ТАКОЙ РАЗНЫЙ КРАХМАЛ: АМИЛОЗА И АМИЛОПЕКТИН
Научно-практическая статья спикера II Международной фитнес-конвенции «Т..Р.И.У.М.Ф.» Софьи Кирюхиной
С каждый днем сторонников правильного питания и здорового образа жизни становится все больше. Люди стали больше обращать внимание на собственное здоровье, внешний вид. Часто сети интернет можно встретить информацию от докторов, тренеров и фитнес моделей о том, что крахмал является причиной многочисленных болезней, связанных с нарушением обмена веществ. Люди бездумно этому верят и интересуются, в каких продуктах содержится крахмал, и стараются оградить себя от его употребления. Действительно ли крахмал так вреден?
Крахмалы с древних времен являлись наиболее часто употребляемым типом углеводов и важным источником энергии для многих людей, наши предки получали из продуктов, содержащих крахмал, множество питательных веществ и микронутриентов.
Цель данной работы: проанализировать содержание крахмала в различных продуктах питания.
Крахмал состоит из молекул глюкозы и является сложным углеводом, в котором присутствуют два типа полимеров: амилоза и амилопектин, разница лишь в том, что амилопектин состоит из разветвленных молекул глюкозы  В зависимости от того, какой из полимеров преобладает крахмалы разделяют на гликемические, которые легко перевариваются, и резистентные.
В зависимости от того, какой из полимеров преобладает крахмалы разделяют на гликемические, которые легко перевариваются, и резистентные.
Гликемические крахмалы расщепляются в желудочно-кишечном тракте под действием амилазы, выделяемого поджелудочной железой и, в зависимости от скорости переваривания, рассматриваются как быстро- и медленно перевариваемые. Например, быстро и полностью перевариваемым в тонком кишечнике считается картофельное пюре, так как в нем крахмал находится в желатинизированном состоянии. Медленно, но полностью перевариваемые крахмалы содержатся, например, в макаронных изделиях, особенно в приготовленных по классическому итальянскому рецепту, то есть аль-денте, это позволяют крахмалу долго растворяться в организме, постепенно переходя в глюкозу.
Самым полезным для человеческого организма является устойчивый (резистентный) крахмал. По данным исследования: потребление резистентных крахмалов улучшает ряд физиологических показателей человека: гликемический и инсулиновый индекс, липидный состав крови, увеличивает чувство насыщения.
Подтверждение этому следующее, из 1 г устойчивых крахмалов тело способно извлечь только 2 ккал — при том, что в 1 г углеводов (а крахмалы — это углеводы) всегда около 4 ккал, все потому что углеводы расщепляются до глюкозы, а устойчивый крахмал бактерии не способны расщепить до глюкозы, он расщепляется до масляной и иных короткоцепочечных жирных кислот —он превращается не в сахар, а в очень полезный жир. Тип углеводов и крахмалов, которые мы употребляем, определяет, сколько калорий мы получаем, стоит только сравнить картофельные чипсы и вареную картошку, батончик шоколада со злаками и овсянку.
Существует четыре вида резистентного крахмала:
RS1-Крахмалы, содержащиеся в продуктах, имеющих твердую волокнистую оболочку, которые не поддаются расщеплению с помощью энзимов в ЖКТ.
|
Содержание устойчивого крахмала в продуктах |
|
|
Название |
Содержание устойчивого крахмала (г) на 100 г продукта |
|
Сырой овес |
11,8г. |
|
Овсяные хлопья, сырые |
11г. |
|
Хлопья зародышей пшеницы |
6,1г. |
|
Фасоль белая вареная |
4,0г. |
|
Нут, приготовленный |
3,0г. |
|
Чечевица, приготовленная |
3,5г. |
|
Горох, приготовленный |
3,0г. |
|
Ячмень |
2,2г. |
|
Фасоль, приготовленная |
2,0г. |
|
Консервированная фасоль или горох |
0г. |

RS2 — Крахмалы, которые ферменты пищеварительной системы человека не могут расщепить на белки, жиры и углеводы. Сюда можно отнести сырой картофель и недозрелые бананы, кукурузу, с высоким содержанием амилозы. Например, в неспелых плодах банана 4г. резистентного крахмала на 100г., который при созревании превращается в обычный.
Стоит быть аккуратнее с кукурузой, так как количество резистентного крахмала еще зависит от качества произведенный продукции и сорта. Например, в некоторых разновидностях кукурузы амилоза присутствует менее, чем на 1% (например, восковидная кукуруза).Другие виды кукурузы, напротив, содержат от 55 до 80% амилозы, но их очень редко выращивают, так как чем выше содержание амилозы, тем ниже урожайность культуры, в среднем кукуруза содержит 15-20% резистентного крахмала.
RS3- «Ретроградные» крахмалы, их особенность в том, что они при остывании меняют свою структуру и снова становятся более устойчивыми к пищеварению. Образуется в хлебе, зерновых (кукурузных хлопьях, в приготовленном и охлаждённом картофеле, холодном рисе, охлаждённых макаронах, сухариках и др.), доля резистентного крахмала в таких продуктах в незначительном количестве – не более 5%.
|
Содержание устойчивого крахмала в продуктах |
|
|
Название |
Содержание устойчивого крахмала (г) на 100 г продукта |
|
Итальянский хлеб, тост |
4,5г. |
|
Ржаной хлеб |
3,8г. |
|
Мюсли |
3,5г. |
|
Кукурузные хлопья |
3,0г |
|
Кукурузная тортилья |
2,5г. |
|
Перловая крупа |
2,3 |
|
Хлеб на закваске |
2,2г |
|
Приготовленное просо |
1,8г. |
|
Измельченная пшеница |
1,5г. |
|
Макаронные изделия из пшеницы |
1,6г. |
|
Пита, пшеничная |
1,0г. |
|
Отруби |
0,8г. |
|
Цельнозерновой хлеб |
0,7 г. |
|
Круассаны, булочки, батончик мюсли, сахарное печенье. |
<0,2 г. |

В различных сортах риса содержится от 0,5 до 4,5% резистентного крахмала, примерно 1 — 3,5г резистентного крахмала на 100г, неочищенном(буром) рисе 1,7 г. Консистенция сваренного риса и его глянец определяются в основном отношением «амилоза – амилопектин» в крахмале. Если рис после варки рассыпчатый то он содержит в себе высокое или среднее количество амилазы, устойчивого крахмала, а низкоамилозный рис – клейкий или полурассыпчатый. Чаще всего в круглозернистом рисе меньше амилозы, поэтому он слипается. Но в рисе можно повысить содержание резистентного крахмала, что доказал Судхаир Джеймс, студент шри-ланкийского Колледжа Химических Наук, для этого нужно — сварить рис с добавлением небольшого количества жира, например, кокосового масла, дать ему остыть и убрать на 12 часов в холодильник, эта процедура повышает содержание резистентного крахмала в рисе в 10 раз.
Консистенция сваренного риса и его глянец определяются в основном отношением «амилоза – амилопектин» в крахмале. Если рис после варки рассыпчатый то он содержит в себе высокое или среднее количество амилазы, устойчивого крахмала, а низкоамилозный рис – клейкий или полурассыпчатый. Чаще всего в круглозернистом рисе меньше амилозы, поэтому он слипается. Но в рисе можно повысить содержание резистентного крахмала, что доказал Судхаир Джеймс, студент шри-ланкийского Колледжа Химических Наук, для этого нужно — сварить рис с добавлением небольшого количества жира, например, кокосового масла, дать ему остыть и убрать на 12 часов в холодильник, эта процедура повышает содержание резистентного крахмала в рисе в 10 раз.
Рассмотрим картофель – один из главных источников резистентного крахмала, но только при правильном приготовлении, так, например, только что приготовленный картофель содержит в себе 1,6 грамма резистентного крахмала на 100г, а батат 1,8 г. Однако, приготовление и последующее охлаждение картофеля или батата приводит к почти двукратному увеличению резистентного крахмала, что составляет 5 грамм на 100 грамм продукта, почти не теряется крахмал в картофеле в процессе жарки, а вот, как отмечалось ранее, в картофельном пюре резистентного крахмала почти нет, он превращается в гликимический.
|
Содержание устойчивого крахмала в картофеле и батате |
|
|
Название |
Содержание устойчивого крахмала (г) на 100 г продукта |
|
Вареный картофель |
1,6г. |
|
Вареный батат |
1,8г. |
|
Вареный и охлажденный картофель или батат |
5,0г. |
|
Картофельные чипсы |
3,5г. |
|
Картофельное пюре |
0 г. |
Чтобы получить максимальный эффект от устойчивых крахмалов — надо потреблять их по возможности в наименее обработанном виде, нагревание разрушает сложные крахмалы, поэтому ряд народных советов приобретают значение: замачивание или проращивание круп значительно укорачивает время их приготовления и сохраниет в целосности сложные углеводы. Только правильное приготовление и употребление продуктов, содержащих резистентный крахмал, убивает в кишечнике предраковые клетки», — говорит Джанин Хиггинс, кандидат наук, исследователь Онкологического центра при Университете Колорадо. (Правильное употребление – это при комнатной температуре или ниже).
Только правильное приготовление и употребление продуктов, содержащих резистентный крахмал, убивает в кишечнике предраковые клетки», — говорит Джанин Хиггинс, кандидат наук, исследователь Онкологического центра при Университете Колорадо. (Правильное употребление – это при комнатной температуре или ниже).
Существует еще один вид резистентного крахмала RS4, который не встречается в природе, а производится промышленным способом.
В современном мире продукты все чаще перерабатывают, так сейчас из различных крахмалосодержащих продуктов получают крахмал с высоким содержанием амилозы или амилопектина. Например, крахмал зерновых культур может содержать от 15% до 28% амилозы, крахмал кукурузы на 75–80% состоит из амилопектина и лишь 20–25% из амилозы, крахмалы клубневых (картофель, например) содержат 17 — 22% амилозы, тогда как в крахмалах бобовых (чечевица, фасоль, нут…) её гораздо больше — от 33 до 66%.
В процессе приготовления хлебобулочных изделий используют чаще всего пшеничную муку высшего сорта для повышения качества мучных изделий, такая мука состоит почти из одного амилопектина, а значит, является «быстрым углеводом». Исследования, проведенные в Белгородском научно-исследовательском институте сельского хозяйства Россельхозакадемии и Белгородском государственном национальном исследовательском университете, установили, что готовая продукция из муки пшеницы с низким содержанием амилозы устойчива к черствению. Часто даже используют муку пшеницы вакси, особенностью которой является отсутствие в составе ее крахмала амилозы, что увеличивает сроков сохранения свежести продуктов. Особые сорта зерновых и кукурузы (waxy, восковые), а также ячменя (шведская компания Swecarb AB, торговая марка Vitargo) содержащие только амилопектин требуемого размера. Так же очень много подобного крахмала содержат в себе каши быстрого приготовления и манка.
Исследования, проведенные в Белгородском научно-исследовательском институте сельского хозяйства Россельхозакадемии и Белгородском государственном национальном исследовательском университете, установили, что готовая продукция из муки пшеницы с низким содержанием амилозы устойчива к черствению. Часто даже используют муку пшеницы вакси, особенностью которой является отсутствие в составе ее крахмала амилозы, что увеличивает сроков сохранения свежести продуктов. Особые сорта зерновых и кукурузы (waxy, восковые), а также ячменя (шведская компания Swecarb AB, торговая марка Vitargo) содержащие только амилопектин требуемого размера. Так же очень много подобного крахмала содержат в себе каши быстрого приготовления и манка.
То же касается и киселя, который изготавливается из крахмала, чаще всего картофельного или кукурузного. Крахмал в процессе приготовления киселя желатинируется, если это происходит сильнее, значит содержание амилозы ниже и тем лучше он может быть расщеплен альфа-амилазами-пищеварительными фирментами, т. е. фактически при нагревании крахмала мы получаем чистую глюкозу.
е. фактически при нагревании крахмала мы получаем чистую глюкозу.
Но настоящую опасность таит в себе рафинированный и модифицированный крахмал. Он не даст организму ничего, из чего тот мог бы извлечь выгоду, именно в таком веществе наблюдается отсутствие пищевых волокон, так необходимых организму, и слишком малое количество полезных веществ, но он дает человеческому организму энергию и не перегружает ЖКТ. Модифицированные крахмалы нашли применение и в различных отраслях пищевой промышленности. Например, при производстве детского питания используют модифицированный крахмал от 3 до 6%: рисовый, кукурузный или картофельный, его изменяют для того, чтобы такой крахмал быстрее расщепился до глюкозы, и не вызывал у ребенка проблемы с пищеварением.
В России в производстве мясных продуктов используются нативные крахмалы: картофельный, пшеничный и импортный тапиоковый, структура таких крахмалов включает преимущественно полисахарид – ветвистый амилопектин, который быстро расщепляется и повышает уровень глюкозы в крови.
В спортивном питании для разных целей используют крахмал. Углвеводные напитки делают именно из амилопектина, дабы не «грузить» ЖКТ и дать возможность быстрейшему поступлению углеводов в кровоток на тренировке, такие углеводы на 70% быстрее восполняют потери гликогена (Скандинавский журнал Гастроэнтерологии (Scand. J. Gastroenterol 2000;35:1143-1149.) и Европейский журнал Прикладной Физиологии (Eur J Appl Physiol 81:346-351.) В диетологии же особенность более долгого переваривания полимеров амилозы используют наоборот для придания углеводам «долготы», т.е. более пролонгированного переваривания. Есть ухищрения эту «долготу» еще и удлинять, воздействую химическим способом. К примеру, разработан такой крахмал со сложно проговариваемым названием Гидроксипропил дистракционый фосфат, сокращенно HDP-крахмал.
В современном мире наблюдается тенденция к уменьшению употребления устойчивых крахмалов. Это объясняется широким распространением продуктов промышленной переработки, которые содержат существенно меньше данного крахмала. Люди едят чипсы вместо картофеля, овсяные батончики вместо овсяных хлопьев, бургеры вместо мяса. Минимальная норма резистентного крахмала составляет от 10грамм в сутки, но она может доходить до 50 г. Добавив в свой рацион устойчивый крахмал, уже через четыре недели можно увидеть, что чувствительность к инсулину увеличивается на 33-50 процентов( Nutrients. 2013).Устойчивые крахмалы смягчают состояние при запорах, синдроме возбудимого кишечника, язвенном колите. Исследование австралийских ученых показало, что неперевариваемый крахмал может помочь компенсировать эффект от частого употребления красного мяса, повышающего риск развития рака кишечника Результаты проведенной работы опубликованы в журнале Cancer Prevention Research). Устойчивые крахмалы помогают чувствовать сытость.
Люди едят чипсы вместо картофеля, овсяные батончики вместо овсяных хлопьев, бургеры вместо мяса. Минимальная норма резистентного крахмала составляет от 10грамм в сутки, но она может доходить до 50 г. Добавив в свой рацион устойчивый крахмал, уже через четыре недели можно увидеть, что чувствительность к инсулину увеличивается на 33-50 процентов( Nutrients. 2013).Устойчивые крахмалы смягчают состояние при запорах, синдроме возбудимого кишечника, язвенном колите. Исследование австралийских ученых показало, что неперевариваемый крахмал может помочь компенсировать эффект от частого употребления красного мяса, повышающего риск развития рака кишечника Результаты проведенной работы опубликованы в журнале Cancer Prevention Research). Устойчивые крахмалы помогают чувствовать сытость.
Подводя итог, хочется сказать, что устойчивый крахмал необходим нашему организму, так как он обладает многими полезными свойствами, поэтому каждому, кто заботиться о своем здоровье необходимо добавить в свой рацион: бобовые, зерновые, картофель, неспелые бананы и другие продукты, с высоким содержанием амилозы и ограничить употребление обработанных продуктов, например, злаковых батончиков. Особое внимание следует уделить процессу приготовления крахмалосодержащих продуктов, для того чтобы в процессе обработки не потерять самый ценный для нашего организма резистентный крахмал.
Особое внимание следует уделить процессу приготовления крахмалосодержащих продуктов, для того чтобы в процессе обработки не потерять самый ценный для нашего организма резистентный крахмал.
Если заметили ошибку, выделите фрагмент текста и нажмите Ctrl+Enter
Картофельный Союз | Крахмал: насколько обоснован страх поправиться?
Мы привыкли, что окружающие говорят о вреде крахмала, в связи с этим мы начинаем обходить стороной продукты, которые содержат этот компонент. А вы задумывались, что нас так пугает? Давайте поговорим подробнее об этом компоненте и узнаем, есть ли основания его бояться или же нужно дать ему зеленый свет.Следует сказать, что крахмал является самым распространенным углеводом, который употребляется вместе с пищей. Есть несколько видов крахмалов, которые оказывают разные действия на наш организм. Одни быстро расщепляются, перевариваются в тонкой кишке, и глюкоза достаточно быстро всасывается в кровь. Есть также устойчивый крахмал, который наоборот медленно расщепляется в организме и доходит до толстой кишки, там такой крахмал служит отличным источником пищи для бактерий. Именно благодаря этому крахмалу человек чувствует насыщение после употребления. Только вот потребление продуктов с высоким содержанием крахмала влияет на поджелудочную железу и уровень инсулина, который становится немного ниже. Поэтому, если держать диету, в которой мы будете употреблять продукты с таким крахмалом нет гарантии, что инсулин в крови будет находится в норме постоянно. А значит организм начинает скапливать в себе меньше жира, если человек будет употреблять продукты, у который очень высокий гликемический индекс или продукты, которые содержат легкоусваеваемый крахмал. Поэтому организм начинает вырабатывать больше инсулина для того, чтобы снизить содержание сахара в крови. Только мало кто знает, что именно устойчивый крахмал является полезным для здоровья человека. Благодаря ему улучшается чувствительность к инсулину, уровень сахара и холестерина становятся ниже, а также аппетит становится умеренным. Согласитесь, интересное открытие. Потому что все незнающие твердят о том, что любой крахмал вреден для организма.
Именно благодаря этому крахмалу человек чувствует насыщение после употребления. Только вот потребление продуктов с высоким содержанием крахмала влияет на поджелудочную железу и уровень инсулина, который становится немного ниже. Поэтому, если держать диету, в которой мы будете употреблять продукты с таким крахмалом нет гарантии, что инсулин в крови будет находится в норме постоянно. А значит организм начинает скапливать в себе меньше жира, если человек будет употреблять продукты, у который очень высокий гликемический индекс или продукты, которые содержат легкоусваеваемый крахмал. Поэтому организм начинает вырабатывать больше инсулина для того, чтобы снизить содержание сахара в крови. Только мало кто знает, что именно устойчивый крахмал является полезным для здоровья человека. Благодаря ему улучшается чувствительность к инсулину, уровень сахара и холестерина становятся ниже, а также аппетит становится умеренным. Согласитесь, интересное открытие. Потому что все незнающие твердят о том, что любой крахмал вреден для организма.
Давайте тогда узнаем, в каких продуктах содержится полезный крахмал? Нужно сказать, что устойчивый крахмал, его еще называют резистентным, делится на четыре вида:
Первый содержится в зерновых, бобовых и семенах;
Второй — в сыром картофеле, зеленых бананах;
Третий — содержится в охлажденном рисе и картофеле;
Четвертый, полученный в результате химической реакции.
Но некоторые виды крахмала могут встречаться в одном продукте. Так, в перезревших бананах полезный крахмал превращается в обычный. А еще количество этого вещества в продукте зависит от способа его приготовления.
А где содержится высокая доза крахмала? Резистентный крахмал в большом количество содержится в фасоли и чечевице. Дальше по количеству содержания идут цельные злаки. Как правило, это различные крупы, гречка, рис и овес. Картофель также является продуктом, о котором знают все и о том, что он богат крахмалом. Небольшое содержание крахмала есть во многих фруктах и овощах, главное, чтобы вы сами понимали, что всему есть своя мера, потому что в первую очередь крахмал — это углеводы.
Кстати, немного о картофеле. Количество полезного крахмала зависит напрямую от того, каким способом вы готовите еду. Если вы приготовили картошку, а после ее охладили, что количество крахмала увеличивается в два раза. Продукты, которые богаты крахмалом не стоит готовить с помощью жарки, потому начинает образовываться химическое вещество — акриламид. Это вещество может быть вредным для человека. Крахмалистые продукты лучше всего сочетать с сырыми овощами.
Поэтому, можно сделать вывод, что крахмала не стоит бояться, нужно лишь выбирать правильный, а это зависит от продуктов!
Источник: http://www.medpulse.ru/health/yourshealth/spec/378586.html
Комментариев: 0 |
Write a comment
- Required fields are marked with *.
Ученые назвали продукты, которые нельзя хранить в холодильнике
Специалисты в области правильного питания назвали продукты, которые категорически нельзя хранить в холодильнике, в основном из-за их быстрой порчи и потери вкусовых и полезных качеств. Вот примеры продуктов, которые журнал My Health Reads советуют не класть в холодильник.
Вот примеры продуктов, которые журнал My Health Reads советуют не класть в холодильник.
Хранение картофеля в холодильнике негативно влияет на него. Например, меняется его вкус, при замораживании крахмал превращается в сахар, и картофель становится бесцветным. Таким образом, вместо того, чтобы замораживать картофель, вы можете хранить его разными способами. Во-первых, держите клубни в бумажных пакетах. Только убедитесь, что вы не используете полиэтилен, так как он ускоряет разложение картофеля. Как вариант, храните их немытыми и вдали от прямых солнечных лучей. Вы также должны держать их подальше от экстремальных температур. Лучше всего использовать мешковину или кладовую. Применительно к таким продуктам, как ягоды, учитываются различные факторы, включая продолжительность хранения ягод, а также продолжительность их употребления. Если вы не собираетесь есть эти ягоды в ближайшее время, лучше не хранить их в холодильнике. Помните, очень важно то, как вы храните ягоды даже вне холодильника. Следует быть осторожным и хорошо промыть ягоды, чтобы не образовалась плесень.
Следует быть осторожным и хорошо промыть ягоды, чтобы не образовалась плесень.
Чеснок ни в коем случае нельзя хранить в холодильнике, потому что он быстрее разлагается в условиях повышенной влажности. Однако, если вы очистите чеснок от кожуры, он будет хорошо сохраняться. Но, как правило, из-за влажности чеснок теряет аромат. Это также ведет к росту плесени. Чтобы чеснок был в хорошем состоянии, храните его в прохладном, сухом месте, вдали от света. Лук также необходимо хранить в прохладном месте, вдали от света, а не в холодильнике. Лук содержит крахмал, и когда вы храните его в местах с большим содержанием влаги, он становится влажным. Это способствует образованию плесени. Вы должны хранить лук отдельно от картофеля, потому что их совместное хранение может привести к выделению газов, которые способствуют гниению лука. Вместо этого храните их в сетчатом мешке.
Эксперты также предостерегают от того, чтобы класть оливковое масло в холодильник. Поскольку холодильник влажный и в нем образуется конденсат, оливковое масло легко может потерять свой естественный вкус. Эта среда также может сделать оливковое масло мутным. Держите его подальше от солнечных лучей и храните в шкафу или другом прохладном и сухом месте, советуют специалисты. Еще один крайне восприимчивый продукт к холоду — это мед. Каждый раз, когда вы подвергаете мед воздействию низких температур, он затвердевает. Затем мед кристаллизуется и становится очень комковатым. Из-за комковатости мед будет трудно использовать. Поэтому правильно хранить мед в кладовой.
Эта среда также может сделать оливковое масло мутным. Держите его подальше от солнечных лучей и храните в шкафу или другом прохладном и сухом месте, советуют специалисты. Еще один крайне восприимчивый продукт к холоду — это мед. Каждый раз, когда вы подвергаете мед воздействию низких температур, он затвердевает. Затем мед кристаллизуется и становится очень комковатым. Из-за комковатости мед будет трудно использовать. Поэтому правильно хранить мед в кладовой.
Уксус — один из продуктов, которые производятся с очень длительным сроком хранения. Таким образом, вам не нужно хранить уксус в холодильнике. Вместо этого храните его в кладовой или в сухом прохладном месте, например, в буфете. Также не рекомендуется хранить хлеб в холодильнике. Хлеб насыщен крахмалом, поэтому, когда вы кладете его в холодильник, он становится сухим и портится быстрее. Если вы храните его в холодильнике в течение длительного периода времени, вы, скорее всего, обнаружите плесень на хлебе. Когда вы храните хлеб при комнатной температуре, он будет оставаться в хорошем состоянии в течение значительного времени. Когда тыква хранится в холодных условиях, она гораздо быстрее повреждается. Поэтому не храните тыквы в холодильниках. Любая среда с температурой ниже 10 градусов не подходит для тыквы.
Когда тыква хранится в холодных условиях, она гораздо быстрее повреждается. Поэтому не храните тыквы в холодильниках. Любая среда с температурой ниже 10 градусов не подходит для тыквы.
Также эксперты не советуют хранить в холодном месте специи, арахисовое масло, авокадо, который лучше порезать на мелкие кусочки, орехи и сухофрукты, а также заправку для салатов, персики, мяту, петрушку, укроп и базилик.
От чего зависят значения гликемических индексов?
От чего зависят значения гликемических индексов?
