Можно годами изнурять себя диетами и часами потеть в спортзале, но результат будет незаметным. Почему одни худеют легко, а у других ничего не получается? Возможно, в этом виноваты гормоны.
Какие гормоны могут помогать нам толстеть?
Влияние гормонов на жизнь человека отражается в скорости метаболизма, наличии аппетита, потере или наборе веса и других физиологических процессах. И первое, что нужно сделать, прежде чем заниматься «самохудением», — обратиться к эндокринологу.
Лептин — гормон насыщения и полноты Низкий уровень гормона повышает аппетит, высокий — даёт команду «Хватит есть!».
Грелин — гормон чувства голода
Частые перекусы + низкий показатель гормона = тревога и стресс. Высокий уровень гормона + отсутствие еды = снижение тревоги. Итог: зверский аппетит после диеты — это замкнутый круг. Необходим контроль гормона: 4-5 приёмов, завтрак обязателен.
Эстроген и прогестерон — гормоны жиро- и водонакопления
Активизируются чаще всего у женщин после 45 лет. Низкий эстроген — накопление жира в области живота. Низкий прогестерон — задержка жидкости.
Тестостерон — гормон обмена веществ
Высокий уровень гормона — оволосение, бесплодие, набор веса.
Инсулин — гормон расщепляющий сахар
Больше сладкого = выше уровень гормона = полнеем в боках.
Тиреоидные гормоны — гормоны расщепления жиров
Гипотиреоз (низкий) — неравномерная полнота во всех местах. Гипертиреоз (высокий) — истощение организма.
Соматотропин — гормон роста
Низкий уровень гормона — затормаживает все процессы организма.
Кортизол — гормон стресса
Высокий уровень — повышает аппетит. Не заедайте проблемы и стресс. Контролируйте уровень гормона.
В 70% случаев причиной лишнего веса становится гормональный дисбаланс. Не ленитесь пройти обследование, прежде чем урезать рацион питания или записываться в тренажёрный зал. Иногда пара лишних килограммов лучше, чем испорченное здоровье.
Запишитесь к эндокринологу в клинику «Академия здоровья» по телефонам:
(8182) 43-96-96, +7 (8182) 43-97-97,+7 (911) 555-96-96,+7 (800) 550-63-98.
10 гормонов, которые влияют на нашу красоту
Наш организм – это сложная система, в которой все процессы взаимосвязаны и отлажены природой. Малейшие сбои в его работе отражаются на функционировании всей системы, в том числе и нашей внешности. Изначальные данные, такие как цвет волос и тип телосложения, записаны в ДНК-коде и не меняются в течение жизни, а то, насколько упругой выглядит наша кожа и стройным тело, во многом зависит от уровня гормонов. Главных гормонов, которые влияют на нашу красоту, десять. Мы решили выяснить, что именно делает каждый из них и как гормональный дисбаланс отражается на внешности.
Эстроген
Эстрогены – это целая группа женских половых гормонов, к которым относятся эстрон, эстриол и эстрадиол. Все они вырабатываются в яичниках, отвечают за формирование фигуры по женскому типу, развитие половых органов и регулируют менструальный цикл. «Если эстроген вырабатывается в нормальном количестве, он очень положительно влияет на внешний вид женщины, – рассказывает Ирина Вяткина, врач гинеколог-эндокринолог «Клиники Марины Рябус». – Этот гормон участвует в процессе обновления клеток всего организма, в том числе и кожи. Эстроген положительно влияет и на красоту волос: придает им блеск, сохраняет молодость и здоровье. Эстрогены поднимают настроение, располагают к флирту и кокетству, препятствуют отложению холестерина в стенках сосудов, заставляют блестеть глаза, разглаживают морщинки, делают кожу эластичной и упругой, тем самым делая женщину красивой. Кроме того, эстрогены поднимают настроение и располагают к флирту и кокетству – они способствует развитию влечения у женщин к лицам мужского пола. На фоне увеличения выработки эстрогенов улучшается координация движений и тем самым походка, меняется качество речи. Она становится более приятной для восприятия».
Эстроген по праву считается главным женским гормоном красоты. Его дефицит сказывается не только на самочувствии, но и на внешности: волосы становятся тусклыми и могут расти в ненужных местах (например, на подбородке или груди), а кожа выглядит бледной и начинает рано увядать. Именно с уровнем эстрогена связывают такой феномен, как «осеннее выпадение волос». Весной и летом уровень гормонов естественным образом повышается, а к осени – снижается. Сезонное выпадение волос в разумных пределах – это нормальное состояние организма, и нет повода переживать из-за этого.
Бывают ситуации, когда уровень эстрогена у женщины повышен. Это сопровождается нарушением половой функции (пропадают месячные или овуляция), образованием жира именно на животе и бедрах, а также предменструальным синдромом.
Прогестерон
Основная задача прогестерона – это развитие яйцеклетки и ее размещение в матке, поэтому его часто называют гормоном беременности. У небеременных женщин прогестерон также присутствует и повышается во второй фазе менструального цикла, что, увы, негативно сказывается на внешности. «Прогестерон способствует задержке жидкости в организме, повышает проницаемость сосудистой стенки, кожа становится растяжимой, повышается выделение кожного сала и появляются угревые высыпания. Может появляться одутловатость лица за счет задержки жидкости», – говорит Ирина Вяткина. Прогестерон также активно способствует запасанию жира, подготавливая нас к беременности, даже если мы ее не планируем. Поэтому во второй фазе цикла многие набирают один-два лишних килограмма. Кроме того, прогестерон снижает сопротивляемость организма инфекциям, что благотворно влияет на патогенную микрофлору кожи из-за ослабления иммунитета и зачастую приводит к образованию прыщей.
