматической мембране, относящимся к до- и постсинаптическим нейрональным процессам. Это связывание вызывает открытие ионных каналов, позволяя потоку отрицательно заряженных ионов хлора проникать в клетку или осуществляя вывод положительно заряженных ионов калия из клетки. Это приводит к негативным изменениям трансмембранного потенциала, и, как правило, вызывает гиперполяризацию. Известно два общих класса рецепторов ГАМК: ГАМКА, где рецептор является частью лиганд-закрытого комплекса ионных каналов; и метаботропные рецепторы ГАМКB, представляющие собой G-белковые рецепторы, которые открывают или закрывают ионные каналы через действие посредников (G белков).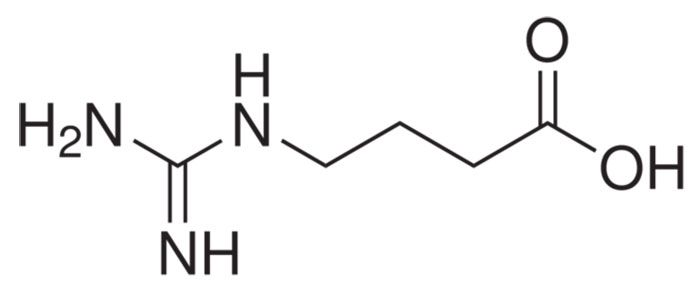 Нейроны, которые производят ГАМК, называются ГАМКергическими нейронами. Они проявляют в основном тормозящее действие на рецепторы у взрослых позвоночных. Средние шипиковые клетки – это типичный пример ингибирующих ГАМКергических клеток ЦНС. В противоположность этому, ГАМК оказывает возбуждающее и ингибирующее воздействие на насекомых, опосредуя мышечную активацию в синапсах между нервными и мышечными клетками, а также стимулируя некоторые железы. У млекопитающих, некоторые ГАМКергические нейроны, такие как канделябровидные клетки, также могут возбуждать их глутаматергические посредники. 5) ГАМКА-рецепторы представляют собой лиганд-активированные хлоридные каналы; то есть, активируясь ГАМК, они позволяют потоку хлорид-ионов проникать через мембрану клетки. Является ли поток хлоридов возбуждающим/деполяризующим (нейтрализующим отрицательное напряжение на мембране клетки), маневренным (не оказывающим никакого влияния на мембрану клетки) или ингибирующим/гиперполяризующим (делающим мембрану ячейки более отрицательной), зависит от направления потока хлорида.
Нейроны, которые производят ГАМК, называются ГАМКергическими нейронами. Они проявляют в основном тормозящее действие на рецепторы у взрослых позвоночных. Средние шипиковые клетки – это типичный пример ингибирующих ГАМКергических клеток ЦНС. В противоположность этому, ГАМК оказывает возбуждающее и ингибирующее воздействие на насекомых, опосредуя мышечную активацию в синапсах между нервными и мышечными клетками, а также стимулируя некоторые железы. У млекопитающих, некоторые ГАМКергические нейроны, такие как канделябровидные клетки, также могут возбуждать их глутаматергические посредники. 5) ГАМКА-рецепторы представляют собой лиганд-активированные хлоридные каналы; то есть, активируясь ГАМК, они позволяют потоку хлорид-ионов проникать через мембрану клетки. Является ли поток хлоридов возбуждающим/деполяризующим (нейтрализующим отрицательное напряжение на мембране клетки), маневренным (не оказывающим никакого влияния на мембрану клетки) или ингибирующим/гиперполяризующим (делающим мембрану ячейки более отрицательной), зависит от направления потока хлорида. При вытекании чистого хлорида из клетки, ГАМК является возбуждающим или деполяризующим; когда чистый хлорид впадает в клетку, ГАМК является ингибирующим или гиперполяризующим. Когда чистый поток хлорида близок к нулю, действие ГАМК является маневренным. Маневренное ингибирование не оказывает прямого влияния на мембранный потенциал клетки; однако, оно минимизирует влияние любых совпадающих синаптических вхождений, главным образом, за счет снижения электрического сопротивления клеточной мембраны (в сущности, это эквивалентно закону Ома). Изменение развития в молекулярной концентрации техники управления хлорида внутри клетки – и, следовательно, направление этого потока ионов, отвечает за изменения в функциональной роли ГАМК у новорожденных и взрослых. То есть, по мере развития мозга в зрелом возрасте, ГАМК меняет свою роль от возбуждающей к ингибирующей.
При вытекании чистого хлорида из клетки, ГАМК является возбуждающим или деполяризующим; когда чистый хлорид впадает в клетку, ГАМК является ингибирующим или гиперполяризующим. Когда чистый поток хлорида близок к нулю, действие ГАМК является маневренным. Маневренное ингибирование не оказывает прямого влияния на мембранный потенциал клетки; однако, оно минимизирует влияние любых совпадающих синаптических вхождений, главным образом, за счет снижения электрического сопротивления клеточной мембраны (в сущности, это эквивалентно закону Ома). Изменение развития в молекулярной концентрации техники управления хлорида внутри клетки – и, следовательно, направление этого потока ионов, отвечает за изменения в функциональной роли ГАМК у новорожденных и взрослых. То есть, по мере развития мозга в зрелом возрасте, ГАМК меняет свою роль от возбуждающей к ингибирующей.
Развитие мозга
В то время как ГАМК является ингибирующим медиатором в зрелом мозге, в развивающемся мозге его действие в первую очередь является возбуждающим. Градиент хлорида восстанавливается в незрелых нейронах, и его потенциал реверсии выше, чем мембранный потенциал покоя клеток; активация ГАМК-А рецептора, таким образом, приводит к оттоку Cl-ионов из клетки, т.е. деполяризующего тока. Дифференциальный градиент хлорида в незрелых нейронах, в первую очередь, зависит от более высокой концентрации ко-транспортеров NKCC1 относительно ко-транспортеров KCC2 в незрелых клетках. Сам ГАМК является частично ответственным за созревание ионных насосов. ГАМКергические интернейроны быстрее созревают в гиппокампе и сигнальное устройство ГАМК возникает раньше глутаматергической передачи. Таким образом, ГАМК является основным возбуждающим нейромедиатором во многих областях головного мозга перед созреванием глутаматэргических синапсов.
Однако эта теория была поставлена под сомнение на основании результатов, показывающих, что в срезах мозга незрелых мышей, инкубированных в искусственную спинномозговую жидкость (с изменениями, учитывающими нормальный состав нейронной среды путем добавления альтернативы энергетического субстрата бета-оксибутирата в глюкозу), ГАМК меняет свое действие с возбуждающего на ингибирующее.
Градиент хлорида восстанавливается в незрелых нейронах, и его потенциал реверсии выше, чем мембранный потенциал покоя клеток; активация ГАМК-А рецептора, таким образом, приводит к оттоку Cl-ионов из клетки, т.е. деполяризующего тока. Дифференциальный градиент хлорида в незрелых нейронах, в первую очередь, зависит от более высокой концентрации ко-транспортеров NKCC1 относительно ко-транспортеров KCC2 в незрелых клетках. Сам ГАМК является частично ответственным за созревание ионных насосов. ГАМКергические интернейроны быстрее созревают в гиппокампе и сигнальное устройство ГАМК возникает раньше глутаматергической передачи. Таким образом, ГАМК является основным возбуждающим нейромедиатором во многих областях головного мозга перед созреванием глутаматэргических синапсов.
Однако эта теория была поставлена под сомнение на основании результатов, показывающих, что в срезах мозга незрелых мышей, инкубированных в искусственную спинномозговую жидкость (с изменениями, учитывающими нормальный состав нейронной среды путем добавления альтернативы энергетического субстрата бета-оксибутирата в глюкозу), ГАМК меняет свое действие с возбуждающего на ингибирующее.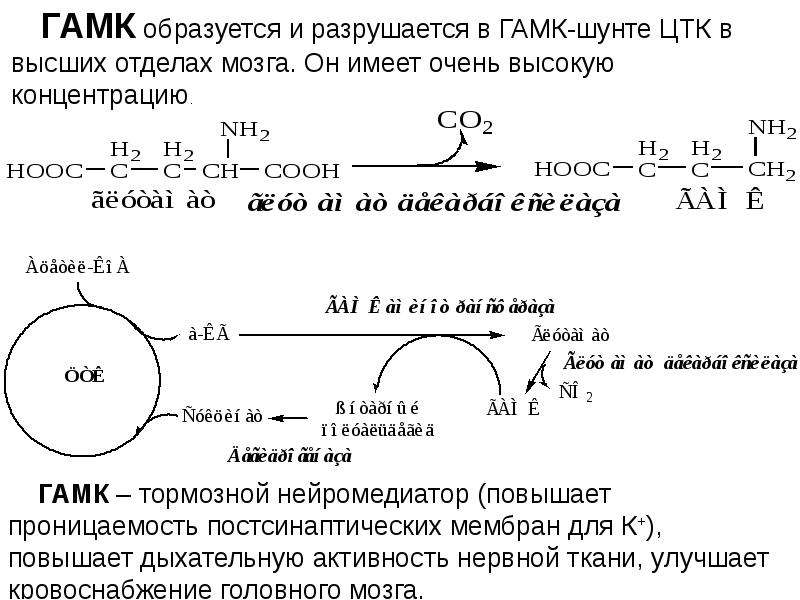 6) Этот эффект был позже повторен с использованием других энергетических субстратов, пирувата и лактата, дополняющих глюкозу в среде. Более поздние исследования метаболизма пирувата и лактата показали, что первоначальные результаты были связаны не с источником энергии, а с изменением рН в результате того, что субстраты действовали как «слабые кислоты». Эти аргументы были позже опровергнуты дальнейшими выводами, которые показывают, что изменения рН, большие, чем изменения, вызванные энергетическими субстратами, не влияют на ГАМК-сдвиг в присутствии энергетического субстрата ACSF, и что механизм действия бета-гидроксибутирата, пирувата и лактата (оцениваемый путем измерения NAD(P)H и утилизации кислорода) был связан с энергетическим метаболизмом.
В стадии развития, предшествующей формированию синаптических контактов, ГАМК синтезируется нейронами и действует в качестве аутокринного (воздействующего на ту же клетку) и паракринного (действующего на близлежащие клетки) сигнализационного медиатора.
6) Этот эффект был позже повторен с использованием других энергетических субстратов, пирувата и лактата, дополняющих глюкозу в среде. Более поздние исследования метаболизма пирувата и лактата показали, что первоначальные результаты были связаны не с источником энергии, а с изменением рН в результате того, что субстраты действовали как «слабые кислоты». Эти аргументы были позже опровергнуты дальнейшими выводами, которые показывают, что изменения рН, большие, чем изменения, вызванные энергетическими субстратами, не влияют на ГАМК-сдвиг в присутствии энергетического субстрата ACSF, и что механизм действия бета-гидроксибутирата, пирувата и лактата (оцениваемый путем измерения NAD(P)H и утилизации кислорода) был связан с энергетическим метаболизмом.
В стадии развития, предшествующей формированию синаптических контактов, ГАМК синтезируется нейронами и действует в качестве аутокринного (воздействующего на ту же клетку) и паракринного (действующего на близлежащие клетки) сигнализационного медиатора.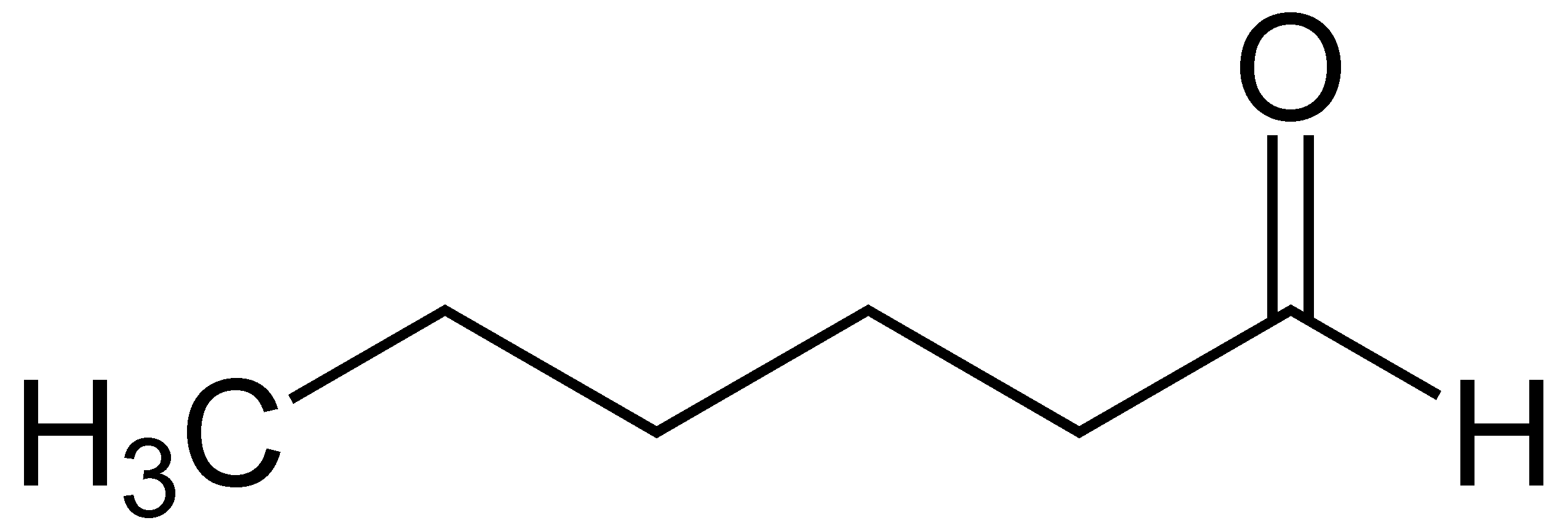 Ганглиозные возвышения также в значительной степени способствуют наращиванию ГАМКергической корковой клеточной популяции.
ГАМК регулирует пролиферацию нервных клеток-предшественников, миграцию и дифференцировку, удлинение нейритов и формирование синапсов. 7) ГАМК также регулирует рост эмбриональных и нервных стволовых клеток. ГАМК может влиять на развитие нервных клеток-предшественников с помощью экспрессии мозгового нейротрофического фактора. ГАМК активизирует рецептор ГАМКА, вызывая остановку клеточного цикла в S-фазе, ограничивая рост.
Ганглиозные возвышения также в значительной степени способствуют наращиванию ГАМКергической корковой клеточной популяции.
ГАМК регулирует пролиферацию нервных клеток-предшественников, миграцию и дифференцировку, удлинение нейритов и формирование синапсов. 7) ГАМК также регулирует рост эмбриональных и нервных стволовых клеток. ГАМК может влиять на развитие нервных клеток-предшественников с помощью экспрессии мозгового нейротрофического фактора. ГАМК активизирует рецептор ГАМКА, вызывая остановку клеточного цикла в S-фазе, ограничивая рост.
Действие ГАМК вне нервной системы
ГАМКергические механизмы были продемонстрированы на различных периферических тканях и органах, включая кишечник, желудок, поджелудочную железу, фаллопиевы трубы, матку, яичники, семенники, почки, мочевой пузырь, легкие и печень.
В 2007 году была описана возбудительная ГАМКергическая нервная система в эпителии дыхательных путей. Система активирует последующее воздействие аллергенов и может участвовать в механизмах астмы. ГАМКергические системы были также обнаружены в яичках и в хрусталике глаза.
ГАМКергические системы были также обнаружены в яичках и в хрусталике глаза.
Структура и конформация
ГАМК существует в основном в виде цвиттер-иона, то есть, с депротонированной карбоксигруппой и протонированной аминогруппой. Его конформация зависит от окружающей его среды. В газовой фазе, высокая конформация более предпочтительна из-за электростатического притяжения между двумя функциональными группами. Стабилизация составляет около 50 ккал / моль, согласно квантовым химическим расчетам. В твердом состоянии конформация более расширена, с транс-конформацией на амино-конце и гош-конформацией на карбоксильном конце. Это связано с взаимодействиями с соседними молекулами. В растворе пять различных конформаций (некоторые из которых сложенные, и некоторые – расширенные), присутствуют благодаря эффектам сольватации. Конформационная гибкость ГАМК имеет важное значение для его биологической функции, поскольку было установлено, что ГАМК связывается с различными рецепторами с различными конформациями. Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, и лучше контролируют связывание. 8)
Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, и лучше контролируют связывание. 8)
История
Гамма-аминомасляная кислота была впервые синтезирована в 1883 году, и изначально была известна только в качестве растения и продукта метаболизма микробов. В 1950 году, однако, было обнаружено, что ГАМК является неотъемлемой частью центральной нервной системы млекопитающих.
Биосинтез
ГАМК не проникает через гематоэнцефалический барьер; он синтезируется в мозге из глутамата с участием фермента L-глутаминовой кислоты декарбоксилазы и пиридоксаль фосфата (который является активной формой витамина В6) в качестве кофактора. ГАМК преобразуется обратно в глутамат в метаболическом пути под названием ГАМК шунт. В ходе этого процесса глутамат, основной возбуждающий нейромедиатор, преобразуется в главный тормозной нейромедиатор (ГАМК).9)
Катаболизм
Фермент ГАМК-трансаминазы катализирует превращение 4-аминобутановой кислоты и 2-оксоглутарата в янтарный полуальдегид и глутамат. Янтарный полуальдегид затем окисляют в янтарную кислоту при помощи янтарной полуальдегиддегидрогеназы. Как таковое, вещество входит в цикл лимонной кислоты в качестве полезного источника энергии.
Янтарный полуальдегид затем окисляют в янтарную кислоту при помощи янтарной полуальдегиддегидрогеназы. Как таковое, вещество входит в цикл лимонной кислоты в качестве полезного источника энергии.
Фармакология
Препараты, которые действуют как аллостерические модуляторы ГАМК-рецепторов (так называемые ГАМК аналоги или ГАМКергические препараты) и препараты, увеличивающие доступный объем ГАМК, обычно оказывают успокаивающее, антистрессовое и антисудорожное действие. Многие из перечисленных ниже веществ вызывают антероградную амнезию и ретроградную амнезию.
ГАМК не может пересекать гематоэнцефалический барьер, хотя некоторые области мозга, которые не имеют эффективного гематоэнцефалического барьера, например, перивентрикулярное ядро, могут быть доступны воздействию ГАМК при системном введении. По крайней мере, одно исследование показывает, что при пероральном приеме ГАМК увеличивает количество гормона роста человека. При впрыскивании ГАМК непосредственно в мозг, вещество проявляет как стимулирующее, так и тормозящее действие на производство гормона роста, в зависимости от физиологии человека. Были разработаны некоторые пролекарства ГАМК (напр. пикамилон), способные проникать через гематоэнцефалический барьер, и делиться на ГАМК и молекулу-носитель уже внутри мозга. Это позволяет прямо увеличивать уровень ГАМК во всех областях мозга.
Были разработаны некоторые пролекарства ГАМК (напр. пикамилон), способные проникать через гематоэнцефалический барьер, и делиться на ГАМК и молекулу-носитель уже внутри мозга. Это позволяет прямо увеличивать уровень ГАМК во всех областях мозга.
ГАМКергические препараты
Лиганды рецепторов ГАМКА
Агонисты / Позитивные аллостерические модуляторы: этанол, барбитураты, бензодиазепины, каризопродол, хлоралгидрат, этаквалон, этомидат, глутетимид, кава, метаквалон, мусцимол, нейроактивные стероиды, Z-препараты, пропофол, Scullcap, валериана, теанин, летучие / ингаляционные анестетики.
Антагонисты / Отрицательные аллостерические модуляторы: бикукуллин, цикутоксин, флумазенил, фуросемид, габазин, оэнантотоксин, пикротоксин, RO15-4513, туйон.Лиганды рецепторов ГАМКB
Агонисты: [[баклофен|баклофен]], ГБЛ, пропофол, ГОМК, фенибут.
Антагонисты: факлофен, саклофен. Ингибиторы обратного захвата ГАМК: дерамциклан, гиперфорин, тиагабин.
Ингибиторы ГАМК-трансаминазы: габакулин, фенелзин, вальпроат, вигабатрин, мелисса
Аналоги ГАМК: прегабалин, габапентин.
Другие: ГАМК (сам), L-глутамин, пикамилон, прогабид.