Большая часть углеводсодержащих продуктов, входящих в рацион современного человека, представляют собой сложные углеводы, преимущественно состоящие из крахмала. Такого рода продукты можно разделить на четыре семейства:
|
Семейства продуктов, преимущественно |
|||
| Злаки | Клубни | Бобовые | Фрукты |
|
Пшеница твердых и мягких сортов Рис Кукуруза Овёс Ячмень Рожь Сорго Просо (пшено) |
Картофель Батат (сладкий картофель) Маниок Ямс Таро (колоказия) Маланга |
Фасоль Горох Нут (турецкий горох) Чечевица Бобы |
Банан Манго Яблоко |
Для того чтобы крахмал, входящий в состав этих продуктов, был поглощен стенками кишечника и поступил в кровь, он предварительно должен быть преобразован в глюкозу. Расщепление осуществляется благодаря пищеварительным ферментам, в частности, амилазе. Переваривание крахмалов начинается в ротовой полости с пережевывания и смачивания слюной и продолжается в тонкой кишке после их прохождения через желудок.
Расщепление осуществляется благодаря пищеварительным ферментам, в частности, амилазе. Переваривание крахмалов начинается в ротовой полости с пережевывания и смачивания слюной и продолжается в тонкой кишке после их прохождения через желудок.
Величина гипергликемии свидетельствует об уровне всасывания глюкозы и, следовательно, о степени усвояемости крахмала.
Подробнее о переваривании и всасывании пищи
Гликемические индексы отражают величину повышения гликемии после переваривания определенного продукта. Многочисленные наблюдения показали, что при одинаковом количестве чистого углевода в разных продуктах, уровень гипергликемии после их употребления может существенно различаться. Большая или меньшая степень всасывания углеводсодержащей пищи связана с тем, что определённая часть крахмала не поддаётся расщеплению.
На степень всасываемости крахмалов, выраженную в гликемических индексах, влияет ряд факторов.
Подробнее о том, что обозначают гликемические индексы
Структура крахмала:
Зерно крахмала состоит из молекулярных соединений двух типов: амилозы и амилопектина. Они могут сопровождаться небольшим количеством неуглеводных компонентов: липидами, протеинами, пищевыми волокнами, а также микроэлементами (витаминами, минеральными солями…).
Они могут сопровождаться небольшим количеством неуглеводных компонентов: липидами, протеинами, пищевыми волокнами, а также микроэлементами (витаминами, минеральными солями…).
Физико-химическая природа крахмалсодержащих продуктов и их влияние на организм человека определяются в основном количественным соотношением между амилозой и амилопектином, присутствующими в этих продуктах.
Соотношение амилоза/амилопектин может существенно меняться как от одного семейства к другому, так и от одной разновидности к другой внутри одного семейства.
Крахмал зерновых культур может содержать от 15% до 28% амилозы. Но в некоторых разновидностях кукурузы амилоза присутствует менее, чем на 1% (например, восковидная кукуруза, экстракт которой используется в пищевой промышленности в качестве загустителя).
Другие виды кукурузы, напротив, содержат от 55 до 80% амилозы, но их очень редко выращивают, так как чем выше содержание амилозы, тем ниже урожайность культуры.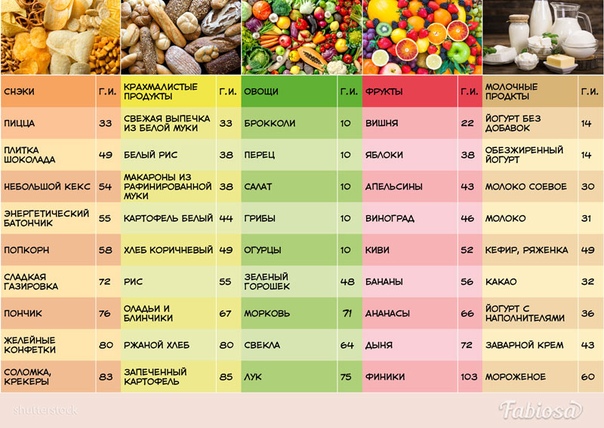
Крахмалы клубневых (картофель, например) содержат 17 — 22% амилозы, тогда как в крахмалах бобовых (чечевица, фасоль, нут…) её гораздо больше — от 33 до 66%.
Изменения гликемического индекса
Гликемический индекс крахмалсодержащего продукта зависит от следующих параметров:
- Соотношение амилоза/амилопектин
При нагревании с водой структура крахмала изменяется. Зерна крахмала, впитывая воду, постепенно набухают, и часть амилопектина переходит в раствор. По мере дальнейшего нагревания в раствор переходит и часть амилозы.
Результатом является более или менее значительная вязкость взвеси. Этот феномен называется желатинированием крахмала.
Чем меньше количество амилозы в продукте, тем более интенсивно желатинирование, и наоборот. Было доказано, что чем более желатинируется крахмал (вследствие низкого содержания амилозы), тем легче он гидролизуется под действием амилаз (пищеварительных ферментов, расщепляющих крахмал в организме человека), тем большая его часть преобразуется в глюкозу, и тем значительнее повышается гликемия.

Другими словами, чем меньше амилозы в крахмалсодержащем продукте, тем выше его гликемический индекс. И наоборот, чем больше амилозы содержит продукт, тем меньше степень желатинирования, тем меньше глюкозы образуется при расщеплении и тем ниже гликемический индекс.
Так, у картофеля, содержащего малое количество амилозы, гликемический индекс высокий, а у чечевицы – низкий из-за значительного количества амилозы, входящей в её состав.
Довольно интересен пример кукурузы. Восковидная кукуруза (Waxy), практически лишенная амилозы, широко культивируется именно из-за высокой вязкости её крахмала. Он обычно используется в качестве загустителя различных фруктовых желе, а также для придания желаемой консистенции консервам и замороженным продуктам. На упаковках в разделе «состав» он так и называется: «кукурузный крахмал».
Входя в состав многих индустриальных блюд, кукурузный крахмал, имеющий очень высокий гликемический индекс (близкий к 100) существенно повышает гликемию человека.
Любопытный факт: в Австралии на одном из хлебопекарных предприятий для снижения гликемического индекса хлеба в тесто был добавлен крахмал кукурузы с высоким содержанием амилозы (более 80%). Получившийся хлеб очень полюбился покупателям, особенно детям, которые, как правило, отказываются от хлеба из муки грубого помола.

- Тип механической и термической обработки, которой подвергается продукт питания
Нагревание в водной среде повышает гликемический индекс продукта. Например, морковь в сыром виде имеет ГИ 20, а после варки её гликемический индекс поднимается до 50 из-за желатинирования содержащегося в ней крахмала.
Некоторые виды промышленной обработки максимизируют желатинирование продуктов. Это относиться, например, к изготовлению различных хлопьев (картофельное пюре быстрого приготовления, кукурузные хлопья для завтрака…) и связующих веществ, таких как модифицированные крахмалы и декстрины.
В результате подобных операций гликемический индекс продуктов значительно повышается (85 для кукурузных хлопьев, 95 для пюре из картофельных хлопьев, 100 для модифицированных крахмалов).
По такому же принципу, при изготовлении попкорна разрыв кукурузного зерна (или рисового зерна для получения воздушного риса) приводит к 15-20-процентному повышению гликемического индекса исходного продукта.«Пастификация» снижает гликемический индекс
Итак, из одной и той же муки можно получить продукты с разными гликемическими индексами (пельмени/вареники — 70, спагетти — 40).
Однако существует и такая обработка продуктов, которая замедляет гидратацию крахмала. Это процесс «пастификации» твёрдых сортов пшеницы. Экструзия (вытягивание) теста через фильеру (формующий элемент в матрице) приводит к нагреванию, результатом которого является образование защитной плёнки, впоследствии замедляющей желатинирование крахмалов во время варки.
Это относится к спагетти и некоторым видам лапши, полученным путём «пастификации», то есть экструзии под высоким давлением, но не касается ни пельменей, ни теста для лазаньи, ни свежей лапши, приготовленной вручную, несмотря на то, что все эти продукты также делаются из муки твёрдых сортов пшеницы.
Но это ещё не всё. Способ приготовления этих продуктов в домашних условиях непосредственно перед их употреблением в пищу также влияет на их гликемический индекс.
Немного недоваренные, слегка похрустывающие на зубах спагетти (после 5-6 мин. варки) будут иметь самый низкий возможный для них гликемический индекс. Это так называемые спагетти аль денте (al dente), что в буквальном переводе с итальянского значит «на зуб». Продолжительная варка (15-20 мин.) ускорит желатинирование крахмала, из-за чего гликемический индекс спагетти неизбежно повысится.


- Ретроградация – процесс обратный желатинированию
Пройдя термическую обработку (варку, жарку…), приводящую к его желатинированию, крахмал при остывании подвергается новым изменениям.
Находящиеся в желеобразном состоянии макромолекулы амилозы и амилопектина вновь постепенно перестраиваются.
Длительное хранение крахмалсодержащих продуктов (готовые блюда в вакууме) при низкой температуре (5°С) благоприятствует ретроградации. Высушивание некоторых продуктов имеет тот же эффект. Например, чем больше черствеет хлеб, тем больше влаги он теряет и тем большей ретроградации подвергается входящий в него крахмал. То же самое произойдет, если подсушить хлеб в духовке или тостере.Ретроградация, хотя и не ведёт к полной обратимости желатинирования, всё-таки позволяет существенно снизить гликемический индекс. Так спагетти (даже из белой муки), приготовленные аль денте и употребляемые в охлажденном виде в салате, будут иметь гликемический индекс 35.
Сделанный из той же белой муки хлеб, в зависимости от того, свежий он (и ещё тёплый), чёрствый или подсушенный в тостере, будет иметь разный гликемический индекс.
Следуя этой логике, можно считать, что замораживание свежего хлеба и его последующее размораживание при комнатной температуре ощутимо снижают его исходный гликемический индекс.
Интересно, что охлаждённая зелёная чечевица (и тем более после 24 часов в холодильнике) имеет ещё более низкий гликемический индекс, чем только что сваренная (ГИ между 10 и 15). Это связано с тем, что чем больше амилозы содержится в исходном крахмале, тем эффективнее проходит процесс ретроградации.
В то же время было доказано, что добавление липидов к крахмалу, подвергающемуся желатинированию, замедляет последующую ретроградацию.
Заметим, что при нагревании крахмал, ранее подвергшийся ретроградации, частично теряет свою способность к последующему желатинированию. Часть такого крахмала (примерно 10%) приобретает тепловую устойчивость. Так что разогревание углеводсодержащего продукта после его хранения в холодильнике также снижает его гликемический индекс.
Необходимо также сказать, что крахмал в своей природной форме (сырой и необработанный) встречается не только в сырых продуктах. Иногда он может сохранять свою первозданную структуру даже при термической обработке. Это происходит из-за отсутствия достаточного для желатинирования количества влаги. Например, присутствующий в хлебной корочке и песочном печенье крахмал частично сохраняет зернистость своей структуры даже после выпечки, и это снижает его гликемический индекс относительно желатинированного крахмала (например, в хлебном мякише).
Так, при тушении или варке на пару доступ влаги к продукту ограничен, что объясняет меньшую степень желатинирования входящих в него крахмалов, в сравнении с варкой в воде.
 Так происходит ретроградация крахмала – то есть возвращение (более или менее значительное) к предшествующей желатинированию молекулярной структуре. Ретроградация прогрессирует с течением времени и по мере понижения температуры.
Так происходит ретроградация крахмала – то есть возвращение (более или менее значительное) к предшествующей желатинированию молекулярной структуре. Ретроградация прогрессирует с течением времени и по мере понижения температуры.

- Присутствие в продукте протеинов и пищевых волокон
Естественно присутствующие в некоторых углеводсодержащих продуктах (например, в злаковых) протеины могут уменьшать гидролиз (расщепление) крахмалов и вследствие этого снижать гликемический индекс продукта.
Хорошим примером этого феномена служат макароны. Присутствующая в них клейковина (глютен) замедляет действие пищеварительных ферментов, что в свою очередь ограничивает всасывание глюкозы.
Рафинирование современных злаков уменьшает и без того низкое содержание в них клейковины, что существенно повышает гликемию человека, употребляющего их в пищу.
С другой стороны, присутствие в крахмалсодержащем продукте пищевых волокон может ограничивать действие на него пищеварительных ферментов (амилаз) и таким образом уменьшать всасывание глюкозы. Особенно эффективна в этом смысле растворимая клетчатка, встречающаяся, например, в бобовых и овсе. Пищевые волокна являются прямым или косвенным барьером всасыванию глюкозы и благодаря этому снижают гликемический индекс данного крахмала.
Подробнее о переваривании и всасывании пищи

- Степень зрелости и длительность хранения
Гликемический индекс крахмалсодержащих фруктов повышается по мере их созревания.
Для полноты картины, добавим, что длительность хранения некоторых продуктов влияет на их гликемический индекс по причине естественной трансформации входящих в них крахмалов. Так ГИ картофеля, хранимого в течение нескольких месяцев, выше, чем ГИ свежего картофеля.
 Эти изменения более или менее значительны в зависимости от фрукта. Например, ГИ банана повышается гораздо больше, чем ГИ яблока. Зелёный банан имеет низкий гликемический индекс (приблизительно 40), но по мере созревания содержащийся в нём крахмал становится всё менее устойчивым, поэтому гликемический индекс зрелого банана вырастает до 65. Тот же феномен наблюдается при термической обработке банана каким бы то ни было способом.
Эти изменения более или менее значительны в зависимости от фрукта. Например, ГИ банана повышается гораздо больше, чем ГИ яблока. Зелёный банан имеет низкий гликемический индекс (приблизительно 40), но по мере созревания содержащийся в нём крахмал становится всё менее устойчивым, поэтому гликемический индекс зрелого банана вырастает до 65. Тот же феномен наблюдается при термической обработке банана каким бы то ни было способом.-
Размер частиц крахмала
При измельчении крахмалсодержащего продукта, чем меньше становятся частицы крахмала, тем проще он будет расщепляться впоследствии, что говорит о повышении его гликемического индекса.
Это в частности относится к злакам, превращаемым в муку.
Так рисовая мука имеет более высокий ГИ, нежели рис, из которого она делается.
В прошлом зёрна пшеницы размельчались между каменными мельничными жерновами, так что частицы муки получались достаточно крупными. Просеивание было нетщательным, что давало муку довольно грубого помола коричневато-серого цвета (пеклеванную). Так называемый тогда «белый» хлеб имел ГИ между 60 и 65, что в принципе ещё допустимо. Одним из редких хлебов, соответствующих сегодня этим стандартам, является знаменитый хлеб Пуалан (фр. Poilâne). Он полезен ещё и тем, что делается исключительно из натуральной хлебной закваски, что способствует снижению гликемического индекса хлеба.
Хлеб для простолюдинов раньше делался из непросеянной муки грубого помола, которая сохраняла все составляющие пшеничного зерна, поэтому этот хлеб назывался «цельнозерновым». Частицы муки были довольно крупными, в ней сохранялись все пищевые волокна и протеины, и кроме того хлеб делался на закваске – поэтому его гликемический индекс был низким (между 35 и 40).
Цельнозерновой хлеб Монтиньяка, который можно приобрести в частности в сети булочных «Премьер Муассон» («Première Moisson») в Квебеке, соответствует именно этому стандарту хлеба.Подробнее о цельнозерновом хлебе Монтиньяка в Квебеке

 Цельнозерновой хлеб Монтиньяка, который можно приобрести в частности в сети булочных «Премьер Муассон» («Première Moisson») в Квебеке, соответствует именно этому стандарту хлеба.
Цельнозерновой хлеб Монтиньяка, который можно приобрести в частности в сети булочных «Премьер Муассон» («Première Moisson») в Квебеке, соответствует именно этому стандарту хлеба.| Питательные вещества | Цельнозерновая мука на 100гр. | Белая мука (T55) на 100 |
| Протеины |
12 гр. |
8 гр. |
| Липиды |
2.5 гр. |
1 гр. |
| Углеводы |
60 гр. |
74 гр. |
| Пищевые волокна |
10 гр. |
3 гр. |
| Вода |
15.5 гр. |
14 гр. |
| Размер частиц |
Крупные |
Мелкие |
| Гликемический индекс |
40 |
70 |

После изобретения в 1870 году вальцовой мельницы изготовление белой муки начинается повсеместно, сначала на западе, а позже во всех странах мира. Распространение этой технической новинки, принимавшейся, конечно, за шаг вперёд, приводит к обеднению хлеба питательными веществами.
По мере дальнейшего усовершенствования мельничного оборудования, мука становится все более рафинированной. В плане питательности это выражается в следующем: в состав муки входит всё меньше пищевых волокон, протеинов и микроэлементов (витаминов, минералов, основных жирных кислот…), и всё более мелкими становятся её частицы. Этим объясняется повышение гликемического индекса тех продуктов, основой которых является такая в высшей степени рафинированная мука.
В плане питательности это выражается в следующем: в состав муки входит всё меньше пищевых волокон, протеинов и микроэлементов (витаминов, минералов, основных жирных кислот…), и всё более мелкими становятся её частицы. Этим объясняется повышение гликемического индекса тех продуктов, основой которых является такая в высшей степени рафинированная мука.
Заключение
Питательные свойства углеводов могут значительно изменяться под действием определённых факторов.
Нельзя забывать, что существует не один, но множество различных видов крахмалов.
Крахмалы отличаются по своей изначальной молекулярной структуре (соотношение амилоза/амилопектин), а также по природе и свойствам других питательных веществ (белки, пищевые волокна), входящих в их состав.
Физико-химические характеристики крахмалов постоянно изменяются под влиянием воды, времени (длительности хранения) и температурных перепадов.
Любая гидротермическая обработка, будь она промышленной или кулинарной, ведёт к преобразованиям внутри продукта, влияющим на его специфические свойства и удобоваримость.
Эти изменения воздействуют на процесс всасывания, за которым следует соответствующий гликемический, а затем и инсулиновый ответ организма.
Гликемический индекс продукта – это результат многочисленных параметров, которые необходимо учитывать в нашем повседневном выборе продуктов питания и способов их приготовления.
Пренебрегая этими научными понятиями, известными уже с конца 80-х годов ХХ столетия, традиционная диетология позволила аграрно-пищевой промышленности не только беспрепятственно вывести подозрительные разновидности растений, но и ввести в практику такие способы обработки и консервации продуктов, которые косвенно способствуют слишком большому повышению гликемии у современного человека.
Гиперинсулинизм, развивающийся в результате регулярного потребления таких продуктов, лежит в основе многочисленных сердечно-сосудистых недугов, а также участившихся заболеваний ожирением, диабетом.
Теперь очевидно, насколько неосторожным со стороны современных диетологов является утверждать, что углеводы должны составлять 50-55 процентов повседневного рациона современного человека, и при этом не уточнять, о каких именно углеводах идёт речь. Хуже того: если и делается попытка объяснить, какие продукты следует выбирать, то при этом обычно цитируется абсолютно ошибочная классификация углеводов на «быстрые» и «медленные сахара».
Подробнее о необоснованности понятий «быстрых» и «медленных сахаров»
По словам профессора Уалтера Уиллетта (Walter WILLETT) из Медицинской Школы Гарварда, эти рекомендации никогда не сопровождаются дополнительными объяснениями относительно выбора и обработки (приготовления, хранения…) содержащих углеводы продуктов с учётом их гликемического индекса.
Максимум, что может быть «официально» рекомендовано, — употреблять в пищу преимущественно сложные углеводы, что практически ничего не значит с точки зрения современных знаний о питании.
Как справедливо отмечают учёный Ф. Борне (F. Bornet) и профессор Ж. Слама (G. Slama), кроме того, что «сложные углеводы не взаимозаменяемы», как считалось ранее, нужно учитывать и то, что «некоторые крахмалы и крахмалсодержащие продукты, несмотря на сложность их структуры, вызывают гораздо более сильную гипергликемию, чем простые сахара». Например, употребление жареного картофеля (ГИ 95) провоцирует гораздо более высокую гликемию, чем употребление сахара (ГИ 70).
Мишель Монтиньяк – первый в мире нутриционист, предложивший использовать гликемические индексы продуктов питания для регулирования веса. Уже с конца 80-х годов ХХ века в своих публикациях он объяснял, как изменение привычек питания привело к значительному учащению случаев заболевания ожирением во всем мире.
Человек постепенно перешёл от пищи, вызывающей несущественное повышение гликемии, к пище, состоящей в основном из углеводов с высоким гликемическим индексом. Результат этой перемены — повсеместно наблюдаемые сегодня патологии обмена веществ, и в частности гиперинсулинизм — фактор появления лишнего веса и развития диабета.
Подробнее о Мишеле Монтиньяке и первом применении гликемических индексов
Бутерброды и пельмени — нельзя? Названы несовместимые сочетания продуктов
https://rsport.ria.ru/20210505/pitanie-1731226460.html
Бутерброды и пельмени — нельзя? Названы несовместимые сочетания продуктов
Бутерброды и пельмени — нельзя? Названы несовместимые сочетания продуктов — Спорт РИА Новости, 05.05.2021
Бутерброды и пельмени — нельзя? Названы несовместимые сочетания продуктов
Употребление комбинаций некоторых продуктов может привести к вздутию и несварению. Диетолог Елена Кален в своем Instagram назвала неудачные сочетания и раскрыла Спорт РИА Новости, 05.05.2021
2021-05-05T19:43
2021-05-05T19:43
2021-05-05T19:43
зож
здоровье
питание
здоровый образ жизни (зож)
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
Россиянам назвали несовместимые сочетания продуктов
https://cdn24.img.ria.ru/images/07e5/05/05/1731234303_0:98:1920:1178_1920x0_80_0_0_5ea0d0f264a95995656393acf2b3ab6a.jpg
МОСКВА, 5 мая — РИА Новости. Употребление комбинаций некоторых продуктов может привести к вздутию и несварению. Диетолог Елена Кален в своем Instagram назвала неудачные сочетания и раскрыла причины несовместимости отдельных продуктов.На сочетаемость продуктов влияют их количество, состав, время и ферменты, затрачиваемые организмом на переваривание.Диетолог рекомендует не использовать кислые фрукты, в частности, апельсины, ананасы, а также чернослив в маринаде мяса. Скорость их переваривания сильно разнится, использование таких ингредиентов может повлечь за собой тяжесть в животе, изжогу и несварение.С картофелем Кален не советует есть сыр и помидоры, так как эти овощи нивелируют полезные свойства друг друга, а их сочетание с сыром приводит к боли в суставах. В качестве альтернативы она предлагает употреблять мясо с овощами, которые не содержат крахмал, — морковь, брокколи, салат, спаржа.Врач отметила, что не следует есть бутерброды с сыром и колбасой, так как они перевариваются достаточно долго и перегружают пищеварительную систему. По схожей причине в список нежелательных продуктов попали пельмени, вареники и манты.Диетолог предупредила об опасности фруктов как заключительного приема пищи, поскольку они следуют за основными блюдами. На их усвоение организм тратит продолжительное время, в связи с чем может образовываться вздутие. По этой причине фрукты рекомендуется употреблять до застолья.Наконец, Кален подчеркнула вред от газированных и алкогольных напитков, которые наносят серьезный удар по печени и организму в целом из-за высокого содержания сахара и калорийности.
https://rsport.ria.ru/20210504/redis-1731106262.html
https://rsport.ria.ru/20210503/myaso-1730979663.html
https://rsport.ria.ru/20210430/zheludok-1730369225.html
Спорт РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2021
Спорт РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://rsport.ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
Спорт РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdn25.img.ria.ru/images/07e5/05/05/1731234303_110:0:1811:1276_1920x0_80_0_0_5f6eec4148f2d1677a5e6ef521215640.jpgСпорт РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Спорт РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
здоровье, питание, здоровый образ жизни (зож)
МОСКВА, 5 мая — РИА Новости. Употребление комбинаций некоторых продуктов может привести к вздутию и несварению. Диетолог Елена Кален в своем Instagram назвала неудачные сочетания и раскрыла причины несовместимости отдельных продуктов.На сочетаемость продуктов влияют их количество, состав, время и ферменты, затрачиваемые организмом на переваривание.
Диетолог рекомендует не использовать кислые фрукты, в частности, апельсины, ананасы, а также чернослив в маринаде мяса. Скорость их переваривания сильно разнится, использование таких ингредиентов может повлечь за собой тяжесть в животе, изжогу и несварение.
Диетолог рассказала, кому нельзя есть редисС картофелем Кален не советует есть сыр и помидоры, так как эти овощи нивелируют полезные свойства друг друга, а их сочетание с сыром приводит к боли в суставах. В качестве альтернативы она предлагает употреблять мясо с овощами, которые не содержат крахмал, — морковь, брокколи, салат, спаржа.
Врач отметила, что не следует есть бутерброды с сыром и колбасой, так как они перевариваются достаточно долго и перегружают пищеварительную систему. По схожей причине в список нежелательных продуктов попали пельмени, вареники и манты.
Диетолог рассказала, какими продуктами можно заменить мясоДиетолог предупредила об опасности фруктов как заключительного приема пищи, поскольку они следуют за основными блюдами. На их усвоение организм тратит продолжительное время, в связи с чем может образовываться вздутие. По этой причине фрукты рекомендуется употреблять до застолья.
Наконец, Кален подчеркнула вред от газированных и алкогольных напитков, которые наносят серьезный удар по печени и организму в целом из-за высокого содержания сахара и калорийности.
30 апреля, 04:30ЗОЖЭндокринолог рассказала, из-за чего на самом деле растягивается желудокАллерген f35 — картофель, IgE (ImmunoCAP)
Количественное определение в крови специфических антител, иммуноглобулинов класса E, появляющихся при наличии аллергической реакции к картофелю.