ДГЭА (дегидроэпиандростерон)
«ДГЭА – гормон, выделяемый надпочечниками. Он является предшественником половых гормонов, отвечает за половое влечение, ясный ум, твердую память, мышечную силу и физическую выносливость и улучшает настроение – все это является неотъемлемой частью молодости и красоты», – говорит Елена Шаткарь, эндокринолог резиденции красоты и долголетия GLMED. Пониженное содержание дегидроэпиандростерона негативно сказывается как на всем гормональном фоне, так и непосредственно на внешности и самочувствии. В первую очередь дефицит ДГЭА проявляется ломкостью ногтей и выпадением волос, плохим настроением и изменением продолжительности менструального цикла.
Мелатонин
Мелатонин, известный как гормон сна, влияет и на многие другие функции организма. Он участвует в работе желудочно-кишечного тракта, эндокринной и иммунной систем, отвечает за нормальное функционирование клеток мозга и защиту от свободных радикалов. Да, мелатонин – это мощный антиоксидант, уровень которого влияет на то, как кожа и остальные органы могут самостоятельно противостоять свободным радикалам, разрушающим клетки. И это еще не все. «Действие мелатонина затрагивает сжигание жира во время сна, – рассказывает Елена Шаткарь. – Выработка гормона происходит во время сна в его наиболее глубокой фазе – примерно с 12 часов ночи до 4 часов утра. Главное существенное условие для его выработки – человек должен спать в темном помещении. Критерием темноты может служить факт невозможности рассмотреть свою вытянутую руку. Любой свет – это враг мелатонина, в том числе циферблат электронного будильника, не говоря уже об экране смартфона, компьютера, телевизора или фонаре на улице. Уменьшение уровня мелатонина – это сигнал другим органам и железам, что настало время для отдыха. Яичники у женщин прекращают работу, сильно снижается уровень эстрогена, приходит менопауза. У мужчин значительно снижается количество гормона тестостерона». Таким образом, ночной образ жизни напрямую связан не только с ухудшением работы всего организма, но и с лишним весом и быстрым увяданием кожи.
Кортизол
Мелатонин влияет на выработку кортизола – гормона стресса, который синтезируется надпочечниками и помогает выжить в экстренных ситуациях. «При изменении выработки кортизола меняются углеводный обмен и выработка инсулина, что приводит к изменению внешнего вида и качества кожи. При высоком уровне инсулина происходит нарушение работы сальных и потовых желез, кожа теряет свои естественные защитные свойства, легко поражается бактериальной и грибковой инфекцией, теряет свою упругость, эластичность, снижается синтез коллагена. При повышении уровня кортизола кожа истончается и становится склонной к появлению пигментации. При нормальной выработке кортизола кожа упругая, ровная и красивая», – рассказывает Ирина Вяткина. Повышенный кортизол способствует накоплению жира в области лица и живота, поэтому справедливо выражение, что стресс мешает похудеть.
Тестостерон
Тестостерон, хоть и считается условно мужским гормоном, у женщин также вырабатывается при помощи надпочечников и половых желез. Тестостерон имеет непосредственное влияние на красоту кожи. «Он повышает скорость обновления клеток эпидермиса и увеличивает выработку коллагена за счет стимуляции соединительной ткани, производящей протеины, необходимые для синтеза коллагена, – говорит Ирина Вяткина. – С возрастом наблюдается снижение синтеза тестостерона. В результате замедляется регенерация кожи, снижаются ее защитные функции и упругость». Такие же симптомы могут наблюдаться и в молодом возрасте из-за гормональных нарушений. Повышение уровня тестостерона в норме происходит во второй фазе цикла, что вместе с прогестероном провоцирует образование прыщей. Сейчас набирает популярность тестостероновая терапия, которая улучшает качество кожи при возрастном снижении синтеза гормона. Врачи предупреждают, что самостоятельным приемом тестостерона можно сильно навредить своему здоровью – такая терапия проводится только по показаниям под наблюдением эндокринолога.
Соматотропин
Соматотропин известен как гормон роста. Его влияние на красоту и молодость сложно переоценить. «Соматотропин отвечает за рост и восстановление костей, помогает нормализовать обменные процессы, усиливает сжигание жира и помогает печени вырабатывать энергию для всего организма, – рассказывает Елена Шаткарь. – С возрастом выработка гормона роста замедляется, в связи с этим начинаются процессы старения. Повысить уровень соматотропина помогают регулярные физические нагрузки, правильное питание и полноценный ночной сон». Выработка гормона роста начинает снижаться уже после 25 лет, а к 45 годам его синтез в два раза меньше, чем в детстве. Это сопровождается снижением качества сна, нарушением аппетита и увеличением массы тела.
Тиреотропный гормон, тироксин и трийодтиронин
Тиреотропный гормон, тироксин и трийодтиронин – это все гормоны щитовидной железы, которые работают сообща. Они участвуют практически во всех обменных процессах организма, регулируют метаболизм, скорость синтеза витаминов и поддерживают нормальную функцию дыхания. «При нехватке гормонов щитовидной железы (гипотиреозе) кожа становится сухой, ногти – ломкими, выпадают волосы. А при избытке гормонов щитовидной железы (гипертиреозе) кожа становится влажной, повышается потливость, а, несмотря на здоровый аппетит и достаточный прием пищи, человек теряет в весе. Однако сброшенные килограммы совсем не радуют, так как при этом серьезно нарушается работа сердца, – говорит Юрий Потешкин, к. м. н., врач-эндокринолог клиники «Атлас». – При первых симптомах, указывающих на неполадки с функционированием щитовидной железы, следует сделать анализ крови на тиреотропный гормон. Именно он регулирует функции тироксина и трийодтиронина и отражает их уровень в организме».