ГАМК в качестве дополнения
Ряд коммерческих источников продают формулы ГАМК для использования в качестве пищевой добавки, иногда для подъязычного введения, несмотря на то, что еще не была продемонстрирована эффективность ГАМК в качестве транквилизатора. Однако, есть также более научные и медицинские доказательства того, что чистый ГАМК не пересекает гематоэнцефалический барьер в терапевтических значимых дозах. Единственным способом эффективной доставки ГАМК является обхождение гематоэнцефалического барьера. В действительности, существует небольшое и ограниченное количество отпускаемых без рецепта (в США) добавок, которые являются производными ГАМК, таких как фенибут и пикамилон. Пикамилон – это сочетание ниацина и ГАМК. Вещество пересекает гематоэнцефалический барьер в качестве пролекарства, которое позже гидролизуется в ГАМК и никотиновую кислоту.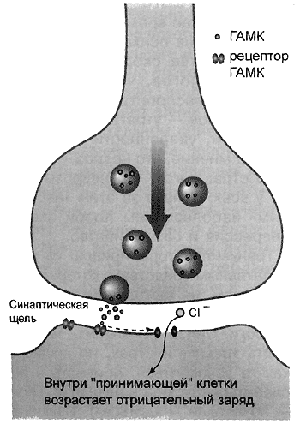
В растениях
ГАМК также содержится в растениях. Это наиболее распространенная аминокислота в апопласте томатов. Она также может играть определенную роль в клеточной сигнализации у растений.
Купить гамма-аминомасляную кислоту (ГАМК), Аминалон
Гамма-аминомасляная кислота (ГАМК) является одним из основных медиаторов торможения как в периферической, так и в центральной нервной системе.Она играет существенную роль в углеводном и аминокислотном обмене в головном мозге, способствует нормализации метаболических процессов в нервной системе. Под влиянием ГАМК активируются энергетические процессы мозга, повышается дыхательная активность тканей, улучшается утилизация мозгом глюкозы, улучшается кровоснабжение. ГАМК оказывает стимулирующее действие на цикл Кребса. Она может служить субстратом окисления в мозговой ткани, вызывает общее увеличение содержания аминокислот в мозгу. Существуют данные о том, что эта кислота участвует в пресинаптическом торможении в качестве медиатора в аксо-аксонных синапсах. В России ГАМК отпускается из аптек под торговой маркой Аминалон. Препарат отпускается по рецепту врача.
Кроме того, существуют следующие аналоги ГАМК: Гаммалон, Пентоган, Фенибут.
В России ГАМК отпускается из аптек под торговой маркой Аминалон. Препарат отпускается по рецепту врача.
Кроме того, существуют следующие аналоги ГАМК: Гаммалон, Пентоган, Фенибут.
:Tags
Читать еще: Аппендицит , Витамины группы B , Леводопа (L-ДОФА) , Эргостерол , Фенхель (Эфирное масло фенхеля) ,
Список использованной литературы:
1) Petroff OA. GABA and glutamate in the human brain. Neuroscientist. (2002) 2) Transport of GABA at the Blood-CSF Interface 3) GABA in plasma and cerebrospinal fluid of different species. Effects of γ-acetylenic GABA, γ-vinyl GABA and sodium valproate 4) Efflux of a suppressive neurotransmitter, GABA, across the blood–brain barrier 5) Blood-brain barrier to h4-γ-aminobutyric acid in normal and amino oxyacetic acid-treated animals 6) Cavagnini F, et al. Effect of acute and repeated administration of gamma aminobutyric acid (GABA) on growth hormone and prolactin secretion in man.
 Acta Endocrinol (Copenh). (1980) 7) Shyamaladevi N, et al. Evidence that nitric oxide production increases gamma-amino butyric acid permeability of blood-brain barrier. Brain Res Bull. (2002) 8) Cavagnini F, et al. Effect of gamma-aminobutyric acid on growth hormone and prolactin secretion in man: influence of pimozide and domperidone. J Clin Endocrinol Metab. (1980) 9) Cavagnini F, et al. Effect of gamma-aminobutyric acid on growth hormone and prolactin secretion in man: influence of pimozide and domperidone. J Clin Endocrinol Metab. (1980)
Acta Endocrinol (Copenh). (1980) 7) Shyamaladevi N, et al. Evidence that nitric oxide production increases gamma-amino butyric acid permeability of blood-brain barrier. Brain Res Bull. (2002) 8) Cavagnini F, et al. Effect of gamma-aminobutyric acid on growth hormone and prolactin secretion in man: influence of pimozide and domperidone. J Clin Endocrinol Metab. (1980) 9) Cavagnini F, et al. Effect of gamma-aminobutyric acid on growth hormone and prolactin secretion in man: influence of pimozide and domperidone. J Clin Endocrinol Metab. (1980)гамма-аминомасляная_кислота.txt · Последнее изменение: 2021/05/01 08:59 — dr.cookie
Нейромолекулы: гамма-аминомасляная кислота или просто ГАМК
Работу нервной системы не представляется возможным описать в двух словах. Но если бы неведомая сила всё-таки принудила нас выразить так её сущность, то первым словом было бы «возбуждение», а вторым — «торможение». Ведь именно баланс этих двух процессов порождает всё многообразие реакций живых существ на сигналы извне. Этими явлениями в центральной нервной системе (ЦНС) управляют аминокислоты — базовые биохимические молекулы, которых называют нейромедиаторами. Их три (на самом деле больше, но эти — основные): глутаминовая кислота (наиболее важный медиатор возбуждения), гамма-аминомасляная кислота и глицин (отвечают за торможение). Сейчас мы поговорим о тормозных нейромедиаторах, а точнее — о гамма-аминомасляной кислоте.
Ведь именно баланс этих двух процессов порождает всё многообразие реакций живых существ на сигналы извне. Этими явлениями в центральной нервной системе (ЦНС) управляют аминокислоты — базовые биохимические молекулы, которых называют нейромедиаторами. Их три (на самом деле больше, но эти — основные): глутаминовая кислота (наиболее важный медиатор возбуждения), гамма-аминомасляная кислота и глицин (отвечают за торможение). Сейчас мы поговорим о тормозных нейромедиаторах, а точнее — о гамма-аминомасляной кислоте.
Молекула ГАМК
Давным-давно, когда мир ещё не знал об Интернете, постмодернизме и оружии массового поражения, один из наиболее влиятельных физиологов второй половины XIX в. Иван Михайлович Сеченов в своих работах показал существование в ЦНС явления торможения. В ранних опытах Сеченова стимуляция определённых центров нервной системы лягушки подавляла рефлекторное движение лапки, выполняя таким образом функцию «тормоза» для этого рефлекса. В те времена дать ответ на вопрос о том, как осуществляется торможение в нервной системе на клеточном и молекулярном уровне, было невозможно. Для этого требовалось несколько десятков лет постепенного накопления данных, добытых трудолюбивыми учёными в лабораториях по всему миру, в том числе и в России (вспомнить хотя бы нобелевского лауреата Ивана Петровича Павлова). Именно благодаря их труду мы теперь имеем возможность спуститься по воображаемой иерархической лестнице до уровня молекул и разобраться подробнее, что же именно открыл Сеченов.
В те времена дать ответ на вопрос о том, как осуществляется торможение в нервной системе на клеточном и молекулярном уровне, было невозможно. Для этого требовалось несколько десятков лет постепенного накопления данных, добытых трудолюбивыми учёными в лабораториях по всему миру, в том числе и в России (вспомнить хотя бы нобелевского лауреата Ивана Петровича Павлова). Именно благодаря их труду мы теперь имеем возможность спуститься по воображаемой иерархической лестнице до уровня молекул и разобраться подробнее, что же именно открыл Сеченов.
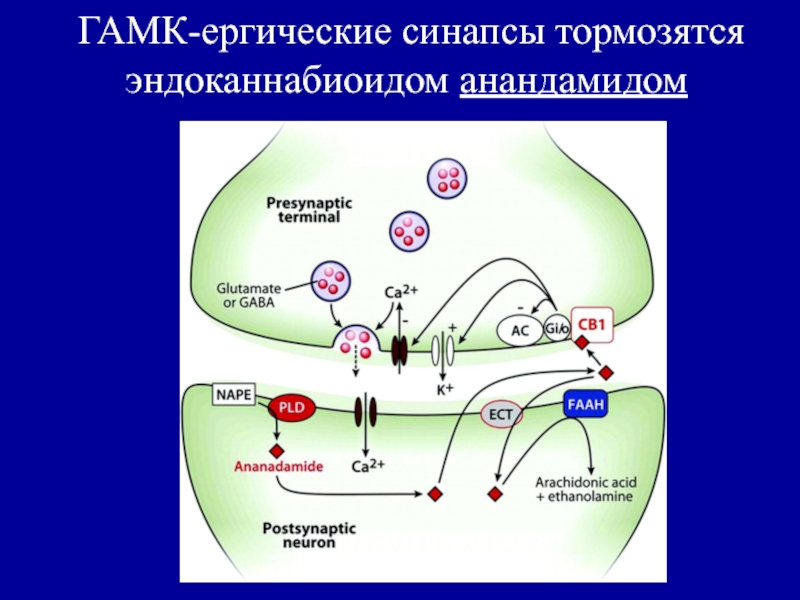
Формула ГАМК
Основной медиатор торможения в ЦНС — аминокислота ГАМК (гамма-аминомасляная кислота). В англоязычных источниках мы видим её под аббревиатурой GABA, поскольку в английском языке она носит более формальное химическое название – гамма-аминобутановая кислота. Что это значит на практике? Почти в любом регионе мозга можно найти нейроны, несущие на себе рецепторы молекул ГАМК. При этом активация этих рецепторов приводит к гиперполяризации мембраны нейрона (об этом процессе мы писали в статье про потенциал действия).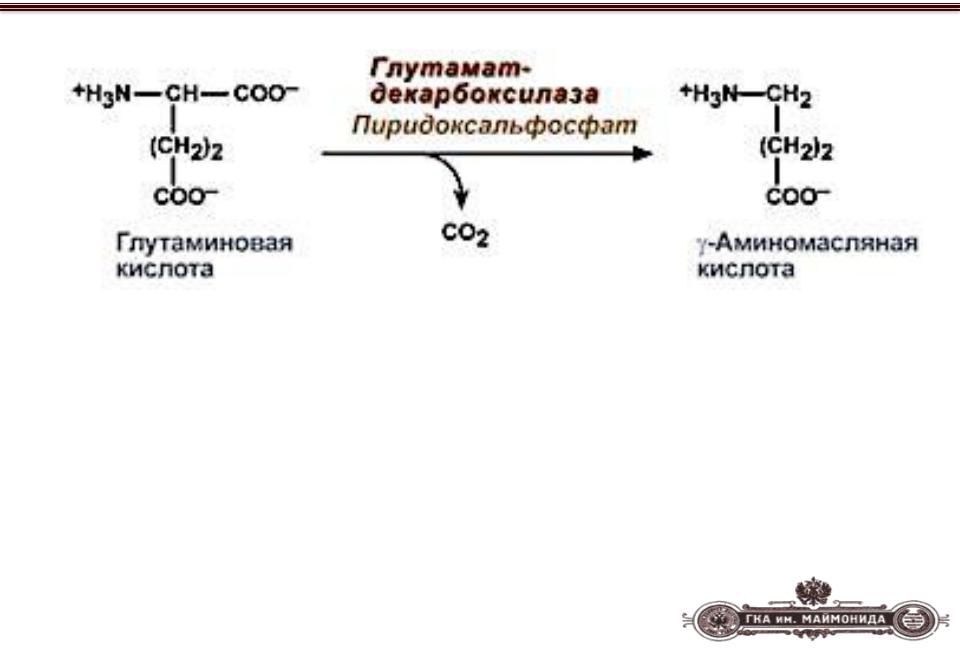 Такой нейрон становится на некоторое время менее восприимчивым к активирующим (возбуждающим) сигналам от других нервных клеток. Таким образом, ГАМК выполняет функцию молекулярного тормоза, регулирующего активность нейронов.
Такой нейрон становится на некоторое время менее восприимчивым к активирующим (возбуждающим) сигналам от других нервных клеток. Таким образом, ГАМК выполняет функцию молекулярного тормоза, регулирующего активность нейронов.
Как нейрон доходит до этого состояния? Рецепторы ГАМК можно разделить на две группы – ионотропные и метаботропные. Метаботропные рецепторы относятся к обширному семейству GPCR (G-protein coupled receptors; рецепторы, сопряженные с G-белками). Они связываются с молекулами ГАМК и пробуждают ферменты, которые активируют калиевые каналы, то есть транспортные белки, занимающиеся поставками калия сквозь мембрану клетки для передачи нервных сигналов.
Ионотропные рецепторы (или лиганд-управляемые ионные каналы) сами представляют собой ионные каналы, проницаемые для ионов хлора. Активация и калиевых, и хлорных каналов преследует одну цель: затормозить нервные импульсы (положительно заряженные ионы калия покидают клетку, делая её заряженной отрицательно относительно внешней среды; отрицательно заряженные ионы хлора проникают внутрь клетки, придавая ей дополнительный отрицательный заряд).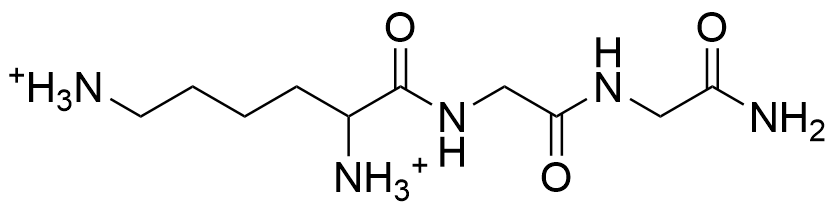
Широкая распространённость рецепторов ГАМК в нервной системе сделала возможной разработку множества лекарств, молекулы которых взаимодействуют с различными типами этих рецепторов. Например, бензодиазепины – лекарства, обладающие успокоительным эффектом – работают как положительные модуляторы ионотропных рецепторов ГАМК. Молекулы бензодиазепинов связываются с рецептором и делают его более чувствительным к воздействию природной гамма-аминомасляной кислоты, что приводит к «затормаживанию» некоторых процессов в мозге и медикаментозному успокоению.
В последние годы идёт разработка и негативных модуляторов рецепторов ГАМК для стимуляции работы нервной системы при состояниях, когда она слишком «заторможена» от природы или в результате воздействия каких-либо веществ, например, алкоголя.
ГАМК-рецептор
Текст: Денис Кудрявцев (научный сотрудник Лаборатории лиганд-рецепторных взаимодействий Института биоорганической химии РАН)
Другие нейромедиаторы:
дофамин
серотонин.
Гамма-аминомасляная кислота | Психология вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
Гамма-аминомасляная кислота (обычно сокращенно ГАМК ) представляет собой тормозящий нейротрансмиттер, обнаруживаемый в нервных системах широко различных видов.Это главный тормозной нейромедиатор в центральной нервной системе, а также в сетчатке. ГАМК — это аминокислота, но не содержится в белках. Хотя некоторое количество ГАМК можно найти в островковых клетках поджелудочной железы и почках, в тканях млекопитающих, кроме тканей нервной системы, нет значительных количеств ГАМК.
Функция []
У позвоночных ГАМК действует на тормозные синапсы в головном мозге. ГАМК действует путем связывания со специфическими трансмембранными рецепторами плазматической мембраны как пре-, так и постсинаптических нейронов.Это связывание вызывает открытие ионных каналов, позволяя потоку либо отрицательно заряженных ионов хлорида в клетку, либо положительно заряженных ионов калия из клетки. Это действие приводит к отрицательному изменению трансмембранного потенциала, обычно вызывая гиперполяризацию. Известны три основных класса рецепторов ГАМК: ГАМК A и ГАМК C ионотропные рецепторы, которые сами являются ионными каналами, и метаботропные рецепторы ГАМК B , которые представляют собой рецепторы, связанные с G-белком, которые открываются. ионные каналы через посредников (G-белки).
Нейроны, вырабатывающие ГАМК в качестве продукции, называются ГАМКергическими нейронами, и они оказывают главным образом ингибирующее действие на рецепторы у взрослых позвоночных. Средние колючие клетки являются типичным примером ингибирующих ГАМКергических клеток ЦНС. ГАМК проявляет возбуждающее действие у насекомых, опосредуя активацию мышц в синапсах между нервами и мышечными клетками, а также стимулируя определенные железы. В гиппокампе и неокортексе мозга млекопитающих ГАМК оказывает в первую очередь возбуждающее действие на ранней стадии развития и фактически является основным возбуждающим нейромедиатором во многих областях мозга до созревания глутаматных синапсов — См. развивающаяся кора .Является ли ГАМК возбуждающей или тормозящей, зависит от направления (внутрь или из клетки) и величины ионных токов, контролируемых рецептором ГАМК A . Когда чистый положительный ионный ток направлен в клетку, ГАМК является возбуждающим, когда чистый положительный ток направлен из клетки, ГАМК оказывает тормозящее действие. Переключатель развития в молекулярном механизме, контролирующий полярность этого тока, ответственен за изменения функциональной роли ГАМК между стадиями новорожденных и взрослых.
Средние колючие клетки являются типичным примером ингибирующих ГАМКергических клеток ЦНС. ГАМК проявляет возбуждающее действие у насекомых, опосредуя активацию мышц в синапсах между нервами и мышечными клетками, а также стимулируя определенные железы. В гиппокампе и неокортексе мозга млекопитающих ГАМК оказывает в первую очередь возбуждающее действие на ранней стадии развития и фактически является основным возбуждающим нейромедиатором во многих областях мозга до созревания глутаматных синапсов — См. развивающаяся кора .Является ли ГАМК возбуждающей или тормозящей, зависит от направления (внутрь или из клетки) и величины ионных токов, контролируемых рецептором ГАМК A . Когда чистый положительный ионный ток направлен в клетку, ГАМК является возбуждающим, когда чистый положительный ток направлен из клетки, ГАМК оказывает тормозящее действие. Переключатель развития в молекулярном механизме, контролирующий полярность этого тока, ответственен за изменения функциональной роли ГАМК между стадиями новорожденных и взрослых.
При спастическом церебральном параличе у человека ГАМК не может должным образом абсорбироваться поврежденными нервными корешками, ведущими к определенным мышцам; это приводит к гипертонусу в этих мышцах.
Строение и внешний вид []
ГАМК находится в основном в виде цвиттериона, то есть с депротонированной карбоксильной группой и протонированной аминогруппой. Его внешний вид зависит от окружающей среды. В газовой фазе сильно сложена конформация из-за электростатического притяжения между двумя функциональными группами.Согласно расчетам квантовой химии, стабилизация составляет около 50 ккал / моль. В твердом состоянии обнаруживается более протяженная конформация с транс-конформацией на амино-конце и гош-конформацией на карбоксильном конце. Это связано с взаимодействием упаковки с соседними молекулами. В растворе в результате эффектов сольватации обнаруживаются пять различных конформаций, некоторые из которых свернуты, а некоторые расширены. Конформационная гибкость ГАМК важна для ее биологической функции, поскольку было обнаружено, что она связывается с разными рецепторами с разными конформациями. Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, чтобы лучше контролировать связывание. [1] [2]
Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, чтобы лучше контролировать связывание. [1] [2]
История []
Гамма-аминомасляная кислота была впервые синтезирована в 1883 году и сначала была известна только как продукт метаболизма растений и микробов. Однако в 1950 году было обнаружено, что ГАМК является неотъемлемой частью центральной нервной системы млекопитающих. [3]
Синтез []
Организмы синтезируют ГАМК из глутамата, используя в качестве кофактора фермент декарбоксилазу L-глутаминовой кислоты и пиридоксальфосфат.Стоит отметить, что этот процесс превращает основной возбуждающий нейромедиатор (глутамат) в основной тормозной (ГАМК).
Фармакология []
ГАМК — нейромедиатор, нацеленный на анксиолитические препараты, такие как валиум. Валиум имеет сайт связывания на рецепторах ГАМК, отличный от эндогенного нейромедиатора ГАМК. В результате валиум искусственно подавляет потенциал действия в этих передачах.