Синонимы русские
Специфический иммуноглобулин класса Е к картофелю.
Синонимы английские
Specific immunoglobulin E to the potato, Spec. IgE to the potato (serum).
Метод исследования
Иммунофлюоресценция на твердой фазе (ImmunoCAP).
Единицы измерения
кЕдА/л (килоединица аллергена на литр).
Какой биоматериал можно использовать для исследования?
Венозную или капиллярную кровь.
Как правильно подготовиться к исследованию?
- Не курить в течение 30 минут до исследования.
Общая информация об исследовании
Аллерген – это вещество, вызывающее аллергическую реакцию. Существует огромное количество веществ природного или искусственного происхождения, каждое из которых может стать аллергеном для человека.
Между собой аллергены различаются по растворимости, ферментативной и температурной стабильности. Пищевые аллергены бывают животного или растительного происхождения. Их особенностью является способность изменять аллергенные свойства в процессе кулинарной обработки. При этом одни пищевые продукты теряют аллергенность, а другие становятся опасными для человека. Иногда термическая обработка приводит к потере родной белковой структуры, но иммуноглобулины класса Е все еще могут реагировать с разрушенными пищевыми белками.
Основной участник аллергической реакции немедленного типа (1-го типа) – иммуноглобулин класса Е (IgE). Для каждого аллергена существует специфический иммуноглобулин Е. Целью данного теста является определение аллергической реакции к картофелю.
Состав картофеля достаточно разнообразен, в него входят крахмал, азотистые вещества, сахара, клетчатка, жиры, титруемые кислоты, пектиновые вещества, нуклеиновые кислоты, минералы, витамины С, В1, В2, В6, РР, К и каротиноиды.
По биологической ценности белки картофеля превосходят белки многих зерновых культур. В них содержатся все аминокислоты, встречающиеся в растениях, в том числе незаменимые: лизин, метионин, треонин, триптофан, валин, фенилаланин, лейцин, изолейцин. При этом в картофеле также есть много аллергенов, в том числе и принадлежащих к семейству профилинов. Главными аллергенами являются пататин и туберин. Кроме того, в составе присутствуют белки – ингибиторы цистеиновой и аспартатной протеаз и катепсин D, принадлежащие к семейству трипсина сои. Картофельная мука и крахмал обычно не содержат аллергены.
Не исключаются перекрестные реакции на пыльцу яблони, березы, ольхи, а также на вещества, входящие в состав овощей семейства пасленовых (баклажан, томаты, сладкий перец и др.).
Аллергия на картофель, как правило, проявляется нарушением функции пищеварительной системы (хейлит, гастрит, колит, гастроэнтерит, синдром раздраженной кишки), кожными проявлениями (атопический дерматит, крапивница, отек Квинке) и реже нарушениями функции дыхательной системы (аллергический ринит, бронхиальная астма).
Выполнение анализа безопасно для пациента по сравнению с кожными тестами (in vivo), так как исключает контакт с аллергеном. Кроме того, прием антигистаминных препаратов и возрастные особенности не влияют на качество и точность исследования.
Количественное определение специфических IgE-антител позволяет оценить взаимосвязь между уровнем антител и клиническими проявлениями аллергии. Низкие значения этого показателя указывают на низкую вероятность аллергического заболевания, в то время как высокий уровень имеет высокую корреляцию с клиническими проявлениями заболевания. При выявлении высоких уровней специфических IgE возможно предсказать развитие аллергии в будущем и более яркое проявление ее симптомов. Однако концентрация IgE в крови нестабильна. Она меняется с развитием заболевания, с количеством получаемой дозы аллергенов, а также на фоне лечения. Рекомендуется повторить исследование при изменении симптомов и при контроле проводимого лечения. О необходимости повторного исследования нужно консультироваться с лечащим врачом.
ImmunoCAP характеризуется высокой точностью и специфичностью: в малом количестве крови обнаруживаются даже очень низкие концентрации IgE-антител. Исследование является революционным и основано на иммунофлюоресцентном методе, что позволяет увеличить чувствительность в несколько раз по сравнению с другими анализами. Всемирная организация здравоохранения и Всемирная организация аллергологов признают диагностику с использованием ImmunoCAP как «золотой стандарт», так как она доказала свою точность и стабильность результатов в независимых исследованиях. В Российской Федерации до настоящего момента методика не получила широкого распространения, хотя во всем мире до 80 % анализов на специфические иммуноглобулины класса Е выполняется с помощью ImmunoCAP.
Таким образом, выявление специфических IgE с помощью данной методики выводит аллергодиагностику на качественно новый уровень.
Для чего используется исследование?
- Для диагностики аллергических заболеваний (пищевая аллергия, атопический дерматит, бронхиальная астма, аллергический ринит, респираторный аллергоз).
- Для оценки риска развития аллергических реакций на картофель.
Когда назначается исследование?
- При наличии следующих, указывающих на аллергический характер, симптомов после употребления в пищу картофеля и продуктов, в приготовлении которых он использовался: покраснение и зуд кожи, ангионевротический отек, риноконъюнктивит, отек гортани, кашель и бронхоспазм, тошнота, рвота, боль в области живота и диарея и др.
- При непрерывно рецидивирующем заболевании без периодов ремиссии.
- При невозможности отмены препаратов для лечения основного заболевания, так как они могут повлиять на результаты кожного тестирования.
- Детям – если их родители страдают аллергическими заболеваниями.
- Детям в возрасте до 5 лет (при невозможности выполнения кожных тестов).
- При поражении кожных покровов, в острый период заболевания.
- При поливалентном характере сенсибилизации, когда нет возможности провести тестирование in vivo с предполагаемыми аллергенами.
- При ложноположительном или ложноотрицательном результате кожного тестирования.
Что означают результаты?
Референсные значения
|
Значение показателя, кЕдА/л |
Класс |
Уровень аллергенспецифических антител IgE |
|
0 – 0,35 |
0 |
Отсутствует |
|
0,351 – 0,69 |
1 |
Низкий |
|
0,70 – 3,49 |
2 |
Средний |
|
3,5 – 17,49 |
3 |
Высокий |
|
17,5 – 49,99 |
4 |
Очень высокий |
|
50,0 – 100,0 |
5 |
Насыщенно высокий |
|
Больше 100,0 |
6 |
Крайне высокий |
Причины повышения уровня специфических IgE:
- наличие аллергических реакций к картофелю;
- атопический дерматит, пищевая аллергия, обусловленные сенсибилизацией к аллергенам картофеля.
Причины снижения уровня специфических IgE
При повторном исследовании (в динамике) уровень специфических IgE может снижаться по следующим причинам:
- ограничение или исключение контакта с аллергеном;
- проведение медикаментозного лечения.
Перекрестные реакции
- Перекрестные реакции – это аллергические реакции на одинаковые по структуре компоненты, представленные в составе различных аллергенов. Во избежание необоснованного выполнения ненужных для пациента тестов и для индивидуального выбора возможных перекрестно реагирующих аллергенов необходима консультация врача-аллерголога.
Также рекомендуется
Кто назначает исследование?
Аллерголог, педиатр, пульмонолог, гастроэнтеролог, терапевт, врач общей практики, ревматолог.
Литература
- Reese G., Lehrer S.B. Food Allergenes. In: Food hypersensitivity and adverse reactions. – New York: Marcel Dekker Inc., 1999. – P. 69-97.
- Burney PGJ et al. The distribution of total and specific serum IgE in the European Community Respiratory Health Survey. J Allergy Clin Immunol 1997;99:314-22.
- Bergmann RL et al. Atopic diseases in infancy. The German multicenter atopy study (MAS-90). Pediatr Allergy Immunol 1994;5 (Suppl 1):19-25.
- Schäfer T et al. Association between severity of atopic eczema and degree of sensitization to aeroallergens in school children. J Allergy Clin Immunol 1999;104:1280-84.15.
- Dolen W.K., Allergy Review Series X: Progress in diagnosis of allergy in vitro. IgE antibody in the serum – detection and diagnostic significance. Allergy 2003; 58: 717-723.
- Sampson H.A., Ho D.G., Relationship between food-specific IgE concentrations and the risk of positive food challenges in children and adolescents. J Allergy Clin Immunol, 1997; 100: 444-51.
- Yunginger JW, Ahlstedt S, Eggleston PA, Homburger HA, Nelson HS, Ownby DR, Platts-Mills TAE, Sampson HA, Sicherer SH, Weinstein AM, Williams PB, Wood RA, Zeiger RS., Quantitative IgE antibody assays in allergic diseases. J Allergy Clin Immunol 2000; 105:1077-84.
Картофельный крахмал — польза и вред — «Гиорд»
Каждый из нас хоть раз в жизни сталкивался или слышал о таком продукте, как картофельный крахмал — белый иногда желтоватый порошок, не имеющий ни вкуса, ни запаха. Сам по себе крахмал относится к сложным углеводам и входит в состав фруктов, овощей, круп, фасоли, орехов. Этот продукт легко усваивается организмом и не вызывает побочных эффектов и аллергических реакций.
Если в картофельный крахмал добавить жидкость, то получится клейкое вещество. Именно поэтому он очень популярен при приготовлении разных кулинарных блюд. Одним из свойств картофельного крахмала, при соединении с водой, является увеличение его в объёме. Если добавить картофельный крахмал в подливку или соус, он будет способствовать загустению конечного продукта. Недостатком является то что такой соус или подливка в случае длительного хранения расслаиваются и становятся мутными, следовательно, употребить такой продукт нужно как можно быстрее. Также картофельный крахмал используют в выпечки, его добавляют к муке. Благодаря этому улучшается структура выпечки, готовый продукт становится рассыпчатым.
В 100 г продукта содержится приблизительно 313 ккал.
Картофельный крахмал — это весьма полезный и питательный для здоровья продукт. Регулярное употребление крахмала в пищу, ведет к стабилизации уровня холестерина в крови. Что значительно снижает возможность возникновения такого заболевания, как атеросклероза. Калий, который содержится в картофеле, а следовательно, и в крахмале, улучшает деятельность сердечной мышцы и функцию почек, способствует выведению лишней жидкости из организма.
Также картофельный крахмал получил широкое распространение в народной медицине, в которой он используется как заживляющее средство при язвах и ранах. Он имеет обволакивающие и противовоспалительное свойство. Проведённые научные исследования, доказали, что крахмал способствует развитию процесса синтеза рибофлавина, что нормализует обмен веществ организма.
Сложные углеводы бывают двух типов: натуральные, получаемые из пищевых продуктов, и синтетические. Синтетические углеводы ещё носят название рафинированные и производятся промышленным путём. Рафинированные углеводы — это мука высокого качества, крахмал, или продукты, которые приготовлены из этих ингредиентов. Искусственный крахмал относится к вредным веществам, потому что значительно повышает содержание инсулина в крови. В свою очередь, повышенный инсулин способствует развитию тяжёлых болезней и гормональному сбою.
Постоянное использование в пищу продуктов, содержащих картофельный крахмал, вредит человеческому здоровью. Казалось бы, такой безвредный продукт, как булочки, пирожные и тортики способствуют возникновению злокачественных раковых опухолей.
Какие формы углеводов организм не может переваривать? | Здоровое питание
Автор: Ян Анниган Обновлено 9 декабря 2018 г.
Пищевые углеводы состоят из сахаров, крахмалов и клетчатки. Некоторые из них вы можете переварить, и они обеспечивают энергию, необходимую для питания ваших мышц и поддержки активности ваших клеток. Углеводы, которые вы не можете переваривать, не являются топливом, но все же приносят пользу вашему здоровью. Они также могут вызывать неприятные желудочно-кишечные симптомы, в зависимости от вашего общего рациона и вашей конкретной физиологии.
Волокно
Пищевые волокна получают из продуктов растительного происхождения, которые содержат целлюлозу, гемицеллюлозу, лигнин, пектин, слизь и камедь. В вашей пищеварительной системе не хватает ферментов для расщепления этих пищевых компонентов, и они проходят через кишечник, не перевариваясь.
Нерастворимая клетчатка, содержащаяся в цельнозерновых, бобовых и овощах, увеличивает объем ваших отходов и ускоряет их прохождение через кишечник, что может уменьшить запоры и развитие геморроя.
Растворимая клетчатка, напротив, замедляет движение пищи через кишечник и помогает регулировать скорость всасывания питательных веществ, таких как холестерин и глюкоза, в кровоток. Таким образом, он может помочь контролировать диабет и предотвратить сердечные заболевания. Овсянка, фрукты, овощи и бобы вносят в ваш рацион растворимую клетчатку.
Устойчивый крахмал
Большая часть съедаемого вами крахмала усваивается, но особый тип, известный как резистентный крахмал, — нет. Некоторые незрелые фрукты, такие как манго и бананы, а также определенные крахмалистые продукты, которые были приготовлены и оставлены для охлаждения — например, макаронные изделия и картофель в холодных салатах — содержат крахмал, который не реагирует на ваши пищеварительные ферменты.Вместо этого эти крахмалы реагируют так же, как клетчатка в пищеварительном тракте. Они могут помочь вам почувствовать сытость и замедлить усвоение питательных веществ. Кроме того, поскольку продукты, содержащие резистентный крахмал, предлагают меньше энергии, чем полностью нестойкие крахмалистые продукты, вы потребляете меньше калорий с этим типом углеводов, что потенциально полезно, если вы пытаетесь похудеть.
Определенные сахара
Некоторые сахара в вашем рационе невозможно переваривать, если вы не производите достаточное количество пищеварительных ферментов, необходимых для их расщепления, или не производите их вообще.Например, рафиноза — это сахар, содержащийся в бобах, брюссельской капусте, брокколи и капусте, и, если вы не можете переварить часть или весь этот сахар, он попадает в толстую кишку. Здесь естественная популяция бактерий переваривает его и при этом выделяет газ в качестве побочного продукта. Точно так же лактоза из молочного сахара не переваривается, если вам не хватает необходимых пищеварительных ферментов. Он также попадает в толстую кишку, где бактерии питаются им, выделяют газы и вызывают метеоризм или дискомфорт в животе.
Рекомендации
Потребляемая клетчатка также может вызывать неприятные симптомы со стороны пищеварительного тракта. Хотя нерастворимая клетчатка помогает регулировать работу кишечника, если вы едите слишком много или слишком быстро увеличиваете потребление, это может вызвать боль в животе, запор или непроходимость кишечника, особенно если вы не увеличиваете потребление воды. Резистентный крахмал не вызывает этой проблемы с желудочно-кишечным трактом.
5.1: Крахмал и целлюлоза — Химия LibreTexts
Полисахариды — это самые распространенные в природе углеводы, которые выполняют множество функций, таких как хранение энергии или как компоненты стенок растительных клеток.Полисахариды — это очень большие полимеры, состоящие из десятков и тысяч моносахаридов, соединенных гликозидными связями. Три наиболее распространенных полисахарида — это крахмал, гликоген и целлюлоза. Эти три упоминаются как гомополимеры , потому что каждый дает только один тип моносахарида (глюкозу) после полного гидролиза. Гетерополимеры могут содержать сахарные кислоты, аминосахары или неуглеводные вещества в дополнение к моносахаридам. Гетерополимеры широко распространены в природе (камеди, пектины и другие вещества), но не будут обсуждаться далее в этом учебнике.Полисахариды являются невосстанавливающими углеводами, не имеют сладкого вкуса и не подвергаются мутаротации.
Крахмал
Крахмал является наиболее важным источником углеводов в рационе человека и составляет более 50% потребляемых нами углеводов. Он встречается в растениях в виде гранул, и их особенно много в семенах (особенно в зернах злаков) и клубнях, где они служат формой хранения углеводов. Распад крахмала до глюкозы питает растение в периоды пониженной фотосинтетической активности.Мы часто думаем о картофеле как о «крахмалистой» пище, однако другие растения содержат гораздо больший процент крахмала (картофель 15%, пшеница 55%, кукуруза 65% и рис 75%). Товарный крахмал представляет собой белый порошок.
Крахмал представляет собой смесь двух полимеров: амилозы и амилопектина. Натуральные крахмалы состоят примерно на 10–30% из амилазы и на 70–90% из амилопектина. Амилоза представляет собой линейный полисахарид, полностью состоящий из единиц D-глюкозы, соединенных α-1,4-гликозидными связями, которые мы видели в мальтозе (часть (а) на рисунке 5.1.1). Экспериментальные данные показывают, что амилоза не является прямой цепочкой глюкозных единиц, а вместо этого свернута, как пружина, с шестью мономерами глюкозы на виток (часть (b) рисунка 5.1.1). При таком свертывании амилоза в ядре имеет достаточно места для размещения молекулы йода. Характерный сине-фиолетовый цвет, который появляется при обработке крахмала йодом, обусловлен образованием комплекса амилоза-йод. Этот цветовой тест достаточно чувствителен, чтобы обнаруживать даже незначительные количества крахмала в растворе.
Рисунок 5.1.1: Амилоза. (а) Амилоза представляет собой линейную цепь из единиц α-D-глюкозы, соединенных вместе α-1,4-гликозидными связями. (б) Из-за водородных связей амилоза приобретает спиралевидную структуру, содержащую шесть единиц глюкозы на оборот.
Амилопектин представляет собой полисахарид с разветвленной цепью, состоящий из единиц глюкозы, связанных главным образом α-1,4-гликозидными связями, но иногда с α-1,6-гликозидными связями, которые ответственны за разветвление.Молекула амилопектина может содержать многие тысячи единиц глюкозы с точками ветвления, встречающимися примерно через каждые 25–30 единиц (рис. 5.1.2). Спиральная структура амилопектина нарушается из-за разветвления цепи, поэтому вместо темно-сине-фиолетовой окраски амилозы с йодом амилопектин дает менее интенсивный красновато-коричневый цвет.
Рисунок 5.1.2 : Представление ветвления амилопектина и гликогена. И амилопектин, и гликоген содержат точки ветвления, которые связаны через α-1,6-связи.Эти точки ветвления чаще встречаются в гликогене.
Декстрины представляют собой полисахариды глюкозы промежуточного размера. Блеск и жесткость, придаемые одежде крахмалом, обусловлены присутствием декстринов, образующихся при глажке одежды. Из-за их характерной липкости при намокании декстрины используются в качестве клея на марках, конвертах и этикетках; как связующие вещества для удерживания пилюль и таблеток вместе; и как пасты. Декстрины перевариваются легче, чем крахмал, и поэтому широко используются при коммерческом приготовлении детского питания.
Полный гидролиз крахмала дает последовательные стадии глюкозы:
крахмал → декстрины → мальтоза → глюкоза
В организме человека несколько ферментов, известных под общим названием амилазы, последовательно расщепляют крахмал до пригодных для использования единиц глюкозы.
Гликоген
Гликоген — это углевод, являющийся энергетическим резервом животных. Практически все клетки млекопитающих содержат некоторое количество хранимых углеводов в форме гликогена, но особенно много его в печени (4-8% от веса ткани) и в клетках скелетных мышц (0.5% –1,0%). Как и крахмал в растениях, гликоген находится в виде гранул в клетках печени и мышц. При голодании животные потребляют эти запасы гликогена в течение первого дня без еды для получения глюкозы, необходимой для поддержания метаболического баланса.
Примечание
Около 70% общего гликогена в организме хранится в мышечных клетках. Хотя процентное содержание гликогена (по весу) выше в печени, гораздо большая масса скелетных мышц хранит большее общее количество гликогена.
Гликоген структурно очень похож на амилопектин, хотя гликоген более разветвлен (8–12 единиц глюкозы между ветвями), а ветви короче.При обработке йодом гликоген дает красновато-коричневый цвет. Гликоген может быть расщеплен на его субъединицы D-глюкозы путем кислотного гидролиза или с помощью тех же ферментов, которые катализируют расщепление крахмала. У животных фермент фосфорилаза катализирует распад гликогена до фосфатных эфиров глюкозы.
Целлюлоза
Целлюлоза, волокнистый углевод, содержащийся во всех растениях, является структурным компонентом стенок растительных клеток. Поскольку земля покрыта растительностью, целлюлоза является самым распространенным из всех углеводов, на ее долю приходится более 50% всего углерода, содержащегося в царстве растений.Волокна хлопка и фильтровальная бумага почти полностью состоят из целлюлозы (около 95%), древесина составляет около 50% целлюлозы, а сухой вес листьев составляет около 10–20% целлюлозы. Наибольшее распространение целлюлоза используется в производстве бумаги и бумажных изделий. Хотя использование нецеллюлозных синтетических волокон растет, вискоза (из целлюлозы) и хлопок по-прежнему составляют более 70% текстильного производства.
Подобно амилозе, целлюлоза представляет собой линейный полимер глюкозы. Однако он отличается тем, что единицы глюкозы соединены β-1,4-гликозидными связями, образуя более протяженную структуру, чем амилоза (часть (а) на рисунке 5.1.3). Эта крайняя линейность позволяет образовывать много водородных связей между группами ОН в соседних цепях, заставляя их плотно упаковываться в волокна (часть (b) на рисунке 5.1.3). В результате целлюлоза слабо взаимодействует с водой или любым другим растворителем. Например, хлопок и дерево полностью нерастворимы в воде и обладают значительной механической прочностью. Поскольку целлюлоза не имеет спиральной структуры, она не связывается с йодом с образованием окрашенного продукта.
Рисунок 5.1,3 : Целлюлоза. (а) В структуре целлюлозы имеется обширная водородная связь. (b) На этой электронной микрофотографии клеточной стенки водоросли стенка состоит из последовательных слоев целлюлозных волокон, расположенных параллельно.
Целлюлоза дает D-глюкозу после полного кислотного гидролиза, но люди не могут метаболизировать целлюлозу как источник глюкозы. В наших пищеварительных соках отсутствуют ферменты, которые могут гидролизовать β-гликозидные связи, содержащиеся в целлюлозе, поэтому, хотя мы можем есть картофель, мы не можем есть траву.Однако некоторые микроорганизмы могут переваривать целлюлозу, потому что они производят фермент целлюлазу, который катализирует гидролиз целлюлозы. Присутствие этих микроорганизмов в пищеварительном тракте травоядных животных (таких как коровы, лошади и овцы) позволяет этим животным разлагать целлюлозу из растительного материала до глюкозы для получения энергии. Термиты также содержат микроорганизмы, выделяющие целлюлазу, и поэтому могут питаться древесной пищей. Этот пример еще раз демонстрирует крайнюю стереоспецифичность биохимических процессов.
Определение и примеры крахмала — Биологический онлайн-словарь
Рецензент: Тодд Смит, доктор философии
Определение крахмала
существительное
множественное число: крахмал, крахмал
крахмал, stɑɹtʃ
Полисахаридный углевод H 10 O 5 ) n, состоящий из большого количества молекул глюкозы, соединенных вместе гликозидными связями, и обнаруживается, в частности, в семенах, луковицах и клубнях
Обзор
Крахмал принадлежит к группе полисахаридов углеводов .Углеводы — это органические соединения, состоящие из углерода, водорода и кислорода, обычно в соотношении 1: 2: 1. Это один из основных классов биомолекул. Как питательные вещества их можно разделить на две основные группы: простых углеводов и сложных углеводов . Простые углеводы, иногда называемые просто , сахар , состоят из одного или двух сахаридных остатков. Они легко перевариваются и служат быстрым источником энергии. Сложные углеводы (такие как целлюлоза , крахмал , хитин и гликоген ) — это те углеводы, которым требуется больше времени для переваривания и метаболизма.Они часто богаты клетчаткой и, в отличие от простых углеводов, с меньшей вероятностью вызывают скачки уровня сахара в крови. В частности, гликоген накапливается в печени для быстрого доступа к энергии, поскольку он сжигается раньше жира.
История и терминология
Крахмал был известен и использовался уже 100 000 лет назад. Считается, что его используют в приготовлении пищи, например, при приготовлении хлеба и каш. Эта гипотеза основана на каменных орудиях, обнаруженных в старых пещерах.Инструменты, вероятно, использовались для очистки и измельчения зерен крахмала дикого сорго. Это наблюдение заставило ученых предположить, что включение крахмала в доисторический рацион первых людей африканских саванн и лесов улучшило качество рациона. Переработка зерна в основной продукт ознаменовала собой сдвиг в доисторическом рационе и считается решающим шагом в эволюции человека. (Ссылка 1) Слово крахмал может происходить от древнеанглийского stearc («резкий, крепкий, грубый»), которое, в свою очередь, могло иметь германское происхождение, т.е.е. starchī , что означает «сильный».
Характеристики
Крахмал — это сложный полисахарид, состоящий из большого количества единиц глюкозы, соединенных гликозидными связями. Это белый порошок без вкуса и запаха. Имеет переменную молярную массу. Не растворяется в спирте и холодной воде. Его химическая формула: (C 6 H 10 O 5 ) n . Два типа молекул содержат чистый крахмал: амилоза и амилопектин .И амилоза, и амилопектин представляют собой полисахариды, состоящие из остатков глюкозы. Они различаются по структуре: амилоза — это линейная цепь молекул глюкозы , соединенная α- (1,4) гликозидными связями, тогда как амилопектин — это разветвленная цепь молекул глюкозы , линейно связанная с α- (1,4) гликозидными связями. и α- (1,6) связи с интервалом от 24 до 30 субъединиц глюкозы. Поскольку крахмал представляет собой полисахарид, состоящий в основном из D-глюкозы, он принадлежит к группе α-глюканов .