Комментарий косметолога
Юлия Щербатова, врач-дерматокосметолог, челюстно-лицевой хирург, главный врач «Клиники современной косметологии Юлии Щербатовой»:
«Любой гормональный дисбаланс – состояние, которое развивалось в организме долго, и его коррекция тоже займет какое-то, зачастую довольно продолжительное время. Все это время косметолог может и должен помогать пациенту решать проблемы, с которыми тот обратился. Мы ни в коем случае не должны ждать завершения лечения, чтобы заниматься косметологией. Работа в этом случае будет, скорее, симптоматическая. Если проблемы связаны с сосудистой сеткой, мы используем лазеротерапию, если пациент жалуется на акне – лечим акне, если его беспокоит пигмент – боремся с пигментом инъекциями и лазером. Словом, действуем по показаниям и состоянию.
Типологизировать проблемы с кожей и их связь с гормональными нарушениями вряд ли возможно, потому что они могут проявляться очень по-разному. Но совершенно точно могу сказать, что коррекция гормонального фона при участии грамотного эндокринолога сделает результат более выраженным и эффективным. А вот желание обойтись одной косметологией, без плановой терапии, – это, как правило, трата денег и времени на процедуры, которые не сработают или сработают недостаточно хорошо. И дело не в самих процедурах, а в патологиях пациента. Так, например, если убрать лазером сосудистую сетку с лица, но не решить проблемы с избытком эстрогена или дефицитом прогестерона, из-за которых она часто возникает, – сосуды быстро вернутся. То же касается и гиперпигментации: казалось бы, бесследно удаленные лазером пигментные пятна будут возвращаться до тех пор, пока не придут хотя бы в относительный порядок стероидные гормоны и гормоны гипофиза, влияющие на клетки-меланоциты. Словом, при эндокринных проблемах, которые сегодня бывают у большого количества пациентов, сочетание косметологии с плановой терапией работает на порядок лучше, чем что-то одно.»
Имейте в виду, что любая попытка самостоятельно исправить гормональный дисбаланс может повлечь за собой еще большие проблемы со здоровьем. Грамотный врач-эндокринолог может вам подобрать нужный тип и дозировку гормонозаместительной терапии. Порой можно обойтись и вовсе без нее: на нарушение работы гормональной системы может влиять любое другое заболевание и даже низкий уровень витаминов и микроэлементов. В таких случаях врач будет корректировать именно первопричину.
Эндокринология и косметология. Тонкая грань проходит по коже
Взаимосвязь гормональной системы и эпидермиса пациента
Состояние кожи, волос и ногтей является результатом целой совокупности нейрогуморальных факторов, среди которых немаловажное место занимают гормоны. Именно они определяют кровоснабжение кожи, толщину эпидермиса, тип оволосения и свойства волос, секреторную активность сальных желез, крепость ногтей.
Главный гормон, отвечающий за красоту и здоровье кожи — эстроген. В переводе с греческого термин означает «живость, яркость». Он обеспечивает упругость кожи, регенерацию после повреждений, укрепляют дермальное соединение, способствует синтезу волокон коллагена и выработке гиалоурановой кислоты.
Поэтому, недостаток эстрогена приводит к быстрому увяданию кожи, что проявляется в сухости, истончении, появлению морщинок и видимости сосудистой сетки. И если после 45 лет, снижение уровня эстрогена у женщины – норма, то в более молодом возрасте недостаток этого гормона– это первый сигнал сбоя гормональной системы. Избыток эстрогена, напротив, увеличивает работу сальных желёз, делая кожу жирной и склонной к воспалительным процессам, закупориванию пор и угревой сыпи.
Однако, несправедливо было бы не упомянуть и иные гормоны, которые неустанно работают на благо женской кожи. Это –
прогестерон и тестостерон (который тоже должен присутствовать у женщины в определённом объеме). Тестостерон контролирует динамику клеточного деления, процессы дифференциации, ороговения и секреции. А еще он придает коже мягкость, обеспечивает защитные бактерицидные функции кожи и оказывает противовоспалительный эффект путем подавления действие протеолитических ферментов, связанных с деградацией коллагена. Именно этот гормон играет ключевую роль в поддержании мышечного тонуса кожи, так как он регулирует синтез коллагена, миозина, актина и других белков.
Прогестерон способствует секреции кожного сала и тормозит синтез тропоколлагена в фибробластах соединительной ткани. В современной косметологии уже активно применяется лечение угревой сыпи при помощи прогестерона. Он эффективен тем, что тормозит биотрансформацию тестостерона в 5а-дигидротестостерон (5а-ДГТ) и конкурирует с 5а-ДГТ за андрогеновые рецепторы в волосяных фолликулах и сальных железах, снижая уровень их активности. Снижение уровня прогестерона проявляется аналогичнор с низким эстроеном: морщины, увядание, сосудистая сетка, и истончение кожи. Повышение уровня прогестерона выше нормы может вести к пигментации кожи, потере тугора и упругости, отечности, возникновению воспалительных процессов. Подобные явления нередко проявляются у беременных женщин вследствие чрезмерно возрастающего уровня прогестерона.
Безусловно, каждый гормон необходим в определенной пропорции и в случае отклонения от нормы, его функции нарушаются, и он может даже начать работать против организма. Если глаза – это зеркало души, то кожа – зеркало гормональной системы. Если гормональная система в балансе — кожа выглядит сияющей и старение замедляется, а если наоборот — старение и увядание кожи происходят очень быстро.
Современная медицина опирается на доказанный многими исследованиями опыт, что клинический дефицит/избыток половых гормонов у женщины является безусловным основанием для назначения врачом гормональной терапии. Открытия ученых в области биогенетики показали, что
гормональная система человека во многом предопределена на уровне ДНК. Поэтому, при назначении гормонов, необходимо с большим вниманием относиться не только к настоящему состоянию и анализам пациента, но и к его генетическими особенностям. Некорректная гормональная терапия в неподходящих пациенту дозах может привести к возникновению серьезных побочных эффектов. Одним из опасных последствий неадекватной гормональной терапии является возникновение тромбозов.