Лекарства, которые действуют как агонисты рецепторов ГАМК (известные как аналоги ГАМК или препараты GABAnergic ) или увеличивают доступное количество ГАМК, обычно обладают расслабляющим, успокаивающим и противосудорожным действием.Известно, что многие из перечисленных ниже веществ вызывают антероградную амнезию и ретроградную амнезию.
Предполагается, чтоГАМК увеличивает количество гормона роста человека. Результаты этих исследований редко воспроизводятся и в последнее время ставятся под сомнение, поскольку неизвестно, может ли ГАМК проходить гематоэнцефалический барьер.
Лекарства, влияющие на рецепторы ГАМК:
Лекарства, влияющие на ГАМК другими способами:
- тиагабин — потенцирует за счет ингибирования поглощения нейронами и глией
- вигабатрин — усиливает действие путем ингибирования ГАМК-Т, предотвращая распад ГАМК.
- вальпроат — потенцирует за счет ингибирования ГАМК-Т
- тетаноспазмин — первичный токсин столбнячных бактерий, блокирует высвобождение ГАМК.
- гиперфорин — подавляет обратный захват ГАМК
ГАМК и настроение []
В то время как гамма-аминомасляная кислота (ГАМК) в качестве тормозного нейромедиатора используется для успокоения нейронов коры и гиппокампа.Глутамат — возбуждающий нейротрансмиттер, подавляемый ГАМК в синапсе. Астроциты регулируют настроение, высвобождая и поглощая глутамат, а также за счет использования рецепторов ГАМК.
См. Также []
Ссылки []
- ↑ Деваши Маджумдар и Сефали Гуха. Конформация, электростатический потенциал и фармакофорная структура ГАМК (гамма-аминомасляная кислота) и некоторых ингибиторов ГАМК. Журнал молекулярной структуры: ТЕОХИМА 1988 , 180 , 125-140.DOI: 10.1016 / 0166-1280 (88) 80084-8
- ↑ Энн-Мари Сапсе. Расчет молекулярных орбиталей аминокислот и пептидов . Биркхойзер, 2000 . ISBN 0817638938.
- ↑ Roth, Robert J .; Купер, Джек Р .; Блум, Флойд Э. (2003). Биохимические основы нейрофармакологии , 416 страниц, Оксфорд [Оксфордшир]: Oxford University Press.
- ↑ Дзитоева С., Димитриевич Н., Манев Х. (2003). Рецептор 1 гамма-аминомасляной кислоты опосредует действия алкоголя у дрозофилы, нарушающие поведение: интерференция РНК взрослых и фармакологические данные. Proc. Natl. Акад. Sci. США 100 (9): 5485-90.
- ↑ Михик С.Дж., Йе Кью, Вик М.Дж., Колчин В.В., Красовски М.Д., Финн С.Е., Маския М.П., Валенсуэла К.Ф., Хэнсон К.К., Гринблатт Е.П., Харрис Р.А., Харрисон Н.Л. (1997). Сайты алкоголя и летучих анестетиков действуют на рецепторы GABA A и глицина. Природа 389 (6649): 385-9.
- ↑ Бем С.Л., Пономарев И, Бледнов Ю.А., Харрис Р.А. (2006). От гена к поведению и обратно: новые взгляды на селективность действия алкоголя субъединицей рецептора GABA A . Adv. Pharmacol. 54 : 171-203.
- ↑ Granger P, Biton B, Faure C, Vige X, Depoortere H, Graham D, Langer SZ, Scatton B, Avenet P (1995). Модуляция рецептора гамма-аминомасляной кислоты типа А противоэпилептическими препаратами карбамазепином и фенитоином. Мол. Pharmacol. 47 (6): 1189–96.
- ↑ Димитриевич Н., Дзитоева С., Сатта Р., Имбеси М., Йылдыз С., Манев Х. (2005). GABA B рецепторы дрозофилы участвуют в поведенческих эффектах гамма-гидроксимасляной кислоты (GHB). Eur. J. Pharmacol. 519 (3): 246-52.
- ↑ Хантер, А (2006). Кава (Piper methysticum) снова в обращении. Австралийский центр дополнительной медицины 25 (7): 529.
 Биохимические основы нейрофармакологии , 416 страниц, Оксфорд [Оксфордшир]: Oxford University Press.
Биохимические основы нейрофармакологии , 416 страниц, Оксфорд [Оксфордшир]: Oxford University Press.  Модуляция рецептора гамма-аминомасляной кислоты типа А противоэпилептическими препаратами карбамазепином и фенитоином. Мол. Pharmacol. 47 (6): 1189–96.
Модуляция рецептора гамма-аминомасляной кислоты типа А противоэпилептическими препаратами карбамазепином и фенитоином. Мол. Pharmacol. 47 (6): 1189–96. Внешние ссылки []
гамма-аминомасляная кислота — Википедия переиздана // WIKI 2
Главный тормозной нейромедиатор в головном мозге млекопитающих
| Имена | |
|---|---|
| Предпочтительное название IUPAC | |
| Идентификаторы | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ChemSpider | |
| Лекарственный банк | |
| ECHA InfoCard | 100. 000,235 000,235 |
| Номер ЕС | |
| КЕГГ | |
| Меш | гамма-аминомасляная кислота + кислота |
| Номер RTECS | |
| UNII | |
| |
| Недвижимость | |
| C 4 H 9 NO 2 | |
| Молярная масса | 103.120 г / моль |
| Внешний вид | белый микрокристаллический порошок |
| Плотность | 1,11 г / мл |
| Температура плавления | 203,7 ° С (398,7 ° F, 476,8 К) |
| Температура кипения | 247,9 ° С (478,2 ° F, 521,0 К) |
| 130 г / 100 мл | |
| журнал P | −3,17 |
| Кислотность (p K a ) |
|
| Опасности | |
| Основные опасности | Раздражающий, Вредный |
| Смертельная доза или концентрация (LD, LC): | |
| 12,680 мг / кг (мышь, перорально) | |
| Если не указано иное, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа). | |
| N проверить (что такое YN ?) | |
| Ссылки на инфобокс | |
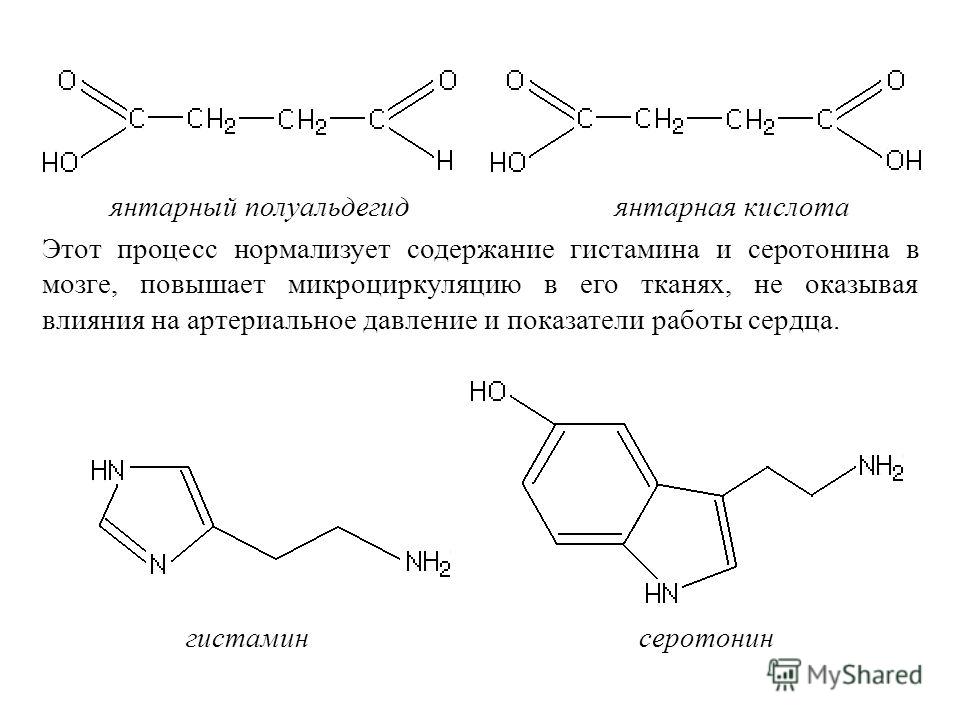 031 (карбоксил; H 2 O)
031 (карбоксил; H 2 O)Химический состав
гамма- Аминомасляная кислота , или гамма-аминомасляная кислота , или ГАМК , является главным тормозным нейромедиатором в центральной нервной системе зрелых млекопитающих.Его основная роль — снижение возбудимости нейронов по всей нервной системе.
ГАМК продается как пищевая добавка во многих странах. Традиционно считалось, что экзогенная ГАМК (т.е. принимаемая в качестве добавки) не проникает через гематоэнцефалический барьер, [2] , однако данные, полученные из более современных исследований [3] , показывают, что это возможно.
Функция
Нейротрансмиттер
Известны два общих класса рецепторов ГАМК: [4]
Цикл высвобождения, обратного захвата и метаболизма ГАМК
нейронов, вырабатывающих ГАМК в качестве продукции, называются ГАМКергические нейроны , и они оказывают главным образом ингибирующее действие на рецепторы у взрослых позвоночных.Средние шиповатые клетки являются типичным примером ингибирующих ГАМКергических клеток центральной нервной системы. Напротив, ГАМК проявляет как возбуждающее, так и тормозящее действие у насекомых, опосредуя активацию мышц в синапсах между нервами и мышечными клетками, а также стимулируя определенные железы. [6] У млекопитающих некоторые ГАМКергические нейроны, такие как клетки люстр, также способны возбуждать свои глутаматергические аналоги. [7]
[6] У млекопитающих некоторые ГАМКергические нейроны, такие как клетки люстр, также способны возбуждать свои глутаматергические аналоги. [7]
GABA A рецепторы представляют собой активируемые лигандами хлоридные каналы: при активации GABA они позволяют потоку хлорид-ионов через мембрану клетки. [8] Является ли этот поток хлорида деполяризующим (делает напряжение на мембране клетки менее отрицательным), шунтирующим (не влияет на мембранный потенциал клетки) или ингибирующим / гиперполяризационным (делает мембрану клетки более отрицательным) зависит от направление потока хлорида. Когда чистый хлорид вытекает из клетки, ГАМК деполяризируется; когда хлорид проникает в клетку, ГАМК тормозит или гиперполяризует. Когда чистый поток хлорида близок к нулю, действие ГАМК является шунтирующим.Ингибирование шунтирования не оказывает прямого воздействия на мембранный потенциал клетки; однако он снижает эффект любого совпадающего синаптического входа за счет уменьшения электрического сопротивления клеточной мембраны.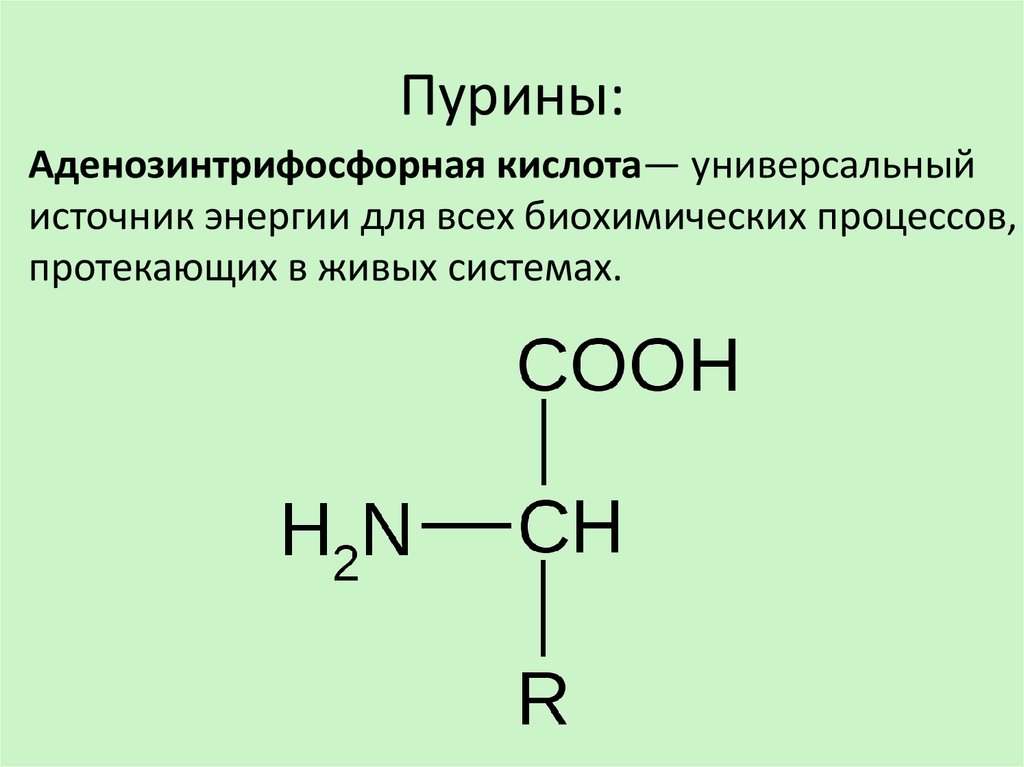 Ингибирование шунтирования может «преодолевать» возбуждающий эффект деполяризующей ГАМК, приводя к общему ингибированию, даже если мембранный потенциал становится менее отрицательным. Считалось, что онтогенетический переключатель в молекулярном механизме, контролирующем концентрацию хлорида внутри клетки, изменяет функциональную роль ГАМК между неонатальной и взрослой стадиями.По мере того как мозг становится взрослым, роль ГАМК меняется с возбуждающей на тормозящую. [9]
Ингибирование шунтирования может «преодолевать» возбуждающий эффект деполяризующей ГАМК, приводя к общему ингибированию, даже если мембранный потенциал становится менее отрицательным. Считалось, что онтогенетический переключатель в молекулярном механизме, контролирующем концентрацию хлорида внутри клетки, изменяет функциональную роль ГАМК между неонатальной и взрослой стадиями.По мере того как мозг становится взрослым, роль ГАМК меняется с возбуждающей на тормозящую. [9]
Развитие мозга
Хотя ГАМК является тормозящим передатчиком в зрелом мозге, считалось, что ее действие в первую очередь возбуждает в развивающемся мозге. [9] [10] Сообщалось, что градиент хлорида обращен в незрелых нейронах, с его обратным потенциалом выше, чем мембранный потенциал покоя клетки; Таким образом, активация рецептора GABA-A приводит к оттоку ионов Cl — из клетки (то есть к деполяризующему току).Было показано, что дифференциальный градиент хлорида в незрелых нейронах в первую очередь обусловлен более высокой концентрацией котранспортеров NKCC1 по сравнению с котранспортерами KCC2 в незрелых клетках. ГАМКергические интернейроны созревают быстрее в гиппокампе, и механизм передачи сигналов ГАМК появляется раньше, чем глутаматергическая передача. Таким образом, ГАМК считается основным возбуждающим нейромедиатором во многих областях мозга до созревания глутаматергических синапсов. [11]
ГАМКергические интернейроны созревают быстрее в гиппокампе, и механизм передачи сигналов ГАМК появляется раньше, чем глутаматергическая передача. Таким образом, ГАМК считается основным возбуждающим нейромедиатором во многих областях мозга до созревания глутаматергических синапсов. [11]
На стадиях развития, предшествующих формированию синаптических контактов, ГАМК синтезируется нейронами и действует как аутокринный (действующий на одну и ту же клетку), так и паракринный (действующий на соседние клетки) медиатор передачи сигналов. [12] [13] Ганглиозные возвышения также вносят большой вклад в создание популяции ГАМКергических кортикальных клеток. [14]
ГАМК регулирует пролиферацию нейральных клеток-предшественников, [15] [16] миграцию [17] и дифференцировку [18] [19] удлинение нейритов [20] и образование синапсов. [21]
ГАМК также регулирует рост эмбриональных и нервных стволовых клеток.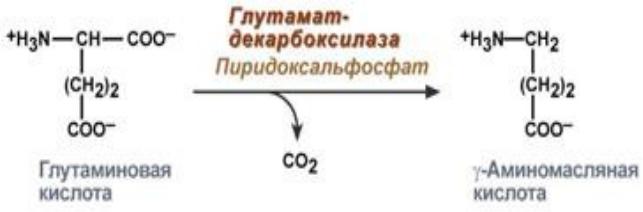 ГАМК может влиять на развитие нервных клеток-предшественников через экспрессию нейротрофического фактора головного мозга (BDNF). [22] ГАМК активирует рецептор ГАМК А , вызывая остановку клеточного цикла в S-фазе, ограничивая рост. [23]
ГАМК может влиять на развитие нервных клеток-предшественников через экспрессию нейротрофического фактора головного мозга (BDNF). [22] ГАМК активирует рецептор ГАМК А , вызывая остановку клеточного цикла в S-фазе, ограничивая рост. [23]
За пределами нервной системы
Помимо нервной системы, ГАМК также продуцируется на относительно высоком уровне в продуцирующих инсулин β-клетках поджелудочной железы. Β-клетки секретируют ГАМК вместе с инсулином, и ГАМК связывается с рецепторами ГАМК на соседних островковых α-клетках и препятствует их секреции глюкагона (что могло бы противодействовать эффектам инсулина). [25]
ГАМК может способствовать репликации и выживанию β-клеток [26] [27] [28] , а также способствовать превращению α-клеток в β-клетки, что может приводить к к новым методам лечения диабета. [29]
Помимо ГАМКергических механизмов, ГАМК также была обнаружена в других периферических тканях, включая кишечник, желудок, фаллопиевы трубы, матку, яичники, семенники, почки, мочевой пузырь, легкие и печень, хотя и на гораздо более низких уровнях, чем в нейронах или β-клетках. [30]
[30]
Эксперименты на мышах показали, что гипотиреоз, вызванный отравлением фтором, можно остановить введением ГАМК. Тест также показал, что щитовидная железа восстановилась естественным путем без дополнительной помощи после того, как фторид был удален ГАМК. [31]
Иммунные клетки экспрессируют рецепторы для ГАМК [32] [33] , и введение ГАМК может подавлять воспалительные иммунные ответы и способствовать «регуляторным» иммунным ответам, так что было показано, что введение ГАМК ингибирует аутоиммунные реакции. болезни на нескольких моделях животных. [26] [32] [34] [35]
В 2018 году было показано, что ГАМК регулирует секрецию большего количества цитокинов. В плазме пациентов с СД1 повышены уровни 26 цитокинов, из которых 16 ингибируются ГАМК в клеточных анализах. [36]
В 2007 году в эпителии дыхательных путей была описана возбуждающая ГАМКергическая система. Система активируется воздействием аллергенов и может участвовать в механизмах астмы. [37] ГАМКергические системы также были обнаружены в яичках [38] и в хрусталике глаза. [39]
Система активируется воздействием аллергенов и может участвовать в механизмах астмы. [37] ГАМКергические системы также были обнаружены в яичках [38] и в хрусталике глаза. [39]
ГАМК встречается в растениях. [40] [41]
Строение и внешний вид
ГАМК находится в основном в виде цвиттер-иона (т.е. с депротонированной карбоксильной группой и протонированной аминогруппой). Его внешний вид зависит от окружающей среды. В газовой фазе сильно сложена конформация из-за электростатического притяжения между двумя функциональными группами. Согласно расчетам квантовой химии, стабилизация составляет около 50 ккал / моль.В твердом состоянии обнаруживается расширенная конформация с транс-конформацией на амино-конце и гош-конформацией на карбоксильном конце. Это связано с взаимодействием упаковки с соседними молекулами. В растворе в результате эффектов сольватации обнаруживаются пять различных конформаций, некоторые из которых складчатые, а некоторые вытянутые.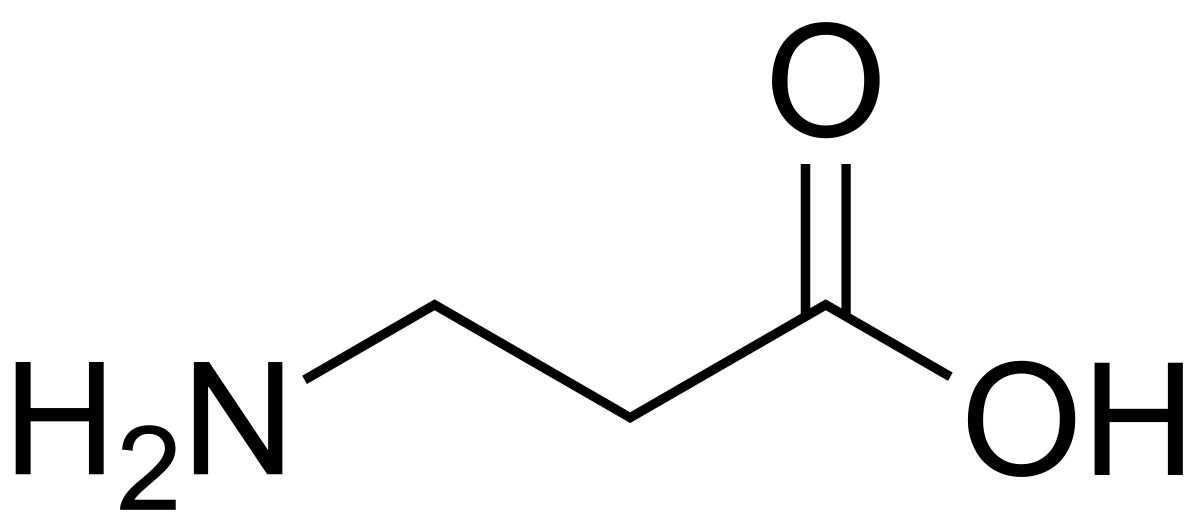 Конформационная гибкость ГАМК важна для ее биологической функции, поскольку было обнаружено, что она связывается с разными рецепторами с разными конформациями. Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, чтобы лучше контролировать связывание. [42] [43]
Конформационная гибкость ГАМК важна для ее биологической функции, поскольку было обнаружено, что она связывается с разными рецепторами с разными конформациями. Многие аналоги ГАМК, применяемые в фармацевтике, имеют более жесткие структуры, чтобы лучше контролировать связывание. [42] [43]
История
В 1883 году впервые была синтезирована ГАМК, и впервые она была известна только как продукт метаболизма растений и микробов. [44]
В 1950 году ГАМК была открыта как неотъемлемая часть центральной нервной системы млекопитающих. [44]
В 1959 году было показано, что при тормозящем синапсе мышечных волокон рака ГАМК действует как стимуляция тормозящего нерва. Как ингибирование нервной стимуляцией, так и применяемая ГАМК блокируются пикротоксином. [45]
Биосинтез
ГАМКергические нейроны, вырабатывающие ГАМК
ГАМК в основном синтезируется из глутамата с помощью фермента глутаматдекарбоксилазы (GAD) с пиридоксальфосфатом (активной формой витамина B6) в качестве кофактора. Этот процесс превращает глутамат (главный возбуждающий нейромедиатор) в ГАМК (главный тормозной нейромедиатор). [46] [47]
ГАМК также может быть синтезирован из путресцина [48] [49] диаминоксидазой и альдегиддегидрогеназой. [48]
Традиционно считалось, что экзогенная ГАМК не проникает через гематоэнцефалический барьер, [2] , однако более современные исследования [3] показывают, что это возможно, или что экзогенная ГАМК (т.е. в форме пищевых добавок) может оказывать ГАМКергическое действие на кишечную нервную систему, что, в свою очередь, стимулирует выработку эндогенной ГАМК. Непосредственное участие ГАМК в глутамат-глутаминовом цикле делает вопрос о том, может ли ГАМК проникать через гематоэнцефалический барьер, несколько вводящим в заблуждение, поскольку глутамат и глутамин могут свободно преодолевать барьер и превращаться в ГАМК в головном мозге.