Амилопектин более растворим в воде и легче переваривается, чем амилоза. Его растворимость обусловлена множеством конечных точек, которые могут образовывать водородные связи с водой. Обычно крахмал содержит 75-80% амилопектина и 20-25% амилозы по весу.
Синтез обезвоживания
Химический процесс соединения моносахаридных единиц называется синтез обезвоживания , поскольку он приводит к выделению воды в качестве побочного продукта. Крахмал производится путем дегидратационного синтеза.Растения хранят глюкозу, которая не используется в качестве крахмала. Сначала глюкоза фосфорилируется до глюкозо-1-фосфат . Гранулы крахмала хранятся внутри амилопластов , расположенных внутри клеток различных органов растений. Гранулы крахмала можно найти в плодах , семенах , клубнях и корневищах . Маргаритки, подсолнухи и топинамбур являются примерами растений, которые хранят инулин (фруктан) вместо крахмала.
Разложение
В растениях разложение крахмала происходит естественным образом в ночное время.Фермент глюкановодикиназа фосфорилирует крахмал, особенно по C-6 одного из остатков глюкозы. Затем другой фермент (фосфоглюкан-водная дикиназа ) фосфорилирует остаток глюкозы по C-3. После фосфорилирования разлагающие ферменты теперь могут воздействовать на крахмал, высвобождая простые сахара. Например, бета-амилаза высвобождает два остатка глюкозы в виде мальтозы . Другой разрушающий фермент — это диспропорционирующий фермент-1 , который в конце процесса деградации высвобождает молекулу глюкозы.Разложение крахмала приводит в основном к мальтозе и меньшему количеству глюкозы. Эти простые сахара затем будут перемещаться из пластиды в цитозоль с помощью транспортеров: транспортер мальтозы для мальтозы и пластидный транслокатор глюкозы для глюкозы. Позже они могут быть использованы в качестве субстрата для биосинтеза сахарозы, которая важна в митохондриальном пути окислительной пентозы , который генерирует АТФ в ночное время. (Ссылка 2)
Гидролиз
Гидролиз — это процесс превращения полисахарида, такого как крахмал, в простые сахарные компоненты.Процесс превращения полисахаридов в моносахариды, в частности, называется осахариванием . У человека сложные углеводы, такие как крахмал, перевариваются посредством ряда ферментативных реакций. Эти ферменты представляют собой амилазы слюны , амилазы поджелудочной железы и мальтазы . Амилаза слюны воздействует на крахмал и расщепляет его до мальтозы . Когда частично переваренные углеводы достигают тонкой кишки, поджелудочная железа выделяет сок поджелудочной железы, который включает амилазу поджелудочной железы .Этот фермент действует на частично переваренные углеводы, расщепляя их на простые сахара. Кайма тонкой кишки высвобождает пищеварительные ферменты, такие как изомальтаза , мальтаза , сахараза и лактаза . Изомальтаза переваривает полисахариды по альфа-1-6 связям и превращает альфа-предельный декстрин в мальтозу. Мальтаза расщепляет мальтозу (дисахарид) на две единицы глюкозы. Сахараза и лактаза расщепляют сахарозу и лактозу на моносахаридные составляющие соответственно.Эпителиальные клетки ( энтероцитов, ) щеточной каймы тонкой кишки поглощают моносахариды, а затем высвобождают их в капилляры. Затем простые сахара переносятся из кровотока в клетки других тканей, особенно в печень. Глюкоза в крови может использоваться организмом для производства АТФ. В противном случае он транспортируется в печень вместе с галактозой и фруктозой (которые в значительной степени превращаются в глюкозу) для хранения в виде гликогена.
Резистентный крахмал
Резистентный крахмал — это форма крахмала, которая противостоит перевариванию в тонком кишечнике человека.Это также пищевых волокон . Вместо этого он метаболизируется в толстом кишечнике микробиотой толстой кишки. Микробы в толстой кишке ферментируют его и производят побочные продукты метаболизма, такие как газы и короткоцепочечные жирные кислоты. В частности, короткоцепочечные жирные кислоты абсорбируются и приносят пользу человеческому организму. Ферментация устойчивого крахмала также способствует росту полезных бактерий.
Растительный крахмал по сравнению с животным крахмалом
Животный крахмал не является крахмалом как таковой .Он относится к компоненту гликогена животного из-за сходства в структуре и составе амилопектина. В то время как растения хранят избыток глюкозы в форме крахмала, животные также хранят это в форме гликогена . Гликоген — это разветвленный полимер глюкозы, который в основном вырабатывается в клетках печени и мышц и функционирует как вторичное долгосрочное хранилище энергии в клетках животных. Как и крахмал, гликоген — это сложный углевод, который в первую очередь служит запасным углеводом.Разница между амилопектином в растениях и амилопектином у животных заключается в том, что последний имеет более обширное разветвление на каждые 8–12 единиц глюкозы.
Биологическое значение
Все семена и клубни растений содержат крахмал, который преимущественно присутствует в виде амилозы и амилопектина. Растения используют крахмал как способ хранения избыточной глюкозы и, таким образом, также используют крахмал в качестве пищи за счет окислительного фосфорилирования митохондрий ночью или когда фотосинтез маловероятен. Растения накапливают избыток крахмала в амилопластах, которые представляют собой лейкопласты, которые функционируют в основном для хранения гранул крахмала за счет полимеризации глюкозы и преобразования этих запасов обратно в более простые сахара (например,г. мальтоза и глюкоза), особенно при отсутствии света. Хлоропласты, пигментированные органеллы, участвующие в основном в фотосинтезе, также способны накапливать крахмал.
Животные не хранят избыток глюкозы в виде крахмала; они хранят их в виде гликогена. Однако некоторые животные питаются крахмалосодержащей пищей.
Диетический крахмал присутствует во многих основных продуктах питания, таких как кукуруза , рис , пшеница , картофель , маниока , ячмень , рожь , таро , ямс , и т. Д. .Он также присутствует в различных пищевых продуктах, таких как злаков , лапша , блины , хлеб , макаронные изделия , и т. Д. . Крахмал обеспечивает около 4,2 килокалорий на грамм. У человека крахмал может служить основным источником глюкозы. Глюкоза важна, так как она участвует в общем метаболизме, например гликолиз (для синтеза энергии), гликогенез (для синтеза гликогена), пентозофосфатный путь (для пентоз и синтеза НАДФН для использования в синтезе нуклеиновых кислот и синтезе липидов, соответственно).
Крахмал имеет множество коммерческих применений, например, в бумажном производстве, в пищевых продуктах, при производстве коммерческого виноградного сахара, для придания жесткости белью в прачечных, для изготовления пасты, в полиграфической промышленности, производстве водорода и т. Д.
Риск для здоровья
Слишком много крахмала в рационе связано с кариесом зубов , ожирением и сахарным диабетом . Крахмал (особенно приготовленный и содержащийся в обработанных пищевых продуктах) может вызывать скачки уровня глюкозы в крови после еды.Таким образом, рекомендуется умеренное потребление крахмала. Людям с глютеновой болезнью и врожденным дефицитом сахаразы-изомальтазы может потребоваться избегать крахмалистых продуктов. (Ссылка 3)
Этимология
- Древнеанглийский stearc («резкий, сильный, грубый»)
Химическая формула
Связанные термины
Сравнить
См. Также
Ссылки:
- Каша была съели 100000 лет назад.(2009, 18 декабря). Получено с telegraph.co.uk/news/uknews/6834609/Porridge-was-eaten-100000-years-ago.html Ссылка
- Соавторы Википедии. (2019, 25 февраля). Крахмал. Получено с en.wikipedia.org/wiki/Starch#Energy-store-of-plants Ссылка
- Крахмал: продукты питания, пищеварение, гликемический индекс. (2016, 4 июня). Получено с сайта nutritiousreview.com/carbs/polysaccharides-starch.html Ссылка
© Biology Online. Контент предоставлен и модерируется онлайн-редакторами биологии
Спросите эксперта: бобовые и устойчивый крахмал | Источник питания
Эксперт: Dr.Гай Кросби
Бобовые, в состав которых входят бобы, горох и чечевица, являются недорогим и здоровым источником белка, калия и сложных углеводов, включая пищевые волокна.
- В среднем бобовые содержат около 20-25% белка по весу в пересчете на сухое вещество, что в 2-3 раза больше белка, чем в пшенице и рисе. Однако они, как правило, содержат мало незаменимой аминокислоты метионина, а иногда и триптофана.
- Бобовые также являются очень хорошим источником пищевых волокон, которые важны для поддержания здоровой функции кишечника.
- Содержание общих углеводов, включая сложные углеводы, колеблется от 65 до 72% по весу в пересчете на сухое вещество, из которых 85% состоит из крахмала, а пищевые волокна составляют от 10 до 20% от веса сушеных бобовых ( 1).
Высокий уровень как крахмала, так и пищевых волокон поднимает очень интересный вопрос. Если большая часть углеводов состоит из крахмала, как бобовые могут быть такими хорошими источниками пищевых волокон? Разве не весь крахмал быстро переваривается до глюкозы? Это было преобладающим представлением о крахмале до 1980-х годов, когда два английских исследователя, Ганс Энглист и Джон Каммингс, обнаружили, что не весь крахмал быстро переваривается в глюкозу в тонком кишечнике (2).
- Они обнаружили, что некоторое количество крахмала устойчиво к перевариванию и попадает в толстую кишку, где большая часть его используется в качестве источника пищи здоровыми бактериями, живущими в нашей толстой кишке. Энглист и Каммингс назвали эту ранее неизвестную форму крахмала «резистентным крахмалом» и пришли к выводу, что он действует аналогично пищевому волокну.
- Существенные исследования с момента их открытия подтвердили, что резистентный крахмал (RS) действует так же, как пищевые волокна в пище (3).
Пищевые волокна содержат две основных формы:
- Одна из них — нерастворимая клетчатка, состоящая из сложных углеводов, таких как целлюлоза, которые нерастворимы в желудочно-кишечной жидкости.Эта форма клетчатки не расщепляется кишечными бактериями, действует как наполнитель и эффективно уменьшает запоры.
- Другая форма клетчатки растворима в желудочно-кишечной жидкости, образуя густую вязкую жидкость, похожую на мед. Растворимая клетчатка легко метаболизируется кишечными бактериями, которые превращают большую ее часть в небольшие молекулы, называемые короткоцепочечными жирными кислотами (SCFA). Клетки, выстилающие толстую кишку, получают около 60-70% своей энергии из SCFA (4).
Хотя резистентный крахмал не увеличивает вязкость желудочно-кишечной жидкости, он является одним из лучших источников SCFA, помогающих поддерживать здоровье клеток толстой кишки.
- Поскольку резистентный крахмал не метаболизируется в тонком кишечнике, он снижает количество глюкозы, попадающей в кровь, тем самым снижая потребность в инсулине, а также снижает калорийность пищи (5).
- Продукты, содержащие значительный уровень резистентного крахмала, повышают чувство насыщения и имеют более низкий гликемический индекс, вызывая меньшее повышение уровня глюкозы в крови, чем продукты с высоким содержанием крахмала, содержащие очень мало резистентного крахмала, такие как печеный картофель, рис и белый хлеб (6).Ясно, что не все углеводы одинаковы.
Бобовые — один из лучших источников устойчивого крахмала. Сырые сушеные бобовые содержат около 20-30% устойчивого крахмала по весу (7). Это означает, что почти половина крахмала в сырых бобовых не переваривается.
Почему крахмал устойчив к перевариванию? Небольшая его часть физически недоступна для пищеварительных ферментов. Но большинство из них устойчиво из-за химической структуры крахмала. Крахмал состоит из двух молекул, называемых амилозой и амилопектином.
- Амилоза представляет собой линейную цепь молекул глюкозы, соединенных встык.
- Амилопектин — гораздо более крупная молекула с многочисленными ответвлениями коротких цепочек молекул глюкозы, связанных с основной цепью, как ветви дерева, растущие из ствола.
Молекулы крахмала, особенно амилоза, образуют кристаллические области, устойчивые к перевариванию ферментами, переваривающими крахмал, в нашем организме (3). По сравнению с другими продуктами с высоким содержанием крахмала, такими как кукуруза, пшеница и рис, крахмал в бобовых очень богат амилозой, составляя до 40% крахмала, что делает их более устойчивыми к перевариванию.
- Важно понимать, что резистентный крахмал — это не отдельная молекулярная структура, такая как глюкоза или холестерин, а концепция, разработанная для объяснения того, почему некоторые крахмалы не перевариваются (8).
- Количество резистентного крахмала, указанное в пищевых продуктах, поэтому сильно зависит от метода, используемого для анализа резистентного крахмала. В 2000 г. стандартизированный тест для определения содержания RS в продуктах питания был одобрен AACC International (9). Цифры, приведенные в этой статье, были определены этим методом (10).
Если бы приготовленные бобовые содержали столько стойкого крахмала, сколько сырые бобовые, нам было бы очень трудно их переваривать. Как и все продукты с высоким содержанием крахмала, при приготовлении бобовых в кипящей воде большие части кристаллических областей разрушаются, уменьшая количество устойчивого крахмала. Но поскольку в бобовых так много амилозы, при варке разрушается меньшее количество резистентного крахмала, чем в других пищевых продуктах, потому что некоторые формы кристаллической амилозы стабильны даже в кипящей воде (8).
- Полностью приготовленные бобовые содержат только около 4-5% от их общего веса (в пересчете на сухое вещество) в виде устойчивого крахмала, независимо от того, как долго они варятся.
- Охлаждение приготовленных бобовых в течение 24 часов в холодильнике увеличивает уровень устойчивого крахмала до 5-6% от общего веса (в пересчете на сухой вес), позволяя некоторым молекулам крахмала перекристаллизоваться. Целые консервированные бобы содержат примерно такое же количество, как и консервированные жареные бобы (10).
Может показаться, что это не очень стойкий крахмал, но он все равно в 4-5 раз выше, чем в других крахмалистых продуктах, таких как белый хлеб и картофель (см. Таблицу ниже).Этого, безусловно, достаточно, чтобы оказать значительное влияние на образование SCFA, гликемический индекс, снижение инсулинового ответа, насыщения и калорийности (11). В общем, бобовые — это здоровый выбор не только из-за высокого содержания белка и других питательных веществ, но и потому, что они содержат один из самых высоких уровней резистентного крахмала среди всех продуктов питания.
| Продукты питания | Крахмал * | RS * | Гликемический индекс ** (12) |
| Картофель запеченный | 94% | <1% | 93 |
| Белый хлеб | 88% | 1% | 70 |
| Фасоль черная, вареная | 60% | 5% | 30 |
| Фасоль пинто, вареная | 61% | 5% | 39 |
- Массовый процент в пересчете на сухое вещество
** Гликемический индекс ниже 55 считается низким показателем G.I. еда (12).
1. Национальная база данных по питательным веществам Министерства сельского хозяйства США для справочной информации (2011 г.).
2. Энглист, Х. Н., Кингман, С. М., и Каммингс, Дж. Х., Классификация и измерение важных для питания фракций крахмала. Евро. J. Clin. Nutr. 1992; 46: 533-550.
3. Кросби Г. А. Устойчивый крахмал делает углеводы лучше. Функциональные продукты питания и нутрицевтики, 2003; 6: 34-36.
4. Топпинг Д. Л. и Клифтон П. М. Короткоцепочечные жирные кислоты и функция толстой кишки человека: роль резистентных крахмальных и некрахмальных полисахаридов. Physiol. Rev. 2001; 81 (3): 1031-1064.
5. Бехолл К. М. и Хоу Дж. К. Вклад клетчатки и устойчивого крахмала в метаболизируемую энергию. Являюсь. J. Clin. Nutr. 1995; 62: 1158S-1160S.
6. Raban, A., et al. Резистентный крахмал: влияние на постпрандиальную гликемию, гормональный ответ и чувство насыщения.Являюсь. J. Clin. Nutr. 1994; 60: 544-551.
7. Bednar, G.E., et al. Фракции крахмала и клетчатки в выбранных пищевых и кормовых ингредиентах влияют на их переваримость и ферментируемость в тонком кишечнике, а также на ферментируемость толстого кишечника in vitro на модели собак. J. Nutr. 2001; 131: 276-286.
8. Томпсон Д. Б. Стратегии производства устойчивого крахмала. Trends Food Sci. Tech. 2000; 11 (7): 245-253.
9. Переа, А., Меда, В., и Тайлер, Р. Т. Устойчивый крахмал: обзор аналитических протоколов для определения содержания устойчивого крахмала в пищевых продуктах. Food Res. Междунар. 2010; 43: 1959–1974.
10. Фаббри А. Д. Т., Шахт Р. В. и Кросби Г. А. Оценка содержания устойчивого крахмала в вареной черной фасоли, фасоли пинто и нуте. NFS Journal 3. 2016; 8-12.
11. Noah, L. et al. Переваривание углеводов из белой фасоли (Phaseolus vulgaris L.) у здоровых людей. J. Nutr.1998; 128: 977-985.
12. Бранд-Миллер, Дж., Волевер, Т. М. С., Колагиури, С., и Фостер-Пауэлл, К. Глюкозная революция: авторитетное руководство по гликемическому индексу. Marlowe & Co., NY, 1999.
Амилопектин — обзор | ScienceDirect Topics
Крахмал, природный субстрат для амилаз
Крахмал обычно состоит из смеси молекул амилозы и амилопектина, и относительное соотношение этих полимеров определяет различные химические свойства гранул крахмала (van der Maarel et al ., 2002). Амилоза (составляющая 10–30% массы гранул) представляет собой линейный полимер, состоящий из до 6000 единиц глюкозы, связанных гликозидными связями α-1,4 (de Souza and de Oliveira Magalhães, 2010). Амилоза имеет единую спиральную структуру, которая случайным образом расположена внутри аморфной ламели и связана с функциональными свойствами крахмала. Напротив, амилопектин представляет собой разветвленный полимер, который состоит из коротких линейных цепей из 10–60 единиц глюкозы, соединенных α-1,4-связями, и связанных боковых цепей α-1,6 из 15–45 единиц глюкозы (Streb and Zeeman, 2012). .
Крахмал в нативном состоянии называется сырым или немодифицированным (гранулированным) крахмалом. Гранулы сырого крахмала имеют плотно уплотненную поликристаллическую структуру, которая сопротивляется ферментативному гидролизу (Lin et al ., 2011). Гранулы крахмала состоят из чередующихся полукристаллических и кристаллических оболочек, и большая часть амилопектина, который образует полукристаллические оболочки, находится в форме двойной спирали, а не в кристаллической форме. Крахмал гидролизуется до более простых углеводов, называемых декстринами, при этом эквивалент декстрозы (DE) указывает на долю гликозидных связей, которая была гидролизована.Например, мальтодекстрин (DE 10–20) представляет собой олигосахарид, используемый в качестве наполнителя и загустителя в пищевой промышленности, кукурузные сиропы (DE 30–70) используются в качестве подсластителей, а декстроза (DE 100) доступна в виде коммерческой глюкозы, полученной полный гидролиз крахмала (Fellows, 2016).
При низких температурах (<50 ° C) гранулированный крахмал находится в твердой фазе и нерастворим в водной среде (Cinelli et al ., 2015). Однако гранулы крахмала претерпевают необратимую клейстеризацию при нагревании выше определенной температуры.Температура клейстеризации крахмала определяется распределением и длиной цепи амилопектина (Jane et al ., 1999) и характеризуется фазовым переходом гранул крахмала из упорядоченного состояния в неупорядоченное (Ubwa et al ., 2012). Разрыв водородных связей приводит к поглощению воды и набуханию молекул, что приводит к нарушению кристаллической структуры крахмала. Происходит выщелачивание амилозы из молекулы крахмала, что приводит к образованию растворимого крахмала, который легко гидролизуется (Daniel et al ., 2000). Условия клейстеризации крахмала определяются типом растения, степенью сшивки амилопектина, а также содержанием соли, сахара и липидов. Большинство крахмалов A-типа, которые обычно встречаются в зерновых крахмалах (таких как пшеница и кукуруза), образуются в теплых и сухих условиях (Ai and Jane, 2015). Напротив, крахмалы B-типа (кристаллиты B-типа) обнаруживаются в клубнях, бананах и крахмале с высоким содержанием амилозы, они образуются в холодных и влажных условиях и имеют более низкую температуру желатинизации ( T 0 из 58.2 ° C), чем крахмалы A-типа (Liu, 2005).
Гидролиз сырого крахмала происходит значительно медленнее, чем гидролиз растворимого крахмала, и для его гидролиза требуются более высокие нагрузки фермента. При сравнении четырех наиболее экономичных нативных крахмалов на предмет их чувствительности к гидролизу они были заказаны (в порядке убывания) кукуруза ≥ пшеница> маниока> картофель (Szymanowska-Powałowska et al ., 2012). Неэффективный катализ может быть результатом различных факторов, включая физическую архитектуру (степень разветвления), молекулярную структуру твердого субстрата и молекулярную конфигурацию фермента.Другие факторы, влияющие на гидролиз крахмала в его гетерогенной фазе, включают диффузию фермента в среде, доступность субстрата, распознавание субстрата, ведущее к адсорбции, образование комплекса фермент-субстрат и каталитическое действие (Gallant et al ., 1997; Tawil ). et al ., 2011).
Образование крахмала в клетках растений
Реферат
Богатые крахмалом культуры составляют основу нашего питания, но растениям еще предстоит раскрыть все свои секреты того, как они производят это жизненно важное вещество.Большой прогресс был достигнут в изучении как сельскохозяйственных культур, так и модельных систем, и мы приближаемся к пониманию ферментативного механизма, ответственного за создание массивных нерастворимых гранул крахмала, обнаруженных в тканях растений. Здесь мы суммируем наше текущее понимание этих биосинтетических ферментов, подчеркивая недавний прогресс в выяснении их конкретных функций. Тем не менее, во многих отношениях мы только прикоснулись к поверхности: остается много неуверенности в том, как эти компоненты работают вместе и контролируются.Мы отмечаем недавние наблюдения, предполагающие значительную степень гибкости во время синтеза крахмала и то, что ранее не подозреваемые неферментативные белки могут иметь значение. Мы пришли к выводу, что исследование крахмала еще не является зрелой темой, и что новые экспериментальные и теоретические подходы будут важны для продвижения в этой области.
Ключевые слова: Arabidopsis thaliana , амилопектин, амилоза, фосфорилирование белков, образование белковых комплексов
Введение
Крахмал представляет собой нерастворимый неструктурный углевод, состоящий из полимеров α-глюкозы.Он синтезируется растениями и водорослями для хранения энергии в плотной, осмотически инертной форме. Крахмал имеет большое значение для человека: он служит основным источником углеводов в сбалансированной диете и возобновляемым сырьем для промышленности. Например, крахмал широко используется в качестве загустителя и текстурирующего агента в обработанных пищевых продуктах, поскольку он желатинизируется с образованием паст при нагревании в воде. Крахмальные пасты также находят бесчисленное количество применений в непищевом секторе, например, в производстве бумаги и картона [1, 2], биоразлагаемых пластиков и упаковочных материалов [3], среди прочего.
В зависимости от его биологических функций крахмал часто подразделяется на два типа: переходный крахмал и запасной крахмал. Крахмал, который синтезируется в листьях непосредственно из фотосинтатов в течение дня, обычно определяется как временный крахмал, поскольку он разлагается следующей ночью для поддержания метаболизма, выработки энергии и биосинтеза в отсутствие фотосинтеза. Если это ночное поступление углеводов сокращается — например, у мутантов с нарушенным синтезом крахмала — растения растут медленнее и испытывают острый голод [4].Крахмал в нефотосинтезирующих тканях, таких как семена, стебли, корни или клубни, обычно хранится в течение более длительных периодов времени и рассматривается как запасной крахмал. Ремобилизация происходит во время прорастания, прорастания или возобновления роста, опять же, когда фотосинтез не может удовлетворить потребность в энергии и углеродном скелете для биосинтеза. Также мутанты с нарушением биосинтеза запасного крахмала часто оказываются в невыгодном положении, а мутантные семена с низким содержанием крахмала или без него могут даже быть нежелательными [5, 6]. Именно этот запасной крахмал мы потребляем в пищу и в качестве экстракта для промышленного использования — на него может приходиться 70–80% сухого веса зерен пшеницы и корней маниоки [7, 8].
Крахмалы из разных растительных источников различаются по своим функциональным свойствам (например, температуре начала желатинизации, конечной вязкости пасты, образованию двухфазных паст или липкости пасты) и, следовательно, по их конечному использованию. Это изменение происходит из-за различий в структуре крахмала, такой как размер гранул крахмала, их состав и молекулярная архитектура составляющих полимеров [9]. Тем не менее, экстрагированный крахмал часто необходимо модифицировать с использованием дорогостоящих и иногда вызывающих отходы химических, физических или ферментативных обработок для придания или улучшения требуемых функциональных свойств [10].Структура крахмала также влияет на его усвояемость в кишечнике. Продукты с пониженной усвояемостью (резистентный крахмал), такие как крахмалы с высоким содержанием амилозы, все больше ценятся из-за их полезного для здоровья эффекта, потенциально служащего профилактической мерой против таких состояний, как колоректальный рак и диабет [11]. Понимание биосинтеза крахмала и его взаимосвязи со структурой и функциональностью представляет огромный интерес, поскольку представляет собой предварительное условие для целевого улучшения крахмальных культур.