На генетическом уровне может быть заложено множество других особенностей эндокринной системы.
Например, высокий риск развития гиперэстрогении или гиперандрогении, то есть переизбытка женских или мужских гормонов. Пример: по результатам генетического анализа по эндокринологии у пациентки выявлен повышенный риск гиперэстрогении, в совокупности с повышенным эстрогеном в реальном времени по результатам анализа крови. При этом, в генетическом анализе «Косметология» имеется высокая предрасположенность к воспалительным процессам и склонность к замедленному метаболизму витаминов группы В. Таким образом, при такой совокупности факторов у кожи очень мало шансов защититься от воспалений, угревой сыпи, акне и дерматитов.
Или, другая ситуация: в генетическом тесте по эндокринологии выявлена сниженная чувствительность рецепторов прогестерона, что означает слабое воздействие этого гормона на организм. Вероятность того, что прогестерон будет снижен в анализе крови очень высока, так как прогестерон очень чувствителен к внешними факторам: стресс, смена климата, недосып, неправильное питание и многое другое. При этом, генетический анализ «Косметология» показал генетическую склонность к слабости сосудистой сетке, сниженному метаболизму витаминов А и Е, ускоренному разрушению коллагена и снижению качества коллагеновых волокон. С таким набором практически неизбежно возникновение ранних морщинок, истонченности и сухости кожи, видимости венозной сетки.
При таком глубоком подходе к гормональному балансу и свойствам кожи конкретного человека, с учетом генетических особенностей организма, удается понять реальную причину тех или иных внешних дефектов кожи. Волшебного крема из рекламы, воспользовавшись которым можно за ночь помолодеть на десять лет и избавиться от всех кожных дефектов – не существует. Только комплексный анализ и терапия позволяют укрепить женскую красоту изнутри, существенно повышая эффективность косметологических процедур.
Вывод: к лечению дерматологического пациента необходимо подходить с разных сторон, учитывая все эпигенетические факторы. Анализировать кожу в отрыве от всего организма — малоэффективно. Для того, чтобы не просто устранить внешние дефекты кожи, а добраться до корня проблемы, следует корректировать нутрициональные и гормональные дефициты. Особое значение имеет коррекция витаминов группы В, А, Е, С омега-3 и омега-6 ПНЖК, железа и половых гормонов. Для полного понимания клинической картины, врачу необходимо провести генетические тестирования пациента и по
эндокринологии, и по косметологии. На основе результатов возможно будет сделать комплексное заключение и разработать наиболее эффективную индивидуальную программу дерматологической и гормональной терапии, позволяющую добиться наилучших результатов в совокупности с косметологическими процедурами.
Какой биоматериал можно использовать для исследования?
Венозную кровь.
Как правильно подготовиться к исследованию?
Исключить из рациона жирную пищу в течение 24 часов до исследования.
Не принимать пищу в течение 8 часов до исследования, можно пить чистую негазированную воду.
Полностью исключить прием лекарственных препаратов в течение 24 часов перед исследованием (по согласованию с врачом).
При отсутствии указаний врача исследование производится на 6-7-й день менструального цикла.
Исключить физическое и эмоциональное перенапряжение в течение 24 часов до исследования.
Не курить в течение 3 часов до исследования.
Общая информация об исследовании
Гормоны яичников, гипофиза и гипоталамуса играют важную роль в процессе наступления и поддержания беременности и естественных родов. В норме в организме женщины репродуктивного возраста происходит циклическое и синхронизированное изменение концентрации этих гормонов, что обеспечивает подготовку ее репродуктивной системы к беременности. При возникновении гормональных сбоев репродуктивная функция нарушается. В некоторых случаях гормональные нарушения могут стать причиной бесплодия. По этой причине при подготовке к беременности проводят гормональные анализы, позволяющие (1) оценить функциональный овариальный резерв и (2) исключить эндокринные причины бесплодия.
Эстрадиол – главный половой гормон женщины репродуктивного возраста, вырабатываемый яичниками. Его основной функцией является подготовка эндометрия матки к имплантации эмбриона и развитие беременности. Нарушение выработки эстрадиола яичниками приводит к ановуляторным циклам и НМЦ и в итоге к полному прекращению менструаций. Постепенное снижение уровня эстрадиола наблюдается при естественном угасании репродуктивной функции женщины при старении.
Фолликулостимулирующий гормон (ФСГ) – гормон аденогипофиза и хорошо известный показатель «репродуктивного возраста» женщины. При снижении функции яичников происходит нарастание концентрации ФСГ. Уровень ФСГ считается повышенным, если его концентрация превышает в два и более раза среднюю концентрацию ФСГ женщины репродуктивного возраста, измеренную в раннюю фолликулярную фазу.
Лютеинизирующий гормон (ЛГ) – гормон аденогипофиза, одна из ролей которого заключается в стимуляции овуляции. Так же, как и в случае с ФСГ, при снижении функции яичников уровень ЛГ возрастает. Стойкое повышение уровня ФСГ и ЛГ при снижении уровня эстрадиола свидетельствует о менопаузе. В случае наступления менопаузы в возрасте до 40 лет говорят о преждевременной недостаточности яичников (прежнее название – ранняя менопауза).
Прогестерон – это стероидный гормон, вырабатываемый в больших количествах желтым телом яичников и при наступлении беременности плацентой. Так же, как и эстрадиол, прогестерон обеспечивает подготовку эндометрия к имплантации эмбриона и способствует развитию беременности. Уровень прогестерона исследуют для косвенной оценки процесса овуляции: низкий уровень этого гормона в лютеиновую фазу (менее 3 нг/мл) говорит об отсутствии овуляции.