Метаболизм
ФерментыГАМК трансаминазы катализируют превращение 4-аминобутановой кислоты (ГАМК) и 2-оксоглутарата (α-кетоглутарата) в янтарный полуальдегид и глутамат. Янтарный полуальдегид затем окисляется в янтарную кислоту янтарной полуальдегиддегидрогеназой и, как таковой, входит в цикл лимонной кислоты в качестве полезного источника энергии. [50]
Фармакология
Лекарства, которые действуют как аллостерические модуляторы рецепторов ГАМК (известные как аналоги ГАМК или препараты GABAergic ) или увеличивают доступное количество ГАМК, обычно обладают расслабляющим, успокаивающим и противосудорожным действием. [51] [52] Известно, что многие из перечисленных ниже веществ вызывают антероградную амнезию и ретроградную амнезию. [53]
Как правило, ГАМК не проникает через гематоэнцефалический барьер, [2] , хотя некоторые области мозга, не имеющие эффективного гематоэнцефалического барьера, такие как перивентрикулярное ядро, могут быть достигнуты путем такие препараты, как системно вводимая ГАМК. [54] По крайней мере, одно исследование предполагает, что пероральный прием ГАМК увеличивает количество гормона роста человека (HGH). [55] ГАМК, непосредственно вводимая в мозг, оказывает как стимулирующее, так и подавляющее действие на выработку гормона роста, в зависимости от физиологии человека. [54] Некоторые пролекарства ГАМК (например, пикамилон) были разработаны для проникновения через гематоэнцефалический барьер, а затем разделения на ГАМК и молекулу-носитель, попавшую в мозг. Пролекарства позволяют напрямую повышать уровень ГАМК во всех областях мозга, следуя схеме распределения пролекарства до метаболизма. [ необходима ссылка ]
ГАМК усиливала катаболизм серотонина в N -ацетилсеротонин (предшественник мелатонина) у крыс. [56] Таким образом, предполагается, что ГАМК участвует в синтезе мелатонина и, таким образом, может оказывать регулирующее влияние на сон и репродуктивные функции. [57]
Химия
Хотя с химической точки зрения ГАМК является аминокислотой (поскольку она имеет как первичный амин, так и функциональную группу карбоновой кислоты), она редко упоминается как таковая в профессиональном, научном или медицинском сообществе.Условно термин «аминокислота», когда он используется без уточнения, относится конкретно к альфа-аминокислоте. ГАМК не является альфа-аминокислотой, что означает, что аминогруппа не присоединена к альфа-углероду. Он также не входит в состав белков, как многие альфа-аминокислоты. [58]
ГАМКергические препараты
GABA A Лиганды рецептора показаны в следующей таблице. [nb 1]
| Деятельность в GABA A | Лиганд |
|---|---|
| Ортостерический агонист | мускимол, [59] ГАМК, [59] габоксадол (THIP), [59] изогувацин, прогабид, пиперидин-4-сульфоновая кислота (частичный агонист) |
| Положительные аллостерические модуляторы | барбитуратов, [60] бензодиазепинов, [61] нейроактивных стероидов, [62] ниацин / ниацинамид, [63] небензодиазепинов (i.е. z-препараты, например золпидем, эзопиклон) [ цитата необходима ] , этамидат, [64] этаквалон [ цитата требуется ] , спирт (этанол), [65] [66] [67] теанин [необходима ссылка ] , метаквалон, пропофол, стирипентол, [68] и анестетики [59] (включая летучие анестетики), глютетимид [требуется ссылка ] |
| Ортостерические (конкурентоспособные ) Антагонист | бикукуллин, [59] габазин, [69] туйон, [70] флумазенил [71] |
| Неконкурентный антагонист (например,грамм. блокировщик каналов) | пикротоксин [необходима ссылка ] , цикутоксин |
| Отрицательные аллостерические модуляторы | нейроактивные стероиды [ необходима ссылка ] (Прегненолон сульфат [ цитата требуется ] ), фуросемид, энантотоксин, аментофлавон |
Кроме того, каризопродол является усилителем активности ГАМК A [ необходима цитата ] .Ro15-4513 является восстановителем активности GABA A [требуется ссылка ] .
ГАМКергические пролекарства включают хлоралгидрат, который метаболизируется до трихлорэтанола, [72] , который затем действует через рецептор ГАМК A . [73]
Тюбетейка и валериана — растения, содержащие ГАМКергические вещества [ необходима цитата ] . Кроме того, растение кава содержит ГАМКергические соединения, включая каваин, дигидрокаваин, метистицин, дигидрометистицин и янгонин. a b Soltani N, Qiu H, Aleksic M, Glinka Y, Zhao F, Liu R, Li Y, Zhang N, Chakrabarti R, Ng T, Jin T, Zhang H , Лу В.Й., Фэн З.П., Прюдом Дж.Дж., Ван Кью (2011). «ГАМК оказывает защитное и регенерирующее действие на бета-клетки островков и обращает вспять диабет». Proc. Natl. Акад. Sci. США . 108 (28): 11692–7. Bibcode: 2011PNAS..10811692S. DOI: 10.1073 / pnas. Erdö SL, Wolff JR (февраль 1990 г.). a b c d e Chua HC, Chebib M (2017). «ГАМК-рецепторы и разнообразие в их структуре и фармакологии». GABAA рецепторы и разнообразие в их структуре и фармакологии . Успехи фармакологии. 79 . С. 1–34. DOI: 10.1016 / bs.apha.2017.03.003. ISBN 28104132 . (a) Herd MB, Белелли Д., Ламберт Дж. Дж. (Октябрь 2007 г.). «Нейростероидная модуляция синаптических и внесинаптических рецепторов ГАМК (А)». Фармакология и терапия . 116 (1): 20–34. DOI: 10.1016 / j.pharmthera.2007.03.007. PMID 17531325. ; (b) Hosie AM, Wilkins ME, da Silva HM, Smart TG (ноябрь 2006 г.). «Эндогенные нейростероиды регулируют рецепторы GABAA через два дискретных трансмембранных сайта». Природа . 444 (7118): 486–9. Bibcode: 2006 Натур.444..486Н. DOI: 10,1038 / природа05324. PMID 17108970. S2CID 4382394. ; (c) Agís-Balboa RC, Pinna G, Zhubi A, Maloku E, Veldic M, Costa E, Guidotti A (сентябрь 2006 г.). «Характеристика нейронов мозга, которые экспрессируют ферменты, опосредующие биосинтез нейростероидов». Труды Национальной академии наук Соединенных Штатов Америки . 103 (39): 14602–7. Bibcode: 2006PNAS..10314602A. DOI: 10.1073 / pnas.0606544103. PMC 1600006. PMID 167. ; (d) Akk G, Shu HJ, Wang C, Steinbach JH, Zorumski CF, Covey DF, Mennerick S (декабрь 2005 г.).«Нейростероидный доступ к рецептору ГАМКА». Журнал неврологии . 25 (50): 11605–13. DOI: 10.1523 / JNEUROSCI.4173-05.2005. PMC 6726021. PMID 16354918. ; (e) Белелли Д., Ламберт Дж. Дж. (июль 2005 г.). «Нейростероиды: эндогенные регуляторы рецептора ГАМК (А)». Обзоры природы. Неврология . 6 (7): 565–75. DOI: 10,1038 / номер 1703. PMID 156. S2CID 125 LP15353-3 Альфа-аминомасляная кислота LP15353-3 Альфа-амиобутират LP32035-5 Креатинин Габриэлла Занетти Также доступна: HTML-версия. Во время стрессовых недель в школе или на работе ваш мозг очень возбужден, и в ответ ваше тело обычно вырабатывает ГАМК, чтобы вернуть нервную систему в состояние покоя. Без ГАМК вы будете становиться все более беспокойными и тревожными, и у вас могут возникнуть судороги. ГАМК — это аминокислота, которая действует как нейротрансмиттер в центральной нервной системе. Естественная функция ГАМК — снижение активности нейронов, с которыми она связывается. Он подавляет нервную передачу в головном мозге, успокаивая нервную деятельность. Это может сделать человека более спокойным и дать ему или ей чувство благополучия. Нервная система состоит из отдельных нервных клеток, называемых нейронами.Они служат проводкой тела. Нервные сигналы передаются по длине нейрона в виде электрического импульса. Когда нервный импульс достигает конца нейрона, он может перепрыгнуть на следующую клетку с помощью химических посредников, называемых нейротрансмиттерами. В центральной нервной системе, состоящей из головного и спинного мозга, нейромедиаторы переходят от нейрона к нейрону. нейротрансмиттеров хранятся в конце каждого нейрона. Когда нейротрансмиттеры достигают соседнего нейрона, нейротрансмиттеры щелкают по специализированным рецепторным участкам так же, как ключ вставляется в замок.Когда к рецепторам присоединяется достаточное количество нейротрансмиттеров, нейрон «срабатывает», посылая электрический импульс по всей своей длине. ГАМК производится в клетках мозга из глутамата и действует как тормозящий нейротрансмиттер, то есть блокирует нервные импульсы. Глутамат действует как возбуждающий нейротрансмиттер и, будучи связанным с соседними клетками, побуждает их «стрелять» и посылать нервный импульс. ГАМК делает обратное и приказывает соседним клеткам не «стрелять», не посылать импульс. Без ГАМК нервные клетки срабатывают слишком часто и слишком легко. Тревожные расстройства, такие как панические атаки, судорожные расстройства и многие другие состояния, включая зависимость, головные боли, синдром Паркинсона и когнитивные нарушения, все связаны с низкой активностью ГАМК. ГАМК препятствует передаче нервных импульсов от одного нейрона к другому. Обладает успокаивающим или успокаивающим действием. Хороший пример, помогающий понять этот эффект, — кофеин.Кофеин подавляет высвобождение ГАМК. Чем меньше ГАМК, тем больше нервных передач. Подумайте, на что похоже слишком много кофе: это ощущение глутамата без достаточного количества ГАМК. Причина, по которой кофеин делает это, заключается в том, что другие молекулы могут связываться с нейроном рядом с сайтом связывания ГАМК и влиять на эффект ГАМК. 1883 : Гамма-аминомасляная кислота была впервые синтезирована, сначала была известна только как продукт метаболизма растений и микробов. 1950 : Было обнаружено, что ГАМК является неотъемлемой частью центральной нервной системы млекопитающих. Многие люди принимают ГАМК в качестве добавки для улучшения настроения и снятия беспокойства. Однако недостаточно исследований, чтобы выявить побочные эффекты и риски добавок ГАМК. Есть мало свидетельств того, что он что-то делает. В последнее время считается, что ГАМК не преодолевает гематоэнцефалический барьер.Гематоэнцефалический барьер — это биологический брандмауэр между общим кровообращением тела и кровообращением, которое снабжает мозг. Он предотвращает попадание в мозг многих химических веществ и лекарств, циркулирующих в крови. Это означало бы, что ГАМК не может проникать из тела в мозг; следовательно, люди сомневаются, сработает ли ГАМК. Тем не менее, люди все еще могут принимать ГАМК, потому что она может действовать как плацебо. Кроме того, это может иметь некоторый эффект, о котором еще не сообщалось, поэтому необходимо провести дополнительные исследования. Вернуться на страницу «Молекула месяца». NCCCC (O) = O InChI = 1S / C4H9NO2 / c5-3-1-2-4 (6) 7 / h2-3, 5h3, (H, 6,7) Путь воздействия: Источник: Биологическое местоположение: Естественный процесс: Косвенная биологическая роль: Биологическая роль Накопление пролина (Pro) в растениях, подвергшихся биотическому / абиотическому стрессу, является хорошо задокументированным и законсервированным ответом у большинства растительных видов.Стрессовые условия вызывают перепроизводство активных форм кислорода, что может привести к повреждению клеток. Анализы in vitro показали, что инактивации фермента гидроксильными радикалами ( · OH) можно избежать в присутствии Pro, предполагая, что эта аминокислота может действовать как поглотитель · OH. Мы применили функциональную теорию плотности в сочетании с моделью поляризуемого континуума, чтобы выяснить, как Pro взаимодействует с · OH. В этой работе мы предполагаем, что Pro благоприятно реагирует с · OH за счет H – отщепления на аминогруппе.Эта реакция вызывает спонтанное декарбоксилирование Pro, приводящее к образованию пирролидин-1-ила. В свою очередь, пирролидин-1-ил легко превращается в Δ 1 -пирролин, субстрат фермента Δ 1 -пирролиндегидрогеназы, который продуцирует γ-аминомасляную кислоту (ГАМК). ГАМК и Pro часто накапливаются в стрессовых растениях, и этим молекулам отведено несколько защитных функций. Таким образом, мы представляем альтернативный неферментативный способ синтеза ГАМК в условиях окислительного стресса.Наконец, эта работа проливает свет на новую полезную роль накопления Pro в поддержании фотосинтетической активности. Образец цитирования: Signorelli S, Dans PD, Coitiño EL, Borsani O, Monza J (2015) Связывание пролина и гамма-аминомасляной кислоты в стрессовых растениях посредством неферментативных реакций. PLoS ONE 10 (3):
e0115349.