В этом обзоре основное внимание уделяется механизмам биосинтеза крахмала и делается попытка предоставить широкий обзор наших текущих знаний, в то же время подчеркивая недавние достижения. Значительные шаги в наших базовых знаниях были сделаны путем анализа модельных систем, таких как растение Arabidopsis thaliana и одноклеточная зеленая водоросль Chlamydomonas reinhardtii. Хотя их крахмалы не имеют прямого промышленного значения, многие аспекты биосинтеза крахмала, по-видимому, широко сохраняются в кладе Viridiplantae.Таким образом, открытия, сделанные в этих системах, вероятно, будут иметь большое значение. Тем не менее, всегда важно помнить о клеточном и метаболическом контексте, в котором образуется крахмал. Различия в условиях между тканями и видами могут иметь сильное влияние на количество и структуру крахмала. Такие различия могут объяснить, почему в некоторых случаях разные фенотипы возникают в результате сходных генетических нарушений. В конечном итоге будет важно понимать как основной процесс биосинтеза крахмала, так и тканеспецифические факторы, которые на него влияют.
Структура крахмала
Крахмал состоит из двух полимеров глюкозы — амилопектина и амилозы, которые вместе образуют нерастворимые полукристаллические гранулы крахмала (см. [12] для более полного обзора). Оба полимера состоят из α-1,4-связанных глюкановых цепей, связанных с α-1,6-точками ветвления, но их структура и биосинтез различны. Амилопектин составляет 75–90% крахмалов дикого типа, имеет степень полимеризации (DP) ~ 10 5 и степень разветвления 4–5% (т.е.е., 4–5% его связей являются точками α-1,6-ветвления) [13]. Амилопектин составляет структурную основу и лежит в основе полукристаллической природы крахмала. Амилоза значительно мельче и слабо разветвлена [13]. Считается, что он заполняет пустоты в полукристаллической матрице, образованной амилопектином, вероятно, делая гранулы крахмала более плотными.
Строение и биосинтез крахмала. a Обзор основного пути биосинтеза крахмала. АДФ-глюкозопирофосфорилаза (AGPase) продуцирует АДФ-глюкозу, субстрат синтаз крахмала (SS).Гранулированная синтаза крахмала (GBSS) синтезирует амилозу, в то время как растворимые SS, ферменты разветвления (BE) и фермент разветвления изоамилазного типа (ISA) совместно синтезируют амилопектин. b Молекулярная структура амилозы и амилопектина (согласно кластерной модели), показывающая характер ветвления и образование вторичных структур. Закрашенные соединенные кружки представляют индивидуальные глюкозильные остатки. c Выравнивание двойных спиралей амилопектина высокого порядка. Каждое кольцо роста ( справа ) имеет толщину прибл.200–400 нм и содержит полукристаллическую и аморфную области. Полукристаллическая область состоит из чередующихся кристаллических пластинок (содержащих линейные части цепей) и аморфных пластин (содержащих большинство точек ветвления), которые складываются с периодичностью ~ 9–10,5 нм ( середина ). В зависимости от точной архитектуры амилопектина, дающего начало кластерам, двойные спирали либо образуют плотно упакованный полиморф A-типа, либо менее плотный гексагональный полиморф B-типа ( вверху ).Смесь A и B также возможна и названа полиморфом C-типа (не показан). Рисунок, составленный из частей из [30] (с разрешения Elsevier) и [284] (thearabidopsisbook.org; Авторское право Американского общества биологов растений)
Относительно простая химическая природа крахмала резко контрастирует со структурной сложностью конечного продукта. гранулы крахмала. Эта химическая простота затрудняет получение окончательной структурной информации о глюканах. Вместо этого методы, используемые для определения численности структурных единиц (например,g., количество точек ветвления или длина цепи), как правило, дают средние показатели, которые маскируют структурную неоднородность. Такие ограничения означают, что мы полагаемся на структурные модели крахмалов. Это неизбежно может повлиять на интерпретацию данных, например на предпочтения ферментов в отношении субстратов или мутантных фенотипов, а также на нашу способность однозначно приписывать определенные функции ферментов. Тем не менее, для многих ферментов данные о различных типах помогли прийти к единому мнению об их роли в синтезе крахмала.
Молекулярная структура амилопектина
Обычно считается, что частота и структура ветвления в амилопектине не случайны. В каждой молекуле составляющие цепи классифицируются в соответствии с их связью с другими цепями: A-цепи — это внешние цепи, которые сами по себе не имеют ответвлений, а B-цепи — это те, которые несут одну или несколько ветвей. C-цепь представляет собой одну B-цепь на молекулу, имеющую свободный восстанавливающий конец. В моделях амилопектина точки ветвления сосредоточены в определенных областях, из которых сегменты линейных цепей отходят в кластеры (рис.). Частотное распределение длин цепей (распределения длин цепей или CLD), выведенное из анализа разветвленных крахмалов, показывает, что большинство цепей содержат от 10 до 20 единиц глюкозы. Они считаются A- и B 1 -цепями (B-цепями, которые участвуют в образовании одного кластера). Однако существуют и более длинные цепочки, которые, как считается, образуют связи между различными кластерами. Обычно считается, что они ориентированы в той же ориентации, что и заполняющие кластеры A- и B 1 -цепи (кластерная модель [14, 15]) и обозначаются как B 2 -, B 3 -, B 4 -цепи для цепочек, охватывающих два, три или четыре кластера соответственно; [16].Однако они могут быть также ориентированы перпендикулярно кластерам (каркасная модель [17]).
Рентгеновский анализ и электронно-микроскопический анализ показывают, что кластеры складываются с периодичностью ~ 9–10 нм [18, 19]. Картины дифракции рентгеновских лучей также показывают, что соседние линейные сегменты цепи внутри кластеров образуют параллельные двойные спирали, причем каждый полный виток имеет 6 единиц глюкозы на цепь и период 2,1 нм. Двойные спирали выстраиваются в плотный полиморф A-типа или менее плотный (и более гидратированный) полиморф B-типа [20, 21] (рис.в). Также наблюдаются крахмалы, содержащие смеси полиморфов A- и B-типа, которые называются полиморфами C-типа. Полиморфы A-типа типичны для зерен злаков и полиморфы B-типа клубневых крахмалов. Однако факторы, ответственные за определение типа полиморфа, полностью не изучены.
Структуры крахмала высокого порядка
Различные микроскопические анализы предполагают уровни организации за пределами 9-нм-повтора (рис. C). Некоторые из самых ранних рисунков и легких микрофотографий гранул крахмала показали концентрические слои внутри гранул.Они были названы «годичными кольцами» из-за внешнего сходства с годичными кольцами деревьев. Обработка гранул растрескавшегося крахмала α-амилазой или кислотой, которая удаляет менее кристаллические области, и анализ с помощью сканирующей электронной микроскопии четко показывают кольца роста как повторяющуюся слоистую структуру с периодом в несколько сотен нанометров. Считается, что каждый из этих устойчивых слоев состоит из множества повторов длиной 9 нм. Предполагается, что восприимчивая аморфная область имеет более низкий порядок [22].Помимо структуры кольца роста, в полукристаллических областях крахмалов наблюдались сферические блоклеты диаметром от 20 до 500 нм [23]. Они могут представлять собой левостороннюю суперспираль амилопектина, предложенную Остергетелем и ван Брюггеном [24] на основе электронно-оптической томографии и криоэлектронографического анализа. В то время как некоторые структурные особенности крахмала широко распространены, такие как образование и упаковка двойных спиралей и наличие годичных колец, другие остаются менее изученными.Следует помнить о возможном появлении артефактов во время пробоподготовки для многих применяемых методов.
Гранулы крахмала из разных видов и тканей сильно различаются по размеру и форме: от относительно небольших частиц диаметром 0,5–2 мкм в семенах амаранта и плоских дисков в листьях арабидопсиса до гладких сфер размером до 100 мкм в клубневых корнях [25 , 26]. Гранулы содержат небольшое количество белка (обычно 0,1–0,7%), который в основном представляет собой гранулированную синтазу крахмала (GBSS), которая производит амилозу, но также и другие ферменты, синтезирующие амилопектин, такие как другие синтазы крахмала (SS) и ферменты разветвления крахмала. (BEs) [27, 28].Многие крахмалы также содержат следы липидов и фосфатных групп (ковалентно связанных в положении C6 или C3 глюкозы) [27]. Уровень фосфорилирования зерновых крахмалов чрезвычайно низкий. В крахмале листьев арабидопсиса он составляет около 0,05% (то есть примерно одна на 2000 единиц глюкозы фосфорилируется), в то время как в клубневом крахмале он может быть во много раз выше (около 0,5% в картофеле). Фосфорилирование, по-видимому, ограничивается амилопектином и обогащено аморфными участками [29]. Высокое содержание фосфатов является промышленно значимым признаком, поскольку оно связано с повышенной гидратацией гранул и пониженной кристалличностью, что дает крахмальные пасты с более высокой прозрачностью, вязкостью и стабильностью при замораживании-оттаивании ([30] и ссылки в нем).
Ферменты биосинтеза крахмала
Крахмал синтезируется в пластидах — хлоропластах в листьях или специализированных амилопластах в запасающих крахмал тканях основных сельскохозяйственных культур. Иное дело у красных водорослей и глаукофитов; их так называемый флоридовый крахмал синтезируется в цитозоле посредством пути, который, по-видимому, механистически отличается от такового у растений и зеленых водорослей [31, 32]. Синтез крахмала в растениях включает три основных ферментативных активности: во-первых, SS удлиняют невосстанавливающие концы глюкозных цепей, используя аденозин-5′-дифосфат-глюкозу (АДФ-глюкозу) в качестве донора глюкозила; во-вторых, БЭ образуют ответвления из существующих цепей посредством реакций глюканотрансферазы; и, в-третьих, ферменты разветвления (DBE) снова гидролизуют некоторые из разветвлений (рис.а). Несмотря на то, что они представлены в последовательной форме, важно воспринимать их как одновременный, взаимозависимый процесс. Ферменты биосинтеза крахмала хорошо сохраняются у разных видов растений, что предполагает общее происхождение [33]. Основной механизм биосинтеза крахмала напоминает механизм гликогена, водорастворимого α-1,4- и α-1,6-связанного полимера глюкозы, синтезируемого во многих бактериях, грибах и животных. Однако, как описано ниже, биосинтез крахмала является более сложным с точки зрения дублирования и специализации SS и BE, привлечения дополнительных ферментов (т.е.е., DBE) и другие недавно описанные белки, которые могут способствовать образованию полукристаллических гранул крахмала.
АДФ-глюкозопирофосфорилаза (AGPase) обеспечивает субстрат для биосинтеза крахмала
Синтез крахмала начинается с производства АДФ-глюкозы, субстрата для SS. В фотосинтетически активных хлоропластах листьев образование АДФ-глюкозы напрямую связано с циклом Кальвина-Бенсона через превращение фруктозо-6-фосфата в глюкозо-6-фосфат (Glc-6-P) (катализируемое фосфоглюкозоизомеразой) через глюкозу. -1-фосфат (Glc-1-P) (катализируется фосфоглюкомутазой).Затем AGPase (EC 2.7.7.27) катализирует превращение Glc-1-P и АТФ в АДФ-глюкозу и пирофосфат (PP i ). По этому пути примерно 30–50% фотоассимилятов листьев Arabidopsis распадаются на крахмал [34]. Каждая из вышеупомянутых реакций термодинамически обратима. Однако in vivo продукт последней реакции PP i метаболизируется пластидиальной щелочной пирофосфатазой, гидролизуя его с образованием двух молекул ортофосфата (P i ) [35, 36].Это делает синтез АДФ-глюкозы в хлоропласте практически необратимым. Действительно, мутанты Arabidopsis с дефицитом SSIV (описанные ниже), которые не могут использовать АДФ-глюкозу для синтеза крахмала, оказывают сильное влияние на фотосинтетический метаболизм, что связано с накоплением АДФ-глюкозы и, как следствие, истощением пула аденилата [37].
Синтез АДФ-глюкозы аналогичен в гетеротрофных тканях, где сахароза импортируется из исходных тканей и метаболизируется с образованием гексозофосфатов в цитозоле.Для биосинтеза крахмала как гексозофосфаты (обычно Glc-6-P, хотя также сообщается о транспорте Glc-1-P), так и АТФ транспортируются в амилопласт, чтобы служить субстратом для синтеза ADP-глюкозы [38, 39]. Транспорт гексозофосфата происходит в обмен на P i , в то время как транспорт АТФ происходит в обмен на ADP и P i . В эндосперме злаков этот путь отличается: здесь основная активность AGPase обнаруживается в цитозоле, а АДФ-глюкоза импортируется непосредственно в пластиду через специальный, специфичный для злаков подкласс переносчиков адениновых нуклеотидов [40–43].
Синтез АДФ-глюкозы с помощью АГФазы часто рассматривается как «обязательный этап» синтеза крахмала. Имеются убедительные доказательства того, что этот шаг регулируется как на транскрипционном, так и на посттрансляционном уровнях, что подробно рассмотрено в другом месте [44]. Вкратце, AGPase представляет собой гетеротетрамер, состоящий из двух больших регуляторных субъединиц и двух небольших каталитических субъединиц. Во многих случаях было продемонстрировано, что фермент аллостерически активируется 3-фосфоглицератом и ингибируется P i (например.г. [45, 46]). Кроме того, фермент чувствителен к окислительно-восстановительной регуляции за счет восстановления межмолекулярного дисульфидного мостика, который образуется между остатками цистеина малой субъединицы [47–49]. Считается, что вместе эти регуляторные свойства обеспечивают производство АДФ-глюкозы и, следовательно, крахмала только при наличии достаточного количества субстратов. Было сделано много попыток стимулировать поток к крахмалу путем экспрессии нерегулируемой AGPase из Escherichia coli или planta (например, [50–57]).Это привело к увеличению содержания крахмала по крайней мере в одном сорте картофеля [50], увеличению общего урожая зерна кукурузы [52, 56] и пшеницы [53] и увеличению биомассы клубневых корней в маниоке [55] (обзор см. В [58]). .
Хотя вышеупомянутый путь производства АДФ-глюкозы хорошо принят, были предложены другие механизмы производства АДФ-глюкозы (см. [34] и ссылки в нем). Однако эти альтернативные пути требуют подтверждения.
Доменная структура синтаз крахмала (SS)
SS (АДФ-глюкоза: 1,4-α-d-глюкан 4-α-d-глюкозилтрансферазы; EC 2.4.1.21) принадлежат к семейству гликозилтрансфераз (GT) 5 (CAZy [59]). Они катализируют перенос глюкозильной части АДФ-глюкозы на невосстанавливающий конец (здесь положение C4) существующей глюкозильной цепи, создавая связь α-1,4 и удлиняя цепь. В биосинтезе крахмала участвуют пять классов SS: четыре растворимы в строме (или частично связаны с гранулами), а один почти полностью связан с гранулами. Растворимые SS (SSI, SSII, SSIII и SSIV) участвуют в синтезе амилопектина, в то время как SS, связанный с гранулами (GBSS), отвечает за синтез амилозы.Существует дополнительный предполагаемый класс SS, названный SSV, который последовательно связан с SSIV, но еще не охарактеризован функционально [60].
SSsсостоят из высококонсервативного С-концевого каталитического домена и вариабельного N-концевого удлинения (рис.). Каталитический домен является консервативным между SS и бактериальными гликогенсинтазами, которые также используют АДФ-глюкозу в качестве субстрата, и содержит как домен GT5, так и домен GT1 (CAZy; [61]). Согласно кристаллической структуре Agrobacterium tumefaciens и E.coli гликогенсинтаз, GBSSI риса и SSI ячменя, каталитический домен принимает складку GT-B, с активным сайтом в щели между двумя доменами GT [62–65]. Связывание АДФ-глюкозы, вероятно, включает один или несколько консервативных мотивов Lys-X-Gly-Gly [66–68] и другие консервативные заряженные / полярные остатки [62, 69–72]. N-концевые расширения классов SS различны. В случае SSIII и SSIV эти удлинения, как было показано, участвуют во взаимодействиях белок-белок, потенциально посредством консервативных мотивов coiled-coil [73–75].N-концевая часть SSIII также содержит три консервативных углеводсвязывающих модуля (CBMs), которые участвуют в связывании субстрата [76, 77].
Доменная структура классов синтаз крахмала (SS). SS из Arabidopsis ( At ) по сравнению с гликоген-синтазами из E. coli ( Ec ) и почкующихся дрожжей ( Sc ). Кукуруза ( Zm ) SSIIIa и SSV включены, поскольку они отличаются по своей структуре от ортологов Arabidopsis. Показаны пластидные транзитные пептиды (N-концевые синие прямоугольники ), внутренние повторы ( серые прямоугольники , RPT), углеводно-связывающие модули семейства 25 ( желтые прямоугольники , CBM), домены спиральной спирали ( зеленые прямоугольники). , C), домены гликозилтрансферазы-5 ( черные ящики, , GT5), домены гликозилтрансферазы-1 ( красных ящиков, , GT1) и домен гликозилтрансферазы-3 (, оранжевое поле, , GT3).Транзитные пептиды были предсказаны с помощью ChloroP [285], мотивы спиральной спирали с помощью Paircoil2 ([286]; значение p <0,05, минимальная длина 21 аминокислота) и всех других мотивов с помощью SMART. Обратите внимание, что длина домена и аннотация зависят от запрашиваемой базы данных. Например, домен GT3 Sc Gsy2P идентифицирован SMART как домен GT1 и был вручную переназначен как GT3 здесь [287]. В то время как Sc Gsy2p является гликозилтрансферазой семейства GT3 и использует UDP-глюкозу в качестве субстрата, все другие показанные синтазы являются гликозилтрансферазами семейства GT5 и используют ADP-глюкозу в качестве субстрата.N-концевые области SSIII, содержащие мотивы спиральной спирали и CBM, высоко консервативны среди различных ортологов, но дополнительно содержат внутренние повторы в некоторых случаях (например, в SSIIIa ячменя и пшеницы). Ортологи SSII часто показывали более слабые предсказания спиральной катушки или их отсутствие. At SSV имеет слабо предсказанный транзитный пептид хлоропласта, но не имеет домена GT1 и его функция неизвестна. SSV из большинства других видов, однако, имеют С-концевое удлинение, включая участок, который был обозначен как предполагаемый GT1-подобный домен [60]. Bar 100 аминокислот (AA)
Дупликации генов привели к появлению нескольких изоформ некоторых ферментов. Кодируемые белки имеют высокую степень сходства последовательностей, но часто по-разному экспрессируются, причем специфические изоформы преобладают в эндосперме или вегетативных тканях [78–84]. Например, для каждого класса SS в зерновых существует несколько изоформ (кроме SSI и SSV). Изоформы «а» SSII и SSIII, по-видимому, являются преобладающими изоформами в эндосперме на основании исследований экспрессии и мутантов (далее мы ссылаемся на эти «а» изоформы SSII и SSIII, если не указано иное).У других видов, включая Arabidopsis и растения, имеющие органы, заполненные запасом крахмала (например, картофель), существует только одна изоформа для каждого класса.
Гранулированная синтаза крахмала (GBSS) синтезирует амилозу.
Мутанты с пониженной или нулевой активностью GBSS, так называемые восковые линии , обычно производят меньше амилозы или вообще не продуцируют ее, например, в эндоспермах кукурузы [85, 86 ], рис [87, 88], пшеница [89], ячмень [90] и амарант [91], корни маниоки [8], картофель [92, 93], семена гороха [94], листья арабидопсиса [95] и С.reinhardtii [96, 97]. Это говорит о том, что GBSS отвечает за синтез амилозы и что никакая другая синтаза не может заменить ее в этой функции.
Вероятно, что GBSS синтезирует амилозу в гранулированном матриксе, образованном амилопектином. Мониторинг распределения амилозы с течением времени в линиях картофеля, в которых экспрессия GBSS и содержание амилозы были подавлены до низкого уровня, позволили предположить, что амилоза была более очевидной по направлению к центру гранулы крахмала и что это ядро, содержащее амилозу, растет вместе с гранулой [98 ].Важно понимать, что хотя и нерастворимы, гранулы гидратированы, и небольшие молекулы, такие как АДФ-глюкоза, очевидно, могут диффундировать в матрикс и использоваться связанными с гранулами белками [99]. Имеются доказательства in vitro, что GBSS действует скорее процессивно, чем распределительно, предпочтительно добавляя единицы глюкозы в одну и ту же цепь вместо переключения между цепями [68, 99]. Активность GBSS также сильно увеличивалась при анализе в концентрациях амилопектина, достаточно высоких для того, чтобы произошла спонтанная кристаллизация глюкана [68].Таким образом, GBSS может синтезировать амилозу, удлиняя отдельные цепи глюкана в среде, окружающей кристаллический или кристаллизующийся амилопектин. Его продукт, вероятно, хорошо защищен от ветвления, что объясняет, почему он в значительной степени линейен. Природа праймера, используемого для синтеза амилозы, полностью не выяснена. Радио-мечение гранул крахмала C. reinhardtii предполагает, что GBSS сначала удлиняет амилопектиновые цепи, а затем высвобождает эти цепи во фракцию амилозы [100].Однако у сосудистых растений это может быть иным, поскольку у Arabidopsis не наблюдалось переноса радиоактивной метки с амилопектина на амилозу [101]. Другим праймером могут быть мальтоолигосахариды, присутствие которых, как было показано, увеличивает активность GBSS и увеличивает его специфичность в отношении синтеза амилозы (в отличие от удлинения амилопектиновых цепей) как in vitro [102], так и in vivo [101].
Хотя GBSS почти полностью связан с гранулами [103], он не содержит каких-либо предполагаемых крахмал-связывающих доменов (рис.). Недавно было показано, что консервативный связывающий крахмал белок, снабженный CBM семейства 48 и длинным мотивом coiled-coil [104], взаимодействует с Arabidopsis GBSS через короткий мотив coiled-coil на GBSS [95]. Это взаимодействие было необходимо как для эффективного связывания GBSS с гранулами, так и для синтеза амилозы.
Данные свидетельствуют о том, что GBSS также способствует синтезу амилопектина. В некоторых исследованиях мутантов waxy сообщалось, что структура амилопектина слегка изменена [91, 105–108], тогда как в других исследованиях — иногда на тех же видах — она казалась нормальной [109–111].Заметным исключением является C. reinhardtii , где отсутствие GBSS вызвало изменение структуры амилопектина: C. reinhardtii содержит низкомолекулярную фракцию амилопектина с характеристиками окрашивания йода, промежуточными между характеристиками амилопектина и амилозы, которые были отсутствует у мутантов GBSS [96]. Отличительная функция GBSS C. reinhardtii по сравнению с GBSS сосудистых растений может быть объяснена наличием уникального С-концевого хвоста и многократной более высокой специфической активностью [112].Интересно, что двойной мутант C. reinhardtii , дефицитный по SSIII и GBSS, пострадал в гораздо большей степени, чем каждый отдельный мутант, поскольку он продуцировал лишь небольшие количества в основном водорастворимого глюкана, который был почти лишен цепей с (DP выше 40 [97 , 113]. Также ранние работы по синтезу крахмала из клубней картофеля предоставили доказательства потенциальной роли GBSS в синтезе крахмала. В репрессорных линиях ssIII , в которых GBSS был активирован, амилопектин имел популяцию сверхдлинных цепей и крахмала. морфология гранул изменена [114].Оба признака, вероятно, являются результатом (повышенной) активности GBSS, поскольку они были отменены, когда активность GBSS подавлялась одновременно с SSIII.
Основной синтез амилопектина: синтазы крахмала от SSI до SSIII
Исходя из мутантных фенотипов, каждый класс SS, по-видимому, играет особую роль во время синтеза амилопектина: проще говоря, SSI и SSII, как полагают, образуют короткие цепочки, заполняющие один кластер. (т.е. A- и B 1 -цепи), в то время как SSIII предлагается для синтеза более длинных цепей B, охватывающих кластер [115].Напротив, SSIV, по-видимому, меньше участвует в определении структуры амилопектина, но выполняет функцию инициации крахмальных гранул и контроля морфологии гранул [116–118]. Однако в действительности ситуация более сложная: есть случаи функционального перекрытия между ферментами [119], нарушения биосинтеза ферментами, расщепляющими крахмал [120–122], и образования комплексов между ферментами [73, 74, 123–127] . Кроме того, существуют значительные пробелы в нашем фундаментальном понимании того, как каждый класс выполняет свою предполагаемую роль на молекулярном уровне, как описано в следующих разделах.
Относительный вклад каждого класса SS варьируется в разных тканях и между видами, что, как полагают, по крайней мере частично объясняет структурные различия между крахмалом из разных источников. В эндосперме кукурузы SSI и SSIII составляют основные очевидные активности растворимых SS [128], тогда как в листьях кукурузы транскрипт SSI не был обнаружен [129]. Напротив, SSII и SSIII являются основными очевидными растворимыми SS в семенах гороха и клубнях картофеля [130–132], тогда как транскрипты SSI картофеля почти исключительно обнаруживаются в листьях [133].В листьях Arabidopsis SSI является основным растворимым SS, о чем судят по оставшейся SS-активности одиночных мутантов ss , за которыми следуют SSIII и SSII [116, 134]. Несмотря на то, что он выражен на разумном уровне, SSIV, по-видимому, вносит лишь небольшой вклад в общую активность SS [116, 135]. Однако важно отметить, что оценки очевидного вклада SS могут быть смещенными; супрессия SSIII, например, часто сопровождается активацией SSI и / или GBSS, что означает, что сравнение активности общей синтазы в присутствии или отсутствии SSIII не только отражает вклад SSIII.Кроме того, сами тесты SS могут предпочтительно измерять один класс по сравнению с другим, что приводит к ошибочным оценкам их относительной активности.