2. Анализы, позволяющие исключить заболевания, приводящие к бесплодию:
Заболевания щитовидной железы: Т4, ТТГ.
Как гипотиреоз, так и гипертиреоз могут приводить к нарушению овуляции и являться причиной бесплодия. С другой стороны, своевременная диагностика и лечение этих заболеваний позволяет полностью восстановить фертильность. Кроме того, исследование функции щитовидной железы также проводят для раннего выявления и лечения у будущей матери гипотиреоза (в том числе субклинического) – состояния, которое ассоциировано с развитием когнитивных нарушений у плода (крайний случай – кретинизм).
Гиперандрогения и синдром поликистозных яичников (СПКЯ): тестостерон, дегидроэпиандростерон-сульфат (ДЭА-SO4).
Хотя андрогены играют важную роль в репродуктивном здоровье женщины, их избыток может приводить к отрицательным последствиям. Кроме того, гиперандрогения является одним из маркеров дисрегуляции системы «гипофиз-надпочечники-яичники». Повышение уровня тестостерона может наблюдаться при СПКЯ, врождённой гиперплазии и опухолях надпочечников, а также гормонпродуцирующих опухолях яичника и поэтому не позволяет судить об источнике гиперандрогении. В отличие от тестостерона, повышение концентрации ДЭА-SO4 – более специфический признак, указывающий на то, что источником гиперандрогении являются надпочечники.
Опухоль гипофиза (пролактинома): пролактин.
Пролактин – это один из гормонов аденогипофиза, по структуре схожий с гормоном роста и хорионическим соматомаммотропином. Одна из функций пролактина заключается в противодействии гонадотропным гормонам ФСГ и ЛГ. Физиологическое повышение уровня пролактина наблюдается во время беременности и кормлении грудью и имеет эволюционное значение, так как приводит к временной ановуляции и в некоторой степени «защищает» женщину от наступления следующей беременности до восстановления ее организма после родов. Заболевания, приводящие к стойкому и существенному повышению пролактина, также могут стать причиной ановуляции, в этом случае – патологической. Наиболее часто причиной гиперпролактинемии являются опухоли гипофиза (пролактинома и другие) и гипотиреоз. Своевременное выявление и коррекция этих заболеваний приводит к полному восстановлению фертильности.
Важно отметить, что концентрация многих гормонов женского организма сильно зависит от фазы менструального цикла, что следует учитывать при интерпретации результата этого комплексного исследования. Это особенно важно для ФСГ, ЛГ и эстрадиола (которые предпочтительнее оценивать в начале цикла – 2-3-й день) и пролактина и прогестерона (за одну неделю до наступления менструации – на 21-й день при 28-дневном цикле). Более полная информация о состоянии здоровья женщины, планирующей беременность, может быть получена при проведении дополнительных комплексных тестов («Планирование беременности – необходимые анализы» и «Планирование беременности – обязательные анализы»).
Для чего используется исследование?
Для оценки функционального овариального резерва и исключения эндокринных причин бесплодия.
Когда назначается исследование?
При подготовке к беременности.
Что означают результаты?
Референсные значения
Важные замечания
Результат комплексного анализа следует интерпретировать с учетом клинических, лабораторных и инструментальных данных;
для получения точного результата необходимо следовать рекомендациям по подготовке к тесту.
Также рекомендуется
[40-044] Планирование беременности — здоровье партнеров (для женщин)
[40-125] Планирование беременности — здоровье партнеров (для мужчин)
[40-047] Планирование беременности — необходимые анализы
Кто назначает исследование?
Акушер-гинеколог, врач общей практики.
Литература
Alan H. DeCherney, Lauren Nathan, Neri Laufer. Current Diagnosis & Treatment Obstetrics & Gynecology, Eleventh Edition. McGraw-Hill Medical, 2007.
Abigail Delaney, Jani R. Jensen, Dean Morbeck. Fertility Testing. How Laboratory Tests Contribute to Successful Infertility Treatments. Clinical Laboratory News, November 01, 2012.
Топ-5 гормонов, из-за которых вы прибавляете в весе
На увеличение массы тела может влиять образ жизни и рацион питания, генетическая предрасположенность и возраст, роды и менопауза. По сути, основная причина нежелательного лишнего веса — гормональный дисбаланс. Мы рассмотрим пять наиболее важных гормонов, влияющих на массу тела, а также поделимся лайфхаками о том, как их контролировать.
Щитовидная железа: гормоны T3, T4 и кальцитонин
За обмен веществ в нашем организме отвечает щитовидная железа. Это маленький, но очень важный орган, поскольку недостаточный объем основных гормонов щитовидки провоцирует гипотиреоз. Один из симптомов этого синдрома — набор веса. Отметим, что объема телу добавляет не жир, а жидкость, которая накапливается в тканях организма.
Как избежать проблем?
Добавьте в рацион йодированную соль, продукты, богатые цинком и витамином D, избегайте блюд из сырых овощей и фруктов.
Поджелудочная железа: инсулин
Гормон инсулин вырабатывает поджелудочная железа. Он участвует в сложном процессе перенесения глюкозы в клетку и трансформации ее в энергию, либо «складирование» в виде жира. Употребление в большом количестве фастфуда, алкоголя, продуктов с искусственными подсластителями может спровоцировать резистенцию к инулину. Это опасно, так как вызывает скачки сахара в крови, которые в свою очередь дают толчок к развитию диабета 2-го типа и ожирению.
Как избежать проблем?