https://doi.org/10.1371/journal.pone.0115349 Академический редактор: Сергей Шабала, Университет Тасмании, АВСТРАЛИЯ Поступила: 4 августа 2014 г .; Одобрена: 22 ноября 2014 г .; Опубликован: 16 марта 2015 г. Авторские права: © 2015 Signorelli et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника Доступность данных: Все соответствующие данные находятся в пределах бумага. Финансирование: Fondo Clemente Estable 6662 Agencia Nacional de Investigación e Innovación (FCE_6662_ANII), Programa de desarrollo de las ciencias básicas (PEDECIBA), http: // www.pedeciba.edu.uy/indice.php, SS. Comisión Académica de Posgrado (CAP), http://www.posgrados.udelar.edu.uy/, SS. Sistema Nacional de Investigadores (SNI): http://www.sni.org.uy/, SS PDD ELC OB JM. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи. Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует. Когда растения подвергаются биотическому / абиотическому стрессу, повреждение клеточных компонентов (белков, липидов, углеводов и ДНК) увеличивается в результате перепроизводства активных форм кислорода (АФК), таких как перекись водорода (H 2). O 2 ), синглетный кислород ( 1 O 2 ), супероксид-анион (O 2 · — ) и гидроксильные радикалы ( · OH) [1].Последние являются наиболее реактивными формами среди АФК и могут образовываться in vivo либо реакцией Фентона, в цикле Габера-Вейсса, либо путем гомолиза H 2 O 2 под УФ-излучением, которому растения подвергаются в высокой степени. [2,3]. Несмотря на короткий срок службы, производство · OH было обнаружено в интактных растениях с использованием методов ЭПР [4,5]. Процессы отвода водорода, присоединения и переноса электрона являются наиболее распространенными каналами реакции для · OH, ведущими к новым радикалам или молекулам с закрытой оболочкой с более низкой реакционной способностью [6]. Клеточная защита от АФК может выиграть от ферментативных или неферментативных антиоксидантных процессов. Считается, что пролин (Pro) участвует в неферментативной антиоксидантной защите растений [7,8]. Накопление Pro в стрессовых растениях до уровня в 100 раз превышающего нормальный уровень является хорошо известным фактом уже более 40 лет [9]. В этом состоянии Pro может достигать концентрации в цитозоле от 120 до 230 мМ (см. Ссылку [10] и ссылки в ней). Сообщалось, что в условиях засухи и высокой солености, УФ / видимого излучения, окислительного стресса, присутствия тяжелых металлов или в ответ на различные виды биотических стрессов [11] накопление Pro за счет синтеза de novo является особенностью являются общими для самых разных организмов, включая бактерии, грибы и растения [12]. В начале 1989 года Sminorff и Cumbes показали, что инактивации фермента · OH можно избежать in vitro за счет присутствия Pro, предполагая, что эта молекула может действовать как поглотитель · OH [13]. Позже было также высказано предположение, что Pro может защищать растения от окисления 1 O 2 [14,15]. Косвенные доказательства такой защитной роли появились при сравнении окислительного повреждения генетически модифицированных растений в условиях стресса, когда трансгены использовались для контроля продукции Pro.В условиях засоления трансгенные растения с повышенной продукцией и накоплением Pro менее подвержены окислительному повреждению [16], в то время как растения, генетически неспособные продуцировать Pro, проявляют значительно более низкую устойчивость к стрессу [17]. Недавно были получены прямые доказательства того, что Pro не тушит синглетный кислород ( 1 O 2 ), как считалось в течение нескольких лет, и был сделан вывод о том, что защитная роль Pro против окислительного повреждения, наблюдаемая у некоторых растений, может быть связана с активность акцептора · OH [18]. Пионерская работа Amici и др. . предположил, что · OH реагирует с Pro, образуя 5-гидроксипролин (5-Hyp), и с 5-Hyp с образованием глутаминовой кислоты [19,20]. В недавней работе мы исследовали реакции, включающие атаку · OH на различные атомы C Pro, чтобы оценить образование 5-Hyp, используя теоретический подход, и предположили, что образование 5-Hyp маловероятно [ 21]. Мы предсказали, что реакция всегда должна происходить на карбоксилатной поверхности ( s-грань ) Pro с образованием либо 3,4-Δ-Pro, либо пирролин-5-карбоксилата (P5C) [21]. В этом сценарии настоящая работа направлена на оценку атаки • OH на атом N, чтобы оценить конкурентоспособность этого пути в отличие от описанных для атомов C [21]. Мы обнаружили, что • ОН-атака на атом N Pro конкурирует с наиболее предпочтительными атаками по атомам C и может привести к образованию γ-аминомасляной кислоты (ГАМК), которая, как сообщалось, накапливается в ответ на абиотический и биотический стресс [22]. Мы исследовали абстракции H-атомов с помощью • OH из 4-эндо и 5-эндо конформера цвиттерионной формы Pro в водном растворе.В конформации 4-эндо атомы C2, C3, C5 и N определяют почти плоскость, в то время как в конформере 5-эндо атомы C2, C3, C4 и N почти совмещены. Как показано на рис. 1, первичная стадия реакции • OH с Pro (стадия 1) характеризуется отщеплением H-атома за счет радикальной атаки на атом N на обеих сторонах пирролидинового кольца (т. Е. От сторона карбоксилатной группы или с противоположной стороны, соответственно обозначенные как s / o грани). На стадии 2 высвобождение карбоксильной группы в виде CO 2 приводит к образованию радикала пирролидин-1-ила (Pyr • ).Pyr • может снова реагировать с • OH — или с другими молекулами, способными отрывать атом водорода от группы NH — с образованием дельта-1-пирролина (Δ 1 -Pyr) и воды. В качестве альтернативы Pyr • может реагировать с молекулой H 2 O 2 и образовывать две молекулы с замкнутой оболочкой: пирролидин (Pyr) и кислотную форму супероксида. Структуры каждого стабильного вида (реагенты, продукты, предреакционные промежуточные комплексы (IC) и переходные состояния (TS)) были полностью оптимизированы в водном растворе на уровне (U) M06 / 6-31G (d, p). [23] в сочетании с моделью поляризуемого континуума IEF-PCM [24–26].Расчеты проводились без наложения ограничений по симметрии и с использованием полостей для растворенных веществ, адаптированных к форме молекулы и построенных с радиусами Бонди [27]. Ультратонкая обрезанная сетка, имеющая 99 радиальных оболочек и 590 угловых точек на оболочку, использовалась для численного интегрирования во всех расчетах M06, чтобы уменьшить возможные ошибки в вычислении энергий и барьеров [28]. Ожидаемые значения оператора спина S 2 были проверены на то, чтобы быть ниже 0,76 и 2,01 для дублетных и триплетных видов с открытой оболочкой, соответственно, чтобы минимизировать спиновое загрязнение.Природа каждой стационарной точки была тщательно проверена путем проверки собственных значений аналитического гессиана в водном растворе. Термохимия была оценена при 298,15 К на основе стандартной обработки для оценки тепловых вкладов (жесткий ротор, гармонические колебания без масштабного коэффициента и т. Д.), Реализованных в программе Gaussian09 [29]. Неэлектростатические вклады (кавитация, дисперсия и отталкивание) [30,31] в свободную энергию растворителя также оценивались при 298,15 К. Координата реакции каждого переходного состояния (TS) визуально проверялась путем анимации собственного вектора, связанного с воображаемым частота.Пути реакции IRC с минимальной энергией [32] к реагентам или продуктам были созданы с помощью алгоритма HPC [33], включающего 45 шагов для каждой стороны с размером шага 2 Бора / а.е.м. 1/2 . Таким образом, все репрезентативные структуры, полученные с каждой стороны пути реакции, использовали в качестве отправной точки для оптимизации структуры соответствующих промежуточных комплексов (IC). Анализ населенности Natural Bond Orbital (NBO) [34,35] был проведен для оценки характеристик связывания между C 1 -C 2 атомами реагентов, TS, I C и продуктов.Ортогональный набор локализованных орбиталей, полученный в результате этого анализа, также использовался для вычисления индекса связи Виберга для процесса декарбоксилирования [36,37]. Одноточечные расчеты на уровне замороженного ядра MP2 (с использованием базиса 6-31G (d, p)) были выполнены для анализа спиновой и электронной плотности всех охарактеризованных структур. Подход «Атомы в молекулах» (AIM) [38] был использован для отслеживания процесса спонтанного декарбоксилирования, нахождения критических точек связи между атомами в плоскости карбоксильной группы в Pro. Все расчеты проводились с использованием Gaussian09, rev. A.1 или B.1 [29], в то время как молекулярные чертежи были построены с использованием Gaussview 5 или VMD 1.9.1 [39]. Анализ AIM был проведен с помощью программы AIM-UC [40]. Были оценены четыре различных пути реакции для ОН-атаки • на атом N Pro цвиттер-иона.Эти пути включают, как для 4-эндо, так и для 5-эндо конформеров Pro, атаку со стороны карбоксильной группы ( s-face ) или ее противоположной стороны ( o-face ). Соответствующие энергетические барьеры (таблица 1) оценивают конкурентоспособность этих путей в отличие от недавно описанных нами [21] для атомов углерода. Самые низкие барьеры с точки зрения свободной энергии для каждого сайта атаки представлены на рис. 2. Примечательно, что после того, как H-отрыв происходит на атоме N 5-эндо Pro, карбоксильная группа дестабилизируется и диссоциирует от кольца (рис. .3). Барьер этих реакций невелик, снижается, когда атака происходит от s-грани . Более того, барьеры, связанные с абстракцией s-face над N, ниже на ∼ на 1 ккал / моль, чем те, которые ранее были описаны для наиболее благоприятных водородных абстракций на атомах C3 / C4 (Таблица 1), причем все они близки к диффузионно-контролируемый предел. Установлено, что предреакционные комплексы, полученные на концах собственного пути координат реакции, являются стабильными с точки зрения энтальпии (6–7 и 2–3 ккал моль –1 для с — и o — лицо соответственно).Однако с точки зрения свободной энергии эти предреакционные комплексы становятся нестабильными из-за неблагоприятных энтропийных вкладов от 2 до 6 ккал / моль -1 . Эти результаты в конечном итоге исключают возможность этих промежуточных комплексов действовать как кинетические ловушки, как это было обнаружено для другого H-отщепления с помощью • OH из дипептида лейцина [41]. Свободная энергия (в ккал-моль -1 ) относительно реагентов была рассчитана в водном растворе при 298.15 К. Энергии для H-отрыва на атомах C взяты из [21]. https://doi.org/10.1371/journal.pone.0115349.g002 Рис. 3. Структуры, участвующие в H-абстракции посредством • OH от атома N 4-эндо и 5-эндо-Pro. Обратите внимание, что IC R и IC P являются промежуточными комплексами для реагентов и продуктов соответственно. https://doi.org/10.1371/journal.pone.0115349.g003 Принимая во внимание, что 4-эндо и 5-эндо формы Pro существуют в растворе в равных пропорциях [42], и что более низкие энергетические барьеры были обнаружены для этих видов, когда атака происходит на s -грани, мы сфокусировались на пути реакции s-face для обоих видов, чтобы детально оценить структурные изменения, которые вызывают высвобождение карбоксильной группы из пирролидинового кольца.На рис. 3 показана структура реагентов, промежуточных комплексов, переходного состояния и продуктов этой реакции. Таблица 2 показывает основные структурные параметры видов, участвовавших в Шаге 1 на s-грани . Как только реакция отрыва достигает TS, расстояние между C α и карбоксильной группой (C α -COO —) увеличивается с 1,529 (TS) до 1,795 Å (продукт) в 4-эндо конформации и от 1,528 (TS) до 2,711 Å (продукт) в случае 5-эндо-конформера.Это свидетельствует об ослаблении связи C 1 -C 2 , в то время как сокращение расстояния C 2 -N в обеих конформациях характерно для двойной связи (Таблица 2). С другой стороны, угол O 1 -C 1 -O 2 карбоксильной группы становится менее острым (особенно в конформере 5-эндо), а расстояния C = O этих групп становятся короче, напоминая к молекуле CO 2 . Когда отрыв происходил от 5-endo Pro, на стороне продукта не было обнаружено минимумов до тех пор, пока не произойдет полная диссоциация карбоксильной группы (конечное расстояние C 1 -C 2 равно 2.711 Å, а значение угла O 1 -C 1 -O 2 составляет 173,4 °). Эти результаты предоставляют убедительные доказательства того, что атака • OH на N-сайт Pro может запускать его декарбоксилирование, давая образование пирролидин-1-ила (Pyr • ). Про декарбоксилирование также было подтверждено экспериментально in vitro в присутствии окислительных частиц, таких как перманганат [43] и медь [44]. Таким же образом Bonifacic et . и .[45] изучили реакцию между • OH и Gly, обнаружив, что атаки • OH происходят исключительно на группу NH 2 , что приводит к образованию HN • -CH 2 -CO 2 — и + H 2 N • -CH 2 -CO 2 — . Последние разлагаются (в наносекундном масштабе времени) до CO 2 и H 2 N- • CH 2 [45]. Для дальнейшей характеристики шагов 1 и 2 (см. Рис.1), H-абстракция и апостериорное декарбоксилирование были проанализированы с использованием индексов Виберга, спиновых плотностей (разницы между альфа- и бета-плотностями) и подхода AIM. В таблице 3 мы представляем индекс Виберга для связи C 1 -C 2 как в 4-эндо, так и в 5-эндо конформации вдоль структур этапа 1. Как показано, результаты согласуются с геометрическими параметрами. (Таблица 2). Связь C 1 -C 2 немного усиливается до TS, а затем быстро ослабевает, почти полностью разрываясь в продукте 5-эндо.В обоих конформерах мы исчерпывающе ищем TS в процессе декарбоксилирования, но все попытки привели к быстрому высвобождению группы CO 2 , что позволяет предположить, что стадия 2 является безбарьерной. Спиновая плотность — хороший квантовый дескриптор для изучения неспаренного электрона на пути реакции. Как показано на рис. 4, неспаренная плотность перемещается от делокализации по атомам кислорода (см. IC R и TS на рис. 4) до локализации по атомам C 2 / Cα и N в продуктах (см. IC P и продукты).Во время H-отрыва спиновая плотность перемещается от атомов кислорода (в основном от • OH) через атом N (образуя катионный радикал) к атому C 2 / Cα, что приводит к декарбоксилированию Pro и усиление связи Cα-N (см. Табл. 2). Конечным результатом является спонтанное образование Pyr • и диоксида углерода (ясно в случае 5-эндо-конформера) в безбарьерной реакции, для которой не требуется энергия активации и невозможно определить TS.Стоит отметить, что декарбоксилирование происходит по-разному в 4-эндо и 5-эндо формах. Декарбоксилирование, по-видимому, хорошо определено в 5-эндо. В реакции 4-эндо спиновая плотность не показывает значительных различий между IC P и продуктом (рис. 4), хотя, как упоминалось ранее, расстояние C 1 -C 2 между ними немного увеличивается. два стабильных вида (см. таблицу 2), что позволяет предположить, что мы обнаружили стабильный комплекс на стороне продукта H-абстракции перед декарбоксилированием (в соответствии с индексами Wiberg, приведенными в таблице 3).В последнем случае анализ AIM также показывает наличие критической точки связи (bcp), предполагая, что все еще существует ковалентная связь, соединяющая карбоксильную группу с пирролидиновым кольцом. Этот bcp отсутствует в продукте 5-эндо реакции, где декарбоксилирование очевидно. В 4-эндо форме молекулы Pyr • и CO 2 , по-видимому, захвачены в локальный минимум, образуя стабильный комплекс, что может частично объяснить низкую скорость разложения, наблюдаемую экспериментально [45]. Рис. 4. Плотность спина и анализ AIM. Для сравнения одна и та же изоповерхность со значением 0,003 | e | / Å 3 , представляющая разницу между плотностями альфа- и бета-электронов, была изображена фиолетовым или зеленым цветом для форм 4-эндо и 5-эндо соответственно. . Для продуктов и промежуточных частиц на стороне продукта (IC P ) наличие критических точек связи (bcp) было подтверждено с помощью AIM. БЦП, соединяющий карбоксильную группу с пирролидиновым кольцом, изображен фиолетовым или зеленым цветом, тогда как остальные БЦП показаны серым цветом.Ядерные критические точки (расположенные в положении ядер), траектории бассейнов и градиентное поле изображены сплошными или пунктирными черными линиями. Связующие пути, определяемые выбранной двухмерной проекцией (плоскостью), показаны красным. https://doi.org/10.1371/journal.pone.0115349.g004 Pyr • может реагировать с различными молекулами с образованием молекул с замкнутой оболочкой. Здесь мы предлагаем два различных механизма: один включает другой H-отрыв с помощью • OH от атома N пирролидинового кольца; а другой связан с отрывом водорода Pyr • от пероксида водорода (см. рис.1). В обоих случаях реакция приводит к образованию Δ 1 -пирролин (Δ 1 -Pyr). Обратите внимание, что после декарбоксилирования невозможно определить s- и o-грань, и по этой причине существует только одна возможная конформация в двух реакциях, исследованных на этапе 3 (рис. 5). Как и ожидалось (на этапе 3) реакция с • OH имеет более низкий энергетический барьер, а также более экзотермична, чем реакция с H 2 O 2 (см. Таблицу 4).Оба предложенных механизма вероятны, и хотя этап 3a необратим, этап 3b может происходить в обоих направлениях. И снова промежуточные комплексы более стабильны с точки зрения энтальпий, но уже не с учетом энтропийных вкладов. В растениях Δ 1 -Pyr, полученный на стадии 3а, является субстратом пирролиндегидрогеназы (PYRR-DH), которая превращает его в γ-аминомасляную кислоту (ГАМК) [46]. Доказано, что ГАМК накапливается в стрессовых условиях [47].По этой причине ГАМК было отведено несколько защитных ролей, таких как вклад в баланс C: N, регуляция цитозольного pH, защита от окислительного стресса, защита от насекомых, осморегуляция и передача клеточных сигналов [47] . Большинство этих защитных функций также приписывается Pro [11]. Основной путь синтеза ГАМК — из глутаминовой кислоты с помощью фермента глутаматдекарбоксилазы [22], в то время как вторичный путь включает полиамины и образование Δ 1 -Pyr [46, 48].Например, мутант CMSII Nicotiana sylvestris обладал пониженной активностью глутаматдекарбоксилазы (GAD), но содержание ГАМК, тем не менее, увеличивалось более чем в два раза по сравнению с базовым уровнем при обработке NaCl [49], что позволяет предположить, что накопление ГАМК могло опосредовано окислением полиаминов. Этот путь требует активности пирролин (γ-аминобутиральдегид) дегидрогеназы, фермента, не продуцируемого в стрессовых условиях [48], что означает, что образование ГАМК должно опосредоваться накоплением предшественников ГАМК.Здесь мы предлагаем альтернативный неферментативный путь (показан синим цветом на рис.6), в котором Pro может вносить вклад в образование предшественника ГАМК Δ 1 -Pyr, давая возможное объяснение связи одновременного накопления Pro и GABA которые возникают в стрессовых условиях. Недавно мы продемонстрировали, что пластидный мутант глутаминсинтетазы Lotus japonicus с неизмененным содержанием глутамата, но более низким содержанием пролина, имел меньшее накопление ГАМК в ответ на осмотический стресс по сравнению с диким животным [50]. Рис. 6. Предлагаемый путь соединения Pro и GABA и его возможное влияние на основные метаболические процессы. Синие линии представляют неферментативные реакции, предложенные для соединения Pro и Δ 1 -Pyr посредством очистки • OH. В неферментативных реакциях захватываются два • OH, а выделившийся CO 2 может войти в цикл C3. Красные и зеленые линии представляют собой катаболические и анаболические пути соответственно. SSA означает янтарный полуальдегид, а AT означает аминотрансферазы. https://doi.org/10.1371/journal.pone.0115349.g006 В хлоропластах совместная локализация Pro, • OH и H 2 O 2 необходима для протекания реакций, описанных в шагах 1 и 3. В этих органеллах утечка электронов O 2 является основным источником O 2 • — [51], что приводит к образованию H 2 O 2 за счет активности СОД в реакции, ограниченной диффузией. • OH можно получить из H 2 O 2 реакцией Фентона или гомолитическим расщеплением под воздействием УФ-излучения. Кроме того, Pro достигает концентрации не менее 160 мМ [52] (даже в нестрессовых условиях) в этих органеллах, становясь более распространенным, чем любой обычный антиоксидант (например: 0,8–2,4 мМ для глутатиона; 12–25 мМ для аскорбата) [ 53]. Помимо продукции H 2 O 2 в хлоропластах, H 2 O 2 также генерируется фотодыханием в пероксисомах [54].Другими источниками H 2 O 2 являются медьсодержащие оксидазы и пероксидазы, которые являются основными ответственными за продукцию H 2 O 2 во время окислительного всплеска в стрессовых условиях у растений [54]. Поскольку H 2 O 2 имеет длительный срок жизни и может диффундировать через органеллы [55], можно предположить, что он доступен во всей клетке с образованием • OH или реагирует с Pyr • . Из-за основной совместной локализации ROS и Pro в хлоропластах CO 2 , высвобождаемый из Pro, накапливается преимущественно в этих органеллах.Там фотосинтетический аппарат может использовать CO 2 путем фиксации через цикл C3 (см. Рис. 6). Несмотря на то, что это гипотеза, которая потребует дальнейших экспериментальных подтверждений, такое локализованное производство CO 2 может быть преимуществом для растений в стрессовых условиях (например, засухе). В таких условиях устьица близки, чтобы избежать потери воды, ограничивая поглощение CO 2 , что приводит к снижению фиксации углерода и накоплению восстанавливающей способности (НАДФН).Все эти комбинированные функции, скорее всего, будут производить АФК за счет утечки электронов. Следовательно, есть основания полагать, что CO 2 , образующийся во время Pro-реакций, поможет снизить накопление NADPH и образование ROS. Pro в зеленых тканях был предложен для регенерации NADP + , помогая поддерживать адекватное соотношение NADP + / NADPH внутри клеток [56], и что Pro мог перемещаться в корни для катаболизации [56] .В дополнение к потреблению НАДФН, производимому во время синтеза Pro, наша работа предполагает, что Pro также может помочь снизить соотношение НАДФ + / НАДФН, высвобождая CO 2 , который войдет в цикл C3 и потребит НАДФН. С другой стороны, известно, что трансаминазы превращают ГАМК в янтарный полуальдегид (SSA), который затем превращается в сукцинат янтарными полуальдегиддегидрогеназами (рис. 6) [57,58]. Эти реакции, известные как шунты ГАМК, производят субстраты митохондриальной дыхательной цепи (сукцинат и НАДН), которые в конечном итоге генерируют АТФ [47].