Крахмалосинтаза I (SSI)
Потеря активности SSI вызывает отчетливые изменения CLD амилопектина, особенно в отношении цепей A и B 1 , составляющих кластеры. Амилопектин из эндоспермов мутантов ssI из риса (в сорте japonica , т.е. рис с неактивным SSIIa и протекающей мутацией в GBSS [136]) и супрессорных линий пшеницы [137] имеет более короткие цепи DP 6 и 7, меньшее количество цепочек DP 8–12 и больше цепочек примерно DP 18.Подобные изменения наблюдались также в крахмале листьев мутантов ssI Arabidopsis [122, 138]. Поразительно, что цепи, которые истощены у мутантов, имеют ту же длину, что и цепи, которые предпочтительно синтезируются с помощью SSI in vitro. Было показано, что SSI из кукурузы, фасоли и риса предпочитает короткие цепи (обычно DP <10) в качестве субстратов [136, 139, 140], а SSI из арабидопсиса более активен в отношении гликогена, чем амилопектина [138]. Эти находки подтверждают, что SSI удлиняет короткие цепи глюкана, происходящие из действия BE (которые в основном представляют собой DP 6), на несколько единиц глюкана (до DP примерно 8-10).Затем эти цепи, вероятно, дополнительно удлиняются SSII и, возможно, другими SS. Однако, поскольку у мутантов ssI и большинство цепей от действия BE все еще являются удлиненными, другие SS, по-видимому, лишь частично зависят от действия SSI.
Интересно, что цепи, удлиненные в отсутствие SSI, кажутся более удлиненными, увеличивая долю цепей вокруг DP 18 за счет более коротких цепей. При использовании модифицированных субстратов гликогена было сообщено, что активность усеченного на N-конце SSI кукурузы резко снижается с увеличением длины внешней цепи, в то время как его связывание с субстратом сильно увеличивается [139].Авторы предложили сценарий, в котором эти характеристики заставят SSI придерживаться удлиненных глюкановых цепей в качестве неактивного фермента, тем самым предотвращая удлинение за счет других синтаз. Действительно, захват SSI внутри гранулы до некоторой степени обычно наблюдается [128, 136, 140, 141]. Неясно, действительно ли степень связывания SSI с гранулами достаточна для блокирования значительной части его короткоцепочечных продуктов. Более того, другие данные свидетельствуют о том, что локализация SSI в гранулах зависит от связывания крахмала его партнера по взаимодействию, SSII (см. Ниже).
Более поздние данные in vitro показали, что рекомбинантный SSI арабидопсиса может фактически синтезировать цепи до DP 15 при инкубации с мальтогептаозой в качестве праймера [142] и, при содействии BE, был способен продуцировать весь спектр длин цепей, обычно присутствующих в один кристаллический слой амилопектина (т.е. цепи A и B 1 [143]). Укороченная на N-конце версия SSI ячменя даже давала цепи DP> 40 при использовании мальтогексозы в качестве праймера [144]. Таким образом, настоящая причина, по которой некоторые короткие цепи, генерируемые SSI, не удлиняются дальше в условиях дикого типа, остается неясной.
Arabidopsis SSI является редокс-чувствительным и требует восстановительных условий для активности [145]. В соответствии с кристаллической структурой SSI ячменя было предложено образование дисульфидного мостика между цистеинами 126 и 506, блокирующего активный центр [65]. Однако одиночная мутация каждого из этих остатков цистеина не делала SSI полностью редокс-нечувствительным, и мутантный белок C506S терял большую часть своей активности.
Крахмалосинтаза II (SSII)
Эффекты дефицита SSII были охарактеризованы для клубней картофеля [133, 146–148], семян гороха [149], эндосперма пшеницы [150], ячменя [151], риса ( я.е., сорт риса japonica [152, 153]) и кукурузы [154] и в листьях Arabidopsis [119, 122, 134]. Наблюдаемые фенотипы удивительно похожи и включают отчетливое изменение тонкой структуры амилопектина: наблюдается повышенное количество цепей вокруг DP 8 и сниженное количество цепей вокруг DP 18, то есть сдвиг в сторону более коротких цепей. Кроме того, мутантные крахмалы ssII часто имеют больше амилозы, измененную морфологию гранул и пониженную кристалличность крахмала. У арабидопсиса также сообщалось, что вместе с крахмалом накапливаются небольшие количества растворимого глюкана [122].
Основываясь на изменениях в CLD, кажется, что SSII удлиняет цепи примерно DP 8 (цепи, удлиненные SSI) до длины около DP 18. Эта прямая интерпретация осложняется тем фактом, что, по крайней мере, в зерновых, SSII взаимодействует с SSI и БЭ класса II (см. Раздел «Роль комплексообразования и фосфорилирования ферментов биосинтеза крахмала»). Следовательно, потеря SSII может иметь плейотропные эффекты на эти ферменты, что затрудняет оценку того, какая часть фенотипа напрямую связана с отсутствием активности SSII.Например, умеренное увеличение амилозы у мутантов ssII может быть вызвано измененной активностью BEII (см. Раздел «Специфика классов BE»). Однако вполне вероятно, что изменения тонкой структуры амилопектина вызваны отсутствием активности SSII. Во-первых, когда рекомбинантный SSII риса инкубировали с амилопектином из мутанта ssII риса , он специфически удлинял аберрантно короткие цепи, так что теперь модифицированный глюкан выглядел более похожим на дикого типа [155]. Во-вторых, потеря активности SSI на фоне мутанта ssII вызвала типичные изменения ssI , что указывает на то, что SSI все еще активен [122, 134, 136].В-третьих, изменения в амилопектине CLD аналогичны у двудольных растений, где нет доказательств образования SSII-содержащих комплексов.
Крахмалосинтаза III (SSIII)
По сравнению с SSI и SSII функция SSIII менее ясна. Его предполагаемые функции включают синтез длинных В-цепей, удлинение цепей, заполняющих кластер (частично дублирующая функция с SSII) и регулирование других ферментов биосинтеза крахмала. Кроме того, SSIII важен для инициации гранул крахмала, по крайней мере, в отсутствие SSIV.В соответствии с мнением о универсальной роли SSIII проявляется как основная активность растворимых SS во всех растениях и тканях, которые были проанализированы на сегодняшний день. Он также несет самое длинное N-концевое удлинение среди всех SS, которое несет домены связывания крахмала и предсказанные домены coiled-coil (Fig.).
Вероятно, наиболее хорошо охарактеризованная функция SSIII заключается в синтезе длинных, охватывающих кластеры B-цепей (например, B 2 , B 3 и т. Д.). Меньшее количество этих цепей наблюдалось у ssIII мутантных крахмалов из клубней картофеля [148], эндоспермов кукурузы [109, 156] и риса [157], а также у C.reinhardtii [97]. Из охарактеризованных мутантов ssIII только мутант Arabidopsis ssIII [158] и мутант ячменя amo1 не проявляют этого фенотипа (обратите внимание, что мутация amo1 не отменяет активность SSIII [159]). .
Изменения профиля короткой цепи амилопектина от мутантов ssIII [147, 148, 157, 158, 160] указывают на то, что SSIII также участвует в синтезе коротких цепей A и B. Эти изменения незначительны по сравнению с изменениями, вызванными отсутствием SSI или SSII.В отсутствие SSII, однако, дополнительная потеря SSIII значительно усиливает мутантный фенотип в зернах риса [80], клубнях картофеля [147, 148] и листьях Arabidopsis [119], подтверждая частично повторяющиеся функции между этими двумя SS. Двойной мутант ssII / ssIII Arabidopsis продуцировал крошечные количества глюканов с резко укороченными цепями, часть которых была растворима в воде [119, 122]. В клубнях картофеля комбинированная репрессия SSII, SSIII и GBSS привела к получению крахмала без амилозы с короткоцепочечным амилопектином, гели которого не подвергались ретроградации при повторных циклах замораживания-оттаивания — предпочтительная характеристика для пищевой промышленности [161].
Мутанты Arabidopsis ssIII , как сообщалось, обладают повышенной общей активностью растворимого SS [158] и производят больше крахмала в течение дня [117, 158] в листе, что позволяет предположить, что он имеет отрицательную регуляторную функцию в отношении других крахмалов -биосинтетические ферменты. Однако в других исследованиях сообщалось о незначительном снижении активности растворимых СС [117, 134] и уровней крахмала [122, 134]. Повышение уровней SSI и / или GBSS наблюдалось у эндоспермов ssIII, кукурузы [128] и риса [157] и у C.reinhardtii [97]. Поэтому возможно, что некоторые изменения в тонкой структуре амилопектина вызваны повышенными уровнями SSI. Более того, наблюдаемое увеличение содержания амилозы в крахмале ssIII из кукурузы [156, 162] и риса [111, 157] может быть связано с повышенной активностью GBSS. Повышенное действие GBSS также может быть причиной других фенотипов, связанных с дефицитом SSIII, таких как повышенное количество сверхдлинных амилопектиновых цепей и трещинных гранул [114, 157]; репрессия GBSS в дополнение к SSIII в клубнях картофеля подавляла эти аспекты фенотипа ssIII [114].
Инициирование образования крахмальных гранул: синтаза крахмала IV (SSIV) и другие факторы
На сегодняшний день мутанты с пониженной активностью SSIV описаны только у риса [80, 163] и арабидопсиса [116–118]. У риса есть две изоформы SSIV: Os SSIVa, которая слабо экспрессируется в листьях и эндосперме, и Os SSIVb, которая обычно экспрессируется выше [79, 80]. Ни одиночные репрессоры Os SSIVa или Os SSIVb [80], ни нуль-мутанты Os SSIVb (у сорта japonica ; [163]) не обнаруживают заметных изменений содержания или структуры крахмала в эндосперме семян.Напротив, нулевые мутанты единственной изоформы SSIV у Arabidopsis демонстрируют изменения суточного содержания крахмала в листьях, имея меньше крахмала в конце дня и больше в конце ночи по сравнению с растениями дикого типа, несмотря на то, что у них нормальный мелкий амилопектин. структура [116]. Мутант ssIV Arabidopsis также имеет значительные изменения в количестве и форме гранул крахмала: вместо примерно шести дискоидных гранул [164] хлоропласты мутантов ssIV имеют ноль, одну или две гранулы, которые увеличены и имеют сферическую форму. менее электронно-плотный центр [116, 118].В молодых листьях ssIV хлоропласты не содержали крахмала [118]. Сверхэкспрессия At SSIV у Arabidopsis увеличивала общее содержание крахмала в листьях, хотя об увеличении количества крахмальных гранул не сообщалось [165].
В целом, Arabidopsis SSIV явно выполняет уникальную функцию в отношении инициации и морфологии крахмальных гранул, а также степени накопления крахмала [116, 118]. Интересно, что уровни АДФ-глюкозы (субстрата для SS) у мутантов ssIV увеличены более чем в 50 раз, что позволяет предположить, что ее потребление сильно ограничено [118].Это особенно вероятно в случае хлоропластов без крахмала, где оставшиеся изоформы SS не имеют субстрата глюкана. Напротив, растения Arabidopsis, имеющие SSIV в качестве единственного растворимого SS (т. Е. мутантов ssI / ssII / ssIII ), продуцируют такое же количество гранул на хлоропласт, что и растения дикого типа, несмотря на то, что в целом они имеют лишь небольшое количество аберрантного крахмала [117]. Было бы интересно посмотреть, есть ли аналогичные воздействия на крахмал листа в мутантных и репрессорных линиях риса ssIVb .
Анализ нескольких мутантов SS у Arabidopsis показывает, что SSIII (но не SSI и SSII) требуется для достижения некоторого синтеза крахмала в отсутствие SSIV: мутанты Arabidopsis ssIII / ssIV почти полностью не способны синтезировать крахмал и проявляют хлороз и задержку роста. рост [37, 117].Несмотря на сильно сниженную скорость фотосинтеза, эти мутанты по-прежнему накапливают примерно в 100 раз больше АДФ-глюкозы, нарушая баланс аденилатного пула и, возможно, объясняя наблюдаемые плейотропные дефекты [37]. Последствия потери SSIIIa и SSIVb в рисе japonica были совершенно иными, хотя все же поразительными. Двойной мутант ssIIIa / ssIVb все еще был способен продуцировать значительные количества крахмала эндосперма, но накапливал сферические и рыхлые гранулы вместо обычно наблюдаемых полиэдрических гранул сложного типа [163].Этот фенотип сопровождался неровной поверхностью амилопласта и измененным галактолипидным составом мембран. Изменения в последнем ранее были вовлечены в переход от гранул простого к сложному типу у мутанта кукурузы opaque5 [166]. Toyosawa и его коллеги предположили, что SSIVb может быть важным для структурной целостности пока еще не охарактеризованных мембраносодержащих структур, подобных перегородке [163], которые могут формировать плесень внутри амилопластов для придания крахмалу многогранной формы [167].Возможные причины фенотипических различий между ssIIIa / ssIVb рисом и ssIII / ssIV Arabidopsis включают присутствие других изоформ SSIII и SSIV в эндосперме риса и / или механистические различия в хранении и переходном образовании крахмальных гранул.
Функция SSIV и механизмы определения количества и формы гранул остаются загадочными. Биосинтез гликогена у других организмов начинается с самогликозилирования либо гликогенсинтаз (в бактериях; [168]), либо специализированных белков, называемых гликогенинами (в эукариотических клетках; [169, 170]).Введение самопрайминга гликогенсинтазы A. tumefaciens в двойные мутанты Arabidopsis ssIII / ssIV действительно восстановило инициацию множественных крахмальных гранул на хлоропласт, но морфология гранул оставалась такой же, как у ssIV [118]. Попытки подтвердить такую самопрайминговую активность в рекомбинантном At SSIV (и At SSIII) не увенчались успехом [117], хотя было обнаружено, что At SSIII катализирует образование непраймированного глюкана в присутствии АДФ-глюкозы [117].Функция автоглюкозилирования может не потребоваться для объяснения инициации полимеров крахмала в фотоавтотрофных тканях, если идеи о синтезе мальтозы de novo во время фотосинтеза подтвердятся [171, 172]: все четыре растворимых SS из Arabidopsis способны удлинять мальтозу и дольше. мальтоолигосахариды (но не глюкоза) [142].
Arabidopsis SSIV, как сообщается, взаимодействует с двумя белками, ассоциированными с пластоглобулами, фибриллинами FBN1a и FBN1b [75]. Это взаимодействие зависело от присутствия некаталитического N-конца SSIV, предположительно содержащего мотивы спиральной спирали (рис.). Исследования локализации SSIV показывают, что это стромальный белок, слабо нацеленный на отдельные точки внутри хлоропласта — первоначально предполагалось, что это края гранул [117]. Однако совсем недавно сообщалось, что он связан с тилакоидными мембранами [75]. В эндосперме риса SSIVb также имел субпластидную локализацию, преимущественно распределяясь в пространстве между гранулами крахмала, хотя отдельные пятна наблюдались редко [163]. Учитывая его предполагаемую роль в определении количества и морфологии гранул, специфическое расположение SSIV в пластиде вполне может оказаться решающим фактором.В свою очередь, понимание факторов, влияющих на его локализацию — в гранулах или мембранах — может дать совершенно новое понимание контроля образования крахмала.
Дополнительные факторы участвовали в контроле количества гранул крахмала. Ряд белков, которые самоглюкозилируются с использованием UDP-глюкозы, были описаны на протяжении многих лет (например, [173–175]), но ни один из них не имел сигналов нацеливания на пластид, но в основном участвовал в биосинтезе клеточной стенки [176]. О белках, гомологичных гликогенинам, сообщалось у Arabidopsis, и подавление белка, содержащего предсказанный сигнал нацеливания на хлоропласты, по-видимому, уменьшало накопление крахмала [177].Это наблюдение, основанное на окрашивании йодом, не было подтверждено. Интересно, что мутантные фенотипы также предполагают участие α-глюканфосфорилазы и DBE изоамилазы как влияющих на количество гранул крахмала (описано в последующих разделах). Также интересно отметить, что один из белков, участвующих в обратимом фосфорилировании глюкана, избыток крахмала 4 (SEX4), активность которого требуется для разложения крахмала в ночное время, также оказывает сильное влияние на морфологию гранул крахмала, поскольку его потеря приводит к очень сильному разложению крахмала. крупные круглые гранулы [26].Об этом фенотипе не сообщалось для других мутантов, затронутых деградацией крахмала, и его основа еще не исследована.
Классы и доменная структура ферментов ветвления (BE)
Ферменты разветвления крахмала (EC 2.4.1.18) относятся к суперсемейству ферментов α-амилазы (также называемого семейством 13 гликозидгидролаз; CAZy: [59]; недавно рассмотрено [ 178] .Они расщепляют цепь α-1,4-глюкана и переносят отщепленную часть в положение C6 единицы глюкозы из той же или другой цепи, создавая ответвление, связанное с α-1,6.При этом BE генерируют дополнительные субстраты для SS (т. Е. Невосстанавливающие концы цепей). BE имеют общую трехдоменную структуру: N-концевой домен, содержащий CBM семейства 48, центральный каталитический домен α-амилазы, характерный для членов семейства Gh23, и C-концевой домен, присутствующий в некоторых α-амилазах (рис.).
Доменная структура ферментов ветвления (BE). Классы ВЕ могут различаться по последовательности, но имеют общую доменную структуру. Изображены пластидные транзитные пептиды (N-концевой , синие прямоугольники ), углеводсвязывающие модули семейства 48 ( оранжевые прямоугольники, , CBM), каталитические домены семейства α-амилазы (, черные прямоугольники, , AMY) и все-β домены, обычно обнаруживаемые на С-конце членов семейства α-амилаз (, синие прямоугольники, , AMY_C).Прогнозы доменов были получены, как показано на рис. Домены coiled-coil не были предсказаны с настройками, описанными на рис. Структура домена сохраняется между классами и ортологами, хотя длина предсказанного каталитического домена семейства α-амилазы варьировала. Поскольку арабидопсис не имеет класса I BE, для сравнения включен BEI кукурузы ( Zm BEI), а также BE гликогена из почкующихся дрожжей ( Sc Glc3p) и E. coli ( Ec GlgB). На BE1 является предполагаемым BE класса III, в котором отсутствует CBM48, и еще не было показано, что он демонстрирует активность ветвления. Bar 100 аминокислот (AA)
В соответствии с сходством последовательностей БЭ делятся на ферменты класса I (или семейство B) и класса II (семейство A). Ферменты класса I обычно встречаются в виде единой изоформы у planta, за исключением Arabidopsis, который не содержит BE класса I [179]. Класс II представлен одним геном у картофеля и гороха, но две изоформы — BEIIa и BEIIb — встречаются в злаках. Они имеют различные паттерны экспрессии: в кукурузе, рисе и ячмене экспрессия BEIIb ограничена зерном, тогда как BEIIa обнаруживается во всех тканях, но часто на более низких уровнях [180–183].Arabidopsis также имеет два БЭ типа II (ВЕ2 и ВЕ3), оба из которых экспрессируются в листьях и кажутся в значительной степени функционально избыточными [179].
Гомология белковых последовательностей выявила предполагаемый третий класс БЭ. Гены, принадлежащие к этому классу III, были обнаружены у Arabidopsis (BE1; [179]), риса, тополя и кукурузы (BEIII; [78, 184]. Несмотря на высокую гомологию внутри этого класса (около 60%), белки разделяют только около 30% идентичности с БЭ из класса I и II [184]. На сегодняшний день функциональный анализ этого предполагаемого БЭ был проведен только на Arabidopsis, где сначала сообщалось, что нокаут-мутанты имеют фенотип дикого типа [179], а затем должны быть неприкосновенный [185].Ни одна из групп не сообщала об активности ветвления. Необходимы дальнейшие исследования, чтобы выявить каталитическую активность и биологическую функцию этих белков.
Специфические классы ферментов ветвления (BE)
Считается, что конкретное размещение ветвей BE является основным фактором, определяющим кластерную структуру крахмала (то есть обогащение ветвей в определенных, регулярно расположенных областях). Однако из-за наших ограничений в оценке фактического распределения точек ветвления, исследования специфичности изоформ BE часто больше фокусировались на длине перенесенных цепей, а не на их размещении, в первую очередь путем сравнения CLD глюканов до и после модификации in vitro. пользователя BE.Соответственно, BE класса I имеют тенденцию переносить несколько более длинные цепи, чем BEII, и более активны по отношению к амилозе, тогда как BE класса II обычно предпочитают амилопектин [186–190]. Кроме того, BEI из риса модифицировал амилопектин, ограничивающий фосфорилазу, что позволяет предположить, что этот BEI может переносить уже разветвленные цепи [190, 191]. Получение ограничивающих фосфорилазу CLD после воздействия БЭ позволяет также анализировать модификации внутренних структур ветвления [191]. Это показало, что все рисовые БЭ производят относительно широкое распределение расстояний между новыми и старыми ветвями [191].Напротив, гликоген BE из E. coli ( Ec GlgB), передавая цепи аналогичной длины растительным BEII, отличался тем, что он предпочтительно помещал новые ветви всего в трех единицах глюкозы от существующей точки ветвления [191] . Этот структурный признак также наблюдался, когда Ec, GlgB экспрессировался в линии Arabidopsis, дефицитной по своим эндогенным БЭ (двойные мутанты be2 / be3 ) [192]. Тем не менее, Ec GlgB восстанавливали синтез нерастворимых глюканов [192, 193], хотя и в меньших количествах и в виде частиц неправильной формы.
Мутанты BEI были проанализированы в эндоспермах кукурузы [194, 195], риса [6, 196] и пшеницы [197], а ген репрессирован в клубнях картофеля [198]. Во всех случаях делеция или уменьшение BEI приводило только к незначительным изменениям в структуре / количестве крахмала в лучшем случае, так что его функция in vivo остается неясной. Снижение большей части или всей активности BEII, однако, вызывало заметные изменения в крахмале и хорошо известном фенотипе амилоза — удлинитель ( ae ). Этот фенотип наблюдался у мутантов beIIb кукурузы [199] и риса [200], у мутантов beIIa твердой пшеницы [201], у мутантов BEII у гороха (давая мутанты с морщинистыми семенами, изученные Грегором Менделем [ 202, 203]) и в линиях двойных репрессоров BEIIa и BEIIb пшеницы и ячменя [204, 205].Фенотип ae характеризуется измененной структурой амилопектина с более длинными внешними и внутренними цепями и, в некоторых случаях, повышенным уровнем амилозы [6, 106, 202, 206–208]. Благодаря своим особым свойствам набухания и желатинизации, а также пониженной перевариваемости в пищеварительном тракте крахмал ae представляет особый интерес как для промышленности, так и для здоровья человека [11, 30]. В зернах риса уровни экспрессии BEIIb коррелировали со структурой амилопектина, а усиление репрессии BEIIb сопровождалось сменой полиморфа A на C и на полиморф B [207, 209].При сверхэкспрессии BEIIb наблюдались глюканы с более низкой кристалличностью, чем обычно, а также накапливались растворимые глюканы [209].
В клубнях картофеля репрессия BEII привела к амилопектину с более длинными цепями [210], но высокое содержание амилозы наблюдалось только при подавлении генов BE как I, так и II класса [211]. Этот крахмал с высоким содержанием амилозы также имел высокий уровень фосфатов — еще одна особенность, представляющая значительный промышленный интерес. Однако общий выход крахмала сильно снизился [211]. Недавно сообщалось, что сверхэкспрессия BEII в картофеле увеличивает количество коротких амилопектиновых цепей (в основном DP 6), снижает температуру желатинизации крахмала и снижает содержание фосфатов [212].
Некоторые мутации в BE влияют на метаболизм крахмала в листьях. Мутанты кукурузы beIIa имеют нормальный крахмал эндосперма, но в основном заменяют крахмал листа глюканом с низкой молекулярной массой и низкой степенью разветвления [213, 214]. Хотя BEIIa является преобладающим БЭ в листьях кукурузы, транскрипты БЭИ также были обнаружены в листе [214], предположительно отвечающие за остаточный синтез глюкана. У Arabidopsis мутанты, разрушенные в обоих БЭ класса II (ВЕ2 и ВЕ3), демонстрируют полную потерю активности БЭ и не способны продуцировать крахмал.Вместо этого они накапливают большое количество мальтозы [179]. Было высказано предположение, что этот водорастворимый глюкан образуется во время одновременного синтеза линейных цепей SS и деградации пластидальной α-амилазой 3 (AMY3) и / или β-амилазами.
В совокупности многочисленные исследования BE показывают, что, как и ожидалось, активность ветвления абсолютно необходима для синтеза амилопектина. Они также показывают, что специфические особенности БЭ различаются и что они значительно влияют на окончательную структуру глюкана.Более того, их активность по сравнению с SSs важна, что иллюстрируется изменением структуры глюкана при экспрессии в разной степени [209, 212]. Тем не менее, БЭ не являются единственными детерминантами для синтеза способных к кристаллизации глюканов, что продемонстрировано способностью БЭ гликогена E. coli восстанавливать синтез некоторых нерастворимых глюканов [192, 193]. Следует также иметь в виду, что БЭ образуют комплексы с другими ферментами биосинтеза крахмала, которые могут влиять на их функции (описанные ниже).