Следует убрать из рациона нездоровую пищу, спиртное, газировку и добавить овощи, зелень, сезонные фрукты. Возьмите за привычку заниматься спортом и пить вдоволь воды ежедневно (не менее 1,5 литров). Позаботьтесь о том, чтобы в вашем меню были источники омега-3-жирных кислот.
Кортизол
Кортизол, вырабатываемый в надпочечниках, именуют гормоном стресса. Он начинает активно «работать», если вы находитесь в опасности, переживаете беспокойство, стресс или страдаете от физического повреждения тела. Его главная задача — снизить уровень стресса через выброс сахара в кровь и регуляцию метаболизма. С учетом того, что современный человек находится в перманентном стрессе, кортизол вырабатывается регулярно и вредит здоровью. Он провоцирует отложение висцерального жира и созревание жировых клеток. Как избежать проблем? Важно спать не менее 8 часов в сутки, посвящать время йоге, медитации, дыхательным упражнениям, избегать спиртных напитков, жареной пищи. Тестостерон Этот гормон называют мужским, но он вырабатывается и в женском организме. Тестостерон отвечает за сжигание жира, крепость костей и эластичность мышц, за активность либидо. Соответственно, если этот гормон перестает вырабатываться в нужном объеме, наблюдается хрупкость костей, потеря мышечной массы и набор веса.
Как избежать проблем?
Позаботьтесь о том, чтобы в вашем рационе было вдоволь клетчатки, избегайте спиртных напитков, регулярно занимайтесь спортом и не забывайте о белковых добавках.
Прогестерон
Прогестерон — это важный женский половой гормон, который влияет на менструальный цикл и беременность. Его уровень в организме женщины должен быть сбалансированным, в противно случае может начаться депрессия и набор веса. На снижение уровня прогестерона влияет прием противозачаточных таблеток, стресс и менопауза.
Как избежать проблем?
Возьмите за правило заниматься спортом, медитируйте и старайтесь избегать стрессовых ситуаций.
Гормоны отвечают за многие физиологические процессы. Поэтому очень важно регулярно проверять уровень наиболее значимых гормонов. Без этого будет проблематично достичь хороших результатов при соблюдении диет, и даже интенсивные физические нагрузки не принесут желаемого эффекта. Рекомендуем сдать Гормональную панель в лаборатории клиники «Актив-Медикал». Вы получите объективную картину своего гормонального фона и сможете выявить причину набора веса. Детали по телефону (0512) 777-888.
Пять самых важных вопросов о мужском здоровье — Российская газета
Оказывается, уже начиная с 30 лет у мужчин постепенно снижается уровень тестостерона — одного из основных мужских гормонов, отвечающих не только за половую функцию, но и многие другие вещи. О том, как сохранить мужское здоровье подольше, и о том, можно ли восстановить угасающие способности, «РГ» рассказал врач-андролог, главный научный сотрудник Института репродуктивной медицины НМИЦ эндокринологии Минздрава РФ, профессор Института высшего и дополнительного профобразования НМИЦ эндокринологии Роман Роживанов.
Не всем известно, но вся наша жизнь — не только половые функции, но и общее самочувствие, и работа всех систем организма — напрямую зависит от гормонов. Их много. Они разные и отвечают за разные вещи. В женском организме обязательно присутствуют мужские гормоны, а мужской не может существовать без женских. Но крайне важно, чтобы все это многообразие существовало в балансе.
«Известен ряд мужских гормонов, связанных с половой функцией, — это ЛГ, ФСГ, ингибин, о которых из неспециалистов мало кто знает. Они необходимы для сперматогенеза. А всем известная классика — это тестостерон и дигидротестостерон, — говорит профессор Роживанов. — Именно они «отвечают» за половое влечение, сексуальное поведение, но не только. Рост мышц, крепость костей, общее состояние организма и даже особенности интеллекта и жизненная активность также зависят от уровня этих гормонов».
1. Что ухудшает выработку гормонов?
Проблемы со сном. На выработку гормонов влияют циркадные ритмы. Поэтому если сон нарушен, циркадные ритмы сбиваются, и при этом нарушается выработка гормонов.
Стресс — классический негативный фактор. «Эволюционно половая функция нужна для получения потомства. А для его выживания необходима благоприятная среда, — поясняет Роман Роживанов. — Но если мы живем в хроническом стрессе, значит, наш организм воспринимает окружающую среду как неблагоприятную. Ему просто не до детей, и он подавляет половую функцию».
Ожирение — тоже огромный минус. Жировая ткань, эндокринологи доказали это давно и убедительно, выступает как отдельный гормональный орган — в ней происходит патологический синтез стероидных гормонов, которые «вытесняют» производство необходимого тестостерона.
Прием стероидных препаратов. Известный факт: спортсмены-бодибилдеры, «выстраивающие» мощную мускулатуру с помощью стероидных добавок, зачастую не в состоянии выполнить элементарные мужские функции. «Анаболические стероиды, которые используют спортсмены, очень похожи на естественные стероидные гормоны, — поясняет Роман Роживанов. — Соответственно, наш гипофиз ошибочно принимает их как собственные половые гормоны. Но раз есть избыток — выработка собственного тестостерона тормозится».
Гормональный баланс для мужского организма важен так же, как и для женского
Интоксикация любого происхождения. «На гормональный фон отрицательно влияют любые вредные химические вещества, попадающие в организм», — говорит профессор.
Гиподинамия. Особенно вредно долгое неподвижное сидение — происходит застой крови в органах малого таза, и это плохо для половой функции.
Регулярный перегрев половых органов. «Яички не случайно находятся в мошонке снаружи, а не внутри организма, — говорит доктор. — Для их нормальной работы нужна температура на 2-3 градуса ниже температуры тела. Поэтому тесное белье, как и подогреваемое сиденье, может оказать негативный эффект».