Шунты ГАМК влияют на окислительно-восстановительный баланс в клетке, потому что сукцинат входит в цикл Кребса, минуя три сайта производства НАДН, тем самым снижая соотношение НАДН / НАД + [49,59,60]. Это снижение отношения НАДН / НАД + активирует ключевые ферменты цикла Кребса [61]. Кроме того, известно, что ферменты сукцинил-КоА-лигаза и α-кетоглутаратдегидрогеназа разлагаются при окислительном стрессе, ограничивая выработку АТФ в цикле Кребса [62]. Шунты ГАМК обеспечивают производство АТФ даже в стрессовых условиях, поэтому шунты ГАМК считались защитой от окислительного стресса [47]. В целом, мы считаем, что неферментативная связь между Pro и GABA, представленная в этой работе, является разумной альтернативой катаболизму Pro с получением восстанавливающей способности, АТФ, углерода и азота, даже в стрессовых условиях, когда катаболические ферменты Pro инактивированы. Эти полезные аспекты Pro и защитная роль против • OH соответствуют многофункциональной роли, которая была принята для Pro. Отрыв водорода от атома N Pro радикалом • OH вызывает декарбоксилирование Pro и приводит к образованию Pyr • .Этот механизм реакции, предложенный здесь впервые, является конкурентным по сравнению с описанным ранее для атомов углерода [21]. Эти теоретические результаты согласуются с экспериментальными данными о декарбоксилировании аминокислот при атаках • OH. Δ 1 -Pyr, продуцируемый H-абстракцией из Pyr • , будет способствовать выработке ГАМК при катализе PYRR-DH, важной молекулы для растений, которая также накапливается в стрессовых условиях. Таким образом, мы предложили альтернативный способ синтеза ГАМК с помощью неферментативной реакции, характерной для растений в условиях окислительного стресса.Наконец, эта работа выдвигает на первый план новую полезную характеристику накопления Pro, как вклад в поддержание фотосинтетической активности у стрессированных растений. Авторы выражают признательность доктору Федерике Баттистини за критическое прочтение рукописи. Задумал и спроектировал эксперименты: SS PDD ELC. Проведены эксперименты: СС ПДД. Проанализированы данные: SS PDD ELC. Предоставленные реагенты / материалы / инструменты анализа: SS PDD ELC OB JM.Написал статью: SS PDD ELC OB JM. Способствовал биологической подоплеке проблемы: SS OB JM.. ; (f) Pinna G, Costa E, Guidotti A (июнь 2006 г.). «Флуоксетин и норфлуоксетин стереоспецифически и избирательно увеличивают содержание нейростероидов в мозге в дозах, которые не действуют на обратный захват 5-HT». Психофармакология . 186 (3): 362–72. DOI: 10.1007 / s00213-005-0213-2. PMID 16432684. S2CID 7799814. ; (ж) Дубровский Б.О. (февраль 2005 г.). «Стероиды, нейроактивные стероиды и нейростероиды в психопатологии». Успехи нейропсихофармакологии и биологической психиатрии . 29 (2): 169–92. DOI: 10.1016 / j.pnpbp.2004.11.001. PMID 156 . S2CID 361 . ; (h) Mellon SH, Griffin LD (2002). «Нейростероиды: биохимия и клиническое значение». Тенденции в эндокринологии и метаболизме . 13 (1): 35–43. DOI: 10.1016 / S1043-2760 (01) 00503-3. PMID 11750861. S2CID 11605131. ; (i) Puia G, Santi MR, Vicini S, Pritchett DB, Purdy RH, Paul SM, Seeburg PH, Costa E (май 1990 г.). «Нейростероиды действуют на рекомбинантные человеческие рецепторы GABAA». Нейрон . 4 (5): 759–65. DOI: 10.1016 / 0896-6273 (90) -Q. PMID 2160838. S2CID 12626366. ; (j) Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM (май 1986).«Метаболиты стероидных гормонов являются барбитуратоподобными модуляторами рецептора ГАМК». Наука . 232 (4753): 1004–7. Bibcode: 1986Sci … 232.1004D. DOI: 10.1126 / science.2422758. PMID 2422758. ; (k) Редди Д.С., Рогавски М.А. (2012). «Нейростероиды — эндогенные регуляторы предрасположенности к приступам и их роль в лечении эпилепсии». В Noebels JL, Avoli M, Rogawski MA и др. (ред.). Основные механизмы эпилепсии Джаспера [Интернет]. 4-е издание. Бетесда (Мэриленд): Национальный центр биотехнологической информации (США) . Roberts MR (сентябрь 2007 г.). «Действует ли ГАМК как сигнал в растениях?: Подсказки молекулярных исследований». Сигнальное поведение предприятия . 2 (5): 408–9. DOI: 10.4161 / psb.2.5.4335. PMC 2634229. PMID 1 16. Внешние ссылки
Эта страница последний раз была отредактирована 6 июля 2021 в 16:21 Ионотропный GABA A GABA A -ρ Metabotropic LOINC 28609-6 — Альфа-аминобутират / креатинин [соотношение] в суточной моче
Описание деталей
Альфа-аминомасляная кислота (AABA) представляет собой изомер аминокислоты аминомасляной кислоты с химической формулой C4 H9 NO2.Есть также гамма-аминомасляная кислота (ГАМК) и бета-аминомасляная кислота. Это ключевой промежуточный продукт в биосинтезе офтальмологической кислоты или офтальмата, который был впервые выделен из хрусталика теленка. Текст об авторских правах доступен по лицензии Creative Commons Attribution / Share-Alike. Подробнее см. Http://creativecommons.org/licenses/by-sa/3.0/. Источник: Википедия, Альфа-аминобутират (Википедия)
Альфа-аминомасляная кислота (AABA), также известная как альфа-амино-N-масляная кислота (A-ANB), представляет собой промежуточное соединение, образующееся при катаболизме метионина и треонина.Повышение AABA происходит вторично по отношению к повышению уровня метионина или треонина. AABA превращается в пропионовую кислоту через альфа-кетомасляную кислоту в присутствии достаточного количества тиамина, витамина B2 (в виде FAD), витамина B3 (в виде NAD), липоевой кислоты и магния. Недостаток любого из них или витамина B6 может вызвать увеличение AABA. Повышенный или пониженный уровень аминокислоты может указывать на врожденный дефект фермента. Текст об авторских правах доступен по лицензии Creative Commons Attribution / Share-Alike.Подробнее см. Http://creativecommons.org/licenses/by-sa/3.0/. Источник: Институт Регенстриф, Альфа-амиобутират (Википедия)
Креатинин или ангидрид креатина — продукт распада креатинфосфата в мышцах. Потеря молекулы воды из креатина приводит к образованию креатинина. Он переносится в почки с плазмой крови, после чего выводится путем клубочковой фильтрации и частичной канальцевой экскреции.Креатинин обычно вырабатывается с довольно постоянной скоростью, и измерение его уровня в сыворотке — простой тест. Повышение уровня креатинина в крови наблюдается только при заметном повреждении функционирующих нефронов; поэтому этот тест не подходит для выявления ранней болезни почек. Креатин и креатинин метаболизируются в почках, мышцах, печени и поджелудочной железе. Copyright © 2005-2009 Genome Alberta (Ссылка на оригинальную публикацию: Wishart DS, Knox C, Guo AC и др. HMDB: база знаний о метаболоме человека.Nucleic Acids Res. 2009 37 (выпуск базы данных): D603-610.) Источник: База данных метаболизма человека, креатинин . Отображение соединения 4-аминомасляной кислоты (FDB008937)
Запись информации Версия 1.0 Дата создания 08.04.2010 22:08:19 UTC 9019 2020-09-17 15:38:19 UTC Первичный ID FDB008937 Вторичные номера доступа Недоступно Химическая информация Химическая информация -Аминомасляная кислота Описание гамма-аминомасляная кислота, также известная как ГАМК или g-амино-бутаноат, принадлежит к классу органических соединений, известных как гамма-аминокислоты и производные.Это аминокислоты, имеющие группу (-Nh3), присоединенную к гамма-атому углерода. гамма-аминомасляная кислота — очень гидрофобная молекула, практически нерастворимая (в воде) и относительно нейтральная. гамма-аминомасляная кислота существует у всех живых существ, от бактерий до людей. Вне человеческого тела гамма-аминомасляная кислота была обнаружена, но не определена количественно в нескольких различных продуктах, таких как лисий виноград, капуста бычьего сердца, садовый лук (разновидность), желтые кабачки и китайская горчица. Это может сделать гамма-аминомасляную кислоту потенциальным биомаркером для употребления этих продуктов.гамма-аминомасляная кислота — потенциально токсичное соединение. Гамма-аминокислота, представляющая собой бутановую кислоту с амино-заместителем, расположенным у C-4. Номер CAS 56-12-2 Структура Синонимы Синоним 4-аминомасляная кислота ChEBI 4Abu ChEBI GABA ChEBI AMIC ChEBI AMIC GAMEB-9018 ChEBI-9018-GAME-9018-GAME4 9018-AMA-9018-GAME4 9018-AMIC-9018-GAME4-9018-GAME4-9018-AMIC-9018-GAME-9018-GAME-4 масляная кислота ChEBI гамма-аминобутанова кислота ChEBI гамма-Aminobuttersaeure ChEBI Омега-аминомасляная кислота ChEBI Piperidic кислота ChEBI Пиперидиновая кислота ChEBI 4-Аминобутират Kegg Gammalon Kegg 4-аминобутаноат Генератор г-АМИНО-бутаноат Генератор AMINO-бутаноат Генератор Генератор Генератор 8 but5 гамма-амино-бутаноат Генератор Г-амино-бутаноат Генератор Г-амино-бутановая кислота Генератор 9018 г-Амино-N-9018 Генератор 9018 9018 9018 Генератор амино-N-9018 г-амино-N-масляная кислота Генератор гамма-амино-N-бутират Генератор Γ-амино-N-бутират Генератор амино масляная кислота Генератор Г-аминобутаноат Генератор г-Аминобутановая кислота Генератор гамма-аминобутаноат Генератор Г-аминобутаноат Генератор Г-аминобутановая кислота Генератор Генератор Генератор g-A-A Омега-аминобутират Генератор Пиперидат Генератор Пиперидинат Генератор g-Aminobutyrate 9018 9018 Генератор 9018 9018 9018 9018 9018 9018 9018 9018 Генератор 9018 9018 9018 9018 9018 9018 9018 Генератор 9018 9018 9018 9018 9018 9018 9018 Генератор 9018 9018 9018 9018 9018 9018 9018 Генератор 9018 9018 9018 гамма-аминомасляная кислота Генератор Г-аминомасляная кислота Генератор Г-аминомасляная кислота Генератор 3-карбоксипропиламин 9018DB 9018DB 9018DB 9018DB 0185 Gaballon HMDB Gamarex HMDB гамма-аминобутират HMDB гамма-аминомасляная кислота HMDB Gammalone HMDB Gammar HMDB Gammasol HMDB Mielogen HMDB Mielomade HMDB W-аминобутират HMDB W-аминомасляная кислота HMDB гамма -Аминомасляная кислота, кальциевая соль (2: 1) HMDB гамма-аминомасляная кислота, гидрохлорид HMDB гамма-аминомасляная кислота, цинковая соль (2: 1) 9018D 4 Аминобутановая кислота HMDB 4 Аминомасляная кислота HMDB Габа лития HMDB гамма-аминомасляная кислота, монолитиевая соль HMDB гамма-аминомасляная кислота Гамма-натриевая соль HMDB гамма-аминомасляная кислота, мононатриевая соль HMDB Кислота, гидрохлорид гамма-аминомасляная смесь HMDB HMDB HMDB 9018DB 9018 G185 9018 G185 9018 G185 9018 G185 9018 G183 9018 H2018 Гидрохлорид гамма-аминомасляная кислота HMDB гамма Аминомасляная кислота гидрохлорид HMDB 4-амино-бутаноат HMDB амино-бутановая кислота биоспайдер 9 0185 4-амино-н-масляная кислота биоспайдер 4-амино-бутилат биоспайдер Аминобутановая кислота биоспайдер биоспайдер биоспайдер Бутановая кислота, 4-амино- (9CI) биоспайдер Масляная кислота, 4-амино- биоспайдер DL-амино-гамма-гамма масляная кислота биоспайдер г-амино-БУТАНОат Генератор г-амино-БУТАНОВАЯ кислота Генератор г-амино-N- 3 г амино-N-масляная кислота Генератор г-аминомасляная кислота биоспайдер гамастан биоспайдер Гамма-аминомасляная кислота биоспайдер Гамма-аминомасляная кислота биоспайдер Гамма-аминомасляная кислота биоспайдер биоспайдер Gamma-амино BUTAN2 BUTAN2 кислота биоспайдер гамма-амино-N-бутират Генератор Гамма-амино-n-масляная кислота биоспайдер Гамма- аминокислота амино-масляная кислота биоспайдер Гамма-аминобутират биоспайдер Гамма-аминомасляная кислота биоспайдер Гамма-аминомасляная кислота 9018 Gammaage3 Гаммалон (TN) биоспайдер 9018 4 Гамулин биоспайдер Реанал биоспайдер γ-амино-бутаноат Генератор γ-амино- Генератор аминокислота аминокислота -бутират Генератор γ-амино-N-масляная кислота Генератор γ-аминобутаноат Генератор γ-γ-γаэбутановая кислота3 Генератор Генератор γ-аминомасляная кислота Генератор γ-аминомасляная кислота Генератор Прогнозируемые свойства Прогнозируемые свойства 92O123 9018 C Химическая формула 9018 C Формула 9018 4-аминобутановая кислота Идентификатор InChI InChI = 1S / C4H9NO2 / c5-3-1-2-4 (6) 7 / h2-3,5h3, (H, 6,7) Ключ InChI BTCSSZJGUNDROE-UHFFFAOYSA-N Изомерные SMILES NCCCC (O) = O Средняя молекулярная масса 103.1198 Моноизотопная молекулярная масса 103.063328537 Классификация Описание Относится к классу органических соединений, известных как гамма-аминокислоты и производные. Это аминокислоты, имеющие группу (-Nh3), присоединенную к гамма-атому углерода. Королевство Органические соединения Супер-класс Органические кислоты и производные Класс Карбоновые кислоты и производные Подкласс 9018 пептиды, аминокислоты пептиды Прямой родитель Гамма-аминокислоты и производные Альтернативные родители Заместители Молекулярный каркас Алифатические ациклические соединения Внешние дескрипторы Онтология Онтология Онтология 9019 Экспериментальный термин Физико-химические свойства — экспериментальные Свойство Значение Ссылка Физическое состояние Твердое тело 203 Физическое описание 9018 9018 Масса Состав Не доступен Точка плавления 203 oC Точка кипения Недоступно Экспериментальная растворимость в воде при 1300 мг / мл 0184 YALKOWSKY, SH & DANNENFELSER, RM (1992) Experimental log P -3.17 HANSCH, C ET AL. (1995) Экспериментальный pKa 4,05 Изоэлектрическая точка Недоступно Заряд Недоступно Оптическое 9018 УФ-данные Недоступно Плотность Недоступно Показатель преломления Недоступно 9018 Spectra Тип Описание Клавиша-заставка Вид EI-MS Масс-спектр (электронная ионизация) splash ГХ-МС 90 184 4-аминомасляная кислота , 3 ТМС , ГХ-МС Spectrum splash20-00dj-1 0000-f831f79dfcaeffa8b177
Spectrum GC-MS TMS , ГХ-МС Spectrum splash20-00di-1 0000-2de9d92a2cfc7bc655f4
Spectrum GC-MS 4-аминомасляная кислота , 3 TMS splash 73bbf2ee0803f058dbed Spectrum ГХ-МС 4-аминомасляная кислота , 3 ТМС , ГХ-МС Spectrum splash20-00di-10000-85drum4182205 9018 9018 GC-MS 4-аминомасляная кислота , 3 TMS , GC-MS Spectrum splash 20-00di-0 0000-6be23968e972a414be51
Spectrum GC-MS 4-аминомасляная кислота , без производных , ГХ-МС Spectrum splash 20-00di-1 0000-9a224763afd8ca892add
Spectrum GC-MS 4-A Спектр ГХ-МС splash20-00di- 00000-d8906d09ca1872a6391c Спектр ГХ-МС 4-аминомасляная кислота — 2 TMS , GC4-MS210db 0107-0195 splash10185-01 Spectrum GC-MS 4-аминомасляная кислота , 3 TMS , GC-MS Spectrum splash20-00di-10000-b047af158215c2b5b8e8 GC5 Spectrum 4-аминомасляная кислота , без производных , GC-MS Spectrum splash20-001i- 00000-21ea76dfb0da62031f1d
Spectrum GC-MS 90 184 4-аминомасляная кислота , недериватизированная , ГХ-МС Spectrum splash 20-00di-00000-5d60b0a446fd8122f613 Spectrum GC-MS 84 безмасляная кислота дериватизированный , ГХ-МС Spectrum splash20-00dj-1 0000-f831f79dfcaeffa8b177
Spectrum ГХ-МС 4-Аминомасляная кислота , Spectrum без разбрызгивания, GC20-004 -1 0000-2de9d92a2cfc7bc655f4
Spectrum GC-MS 4-аминомасляная кислота , без производных , GC-MS Spectrum splash -MS 4-аминомасляная кислота , недериватизированная , GC-MS Spectrum splash20-00di-10000-85d4bd98af8534428b5a Spec trum ГХ-МС 4-аминомасляная кислота , без производных , ГХ-МС Spectrum splash20-00di-0 0000-6be23968e972a414be51
Spectrum GC Spectrum -Аминомасляная кислота , без производных , ГХ-МС Spectrum splash 20-00di-1 0000-9a224763afd8ca892add
Spectrum GC-MS без производных , GC-MS , без производных , GC-, GC-MS , без производных , GC-, без производных , GC-, без производных, GC- -MS Spectrum splash20-00di- 00000-d8906d09ca1872a6391c Spectrum GC-MS 4-аминомасляная кислота , без производных db00000000, Spectrum GC-MS 4-аминомасляная кислота , без производных , GC-MS Spectrum splash 20-00di-1000 0-b047af158215c2b5b8e8 Spectrum GC-MS 4-аминомасляная кислота , недериватизированная , GC-MS Spectrum splash20-00dj-100bex18 MS 4-аминомасляная кислота , недериватизированная , ГХ-МС Spectrum splash20-0udi-0 0000-f7117dfaf9d856c
Spectrum GC203 Масляная кислота splash20-0006-1 0000-e35585a985d8128d044e
Spectrum Прогнозируемая ГХ-МС 4-Аминомасляная кислота Спектрумзед Неорганизованная ГХ3 , Положительный всплеск 20-001i- 00000-d5f55a414ff1e8c65d9d
Spectrum Прогнозируемый GC-MS 4-аминомасляная кислота 922 03, 1 TMS , прогнозируемый спектр ГХ-МС — 70 эВ, положительный всплеск 20-0fk9- 00000-4b28087240dd479 Спектр 02 Клавиша-заставка Просмотр МС / МС Спектр ЖХ-МС / МС — Quattro_QQQ 10 В, положительный (с аннотацией) splash20-0uxr-8 0000-ce0d8f44422836cd 90 / MS Spectrum 90 / MS 90 / MS
Спектр ЖХ-МС / МС — Quattro_QQQ 25 В, положительный (аннотированный) splash20-0005- 00000-8ce5afb97d7c08821da3
Spectrum MS / MS Quattro / MS_Q положительный ) splash20-0005-00000-32c433b2c Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scien tfic), положительный splash20-0udi-0 0000-5831aaabdf53f3132ae5
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic ), положительный -0a4i- 00000-9babfd4a6937ecba7318
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), Положительный splash20c09-0a4 Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-0udi-0 0000-647d55ecf98850e7a875
MS Spectrum MS Spectrum / MS Спектр ЖХ-МС / МС — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-0udi-0 0000-7c107641a38922c88fca
Spectrum MS / MS / МС Спектр — ЖХ-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-000i- 00000-86718b349efad6334e3a
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbit Scient XL ), Положительный splash20-000i- 00000-a1e84e55e4b6c6628d5d
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo 9018-2020), 0006-000 00-29c22bf0ed844d09c0af
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) 10 В, отрицательный Spectrum МС / МС Спектр ЖХ-МС / МС — LC-ESI-QQ (API3000, Applied Biosystems) 20 В, отрицательный splash ЖХ-МС / МС Spe ctrum — LC-ESI-QQ (API3000, Applied Biosystems) 30 В, отрицательный splash20-001i- 00000-a14a52dc59bf9988bb44
Spectrum MS / MS LC-MS / ESI Spectrum — LC-MS / ES Spectrum — API3000, Applied Biosystems) 10 В, положительный splash 20-0udi-5 0000-20c55b2809389d5ad83b
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) splash20-000i- 00000-eca4c5aefca98751a11e
Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) 30V, положительный splash20184-02020 f0783316e092409 Spectrum MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) 40 В, положительный splash20-0005- 00000-818379eb9c МС / МС
ЖХ-МС / М S Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) 50 В, положительный splash20-0005- 00000-8b48126992d7fa242636
Spectrum MS / MS LC-MS / MS Spectrum — LCTO-ES (UPLC Q-Tof Premier, Waters), положительный splash20-000i- 00000-7d4636efbc4e5d75872e
Spectrum Прогнозируемый MS / MS Прогнозируемый спектр LC-MS / MS — 10Vi-203, положительный splash 00000-655f9d93a35bfa537583 Спектр Прогнозируемый МС / МС Прогнозируемый спектр ЖХ-МС / МС — 20 В, положительный splash20-00ku- 00000-4a334d5e272576f62403 9018 МС 9018 Спектр 9018 Прогнозируемый 9018 MS 9018 Спектр ЖХ-МС / МС — 40 В, положительный
splash20-0006- 00000-4a13b03446b3370ccd43
Спектр Прогнозируемый МС / МС Прогнозируемый спектр ЖХ-МС / МС — 10 В, положительный splash20-000i-00000-655f9d93a35bfa537583 Spectrum Прогнозируемый MS / MS Прогнозируемый спектр LC-MS / MS — 20 В, положительный splash20-00ku- 000003276 9018 9018 9018 Spectrum
ЯМР Внешние ссылки ChemSpider ID 116 ChEMBL ID CHEMBL96 CHEMBL96 KEG2 9018 ID соединения 9018 Соединение 9018 ID соединения Pubchem Substance ID Недоступно ChEBI ID 16865 Phenol-Explorer ID Недоступно DrugBank ID 9018M 9018M CRC / DFC (Словарь Foo d Соединения) ID Недоступно EAFUS ID 165 Dr.Duke ID GABA | ГАММА-АМИНОБУТИРИНОВАЯ КИСЛОТА BIGG ID 34652 KNApSAcK ID C00001337 9018 HET VMH ID Недоступно Flavornet ID Недоступно GoodScent ID rw1198861 SuperScent ID 9018 G185 SuperScent ID 9018 Phenol-Explorer Metabolite ID Недоступно Дубликат IDS Недоступно Старое IDS DFC Недоступно Ассоциированные продукты питания 9024 Среднее значение Ссылка Продукты питания Ссылка Биологические эффекты и взаимодействия Влияние на здоровье / биоактивность Описание Каталожный номер аналептик 35337 Любой препарат, усиливающий деятельность центральной нервной системы. DUKE антицефалагическое 52217 Любое вещество, введенное в живой организм с терапевтической или диагностической целью. DUKE антицеребротический 52217 Любое вещество, введенное в живой организм с лечебной или диагностической целью. DUKE антихореический 52217 Любое вещество, введенное в живой организм с лечебной или диагностической целью. DUKE противосудорожное средство 52217 Любое вещество, введенное в живой организм с лечебной или диагностической целью. DUKE антигипертензивное 52217 Любое вещество, введенное в живой организм с лечебной или диагностической целью. DUKE anti insomniac 52217 Любое вещество, введенное в живой организм с терапевтической или диагностической целью. DUKE anti insomnic 52217 Любое вещество, введенное в живой организм с терапевтической или диагностической целью. DUKE антилетаргический 52217 Любое вещество, введенное в живой организм с лечебной или диагностической целью. DUKE антистресс 52217 Любое вещество, вводимое в живой организм с лечебной или диагностической целью. DUKE противошумные DUKE анксиолитические 35474 Анксиолитики — это агенты, которые снимают тревогу, напряжение и тревожные расстройства, оказывают успокаивающее действие. сознание или неврологические состояния. DUKE сердечно-сосудистые 38070 Препарат, используемый для лечения или профилактики сердечных аритмий.Антиаритмические препараты могут влиять на фазу поляризации-реполяризации потенциала действия, на его возбудимость или рефрактерность, или на проведение импульсов или чувствительность мембран в сердечных волокнах. DUKE ингибитор центральной нервной системы 35470 Класс препаратов, оказывающих как физиологические, так и психологические эффекты посредством различных механизмов, затрагивающих центральную нервную систему. DUKE диуретик 35498 Средство, которое способствует выведению мочи за счет его воздействия на функцию почек. DUKE гипотензивный DUKE нейро-ингибитор 35222 Вещество, снижающее скорость химической реакции. Герцог нейротоксичный 50910 Яд, нарушающий функции нервной системы. DUKE нейромедиатор 25512 Эндогенное соединение, которое используется для передачи информации через синапс между нейроном и другой клеткой. DUKE успокоительное 35717 Депрессант центральной нервной системы, используемый для того, чтобы вызвать сонливость или сон, или уменьшить психологическое возбуждение или тревогу. DUKE транквилизатор DUKE Ферменты Имя Gene Имя UniProt ID Глицин amidinotransferase, митохондриальная GATM P50440 4-аминобутират аминотрансферазы, митохондриальная ABAT P80404 глутаматдекарбоксилаза 2 GAD2 Q05329 глутаматдекарбоксилаза 1 GAD1 Q99259 Аргинин декарбоксилаза ADC Q96A70 Глутаминсинтетаза GLUL P15104 Вероятный глутамат — тРНК-лигаза, митохондриальная Q18AHJP EGM 9018 Связанный с рецептором масляной кислоты протеиноподобный 2 GABARAPL2 P60520 Пути Метаболизм Органические свойства ароматы аромат цитаты пикантные мясистый Файлы MSDS Недоступно Справочные материалы Справочные материалы по синтезу Отсутствуют — Герцог, Джеймс.Доктор Фитохимические и этноботанические базы данных Герцога. Министерство сельского хозяйства США ». Служба сельскохозяйственных исследований, по состоянию на 27 апреля (2004 г.). Гамма-аминомасляная кислота (ГАМК) — версия JMol
Гамма-аминомасляная кислота (ГАМК) — версия JMol Молекула релаксации …
Hull Collegiate School, Халл, Великобритания Чувствуете стресс и тревогу? Все, что вам нужно, это ГАМК!