Ферменты разветвления (DBE) и обрезка глюканов
DBE растений гидролизуют α-1,6-связи и высвобождают линейные цепи. Они принадлежат к семейству гликозидгидролаз 13 (CAZy: [59]) и разделяют центральный домен α-амилазы и домен связывания крахмала с BE (рис.,). Далее их можно разделить на два типа: изоамилазы (ISA; E.C. 3.2.1.68) и лимит-декстриназа (LDA; E.C. 3.2.1.41). Эти два типа можно различить по белковым последовательностям и субстратной специфичности, поскольку только LDA могут эффективно разлагать пуллулан [215, 216], дрожжевой глюкан, состоящий из α-1,6-связанных мальтотриозильных единиц (отсюда альтернативное название фермента пуллуланаза или ПУ1).Геномы растений кодируют три класса изоамилазы — ISA1, ISA2 и ISA3 — и один LDA.
Доменная структура ферментов разветвления. Ферменты разветвления Arabidopsis ( At ISA1, At ISA2, At ISA3, At LDA) и фермент разветвления E. coli ( Ec, GlgX) разделяют доменную структуру с BE. Фермент непрямого разветвления из почкующихся дрожжей ( Sc Gdb1p) показан для сравнения. Изображены пластидные транзитные пептиды (N-концевой , синие прямоугольники ), углеводсвязывающие модули семейства 48 ( оранжевые прямоугольники, , CBM), каталитические домены семейства α-амилазы (, черные прямоугольники, , AMY) и домен Pfam. неизвестной функции, DUF3372 ( красный ящик , DUF).Прогнозы доменов были получены, как показано на рис. Домены coiled-coil не были предсказаны с настройками, описанными на рис. Там, где можно было получить полноразмерные последовательности, доменные структуры классов сохранялись среди ортологов у различных видов. Sc Gdb1p содержит три домена фермента, расщепляющего разветвления гликогена (GDE) Pfam (hGDE_N, hGDE_central и GDE_C, от N до C конца). Это характерно для эукариотических GDE, которые объединяют олиго-α-1,4 → α-1,4-глюканотрансферазу (EC 2.4.1.25) и α-1,6-глюкозидаза (EC 3.2.1.33) в рамках одного фермента и обезвоживания гликогена в двухступенчатом механизме. Bar 100 аминокислот (AA)
ISA1 и ISA2 участвуют в разветвлении во время синтеза амилопектина и, таким образом, находятся в центре внимания. Они действуют вместе как гетеромультимерный фермент, и ISA1 дополнительно образует активные гомомультимеры у некоторых видов (описанных ниже) — после этого мы используем «ISA» для обозначения как гомомерной, так и гетеромерной активности. Напротив, ISA3 и LDA в первую очередь разветвляют крахмал во время его деградации, и мутанты этих ферментов часто имеют крахмала — избыток фенотипов ( пол ) [217–220].Тем не менее, ISA3 и LDA действительно влияют на глюкан, вырабатываемый в отсутствие ISA, хотя они не могут заменить его специфическую функцию [121, 218, 221–223].
Мутанты без ISA частично замещают крахмал водорастворимым полисахаридом в эндоспермах кукурузы [224, 225], риса [226] и ячменя [227], в клубнях картофеля [228], листьях арабидопсиса [120, 218] и у C. reinhardtii [229]. С точки зрения уровня разветвления, длины волны максимального поглощения после окрашивания лица с йодом и молекулярной массы этот водорастворимый глюкан напоминает гликоген и, следовательно, был назван фитогликогеном [230].В большинстве случаев все же производится нерастворимый крахмал, хотя и с небольшими структурными изменениями. Только у C. reinhardtii и japonica риса (т.е. риса с фоном ssII ) мутанты isa1 полностью не способны продуцировать крахмал [226, 229]; во втором случае это зависит от мутировавшего аллеля ssII [231].
Распространенной моделью, объясняющей накопление фитогликогена, является модель «обрезки» [232]. Согласно этой модели, ISA удаляет лишние ответвления из вновь созданной аморфной зоны в «пре-амилопектине», так что только соответствующие цепи удлиняются SS.В отсутствие ISA большое количество разветвлений привело бы к стерическим ограничениям, отмене регулярных структур и синтезу дополнительных ламелей. В соответствии с этим фитогликоген Arabidopsis действительно обогащен ветвями, разделенными небольшими расстояниями [120]. Однако, хотя эта модель рассматривает разветвление с помощью ISA как обязательный этап, тщательное наблюдение за мутантами, не имеющими фермента, позволяет предположить, что это не является строго необходимым для создания компетентного к кристаллизации глюкана: не только большинство мутантов с дефицитом ISA вырабатывают некоторое количество крахмала, часто и то, и другое крахмал и фитогликоген производятся в одной и той же пластиде, и некоторые типы клеток / ткани оказываются мало затронутыми (например,г. пучково-влагалищные клетки из листьев Arabidopsis и кукурузы [120, 233, 234]). Streb et al. показали, что крахмал листьев арабидопсиса полностью заменен фитогликогеном у мутанта isa1 / isa2 / isa3 / lda , лишенного всех DBE [121]. Это предполагает некоторое функциональное перекрытие между DBE — результат, согласующийся с предыдущими наблюдениями на арабидопсисе [218, 223], кукурузе [217] и рисе [221, 222]. Однако дополнительная мутация ферментов, разрушающих крахмал, α-амилазы 3 (AMY3) на фоне мутантного разветвления частично восстанавливает синтез гранул крахмала, показывая, что разветвление, хотя и является частью процесса биосинтеза, не является абсолютным требованием [121].Этот результат согласуется с более ранним наблюдением, что фитогликоген склонен к разрушению ферментов [120], и предполагает, что модификации, производимые деградирующими ферментами, также служат для предотвращения образования нерастворимых гранул крахмала на образующихся молекулах. Недавно генетический анализ арабидопсиса [122] и риса [231] показал, что длина цепей, заполняющих кластер, также влияет на то, образуются ли нерастворимые глюканы в отсутствие ISA; относительное увеличение длинных цепей способствует образованию нерастворимых глюканов, тогда как обогащение более коротких цепей полностью их устраняет.Например, двойные мутанты ssI / isa имеют более длинные цепи A и B 1 и продуцируют значительно больше крахмала, чем одиночные мутанты isa [122] (рис.). Таким образом, кажется, что разветвление облегчает или ускоряет кристаллизацию глюканов, а не формально делает это возможным. Сама кристаллизация в твердых гранулах может, в свою очередь, защитить глюканы от преждевременной деградации амилазами [120, 121].
Модуляция фенотипа мутанта isa синтазами крахмала.Просвечивающие электронные микрофотографии хлоропластов арабидопсиса в конце дня. Растения дикого типа (WT) содержат гранулы крахмала ( черных стрелок, ) в мезофилле и эпидермисе. Мутант isa1 / isa2 накапливает преимущественно фитогликоген ( белые стрелки ) в мезофилле, но все же производит крахмал в эпидермальных клетках. Дополнительная потеря SSI восстанавливает образование гранул крахмала в мезофилле, скорее всего, из-за относительного увеличения доли более длинных, заполняющих кластеры цепей, которые способствуют кристаллизации, несмотря на аберрантное разветвление.Напротив, дополнительная потеря SSII полностью отменяет синтез нерастворимых глюканов даже в эпидермальных клетках. Это, вероятно, происходит из-за относительного увеличения более коротких цепей в отсутствие SSII, нарушая образование вторичных и третичных структур. Epi эпидермальная клетка, Pal палисадная клетка, Bars 1 мкм. Адаптировано из [122], Авторское право Американского общества биологов растений (http://www.plantphysiol.org)
Интересно, что недавние исследования показывают, что сама ISA может действовать разрушающим образом при некоторых обстоятельствах.Во-первых, экспрессия At ISA1– At ISA2 в E. coli [235] и S. cerevisiae (BP и SCZ, неопубликованные данные) препятствовала накоплению гликогена вместо образования более амилопектин-подобного глюкана. . Во-вторых, гораздо меньше глюканов обнаружено у двойного мутанта ssII / ssIII Arabidopsis по сравнению с тройным мутантом ssII / ssIII / isa [122]. Примечательно, что глюканы, полученные в ssII / ssIII растениях, показали структурное сходство с гликогеном в том, что они имели гораздо более короткие цепи, чем амилопектин дикого типа, и частично были обнаружены в водорастворимой фракции [122].Эти данные показывают, что для разветвления с помощью ISA для ускорения кристаллизации требуется соответствующий глюкановый субстрат — преамилопектин с адекватной длиной цепи и / или паттерном разветвления. Если структура глюкана несовместима с (быстрой) кристаллизацией при расщеплении ветвей (например, из-за недостаточно длинных цепей), постоянная доступность может вызвать избыточное расщепление разветвлений и, в конечном итоге, деградацию глюкана.
Чисто разрушающее действие ISA может также объяснить образование множества мелких гранул / частиц у мутантов isa [227, 228, 236].Предполагается, что у Arabidopsis этот фенотип является вторичным эффектом, возникающим в результате усиленного α- и β-амилолитического разложения растворимых глюканов: это может давать небольшие растворимые олигосахариды, которые могут служить праймерами для синтеза глюкана или сайтами зародышеобразования для гранул крахмала [ 121]. Однако небольшие разветвленные олигосахариды, возможно, представляющие собой самую раннюю стадию нормального синтеза глюкана, также могут быть отличными мишенями для разветвления под действием ISA, как упоминалось выше. Удаление ветвей и удаление таких глюканов может ограничить пул потенциальных центров зародышеобразования для гранул крахмала.
Отличительные функции гомо- и гетеромультимеров изоамилазы
В зависимости от вида ISA1 действует либо исключительно в гетеромультимерных комплексах, образованных между ISA1 и ISA2, либо как гетеромерные, так и гомомерные комплексы. Мутанты Arabidopsis isa1 , isa2 и isa1 / isa2 обнаруживают идентичные фенотипы [120]. Более того, At ISA1 и At ISA2 имеют высокую нативную молекулярную массу, взаимодействуют друг с другом и стабильны только в присутствии своего партнера [120, 235].Каталитическая активность наблюдалась только тогда, когда присутствовали как At, ISA1, так и At ISA2 [235, 237]. Эти данные убедительно свидетельствуют о том, что ISA1 и ISA2 образуют облигатный гетеромультимерный фермент. ISA2 несет в себе замены в аминокислотных остатках, важных для катализа, что, вероятно, делает его некаталитическим. Следовательно, ISA1, вероятно, будет каталитической субъединицей, при этом ISA2 требуется для стабильности фермента и, возможно, для его специфичности и / или регуляторных свойств [235, 238, 239].Точно так же у картофеля был обнаружен только гетеромультимерный комплекс ISA1 и ISA2 [240], хотя ISA1 картофеля был активен сам по себе, когда рекомбинантно экспрессировался как S-tagged белок в E. coli [238].
У других видов очевидно, что ISA1 также действует как гомомультимер. Один или несколько гомомерных комплексов ISA1 были обнаружены в дополнение по крайней мере к одному комплексу ISA1 / ISA2 в зернах риса и кукурузы и в C. reinhardtii [240–242]. В недавней кристаллической структуре C.reinhardtii ISA1 имеет удлиненную структуру и спарен с др. ISA1 хвостом к хвосту, хотя нельзя исключить существование высших мультимеров [242]. Мутанты кукурузы и риса isa2 , которые имеют только гомомер ISA1, обладают сходным синтезом крахмала в эндосперме, как и растения дикого типа, что позволяет предположить, что гомомеров ISA1 достаточно [241, 243]. Однако на фоне мутанта ssIII потеря ISA2 привела к накоплению фитогликогена в зернах кукурузы [160] — интригующее, хотя и необъяснимое наблюдение.Также сообщалось, что гетеромультимер ISA1 / ISA2 является более термостабильным и имеет более высокое сродство к фитогликогену, чем один ISA1 [240]. Таким образом, как каталитические свойства, так и стабильность ISA1, по-видимому, зависят от ISA2, и как гомомерные, так и гетеромерные ISA могут потребоваться для оптимального синтеза амилопектина, по крайней мере, в некоторых условиях.
Роль комплексообразования и фосфорилирования ферментов биосинтеза крахмала
За последнее десятилетие появилось все больше свидетельств образования комплексов между ферментами биосинтеза крахмала и фосфорилирования этих ферментов.Недавно были идентифицированы два дополнительных комплекса, которые отличаются тем, что они включают белки без очевидных каталитических функций (см. Раздел «Связывающие крахмал белки и другие новые белки»). В одном из ранних исследований, посвященных этой теме, коиммунопреципитация показала, что BEIIb взаимодействует с BEI и фосфорилазой крахмала в эндосперме пшеницы [123]. Обработка белкового экстракта щелочной фосфатазой устраняла эти взаимодействия, тогда как инкубация с АТФ имела противоположный эффект.Таким образом, предполагается, что фосфорилирование является предпосылкой для образования комплекса, что согласуется с наблюдением, что все БЭ пшеницы могут фосфорилироваться [123]. Позже эксклюзионная хроматография (SEC) экстрактов амилопластов эндосперма кукурузы показала, что SSII, SSIII, BEIIa, BEIIb встречаются в различной степени как немономерные формы. Тесты на парное взаимодействие с помощью иммунопреципитации, аффинной очистки и двугибридного дрожжевого теста позволили предположить, что фосфорилированный комплекс SSIII, SSII, BEIIa, BEIIb (и, возможно, SSI) и второй аналогичный комплекс без SSIII [73, 74].Между тем, Tetlow с соавторами сообщили о других зависимых от фосфорилирования комплексах в эндосперме пшеницы, в том числе о комплексе между SSI, SSII и BEIIa или BEIIb [124]. Это наблюдение было также основано на попарной коиммунопреципитации, аналогичных профилях элюирования, полученных с использованием SEC, и последующем вестерн-блоттинге поперечно-сшитых фракций.
Связь SSI, SSII и БЭ класса II, по-видимому, сохраняется, по крайней мере, в эндосперме злаков, что подтверждается также у кукурузы [125], ячменя [126] и риса [127].В семенах пшеницы и ячменя, где BEIIa и BEIIb выполняют повторяющиеся функции, этот комплекс содержит любой из них, а в ячмене, возможно, оба [124, 126]. В эндосперме кукурузы и риса, где BEIIb является доминирующим BE, BEIIa обычно не связан с комплексом [125, 127].
Знание о существовании этих белковых комплексов неизбежно поднимает вопрос об их значении. В частности, важно понимать, чем функции ферментов в комплексе различаются по сравнению с функциями их мономерных форм.Эта информация сильно повлияет на интерпретацию мутантных фенотипов и, следовательно, на наше понимание роли отдельных ферментов: если отсутствие одного фермента отменяет образование всего комплекса и изменяет активность оставшихся теперь исключительно мономерных или частично комплексных ферментов, результирующий фенотип будет результатом не только отсутствия активности фермента, но также и изменения биосинтетического аппарата в целом.
В мутантах ssII SSI и взаимодействующие BEII (s) постоянно появляются меньше во фракции, связанной с гранулами [150, 151, 244].Это также наблюдалось в связывающем крахмал мутанте SSII [245]. Более того, в отсутствие BEIIb у кукурузы в эндосперме образуется модифицированный комплекс: SSI и SSII теперь взаимодействуют с BEI, BEIIa и фосфорилазой [125, 246]. Точно так же репрессированные BEIIa или BEIIb заменялись на BEI и фосфорилазу в комплексе в ячмене [126]. В обоих случаях замена белка отражалась на профиле белков, связанных с гранулами: новые комплексные ферменты становились связанными с гранулами вместе со своими комплексными партнерами и GBSS [125, 126].Это говорит о том, что белки в комплексе обладают более высоким сродством к крахмалу. Тем не менее, стоит отметить, что и BEIIb, и SSI могут связывать крахмал самостоятельно [139, 245, 246], и остается неясным, требуется ли тесная ассоциация или даже захват для оптимальной активности.
Потенциальная биологическая функция для образования комплекса может заключаться в передаче субстратов от одного фермента к другому — вновь созданная ветвь, возникающая в результате действия BE, может быть непосредственно удлинена сначала SSI, а затем SSII, тем самым повышая общую эффективность процесса.Также возможно, что комплекс придает ферментативную специфичность; например, стерические ограничения всего комплекса могут определять длину цепи, которая передается ВЕ, или где именно размещается новая ветвь. Тетлоу и Эмес [115] далее предположили, что образование комплексов может регулировать активность ферментов аллостерически и / или может защищать растущий глюкан от разложения ферментов. Тем не менее, ни одна из этих гипотез еще не проверена.
Сходным образом роль фосфорилирования BE и некоторых SS [28] остается загадочной.Помимо стимулирования образования комплексов, фосфорилирование также увеличивает активность стромальных BEIIa и BEIIb у пшеницы [123]. Мутация консервативных сайтов фосфорилирования BEIIb кукурузы на серин резко снижает активность рекомбинантного белка in vitro [247]. Поскольку эта активация происходит в отсутствие его партнеров по взаимодействию, это показывает, что фосфорилирование не обязательно увеличивает активность BEIIb за счет образования комплексов. Кроме того, авторы представили доказательства участия Ca 2+ -зависимых протеинкиназ в фосфорилировании BEIIb.
Очевидно, что в будущем будет большая потребность в детальном биохимическом и структурном анализе этих белковых комплексов. В частности, создание мутантных белков, которые все еще активны, но не могут взаимодействовать со своими ферментативными партнерами, могло бы помочь в установлении важности комплексов.
Дополнительные факторы с предполагаемым влиянием на синтез крахмала
Обратимое фосфорилирование глюканов
Обратимое фосфорилирование крахмала лучше всего охарактеризовано у Arabidopsis, где оно имеет решающее значение для эффективного разложения.Считается, что фосфорилирование глюкозильных единиц в положениях C6 или C3 (осуществляемое глюканом, водной дикиназой, GWD или фосфоглюканом, водной дикиназой, PWD, соответственно [248, 249]) разрушает и дестабилизирует спиральные структуры, образованные цепями глюкана, делая их доступными для разрушающих ферментов, в частности β-амилаз [250]. Поскольку экзо-действующие β-амилазы не могут разрушать прошлые фосфатные группы, их необходимо гидролизовать одновременно с помощью фосфоглюканфосфатаз SEX4 (гидролизуется по C6 и C3 [251]) и Like-SEX4 2 (LSF2; гидролизуется по C3 [252], недавно рассмотрено в [253]).
Однако фосфорилирование глюкана не ограничивается темным периодом. Крахмал из листьев арабидопсиса, полученный в течение одного фотопериода, фосфорилируется по C6 и C3, что свидетельствует о том, что и GWD, и PWD активны в течение дня. Более того, мутанты lsf2 имеют повышенные уровни фосфорилирования C3 в конце дня, хотя эти мутанты почти полностью разрушают свой крахмал в ночное время. Это говорит о том, что фосфатные группы, введенные в течение дня, обычно также удаляются LSF2 [252].Кроме того, было показано, что количество C6-связанного фосфата в крахмале Arabidopsis в конце дня непостоянно и положительно коррелирует с фотопериодом [254]. Инкубация дисков клубней картофеля с глюкозой и радиоактивно меченным ортофосфатом привела к образованию радиоактивно меченного фосфата в крахмале, что также указывает на то, что фосфат включается во время чистого синтеза запасного крахмала [255].
Актуальность фосфорилирования во время синтеза еще не ясна. Крахмалы из мутантов Arabidopsis gwd , которые практически не содержат фосфатов, имеют нормальную внутреннюю структуру, но отличаются своей зернистой поверхностью, которая нерегулярна и имеет более короткие цепи [256].Однако эти различия могут быть вторичными эффектами неполной деградации крахмала в течение ночи и последующего синтеза глюканов поверх этих измененных гранулированных поверхностей. Мутанты Arabidopsis gwd также имеют лишь незначительный чистый синтез крахмала в течение дня. Это может означать, что отсутствие GWD в целом подавляет синтез крахмала [257]. Однако, подавляя GWD в течение продолжительной ночи, Skeffington и его коллеги получили растения без остатков крахмала и GWD, и они синтезировали количества крахмала дикого типа на следующий день [257].Таким образом, маловероятно, что GWD требуется непосредственно в процессе синтеза крахмала. Скорее, фосфорилирование глюканов GWD для эффективной деградации глюкана в течение ночи может иметь важное значение для последующего синтеза крахмала: либо напрямую путем предоставления надлежащего исходного материала, либо косвенно, влияя на распределение углерода (мутанты gwd голодают в течение ночи, что может влиять на использование фотоассимилируется на следующий день).
α-Глюканфосфорилаза — фосфолиз или синтез?
α-Глюканфосфорилаза (EC 2.4.1.1) катализирует обратимую реакцию (α-1,4-связанная глюкоза) n + глюкозо-1-фосфат ⟷ (α-1,4-связанная глюкоза) n +1 + ортофосфат. У высших растений есть два класса фосфорилазы: пластидиальный и цитозольный. Пластидный класс, названный Pho1, PhoL (из-за его низкого сродства к гликогену) или PHS1, несет вставку из 78 аминокислот рядом с сайтом связывания глюкана и показывает самое высокое сродство к линейным глюканам, особенно мальтоолигосахаридам.Напротив, цитозольный класс, названный Pho2, PhoH или PHS2, предпочитает разветвленные субстраты, такие как гликоген [258, 259].
Функция Pho1 неоднозначна до такой степени, что даже не ясно, действует ли он синтетическим или фосфоролитическим (т. Е. Разлагающим) образом in vivo. Пластидная концентрация ортофосфата обычно превышает концентрацию глюкозо-1-фосфатного многообразия [260], что способствует фосфолизу in vivo. Тем не менее было показано, что рис Pho1 катализирует удлинение цепи, а также ее укорочение при сопоставимых концентрациях in vitro [261, 262].Рисовые мутанты Pho1 продуцируют крахмал эндосперма с немного более короткими цепями, более низкой температурой желатинизации и измененной формой гранул, хотя этот фенотип не был полностью пенетрантным [261]. Дополнительный фенотип наблюдался, когда большинство зерен сморщивалось и содержало мало крахмала, когда растения выращивали при более низких температурах (20 ° C) [261]. Было высказано предположение, что в этих условиях Pho1 может работать в биосинтетическом направлении, помогая инициировать синтез крахмала путем синтеза мальто-олигосахаридных праймеров для SS, что объясняет условную потерю крахмала у мутантов.Однако репрессия Pho1 в ячмене [263] не оказывала очевидного эффекта на инициацию крахмальных гранул, хотя количество гранул не оценивалось.
Тем не менее, экспрессия генов и / или ферментативная активность Pho1 коррелирует с таковой из известных генов биосинтеза крахмала в клубнях картофеля [264] и эндоспермах кукурузы, пшеницы и ячменя [263, 265, 266], что согласуется с ролью в биосинтезе. Действуя в фосфоролитическом направлении, Pho1 может способствовать разложению мальтоолигосахаридов, созданных во время обрезки глюкана изоамилазой, с образованием глюкозо-1-фосфата для последующего обратного превращения в АДФ-глюкозу [267].Учитывая, что Pho1 взаимодействует с BEIIb и BEI в амилопластах пшеницы и кукурузы дикого типа [123, 268] и с SSI, SSIIa и BEI у мутантов beII кукурузы и ячменя [125, 126], Pho1 также может действовать. косвенно, изменяя их ферментативную активность или непосредственно на их продуктах.
В C. reinhardtii , который отличается от сосудистых растений наличием двух пластидных изоформ, PhoA и PhoB, потеря PhoB привела к снижению содержания крахмала. Оставшийся крахмал, кроме того, имел более высокое относительное содержание амилозы и структурные модификации [267].Тем не менее, этот фенотип был ограничен условиями азотного голодания, когда поток в крахмал наиболее высок. PhoB также больше похож на цитозольный, чем на пластидный класс сосудистых растений, поскольку он лишен типичной вставки последовательности и имеет низкое сродство к мальто-олигосахаридам [267].
Также было предположение, что PhoI может участвовать в разложении крахмала. Однако не было отмечено значительного влияния на содержание крахмала в листьях при репрессии листовой изоформы Pho1 у картофеля [269] или у мутанта pho1 Arabidopsis [270–272].Хотя эти отчеты указывают на существенную роль во время деградации преходящего крахмала, локальное накопление крахмала наблюдалось вокруг повреждений листьев, что привело к предположениям о специфической роли фосфорилазы в обеспечении респираторных субстратов во время стрессовых реакций [270, 273]. Совсем недавно генетические доказательства участия Arabidopsis Pho1 в деградации крахмала были получены на основе дефицита Pho1 на фоне мутантов ssIV и мутантов с нарушенным метаболизмом мальтозы [271, 272].
Белки, связывающие крахмал, и другие новые белки
В последние годы было идентифицировано несколько новых белков, которые влияют на структуру и / или количество крахмала. В некоторых случаях воздействие на крахмал может быть косвенным, в то время как в других белки могут быть частью еще не открытого аспекта процесса биосинтеза крахмала. В любом случае анализ лежащих в основе механизмов может дать новое понимание аспектов биосинтеза крахмала, которые выходят за рамки основного ферментного аппарата, таких как регуляция синтеза крахмала или влияние пластидного окружения на образование и морфологию гранул.