Возраст. У всех абсолютно мужчин это не болезнь, а естественный физиологический процесс — после 30 лет уровень тестостерона начинает снижаться. Общий тестостерон — в среднем на 0,4-1,2 процента в год.
2. От чего же зависит возраст, в котором у мужчины начинаются проблемы?
«Во-первых, от максимального уровня тестостерона, который был по окончании периода полового созревания, то есть на пике. И во-вторых, от скорости падения его уровня, — поясняет Роман Роживанов. — И если на первую причину повлиять мы не можем, то со второй (а на нее как раз и влияют все вышеперечисленные факторы) можем успешно бороться».
Главный способ сохранить мужскую силу как можно дольше — убрать воздействие всех негативных факторов.
«Как ни банально, но в большинстве случаев (если нет необратимых изменений, например, травмы яичек) главный способ лечения — соблюдение здорового образа жизни. Среди мужчин среднего возраста без сопутствующих заболеваний около 6 процентов имеют дефицит тестостерона. Но если выделить группу страдающих ожирением или диабетом, проблема будет уже у каждого второго. Если сочетается сразу несколько негативных факторов — и стресс, и гиподинамия, и вредные привычки, дисфункция может возникнуть уже в 30 лет».
Половине пациентов с потенциально обратимым дефицитом тестостерона удается нормализовать его уровень и восстановить половую функцию только за счет похудения, увеличения физической активности и отказа от вредных привычек. «Почему мы говорим только о половине пациентов? Потому что подобрать программу мало, кого-то просто не удалось замотивировать на ЗОЖ, человек элементарно не смог похудеть», — поясняет Роман Роживанов.
3. Какие симптомы указывают на дефицит мужских гормонов?
Главные — это:
снижение либидо;
нарушение эрекции.
Дополнительно — вегетососудистые нарушения:
горячие приливы;
повышенная потливость;
сердцебиение, кардиалгия.
Психоэмоциональные:
быстрая утомляемость;
изменчивое настроение, склонность к депрессии;
снижение творческой продуктивности;
расстройства сна.
Эндокринные:
ожирение, увеличение грудных желез;
уменьшение оволосения на лице.
Костно-мышечные:
снижение или, наоборот, неадекватное физическим нагрузкам увеличение мышечной массы;
боли в костях.
Трофические нарушения:
сухость кожи, появление морщин.
Мочеполовые расстройства:
учащенное мочеиспускание;
бесплодие.
4. Возможно ли медикаментозное лечение?
«Да, фармакологическая помощь возможна, — говорит Роман Роживанов. — Но проведение андрогензаместительной терапии — это строго индивидуальная история. Все препараты рецептурные, и самолечение недопустимо. Есть противопоказания: повышенный уровень гемоглобина, некоторые опухоли. Абсолютное противопоказание — рак предстательной железы (а вот при аденоме простаты лечение возможно). Что касается возраста, о чем часто спрашивают пациенты, тут особых ограничений нет. Если качество жизни не устраивает и мужчина хочет его улучшить и противопоказаний нет, пожалуйста, начать терапию можно в любом возрасте и проводить ее пожизненно».
5. Поможет ли гормонозаместительная терапия сохранить мужскую силу?
Обычно это первый вопрос, который волнует мужчин больше всего остального. Доктор говорит: да, поможет, но это не главное. В первую очередь поддержание уровня тестостерона сохраняет мышечную силу, предотвращает вторичное ожирение, улучшает обмен веществ.
«По поводу восстановления сексуальной функции — любопытный факт. Когда начинаются проблемы, мужчины часто идут не к врачу, а в аптеку, и говорят: дайте мне что-нибудь поэффективнее для поддержания эрекции. Начинают принимать препараты, а результата нет. Наступает полное разочарование, — рассказал профессор. — Но мужчину никто не предупредил (а фармацевт в аптеке и не обязан знать такие тонкости), что стимулирующие препараты плохо работают при гормональной дисфункции. Если устранить гормональный дефицит — эффект будет гораздо лучше».
6 гормонов, которые мешают вам похудеть
Деятельность гормонов часто приводит к неконтролируемому чувству голода Фото: pixabay.com
Исследователи объяснили взаимосвязь между гормональной системой человека и похудением.
Многие уверены: чтобы похудеть, достаточно соблюдать баланс между правильным питанием и физической активностью. Но исследования показывают, что и это не самое главное.
Кроме диеты и образа жизни, на вес влияют гормоны: лептин, грелин, трийодтиронин (Т3), кортизол, эстроген и тестостерон. Одни отвечают за аппетит, другие за отложение жира, и от того, как они себя ведут, зависит индекс массы тела.
6 гормонов, которые влияют на ваш вес:
Лептин. Когда мозг получает достаточно лептина, то думает, что сыт, и запускается процесс сжигания жира. Ученые считают, что у многих людей эта связь нарушена, и организм ошибочно пытается сохранить жир. Чтобы запустить эту цепочку, ешьте больше белка и клетчатки, откажитесь от обработанных продуктов и сахара. Занимайтесь физической активностью и хорошо спите.
Грелин. Сообщает мозгу о чувстве голода. Считается, что этот гормон не способствует ожирению, но его неправильные сигналы могут побудить есть, даже когда не голодны, что приводит к увеличению веса. Лучший способ борьбы с активностью грелина – достаточный сон.
Трийодтиронин или Т3. Вырабатывается щитовидной железой и помогает контролировать использование организмом жира. Низкий уровень T3 может быть признаком гипотиреоза, который, помимо прочего, вызывает увеличение веса.
Кортизол. Гормон стресса кортизол повышает аппетит. В итоге вы едите больше, чтобы справиться со стрессом. Это помогает объяснить, почему люди с высоким уровнем хронического стресса чаще страдают из-за лишнего веса.