Что такое ГАМК?
Как работает ГАМК?
Что бы произошло, если бы ГАМК не существовало?
Что такое история ГАМК
Каковы виды использования ГАМК
Ссылки:
Отображение метабокарты для гамма-аминомасляной кислоты (HMDB0000112)
Информация о записи Версия 4.0 Статус Дата обнаружения и количественная оценка : 48: 42 UTC Дата обновления 2020-11-09 23:12:04 UTC Идентификатор HMDB HMDB0000112 Вторичные номера доступа 9019 9019 Общее название гамма-аминомасляная кислота Описание гамма-аминомасляная кислота (ГАМК) является ингибирующим нейротрансмиттером, обнаруживаемым в нервных системах широко различных видов, включая человека.Это главный тормозной нейромедиатор в центральной нервной системе позвоночных. У позвоночных ГАМК действует на тормозные синапсы в головном мозге. Он действует путем связывания со специфическими трансмембранными рецепторами плазматической мембраны как пре-, так и постсинаптических нейронов. Это связывание вызывает открытие ионных каналов, позволяя либо потоку отрицательно заряженных ионов хлорида в клетку, либо положительно заряженным ионам калия из клетки. Обычно это приводит к отрицательному изменению трансмембранного потенциала, обычно вызывая гиперполяризацию.Известны три основных класса рецепторов ГАМК (PMID: 10561820). К ним относятся ионотропные рецепторы GABA-A и GABA-C, которые сами по себе являются ионными каналами, и метаботропные рецепторы GABA-B, которые представляют собой рецепторы, связанные с G-белком, которые открывают ионные каналы через посредников, известных как G-белки (PMID: 10561820). Активация рецептора ГАМК-В с помощью ГАМК вызывает гиперполяризацию нейрональной мембраны и, как следствие, ингибирование высвобождения нейромедиатора. Помимо сайтов связывания для ГАМК, рецептор ГАМК-А имеет сайты связывания для бензодиазепинов, барбитуратов и нейростероидов.Рецепторы ГАМК-А связаны с каналами хлорид-иона. Следовательно, активация рецептора ГАМК-А вызывает повышенный поток внутрь хлорид-ионов, что приводит к гиперполяризации мембраны и ингибированию нейронов (PMID: 10561820). После высвобождения в синапс свободная ГАМК, которая не связывается ни с рецепторными комплексами ГАМК-А, ни с ГАМК-В, может захватываться нейронами и глиальными клетками. Считается, что четыре различных белка-переносчика ГАМК (GAT-1, GAT-2, GAT-3 и BGT-1), которые различаются по своему распределению в ЦНС, опосредуют захват синаптической ГАМК нейронами и глиальными клетками.Подтип рецептора ГАМК-А регулирует возбудимость нейронов и быстрые изменения возбуждения страха, такие как тревога, паника и реакция на острый стресс (PMID: 10561820). Лекарства, которые стимулируют рецепторы ГАМК-А, такие как бензодиазепины и барбитураты, обладают анксиолитическим и противосудорожным действием за счет снижения возбудимости нейронов, опосредованного ГАМК-А, что эффективно повышает судорожный порог. Антагонисты ГАМК-А вызывают судороги у животных, и при исследовании пациентов с паническим расстройством наблюдается снижение связывания рецептора ГАМК-А при исследовании с помощью позитронно-эмиссионной томографии (ПЭТ).Нейроны, которые производят ГАМК в качестве продукции, называются ГАМКергическими нейронами и оказывают главным образом ингибирующее действие на рецепторы позвоночных. Средние шиповатые нейроны (MSN) являются типичным примером ингибирующих ГАМКергических клеток ЦНС. Было показано, что ГАМК играет возбуждающую роль у позвоночных, особенно в развивающейся коре головного мозга. Организмы синтезируют ГАМК из глутамата с использованием фермента декарбоксилазы L-глутаминовой кислоты и пиридоксальфосфата в качестве кофактора (PMID: 12467378). Стоит отметить, что при этом происходит преобразование основного возбуждающего нейромедиатора (глутамата) в основной тормозной (ГАМК).Лекарства, которые действуют как агонисты рецепторов ГАМК (известные как аналоги ГАМК или ГАМКергические препараты) или увеличивают доступное количество ГАМК, обычно обладают расслабляющим, успокаивающим и противосудорожным действием. Во многих исследованиях экспериментальной эпилепсии и эпилепсии у людей обнаружен дефицит ГАМК в спинномозговой жидкости и головном мозге. Бензодиазепины (такие как валиум) полезны при эпилептическом статусе, потому что они действуют на рецепторы ГАМК. ГАМК увеличивается в головном мозге после приема многих противосудорожных препаратов. Следовательно, очевидно, что ГАМК является противоэпилептическим питательным веществом.Ингибиторы метаболизма ГАМ также могут вызывать судороги. Синдромы спастичности и непроизвольных движений, такие как болезнь Паркинсона, атаксия Фридрейха, поздняя дискинезия и хорея Хантингтона, при исследовании уровня аминокислот характеризуются низким уровнем ГАМК. Испытания от 2 до 3 г ГАМК, вводимых перорально, оказались эффективными при различных синдромах эпилепсии и спастичности. Агенты, повышающие уровень ГАМК, также полезны для снижения гипертонии. Три грамма внутрь эффективны для контроля артериального давления. ГАМК снижается при различных энцефалопатиях.ГАМК может снизить аппетит и снижается при гипогликемии. ГАМК снижает уровень сахара в крови у диабетиков. Хронические мозговые синдромы также могут быть отмечены дефицитом ГАМК. Витамин B6, марганец, таурин и лизин могут увеличивать как синтез ГАМК, так и эффекты, в то время как аспарагиновая кислота и глутаминовая кислота, вероятно, подавляют эффекты ГАМК. Сообщалось о низком уровне ГАМК в плазме у некоторых пациентов с депрессией, и он может быть полезным маркером аффективных расстройств. ГАМК играет важную роль в эмбриональном развитии, особенно в развитии лица, что подтверждается ассоциацией волчьей пасти у трансгенных мышей с дефицитом GAD67 (глутаматдекарбоксилазы).Недавнее исследование населения Японии показало связь у пациентов с несиндромной расщелиной губы с расщелиной неба или без нее и со специфическими гаплотипами GAD67 (PMID: 23842532). Необычно высокие уровни ГАМК (особенно в головном мозге) могут быть токсичными, а ГАМК может действовать как нейротоксин и метаботоксин. Нейротоксин — это соединение, которое повреждает мозг и / или нервную ткань. Метаботоксин — это эндогенно продуцируемый метаболит, который вызывает неблагоприятные последствия для здоровья при хронически высоких уровнях. Хронически высокие уровни ГАМК связаны по крайней мере с пятью врожденными ошибками метаболизма, включая D-2-гидроксиглутаровую ацидурию, дефицит 4-гидроксимасляной ацидурии / янтарного полуальдегиддегидрогеназы, дефицит ГАМК-трансаминазы, гомокарнозиноз и гипербета-аланинемию.Почти все эти состояния связаны с судорогами, гипотонией, интеллектуальным дефицитом, макроцефалией, энцефалопатией и другими серьезными неврологическими или нервно-мышечными проблемами. Повышенные уровни ГАМК, по-видимому, изменяют функцию рецептора ГАМК-В, который может играть роль в тонико-клонических припадках, которые часто наблюдаются у пациентов с вышеуказанными расстройствами. ГАМК также является микробным метаболитом, ГАМК с мочой продуцируется Lactobacillus и Bifidobacterium (PMID: 24621061). Структура Синонимы Значение Источник 4-Аминобутановая кислота Кислота E E E 4Abu ChEBI GABA ChEBI ГАММА-АМИНО-БУТАНОВАЯ КИСЛОТА ChEBI 8-Гамма-бутановая кислота -N-AB5 -гамма-бута-A ChEBI гамма-Aminobuttersaeure ChEBI Омеги-аминомасляная кислота ChEBI Piperidic кислота ChEBI пиперидиновой кислота ChEBI Гаммалон KEGG 4-аминобутират Кегг 4-аминобутаноат Генератор г-АМИНО-бутаноат Генератор г-АМИНО-бутановая кислота г-АМИНО-бутановая кислота Генератор AMO-бутановая кислота Генератор Гамма Генератор Генератор Г-амино-бутаноат Генератор Г-аминобутановая кислота Генератор г-амино-N-бутират Генератор Генератор гамма-амино-N-бутират Генератор Γ-амино-N-бутират Генератор Γ-амино-N- 3 Генератор 9018 3 9018 масляная кислота 9018 g-аминобутаноат генератор g-аминобутановая кислота генератор гамма-аминобутаноат генератор 901 84 Γ-аминобутаноат Генератор Γ-аминобутановая кислота Генератор g-Aminobuttersaeure Генератор 9020buttersaeure Генератор 9020buttersaeure аминокислота Генератор Пиперидат Генератор Пиперидинат Генератор g-Aminobutyrate Generator g-Aminobutyrate Генератор g18 Aminobuty Γ-аминомасляная кислота Генератор Γ-аминомасляная кислота Генератор 3-карбоксипропиламин HMDB HMDB 9018 Hb203 9018 Hb3 9018 Hb204 9018 Hb3 0182 Gamarex HMDB гамма-аминобутират HMDB гамма-аминомасляная кислота HMDB Gammalone HMDB Gammar HMDB Gammasol HMDB Mielogen HMDB Mielomade HMDB W-Aminobutyrate HMDB Aminobutyrate W-Aminobutyrat 1) HMDB гамма-аминомасляная кислота, гидрохлорид HMDB гамма-аминомасляная кислота, цинковая соль (2: 1) HMDB3 8282 HMDB382 4 Аминомасляная кислота HMDB Габа-литий 90 184 HMDB гамма Аминомасляная кислота, монолитиевая соль HMDB гамма Аминомасляная кислота, мононатриевая соль HMDB гамма-аммонитриевая кислота кислота, мононатриевая соль HMDB Кислота, гидрохлорид гамма-аминомасляная кислота HMDB Аминалон HMDB GAB183 9018D гамма-аминомасляная кислота, гидрохлорид HMDB 4-амино-бутаноат HMDB гамма-аминомасляная кислота 9018 4 9018 4 9018 4 9018 4 9018 4 9018 4 9018 4 9018 4 9018 4 9018 4 H 9 НЕТ 2 Средний молекулярный вес 103.1198 Моноизотопный молекулярный вес 103.063328537 Название IUPAC 4-аминобутановая кислота Традиционное название гамма (аминокислота) 9018-123 -2 УЛЫБКИ Идентификатор InChI Ключ InChI BTCSSZJGUNDROE-UHFFFAOYSA-N Химическая таксономия Описание , производные органических кислот, принадлежащих к классу органических соединений .Это аминокислоты, имеющие группу (-Nh3), присоединенную к гамма-атому углерода. Королевство Органические соединения Супер-класс Органические кислоты и производные Класс Карбоновые кислоты и производные Подкласс 9018 пептиды, аминокислоты пептиды Прямой родитель Гамма-аминокислоты и производные Альтернативные родители Заместители Молекулярная структура Алифатические ациклические соединения Внешние дескрипторы Онтология Дистрибьюция 9685 Процесс Роль Физические свойства Состояние Твердое тело Экспериментальные свойства Свойство Значение Точка плавления 203 ° C Недоступно Точка кипения Недоступно Недоступно Растворимость в воде 1300 мг / мл 9018 9018 -3.17 HANSCH, C ET AL. (1995) Прогнозируемые свойства Spectra Spectrum Type 9019 Клавиша Splash 9018 Спектр ГХ-МС — GC-EI-TOF (система Pegasus III TOF-MS, Leco; GC 6890, Agilent Technologies) (3 ТМС) splash20-00dj-1 0000-f831f79dfcaeffa8b177
Spectrum GC МС 85 Спектр ГХ-МС — GC-EI-TOF (система Pegasus III TOF-MS, Leco; GC 6890, Agilent Technologies) (3 ТМС) splash20-00di-1 0000-2de9d92a2cfc7bc655f4
Spectrum GC -MS Спектр ГХ-МС — GC-EI-TOF (система Pegasus III TOF-MS, Leco; GC 6890, Agilent Technologies) (3 TMS) splash20-00di-1 0000-73bbf2ee0803f058dbed
Spectrum GC- MS Спектр ГХ-МС — GC-EI-TOF (система Pegasus III TOF-MS, Leco; GC 6890, Agilent Technologies) (3 TMS) splash20-00di-10000-85d4bd98af8534428b5a Spectrum GC-MS GC-MS Spectrum — GC-EI-TOF (Pegasus III TOF-MS system, Pegasus III TOF-MS system, Pegasus III TOF-MS system ; GC 6890, Agilent Technologies) (3 TMS) splash20-00di-0 0000-6be23968e972a414be51
Spectrum GC-MS GC-MS Spectrum — GC-EI-TOF (Pegasus III TOF-MS system, Leco; GC 6890, Agilent Technologies) (без производных) splash20-00di-1 0000-9a224763afd8ca892add
Spectrum GC-MS GC-MS Spectrum — GC-EI-TOF (Pegasus III TOF (Pegasus III TOF) система, Leco; ГХ 6890, Agilent Technologies) (3 ТМС) splash20-00di- 00000-d8906d09ca1872a6391c Spectrum GC-MS GC-MS Spectrum — GC-MS (2 splash204) -0udi-1 0000-54db7e217
045519 Спектр GC-M S Спектр ГХ-МС — ГХ-МС (3 ТМС) splash20-00di-10000-b047af158215c2b5b8e8 Спектр ГХ-МС Спектр ГХ-МС — EI-B (без производных) splash20-001i- 00000-21ea76dfb0da62031f1d
Spectrum GC-MS GC-MS Spectrum — EI-B (без производных) splash20-00di-00000f6f2013128 -MS Спектр ГХ-МС — GC-EI-TOF (без производных) splash20-00dj-1 0000-f831f79dfcaeffa8b177
Спектр GC-MS GC-MS Spectrum-TOF (Без производных) splash20-00di-1 0000-2de9d92a2cfc7bc655f4
Spectrum ГХ-МС Спектр ГХ-МС — GC-EI-TOF (Без производных) -00-80bb-210000 splash0000 Спектр 90 203 ГХ-МС Спектр ГХ-МС — GC-EI-TOF (без производных) splash 20-00di-10000-85d4bd98af8534428b5a Спектр ГХ-МС Спектр ГХ-ЭМС -TOF (без производных) splash20-00di-0 0000-6be23968e972a414be51
Spectrum GC-MS GC-MS Spectrum — GC-EI-TOF (Non-Derivatized) splash20 -9a224763afd8ca892add Спектр ГХ-МС Спектр ГХ-МС — GC-EI-TOF (без производных) splash20-00di- 00000-d8906d09ca1872a418204 9018 9018 MS Spectrum 9018 Спектр ГХ-МС — ГХ-МС (без производных) splash20-0udi-1 0000-54db7e217
045519 Спектр ГХ-МС Спектр ГХ-МС — ГХ-МС (без производных3) 9020 -00di-10000-b047af158215c2b5b8e8 Спектр ГХ-МС Спектр ГХ-МС — GC-EI-TOF (без производных) splash20-00dj-1 0000-1219470a0be188da64e6
Spectrum Spectrum GC203 Spectrum GC203 Spectrum GC203 Spectrum GC203 Spectrum GC203 Spectrum GC-MS GC-EI-TOF (без производных) splash20-0udi-0 0000-f7117dfaf9d856c
Spectrum GC-MS GC-MS Spectrum — GC-EI-TOF (Non- splash) 0006-10000-e35585a985d8128d044e
Спектр Прогнозируемый спектр ГХ-МС Прогнозируемый спектр ГХ-МС — ГХ-МС (без производных) — 70 эВ, положительный splash20-001if- 00a4-d
Прогнозируемый ГХ-МС Прогнозируемый спектр ГХ-МС — ГХ-МС (1 ТМС) — 70 эВ, положительный брызги 20-0fk9- 00000-4b28087240dd479 Спектр LC-MS LC-MS LC-MS МС / МС Спектр — Quattro_QQQ 10 В, положительный (аннотированный) splash20-0uxr-8 0000-ce0d8f44422836cd9965
Spectrum LC-MS / MS LC-MS / MS Spectrum — Quattro_QQQ000 25V, Positive ( — -8ce5afb97d7c08821da3 splash203-
)
Спектр ЖХ-МС / МС Спектр ЖХ-МС / МС — Quattro_QQQ 40 В, положительный (с аннотацией) splash20-0005-00000-32c43376202c9 MS Спектр ЖХ-МС / МС — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-0udi-0 0000-5831aaabdf53f3132ae5
Spectrum LC-MS203 LC 90-184 МС / МС спектр — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash 20-0a4i- 00000-9babfd4a6937ecba7318
Spectrum LC-MS / MS Spectrum LC-MS — MS / MS LC-ESI-ITFT (LTQ Orbitrap XL, Th ermo Scientfic), положительный splash20-0a4i- 00000-e1c0c1485d846e9b123b
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQr Orbitrap XL) splash20-0udi-0 0000-647d55ecf98850e7a875
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic 90udi ), положительный splash -0 0000-7c107641a38922c88fca
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-000i- Spectrum00000-a1e84e55e4b6c6628d205
ЖХ-МС / МС ЖХ-МС / М S Spectrum — LC-ESI-ITFT (LTQ Orbitrap XL, Thermo Scientfic), положительный splash20-0006-000 00-29c22bf0ed844d09c0af
Spectrum LC-MS / MS Spectrum LC-MS — LC-MS ESI-QQ (API3000, Applied Biosystems) 10 В, отрицательный splash20-0udi-0 0000-1d00adad47e42c60c340
Spectrum LC-MS / MS LC-MS / MS Spectrum (API-ESI-Q) Applied Biosystems) 20 В, отрицательный splash20-0udi-1 0000-47b195fb74720cc99464
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) splash20-001i- 00000-a14a52dc59bf9988bb44
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-QQ (API3000, Applied Biosystems) 10 В, положительный splash204 9018—0204 5 0000-20c55b2809389d5ad83b