Пэн и его коллеги описали рисовый Floury Endosperm6 (FLO6), пластидный белок с CBM48 и способностью связывать крахмал [274]. Зерна, дефицитные по FLO6, откладывают различные гранулы крахмала с изменениями морфологии и структуры амилопектина [274]. Интересно, что FLO6 физически взаимодействовал с ISA1, который сам по себе не связывался с крахмалом. Было высказано предположение, что FLO6 обеспечивает связывание крахмала с ISA1 и что фенотип flo6 является результатом измененной функции ISA1.Это важная гипотеза для проверки, учитывая важность, приписываемую ISA в сокращении образующегося амилопектина для облегчения его кристаллизации.
Другой пластидный белок, содержащий CBM48, Protein Targeting To Starch (PTST), был описан в Arabidopsis [95, 104]. В дополнение к CBM, PTST содержит предсказанные мотивы coiled-coil. Было показано, что PTST связывается с GBSS посредством взаимодействия через мотив спиральной спирали на GBSS и что это взаимодействие позволяет GBSS связываться с гранулами крахмала: потеря PTST приводит к потере GBSS из гранул in vivo и потере синтеза амилозы [ 95].В дополнение к биотехнологической значимости (как высокое, так и низкое содержание амилозы являются желанными характеристиками крахмала), FLO6 и PTST, таким образом, представляют собой захватывающие примеры неферментативных белков, которые, по-видимому, играют важную роль в нацеливании или модуляции активности ферментов биосинтеза крахмала.
Другой класс неферментативных белков совсем недавно был обнаружен у арабидопсиса. Обозначенный Early Starvation1 (EST1), этот связывающий пластидный крахмал белок был идентифицирован, потому что у мутантов, у которых он отсутствует, обнаруживается преждевременное истощение переходного крахмала ночью [275].Это, по-видимому, происходит из-за изменения морфологии крахмальных гранул, что приводит к несоответствующей деградации крахмала в дневное время и ускоренной неконтролируемой деградации крахмала в ночное время. Напротив, растения, сверхэкспрессирующие EST1, развили фенотип с избытком крахмала, указывающий на ингибированное разложение крахмала. Хотя точная молекулярная функция белка неизвестна, генетические данные свидетельствуют против прямого ингибирования деградирующих ферментов EST1. Предполагается, что EST1 может каким-то образом способствовать правильному выравниванию молекул глюкана в гранулированном матриксе — процесс, который до сих пор считался спонтанным.Интересно, что гомолог EST1, подобный EST1 (LEST), по-видимому, выполняет противоположную роль: в то время как , чтобы мутанты не обладали фенотипом дикого типа, сверхэкспрессия приводит к фенотипу с низким содержанием крахмала, подобному таковому у мутантов est1 [ 275].
Изменения морфологии крахмальных гранул в эндоспермах злаковых также были связаны с нарушениями других клеточных процессов, включая стресс ER [276], нарушение транспортировки запасных белков [277] и изменение липидного состава мембран пластид [166].Эти наблюдения подчеркивают зависимость образования гранул крахмала от общей эффективности функционирования пластид и клеток. Локализованное в пластидах некондиционное крахмальное зерно4 (SSG4) также может попадать в эту категорию. Мутанты, лишенные этого белка, имеют увеличенные зерна крахмала и пластиды, потенциально возникающие из-за дисфункции во время развития пластид [278, 279]. Недавно сообщалось, что потеря другого стромального белка, Floury Endosperm7 (FLO7), вызывает структурные и морфологические изменения в крахмале.Этот фенотип был ограничен периферией семян риса, где также преимущественно экспрессируется FLO7, но функция FLO7 остается неизвестной [280].
Наконец, другие исследования начали раскрывать регуляторные факторы, которые, по-видимому, влияют на структуру и / или содержание крахмала, контролируя экспрессию генов биосинтеза крахмала. Например, факторы транскрипции Os bZIP58 и Rice Starch Regulator1 (RSR1), как было показано, регулируют экспрессию ряда генов биосинтеза крахмала в зернах риса, тем самым влияя на структуру крахмала и упаковку гранул [281, 282].Сходным образом Floury Endosperm2 (FLO2), ядерный белок с мотивом TPR, влияет на морфологию крахмальных гранул и экспрессию генов биосинтеза крахмала, предположительно посредством взаимодействия с факторами транскрипции [283].
Этот разнообразный набор недавно идентифицированных белков служит иллюстрацией того, что еще многое предстоит понять о биосинтезе крахмала и клеточном контексте, в котором он происходит. Стоит отметить, что большинство описанных выше белков эволюционно законсервированы у высших растений, и межвидовой анализ будет важным аспектом дальнейших исследований.
Food-Info.net: Углеводы> Крахмал
Food-Info.net> Темы> Пищевые компоненты> Углеводы
Введение
Крахмал — один из самых распространенных в природе углеводов, который присутствует во многих пищевых растениях. Крахмал на сегодняшний день является одним из наиболее потребляемых углеводов в рационе человека. Традиционные основные продукты питания, такие как злаки, корнеплоды и клубни, являются основным источником диетического крахмала. Крахмал — один из основных источников энергии в нашем рационе.
По химическому составу крахмал представляет собой полисахарид, состоящий из большого количества молекул глюкозы.
Крахмал находит множество применений не только в пищевой промышленности. Крахмал и производные крахмала широко используются в производстве бумаги, в клеях, таких как клей для обоев, косметика и даже в качестве смазки при бурении нефтяных скважин. Здесь рассматриваются только пищевые продукты.
Слово крахмал происходит от среднеанглийского sterchen, означающего «застывать», что вполне уместно, поскольку его можно использовать в качестве загустителя при растворении в воде и нагревании.
- Химия крахмала
- Амилоза
- Амилопектин
- Крахмал модифицированный
- Устойчивый крахмал
- Производные крахмала
- Растительные источники
- Приложения
Химия крахмала
Крахмал представляет собой смесь двух полимеров глюкозы и, таким образом, имеет общую химическую формулу (C 6 H 10 O 5 ) n с числом мономеров глюкозы.Молекулы крахмала могут достигать DP (степени полимеризации) от нескольких сотен до многих тысяч мономеров глюкозы.
Два полимера крахмала — это амилоза и амилопектин. В зависимости от растения крахмал обычно содержит от 20 до 25 процентов амилозы и от 75 до 80 процентов амилопектина. Обычно крахмалы зернового происхождения имеют более высокое содержание амилозы, чем крахмалы клубневого происхождения.
Таблица 1: Соотношение амилозы и амилопектина в некоторых крахмалахисточник | амилоза (%) | амилопектин (%) |
картофель | 21 | 79 |
кукуруза | 28 | 72 |
пшеница | 26 | 74 |
тапиока | 17 | 83 |
восковая кукуруза * | – | 100 |
Амилоза
Амилоза представляет собой линейный полимер глюкозы, связанный в основном связями (1-4), которые способствуют образованию спиральной структуры.Он может состоять из нескольких тысяч единиц глюкозы.
Молекулы йода аккуратно вписываются в спиральную структуру амилозы, поэтому обычным тестом на крахмал является смешивание его с небольшим количеством раствора желтого йода. В присутствии амилозы будет наблюдаться сине-черный цвет. Интенсивность окраски можно проверить с помощью колориметра, используя красный фильтр для определения концентрации крахмала, присутствующего в растворе.
Структура амилозы (источник)
Амилопектин
Амилопектин — это сильно разветвленный полимер глюкозы.Единицы глюкозы связаны линейным образом связями (1-4). Разветвление происходит с (1-6) связями, возникающими через каждые 24-30 единиц глюкозы. Молекулы амилопектина могут содержать до двух миллионов единиц глюкозы.
Простая структура амилопектина (источник)
Сложная структура амилопектина, где каждая точка представляет молекулу глюкозы. (источник)
Желатинизация и ретроградацияЖелатинизация — это процесс, при котором крахмал становится растворимым, связывает воду и образует гель.Этот процесс делает крахмал более легкоусвояемым. На этом процессе основано использование крахмала в качестве загустителя.
Крахмал набухает при нагревании и продолжает набухать, впитывая воду и показывая большую вязкость и прозрачность при повышении температуры. В определенный момент была достигнута максимальная вязкость. При дальнейшем нагревании молекулы крахмала будут расходиться дальше друг от друга, и вязкость будет снижаться. Вязкость снова постепенно увеличивается, когда раствор охлаждается, и непрерывное охлаждение делает раствор мутным.Дать раствору отстояться, он образует гель. Прочность геля определяется типом и концентрацией крахмала в продукте.
Когда гель крахмала оставляют на некоторое время, молекулы амилозы теряют воду и связываются друг с другом. Аналогичный процесс происходит, когда продукты, богатые крахмалом, например картофель, будут храниться долгое время. Этот процесс перекристаллизации крахмала называется ретроградацией.
Желатинизация и ретроградация крахмала.A: нативный крахмал, B желатинизированный крахмал, C: ретроградный крахмал
Характеристики клейстеризации (и, следовательно, прочность геля) крахмала зависят от состава крахмала; такие как соотношение между амилозой и амилопектином, присутствие жиров, фосфора и других компонентов в крахмале, а также исходное содержание воды в грануле крахмала.
Таблица 2: характеристики растворов крахмалавязкость | прозрачность | стойкость к перемешиванию | ретроградация | |
картофель | очень высокий | прозрачный | низкий-средний | средний |
кукуруза | средний | непрозрачный | средний | высокий |
пшеница | низкий-средний | мутный | средний | высокий |
восковая кукуруза | средне-высокий | прозрачный | низкий | очень низкий |
Крахмал модифицированный
Модифицированный пищевой крахмал подвергается одной или нескольким химическим или физическим модификациям, которые позволяют ему правильно функционировать при высоких температурах и / или сдвиге, часто встречающихся во время обработки пищевых продуктов.Слово «модифицированный», таким образом, относится к химической структуре молекулы крахмала и не имеет ничего общего с генетически модифицированными растениями или продуктами питания.
Пищевые крахмалы обычно используются в качестве загустителей и стабилизаторов в таких пищевых продуктах, как пудинги, заварные кремы, супы, соусы, подливы, начинки для пирогов и заправки для салатов, но имеют и множество других применений. Во многих пищевых продуктах условия не подходят для правильного желирования крахмала. Путем химической модификации крахмала изменяются свойства молекулы, и крахмал можно использовать в пищевых продуктах, например, с низким pH, или в пищевых продуктах, которые нельзя нагревать.
Модифицированные крахмалы кодируются в соответствии с Международной системой нумерации пищевых добавок (INS) и имеют следующие номера:
Таблица 3: Модифицированные крахмалы и характеристики конечного продукта (Источник)Номер | Модификация | Ограничения процесса | Технические характеристики конечного продукта |
1400 | Декстрин жареный крахмал | Сухая термообработка соляной или ортофосфорной кислотой | Конечный pH 2.5-7,0 |
1401 | Крахмал, обработанный кислотой | Обработка соляной кислотой, ортофосфорной кислотой или серной кислотой | Конечный pH 4,8-7,0 |
1402 | Крахмал, обработанный щелочью | Обработка гидроксидом натрия или гидроксидом калия | Конечный pH 5.0-7,5 |
1403 | Крахмал беленый | Обработка перуксусной кислотой и / или перекисью водорода, или | Добавленная карбонильная группа не более 0,1% |
натрия гипохлорит или хлорит натрия, или | Без остатков реагента | ||
диоксид серы или альтернативные разрешенные формы сульфитов, или | Остаточный диоксид серы не более 50 мг / кг | ||
перманганат калия или персульфат аммония | Марганец остаточный не более 50 мг / кг | ||
Крахмал, обработанный ферментами | Обработка в водном растворе при температуре ниже точки желатинизации одним или несколькими амиолитическими ферментами пищевого качества | ||
E1404 | Крахмал окисленный | Обработка гипохлоритом натрия | Карбоксильных групп не более 1.1% |
E1410 | Монокрахмалфосфат | Этерификация ортофосфорной кислотой, ортофосфатом натрия или калия или триполифосфатом натрия | Фосфат в пересчете на фосфор не более 0,5% для картофеля или пшеницы и не более 0,4% для других крахмалов |
E1412 | Дикихмал фосфат | Этерификация триметафосфатом натрия или оксихлоридом фосфора | Фосфат в пересчете на фосфор не более 0.5% для картофеля и пшеницы и не более 0,4% для других крахмалов |
E1413 | Дистахрамовый фосфат фосфатный | Комбинация обработок монострахмального фосфата и дистахмального фосфата | Фосфат в пересчете на фосфор не более 0,5% для картофеля и пшеницы и не более 0,4% для других крахмалов |
E1414 | Ацетилированный фосфат дистрахмала | Этерификация триметафосфатом натрия или оксихлоридом фосфора в сочетании с этерификацией уксусным ангидридом или винилацетатом | Ацетильные группы не более 2.5%; фосфат в пересчете на фосфор не более 0,14% для картофеля и пшеницы и 0,04% для других крахмалов; и винилацетата не более 0,1 мг / кг |
E1420 | Ацетат крахмала | Этерификация уксусным ангидридом или винилацетатом | Ацетильные группы не более 2,5% |
E1422 | Ацетилированный дистархат адипат | Этерификация уксусным ангидридом и адипиновым ангидридом | Ацетильные группы не более 2.5% и адипинатные группы не более 0,135% |
E1440 | Гидроксипропилкрахмал | Этерификация оксидом пропилена | Гидроксипропильные группы не более 7,0%; пропиленхлоргидрин не более 1 мг / кг |
E1442 | Гидроксипропилдихрамм фосфат | Этерификация триметафосфатом натрия или оксихлоридом фосфора в сочетании с этерификацией оксидом пропилена | Гидроксипропильных групп не более 7.0%; пропиленхлоргидрин не более 1 мг / кг; и остаточный фосфат в пересчете на фосфор не более 0,14% для картофеля и пшеницы и не более 0,04% для других крахмалов |
E1450 | Крахмал октенилсукцинат натрия | Этерификация октенилянтарным ангидридом | Октенилсукцинильные группы не более 3%; и остаточное содержание октенилянтарной кислоты не более 0.3% |
Крахмалы модифицируются несколькими различными способами, которые кратко описаны ниже.
Сшивка улучшает устойчивость процесса, что означает, что крахмал может выдерживать нагревание, кислые ингредиенты, перемешивание, перекачивание и упаковку. Этот крахмал не разрушается во время приготовления.
Стабилизатор помогает продукту противостоять ретроградации. Это означает, что крахмал имеет хорошую стабильность при замораживании / оттаивании и хорошую стабильность во времени.
Два других типа «модифицированного» крахмала используются в пищевой промышленности; прежелатинизированный крахмал и физически модифицированный крахмал. Эти крахмалы не нужно маркировать как модифицированный крахмал на этикетке пищевых продуктов.
Их можно обозначить как крахмал, поскольку они химически не отличаются от натурального крахмала.
Прежелатинизированный крахмал уже готовился в воде, желатинизировался, а затем сушился. Он также известен как предварительно подвергнутый тепловой обработке крахмал, предварительно желатинированный крахмал, быстрорастворимый крахмал, крахмал в холодной воде и набухающий в холодной воде крахмал.Когда порошок добавляется к холодным или теплым жидкостям, он набухает, образуя гель, становится вязким и не требует дальнейшего нагрева. Предварительно желатинизированные крахмалы могут давать гладкую или мясистую консистенцию. Обе текстуры созданы с использованием разных методов сушки. Гладкая текстура используется для супов и соусов. Мякоть мякоти используется в начинках для фруктовых пирогов и томатных начинок для пиццы.
Физически модифицированный крахмал , обработанный производителем без использования химикатов. Крахмалы можно физически модифицировать с помощью валковой сушки, экструзии, распылительной сушки, а также термической и влажной обработки.
Физически модифицированный крахмал используется, например, в пудингах и пирогах.
Крахмал устойчивый
Резистентный крахмал — это крахмал, который не переваривается в тонком кишечнике здоровых людей.
Устойчивый крахмал подразделяется на четыре типа:
- RS1 Физически недоступный или усвояемый устойчивый крахмал, например, содержащийся в семенах или бобовых, а также в необработанном цельном зерне
- RS2 Устойчивый крахмал в естественной гранулированной форме, такой как сырой картофель, мука из зеленых бананов и кукуруза с высоким содержанием амилозы
- RS3 Устойчивый крахмал, который образуется при приготовлении и охлаждении крахмалосодержащих продуктов, например, в хлебе, кукурузных хлопьях, вареном и охлажденном картофеле или ретроградной кукурузе с высоким содержанием амилозы
- RS4 Крахмалы, которые были химически модифицированы, чтобы противостоять перевариванию.Этот тип устойчивых крахмалов может иметь самую разнообразную структуру и не встречается в природе.
Тип RS | Описание | Источники питания | Сопротивление минимизировано
|
RS1 | Физически защищенный | Цельнозерновые или частично измельченные зерна и семена зернобобовых культур
| Измельчение, жевание |
RS2 | Нелатинизированные резистентные гранулы, медленно гидролизуемые α-амилазой
| Сырой картофель, зеленые бананы, бобовые, кукуруза с высоким содержанием амилозы
| Пищевая промышленность и кулинария |
RS3 | Ретроградный крахмал | Картофель вареный и охлажденный, хлеб, кукурузные хлопья, пищевые продукты с многократной влажной термообработкой
| Условия обработки |
RS4 | Химически модифицированные крахмалы за счет сшивки химическими реагентами | Продукты питания, в которых использовались модифицированные крахмалы (например, хлеб, пирожные)
| Менее восприимчивы к усвояемости in vitro |
Резистентный крахмал считается пищевым волокном, поскольку он не разлагается в тонком кишечнике и, таким образом, в неповрежденном виде достигает толстого кишечника, где частично ферментируется.
Устойчивый крахмал широко используется в хлебопекарной промышленности в качестве источника волокна в хлебе, печенье и аналогичных продуктах. Устойчивый крахмал также можно использовать в качестве ингредиента, улучшающего хрусткость пищевых продуктов, в которых поверхность продукта нагревается во время обработки. Он также используется при производстве экструдированных снеков.
В продукте увеличивается образование стойкого крахмала во время хранения, особенно при низких температурах. Это происходит главным образом из-за ретроградации крахмала и, следовательно, из-за типа RS3.
У резистентного крахмала описаны несколько свойств, способствующих укреплению здоровья. Он действует как пищевые волокна и, таким образом, положительно влияет на здоровье толстой кишки. Устойчивый крахмал полностью разрушается и ферментируется бактериями толстого кишечника. Эти бактерии производят относительно большое количество бутирата из резистентного крахмала, и бутират может предотвращать образование рака кишечника. Устойчивый крахмал также может быть легко использован больными диабетом, поскольку он не влияет на уровень глюкозы в крови.Есть также признаки того, что резистентный крахмал положительно влияет на уровень холестерина в крови и может предотвратить образование желчных камней. Наконец, есть признаки того, что резистентный крахмал увеличивает всасывание кальция и железа из толстого кишечника.
Производные крахмала
Крахмал можно гидролизовать до более простых углеводов с помощью кислот, различных ферментов или их комбинации. Степень превращения обычно количественно определяется эквивалентом декстрозы (DE), который примерно представляет собой процент разорванных гликозидных связей в крахмале.К таким пищевым продуктам относятся:
- Декстрин или мальтодекстрин , продукт из слегка гидролизованного крахмала (DE 1020), используемый в качестве наполнителя и загустителя с мягким вкусом.
- Различные глюкоза или кукурузные сиропы (DE 3070), вязкие растворы, используемые в качестве подсластителей и загустителей во многих видах обработанных пищевых продуктов.
- Декстроза (DE 100), техническая глюкоза, полученная путем полного гидролиза крахмала.
- Сироп с высоким содержанием фруктозы (кукурузный) , полученный обработкой растворов декстрозы ферментом глюкозоизомеразой до тех пор, пока значительная часть глюкозы не превратится во фруктозу.
Декстрины образуются в результате разложения амилозы и амилопектина. Таким образом, в результате получается смесь продуктов разложения обеих молекул. Существуют различные методы разложения крахмала, например, ферментами, кислотой или другими химическими веществами. Промышленное производство, как правило, осуществляется путем кислотного гидролиза картофеля или других крахмалов.Декстрины имеют разные применения, и для каждого применения используется определенная смесь. Мальтодекстрины — это класс декстринов меньшего размера.
Декстрины полностью перевариваются в тонком кишечнике и обеспечивают такое же количество энергии (калорий), как глюкоза или сахар. Их нельзя употреблять диабетикам. При производстве из пшеничного крахмала (мальто) декстрины могут все еще содержать следы глютена.
Циклодекстрины — это особый класс кольцевых декстринов, продуцируемых несколькими бактериями.Циклодекстрины находят множество промышленных применений, но редко используются в пищевых продуктах.
бета-циклодекстрин
Кукурузные или глюкозные сиропы представляют собой растворы сильно разложенного крахмала в воде. В США используется в основном кукурузный крахмал, отсюда и использование кукурузного сиропа в пищевых продуктах, производимых в США или США. В других странах можно использовать другие крахмалы. Название «глюкозный сироп» означает раствор чистой глюкозы в воде, но на практике это смесь глюкозы, мальтозы и более мелких мальтодекстринов.
Сиропы глюкозы широко используются в качестве подсластителя и структурного агента во многих пищевых продуктах. Для увеличения сладости глюкозные сиропы можно обработать ферментом глюкозоизомеразой, чтобы произвести фруктозу, которая слаще глюкозы. Эти сиропы известны как сиропы с высоким содержанием фруктозы (кукурузные) (HFS или HFCS)
.Как и декстрины, компоненты глюкозных сиропов перевариваются в тонком кишечнике и дают столько же энергии, сколько и глюкоза. Их нельзя употреблять диабетикам. При производстве из пшеничного крахмала глюкозные сиропы все еще могут содержать следы глютена.
Растительные источники
Крахмал содержится только в растениях, но не в животных. Крахмал содержится в тысячах видов растений, где его основная функция — хранение энергии для растений. Таким образом, высокие концентрации можно найти в корнях, луковицах, семенах и клубнях растений. Все основные продукты питания (пшеница, кукуруза, рис, картофель, маниока и т. Д.) Богаты крахмалом.
Гранулы крахмала (слева направо): восковая кукуруза, тапиока, картофель, кукуруза.
Без исключения крахмал встречается в природе в виде гранул.С помощью микроскопа можно определить источник крахмала, поскольку размер и форма гранул крахмала различаются для каждого вида крахмала.
Таблица 5: характеристики некоторых гранул крахмала (1 мкм = 1,10 -6 мкм)крахмал | тип | размер зерна в мкм | форма | |
диапазон | среднее | овал сферический | ||
картофель | клубень | 5 100 | 40 | овал сферический |
кукуруза | зерно | 2-30 | 15 | круглая многоугольная |
пшеница | зерно | 1–45 | 25 | двояковыпуклая круглая |
тапиока | корень | 4–35 | 25 | усеченный овал |
восковая кукуруза | зерно | 3–26 | 15 | круглая многоугольная |
По сравнению с другими крахмалом картофельный крахмал имеет высокое содержание влаги.Клубневые и корневые крахмалы обычно имеют гораздо более низкое содержание жира, чем зерновые. Эти жиры представляют собой преимущественно сложные эфиры глицерина и жирных кислот (кукуруза) или фосфолипиды (пшеница). Длинные гидрофобные (= неполярные) цепи жирной кислоты сильно взаимодействуют с молекулой крахмала, что частично определяет свойства крахмала. Крахмалы с высоким содержанием жира, такие как пшеничный крахмал, образуют мутные растворы и, как правило, менее вязкие, чем крахмалы с низким содержанием жира. Окисление жира воздухом придает крахмалу прогорклый запах и вкус, что делает эти крахмалы менее подходящими для пищевой промышленности.
Содержание азота (белки, олигопептиды, амиды и аминокислоты) в картофельном крахмале и крахмале тапиоки также ниже, чем в зерновых крахмальных продуктах. Белок придает характерный мучной запах зерновому крахмалу и отвечает за склонность к пенообразованию. Белок в крахмале нежелателен, так как при нагревании он инициирует реакции Майяра, которые вызывают обесцвечивание и формирование вкуса.
Приложения
Крахмал в его основной рафинированной форме или в виде модифицированного крахмала используется в кулинарии для загущения пищевых продуктов, таких как соусы и пудинги.Таким образом, крахмал используется в тысячах пищевых продуктов, помимо продуктов, в которых он присутствует в естественных условиях. Другие применения включают использование в качестве эмульгатора, стабилизатора, заменителя жира, для лучшей адгезии, а также в качестве средства для замутнения и глазури.
Крахмал также используется в пищевой промышленности двумя способами; один — в биоразлагаемых и съедобных покрытиях и упаковочных материалах, другой — в форме формы для приготовления кондитерских изделий. Жевательные конфеты, такие как желейные бобы, лакрица и винные жевательные резинки, не производятся с использованием форм в обычном понимании.Поднос заполняется крахмалом и разравнивается. Затем в крахмал вдавливают положительную форму, оставляя впечатление примерно 1000 желейных бобов. Затем смесь выливается в слепки и ставится в печь для застывания. Этот метод значительно сокращает количество форм, которые необходимо изготовить. Крахмал идеально подходит для этого применения, поскольку он не связывается со сладостями и может использоваться неограниченно долго.
Однако основные области применения крахмала находятся за пределами пищевой промышленности. Крахмал используется в качестве клея, например, для обоев, марок или конвертов, в качестве смазки при бурении нефтяных скважин, в бумажной промышленности для повышения прочности бумаги, в фармацевтических продуктах в качестве наполнителя, в детских подгузниках для увеличения поглощения влаги и т. Д.
Источники и литература
.