Spectrum LC-M S / MS Спектр ЖХ-МС / МС — LC-ESI-QQ (API3000, Applied Biosystems) 20 В, положительный splash20-000i- 00000-eca4c5aefca98751a11e
Спектр LC- LC- LC- LC- LC- -МС / МС-спектр — ЖХ-ESI-QQ (API3000, Applied Biosystems) 30 В, положительный всплеск 20-014j- 00000-f0783316e092
409 Спектр ЖХ-МС / МС Спектр ЖХ-МС / МС-МС / МС Спектр ЖХ-МС / МС LC-ESI-QQ (API3000, Applied Biosystems) 40 В, положительный splash20-0005- 00000-81837eb9c0b926cb0e81
Spectrum LC-MS / MS LC-MS / ES Spectrum — LCQ-Q-LCQ- API3000, Applied Biosystems) 50 В, положительный splash20-0005- 00000-8b48126992d7fa242636
Spectrum LC-MS / MS LC-MS / MS Spectrum — LC-ESI-QTOF (UPLC) Waters), положительный брызги 20-000i- 00000-7d4636efbc4e5d75872e
Spectrum Прогнозируемый спектр ЖХ-МС / МС Прогнозируемый спектр ЖХ-МС / МС — 10 В, положительный Всплеск 20-000i-00000-655f9d93a35bfa537583 Спектр Прогнозируемый ЖХ-МС / МС Спектр МС — 20 В, положительный splash20-00ku- 00000-4a334d5e272576f62403
Спектр Прогнозируемый ЖХ-МС / МС Прогнозируемый спектр ЖХ-МС / МС — 400003333 брызги b000000002043 Спектр Прогнозируемый спектр ЖХ-МС / МС Прогнозируемый спектр ЖХ-МС / МС — 10 В, положительный всплеск 20-000i-00000-655f9d93a35bfa537583 Спектр МС Прогнозируемый спектр ЖХ-МС / МС — 20 В, положительный splash 20-00ku- 00000-4a334d5e272576f62403
Спектр MS Масс-спектр (электронная ионизация) 90 203 splash20-001i-00000-dbf4f9e19a35f953a189
Спектр 1D ЯМР 1H ЯМР спектр Недоступно Спектр 1D Спектр 9018 9018 Спектр NMR 9018 9018 1D ЯМР Спектр 1H ЯМР Недоступно Спектр 1D ЯМР 13C ЯМР-спектр Недоступно Спектр 1D Спектр 1D Спектр 2D ЯМР [1H, 13C] 2D ЯМР-спектр Недоступно Спектр Биологические свойства Внеклеточные местоположения 0 Местонахождение биопрепаратов Расположение тканей Пути пути Нормальные концентрации Кровь Обнаружено, но не определено количественно Не лет Оба Нормальный подробности Кровь Обнаружено и определено количественно 0.110 +/- 0,023 мкМ Взрослый (> 18 лет) Оба Нормальный подробности Кровь Обнаружено и количественно определено 0,120-0,500 мкМ Дети (1-13 лет) ) Оба Нормальный детали Цереброспинальная жидкость (ЦСЖ) Обнаруженная и количественная оценка 0,24 +/- 0,096 мкМ Взрослые (> 18 лет) Оба Нормальные подробности Цереброспинальная жидкость (ЦСЖ) Обнаруженная и количественная оценка 0.0200-0,100 мкМ Новорожденный (0-30 дней) Не указано Нормальный подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0,32 (0,2 -0,44) 0,32 (0,2 -0,44) Взрослый (> 18 лет) Оба Нормальный детали Цереброспинальная жидкость (CSF) Выявленная и количественная оценка 0,23 +/- 0,075 мкМ Взрослый (> 184 года) Оба Нормальный подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0.233 +/- 0,075 мкМ Не указано Не указано Нормальное подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0,093 (0,083184> 9020 мкМ) лет) Оба Нормальный подробности Кал Обнаружены, но не определены количественно Не определены количественно Дети (6-18 лет) Не указаны Нормальные данные Нормальные данные Кал Обнаружен, но не определен количественно Не определен количественно Не определен Оба Нормальный подробности Фекалии Определен, но не определен Не определен Не определен Не определен 9018 Задано Нормальное детали F eces Обнаружено и количественно определено 4363.856 (2909,238-5818,475) нмоль / г влажных фекалий Младенцы (0–1 год) Не указано Нормальные детали Кал Обнаружено и количественно определено 03 281280,2 нмоль / г влажных фекалий Младенцы (0-1 год) Не указано Нормальные детали Кал Обнаружено и количественно определено 4202.232 (2359.715-6044.749) нмоль / г влажных фекалий Младенцы (0-1 год) Не указано Нормальное подробности Кал Обнаружено, но не определено количественно Не определено количественно Взрослые (>> старый) Оба Нормальный подробности Фекалии Обнаружены, но не определены количественно Не определены количественно Взрослые (> 18 лет) Самка Нормальные данные Фекалии Обнаружены и количественно определены 576.999 +/- 520,754 нмоль / г влажных фекалий Не указано Не указано Нормальное подробности Фекалии Обнаружены, но не определены количественно Не определены количественно Нормальный подробности Фекалии Обнаружены, но не определены количественно Количественно не определены Взрослые (> 18 лет) Оба Нормальные Подробные данные Обнаружены колличественные Не определено количественно Взрослый (> 18 лет) Оба Нормальный подробности Кал Обнаружен, но не определен количественно Не определен количественно Взрослый (> 18 лет) Оба Нормальный подробности Слюна 9 0203 Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Мужчина Нормальный подробности Слюна Обнаружен, но не определен количественно 18203> старый) Оба Нормальный подробности Слюна Обнаружено, но не определено количественно Не определено количественно Взрослые (> 18 лет) Оба Нормальные данные Слюна Обнаруженная и количественная оценка > 10 мкМ Взрослые (> 18 лет) Оба Нормальные детали Слюна Обнаружено и количественно определено 1.23 +/- 0,57 мкМ Взрослый (> 18 лет) Женский Нормальный подробности Слюна Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Мужской Нормальный подробности Слюна Обнаружен, но не определен количественно Не определен количественно Взрослый (> 18 лет) Мужчина Нормальный 9018 Слюна подробности Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Мужчина Нормальный подробности Слюна Обнаружен, но не определен количественно Взрослый (> 184 лет) Не определен количественно ) Мужской Нормальный детали Слюна Обнаружено но не определен количественно количественно не определен Взрослый (> 18 лет) мужской нормальный подробные сведения слюна обнаружен, но не определен количественно не определен количественно взрослый (возраст> 18 лет) Мужской Нормальный подробности Слюна Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Мужчина 90iva203 Нормальный Нормальный Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Мужчина Нормальный подробности Слюна Обнаружен, но не определен количественно 18203 9018 (количественно не определено 9018) старый) Наружный Нормальный детали Моча Обнаруженная и количественная оценка <0.68 мкмоль / ммоль креатинин Дети (1-13 лет) Не указано Нормальный подробности Моча Обнаруженный и количественно определенный 0,09 +/- 0,10 мкмоль ммоль креатинин (0-1 год) Оба Нормальный подробности Моча Обнаруженный и количественный <0,461 мкмоль / ммоль креатинин Взрослый (> 18 лет) Оба Оба Нормальный подробности Моча Обнаруженная и количественная оценка <0.57 мкмоль / ммоль креатинин Подростки (13-18 лет) Не указано Нормальное подробности Моча Выявленная и количественная оценка 8,037 +/- 2,502 203 мкмоль / детский креатинин 9018 (1-13 лет) Не указано Нормально подробности Моча Обнаруженный и количественный 0,17 +/- 0,025 мкмоль / ммоль креатинин Взрослый (> 184 ) Женский Нормальный подробности Моча Обнаружено и количественно определено 0.25 +/- 0,20 мкмоль / ммоль креатинина Взрослый (> 18 лет) Не указано Нормальный подробности Моча Выявленный и количественный Креатинин для взрослых <0,57 мкмоль ммоль (> 18 лет) Не указано Нормальное подробности Моча Обнаруженный и количественный 1,3 (0,0-2,8) мкмоль / ммоль креатинина Новорожденный (0-30 дней) Оба Нормальный подробности Моча Обнаруженная и количественная оценка <2,83 мкмоль / ммоль креатинина Младенцы (0-1 год старый) Не указано Нормально детали Моча Выявлено и определено количественно <1,47 мкмоль / ммоль креатинина Дети (1-13 лет) подробности Моча Обнаруженная и количественная оценка 103.89 (72,43 — 169,51) мкмоль / ммоль креатинин Взрослые (> 18 лет) Оба Нормальный подробности Моча Обнаруженный и количественный анализ 2,26 +/- 10,1 ммоль Новорожденный (0-30 дней) Оба Нормальный подробности Моча Обнаруженный и количественный 4,52 +/- 18,09 мкмоль / ммоль креатинин -30 дней Новорожденный (0-30 дней) ) Оба Нормальный подробности Моча Обнаружено и определено количественно <1.24 мкмоль / ммоль креатинина Дети (1-13 лет) Не указано Нормальное подробности Моча Выявленная и количественная оценка 17-20 мкмоль / ммоль креатинин -30 дней) Оба Нормальный подробности Моча Обнаруженный и количественный 2,9 (2,7-3,3) мкмоль / ммоль креатинин Взрослые (> 18 лет) Оба Нормальный подробности Моча Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Оба Нормальный подробности Кровь Обнаруженная и количественная оценка 0 .46 +/- 0,11 мкм Взрослый (> 18 лет) Оба Печеночная энцефалопатия подробности Кровь Обнаружено и количественно Взрослый (0,0-2,0) мкм 18 лет) Оба Дефицит Габа-трансаминаз подробности Кровь Обнаружено и количественно 1,45 (0,00-2,9) мкМ Дети (1-13 лет) Оба Дефицит трансаминаз ГАМК подробности Кровь Обнаружено и количественно определено 1.84 +/- 0,215 мкМ Пожилые люди (> 65 лет) Оба Болезнь Альцгеймера подробности Цереброспинальная жидкость (ЦСЖ) Выявлено и количественно определено 0,13 Взрослый (> 18 лет) Оба Спиноцеребеллярная дегенерация детали Спинномозговая жидкость (CSF) Выявлено и количественно определено 0,011 мкМ Новорожденный Hyperekplexia подробности Цереброспинальная жидкость (CSF) Выявление и количественная оценка 1.5 +/- 1,1 мкМ Взрослый (> 18 лет) Не указано Эпилепсия подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0,1 (0,063 мкМ) 0,1 (0,063 — 0,063) Дети (1-13 лет) Не указано Фебрильный припадок детали Спинномозговая жидкость (CSF) Выявлено и количественно определено 0,52 — 0,57 мкм Дети (1-13 лет ) ) Не указано Эпилепсия подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0.11 (0,067 — 0,176) мкМ Дети (1-13 лет) Не указано Эпилепсия подробности Цереброспинальная жидкость (ЦСЖ) Обнаружено и количественно определено 0,05 (0,00-103) 0,05 uM Взрослые (> 18 лет) Оба Дефицит Габа-трансаминазы подробности Цереброспинальная жидкость (CSF) Выявлено и количественно определено 2,4 (0.00-4,8) мкМ Дети (1-13 лет) Оба Дефицит трансаминазы ГАМК подробности Цереброспинальная жидкость (ЦСЖ) Выявлено и количественно определено 0,1 (0,0M) Дети (1-13 лет) Оба Менингит подробности Цереброспинальная жидкость (CSF) Выявлено и количественно определено 0,016 мкМ Младенцы (0-1 лет) Не указано Типичные пиридоксин-зависимые судороги подробности Цереброспинальная жидкость (CSF) Выявлено и количественно определено 4.0 +/- 4,2 мкМ Взрослые (> 18 лет) Оба Эпилепсия подробности Цереброспинальная жидкость (CSF) Выявлено и количественно определено для взрослых 0,00013203 0,00013203 +/- 90 (> 18 лет) Оба Туберкулезный менингит подробности Цереброспинальная жидкость (ЦСЖ) Выявленная и количественная оценка 0,00013 +/- 0,00002 мкМ Взрослый (возраст> ) Оба Менингит подробности Кал Обнаружены, но не определены количественно Количественно не определены Дети (6-18 лет) Не указаны Болезнь Крона Обнаружено, но не определено количественно Количественно не определено Дети (6-18 лет) Не указано 9018 4 Язвенный колит подробности Кал Обнаружены, но не определены количественно Количественно не определены Дети (6-18 лет) Не указаны Неклассифицированные IBD Обнаружено, но не определено количественно Не определено количественно Не указано Не указано Инфекция Cryptosporidium подробности Кал Обнаружено, но не определено количественно Возраст> Оба Колоректальный рак подробности Кал Обнаружен, но не определен количественно Не определен количественно Взрослые (> 18 лет) Оба Колоректальный рак Кал Обнаружен но не определено количественно количественно не определено взрослые (> 18 лет) оба синдром раздраженного кишечника подробности кал обнаружены, но не определены количественно взрослые старый) Оба Колоректальный рак подробности Слюна Обнаружено, но не определено количественно Количественно не определено Взрослые (> 18 лет) Оба Рак полости рта Слюна Обнаружен, но не определен количественно Количественно не определен Взрослый (> 18 лет) Женский Рак груди подробные сведения Слюна Количественно не определен 9018 Определен, но не определен Взрослый (> 18 лет) Не указано Поджелудочная железа рак подробности Слюна Обнаружено, но не определено количественно Количественно не определено Взрослые (> 18 лет) Не указано Заболевания пародонта 92iva123 подробности 1.08 +/- 0,34 мкМ Взрослые (> 18 лет) Мужчины Болезнь Альцгеймера подробности Слюна Обнаружено и количественно определено 1,78 +/- 0,503 мкМ 90 лет) Мужчина Деменция лобно-височной доли подробности Слюна Обнаружена, но не определена количественно Количественно не определена Взрослые (> 18 лет) Скворечник ) подробности Слюна Обнаружена, но не определена количественно Количественно не определена Взрослые (> 18 лет) Оба Оральная лейкоплакия (OLK) слюна Обнаружено и количественно определено 5.46 +/- 5,62 мкМ Взрослые (> 18 лет) Оба Болезнь с тельцами Леви подробности Моча Выявлено, но не определено количественно Не определено Взрослые (> 18 лет ) Оба Рак мочевого пузыря подробности Моча Обнаружено и количественно определено 24,545 +/- 17,404 мкмоль / ммоль креатинин Дети (1–1318 лет) Einophilic Einophilic эзофагит подробности Моча Обнаружено и количественно определено 0.75 (0,00-1,50) мкмоль / ммоль креатинина Взрослые (> 18 лет) Оба Гипербета-аланинемия подробности Моча Выявлено и количественно определено 3,0 (1,5-4,5) мкмоль / ммоль креатинин Дети (1-13 лет) Оба Гипербета-аланинемия подробности Моча Выявленная и количественная оценка креатинина 118,51 (63,24 — 247,74 мкмоль / 18 ммоль) Взрослые (> 18 лет) Оба Сахарный диабет 1 типа подробности Сопутствующие расстройства и заболевания Болезни болезни Печеночная энцефалопатия Дефицит габа-трансаминазы Туберкулезный менингит Менингит Эпилепсия Фебрильные припадки Пиридоксин-зависимая эпилепсия Hyperekplexia Синдром раздраженного кишечника Колоректальный рак Болезнь Крона Язвенный колит Применение периллилового спирта для лечения рака Рак поджелудочной железы Заболевания пародонта Лобно-височная деменция Болезнь с тельцами Леви Гипер бета-аланинемия Сахарный диабет 1 типа Эозинофильный эзофагит Связанные идентификаторы OMIM Внешние ссылки DrugBank ID DB02530 Phenol Explorer Compound ID 90DB C00001337 Chemspider ID 116 Идентификатор соединения KEGG C00334 BioCyc ID 4-AMINO -182 9018 BUTYRATE 9018 BUTYRATE 9018 BUTYRATE 9018 BUTYRATE 9018 5 Гамма-аминомасляная_кислота METLIN ID Недоступно PubChem Compound 119 PDB ID Не доступно 9018 Chology 9018 Биология Недоступно VMH ID Недоступно MarkerDB ID MDB00013416 Ссылки Synthesis Reference Minoshima.Приготовление гамма-аминомасляной кислоты из незрелых бобов. PCT Int. Прил. (2007), 25 стр. Паспорт безопасности материала (MSDS) Недоступно Общие ссылки Связывание пролина и γ-аминомасляной кислоты в стрессовых растениях посредством неферментативных реакций
Abstract
Введение
Методы
Молекулярные системы
Теоретический уровень
Результаты и обсуждение
Аминогруппа Pro является одним из наиболее предпочтительных реакционных центров для H-отщепления гидроксильным радикалом
Атака гидроксила атома N запускает декарбоксилирование Pro
Дополнительные реакции для получения частиц с закрытой оболочкой
Подключение Pro к GABA
Биологическое значение реакции Pro с
• OH в растениях при стрессе Выводы
Благодарности
Вклад авторов
Ссылки
2 [pii] pmid: 15699356 . pmid: 108 0. pmid: 16 0