Эстроген. Женский гормон вырабатывается в жировых клетках, особенно в висцеральном жире вокруг живота, в том числе и у мужчин. У женщин преобладание эстрогена во время постменопаузы способствует увеличению веса.
Тестостерон. Мужской половой гормон есть и у женщин, но меньше. Высокий уровень тестостерона помогает похудеть и поддерживать вес в норме.
Ранее «Кубанские новости» рассказали, сколько надо гулять, чтобы похудеть.
Hormone Response Element — обзор
C КЛЕТОЧНО-СПЕЦИФИЧЕСКИЕ ЭЛЕМЕНТЫ (CSE) И ЭЛЕМЕНТ ОТВЕТА НА ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ (TRE), КАК СОСТАВЛЯЮТ УЛУЧШИТЕЛЬ-ПОДОБНОЕ УСТРОЙСТВО, КОТОРОЕ ОБЕСПЕЧИВАЕТ ПОВРЕЖДЕНИЕ СПЕЦИФИЧНОСТИ ГЕРОФОРМЫ КЛЕТОЧНОГО ТИПА И ТЕМПЕРАТУРЫ ЭКСПРЕССИЯ
Элемент ответа на гормон щитовидной железы, содержащийся во фрагменте -236/146, наиболее эффективно работает с промотором гомологичного гена гормона роста крысы, и это не зависит от спиральной связи между двумя областями (Flug et al., 1987). Стимуляция l-T3 не происходила, когда фрагмент -236 / -146 был лигирован (Flug et al. , 1987) с безхансерным промотором SV40 в pA 10 -cat 2 (Laimins et al. , 1984). ). В исследованиях, представленных здесь, стимуляция l-T3 также не была обнаружена, когда ДНК от -236 до -146 была помещена перед безхансерным промотором RSV [pRSVΔE-cat (2)]. Однако гормонально-регулируемая экспрессия имела место, когда фрагмент, простирающийся от -236 до -47, был лигирован с промотором RSV либо в нормальной [pRSVΔE-cat (3)], либо в инвертированной [pRSVΔE-cat (4)] ориентации, но не в 3 ‘ориентации ген CAT [pRSVΔE-cat (5)] (фиг.11 и 12). Мы также недавно обнаружили, что и гормон щитовидной железы, и клеточно-специфические элементы необходимы для передачи регулируемой l-T3 экспрессии на неинхансерном промоторе SV40 pA 10 -cat 2 (неопубликованное наблюдение). Эти результаты показывают, что как дистальный элемент ответа гормона щитовидной железы, так и клеточно-специфический базальный элемент необходимы для регулируемой l-T3 экспрессии гена гормона роста. Поскольку эти последовательности функционируют независимо от ориентации (рис.11 и 12), они, по-видимому, действуют как функциональная энхансерная единица, которая может обеспечивать как клеточно-специфическую, так и регулируемую тироидными гормонами экспрессию.
Чтобы учесть наши наблюдения, мы предлагаем механизм регуляции гена гормона роста крысы, в котором для стимуляции l-T3 требуются два элемента, в которых происходят белковые взаимодействия, специфичные для гормона-рецептора-клетки. В этой модели мы предполагаем, что рецептор связывается с последовательностями в области -208 / -178 (рис. 20B) и что комплекс l-T3-рецептор действует, «стабилизируя» или «усиливая» взаимодействия белок-ДНК клетки. -специфический базальный элемент.Это приводит к образованию более «активного» или «стабильного» транскрипционного комплекса, который стимулирует уровень экспрессии генов. Хотя мы не подтвердили, что функциональный элемент в области -208 / -178 связывает рецептор, 31 нуклеотидная последовательность содержит интересную область обратной симметрии диады (рис. 20B). Эта область также демонстрирует некоторую гомологию с 5′-фланкирующей ДНК других генов крыс, которые стимулируются l-T3 после введения in vivo (Flug et al. , 1987).
Недавние исследования промотора вируса опухоли молочной железы мыши (MMTV) подтверждают аналогичную двухэлементную модель для стимуляции экспрессии гена глюкокортикоидным гормоном. Инактивация сайта связывания фактора транскрипции NF-I (или белка с аналогичной последовательностью распознавания, например, TGGCA-связывающего белка) заметно снижает стимуляцию глюкокортикоидными гормонами без значительного изменения базальной активности гена (Buetti and Kuhnel, 1986 ; Miksicek и др. , 1987).Кроме того, отпечаток in vivo указывает на то, что инкубация глюкокортикоидного гормона увеличивает ассоциацию NF-I с его родственной последовательностью (Cordingley et al. , 1987), предполагая, что рецептор глюкокортикоидов усиливает связывание NF-I или взаимодействует с NF-I для образуют комплекс транскрипции, который активирует экспрессию промотора MMTV. Поскольку контрольные элементы других генов, чувствительных к тироидным гормонам, еще не идентифицированы, невозможно определить, необходимы ли два элемента для регулируемой экспрессии l-T3 в других системах.Однако он обеспечивает механизм, объясняющий, как гормон щитовидной железы, предположительно действующий через один и тот же рецептор, может положительно (например, гормон роста крысы) или отрицательно (например, гормон, стимулирующий щитовидную железу) (Kourides et al. , 1984; Shupnik и др. , 1985; Carr и др. , 1987) регулируют экспрессию генов. Происходит ли положительная или отрицательная регуляция, будет зависеть от комплекса гормон-рецептор, действующего в цис-системе, чтобы усилить или подавить эффект второго транс-действующего регуляторного белка (ов), который играет центральную роль в определении скорости экспрессии гена.
Для подтверждения этой гипотезы требуются дополнительные исследования. Например, положительная и отрицательная регуляция может опосредоваться структурно схожими, но разными рецепторами гормонов щитовидной железы. Наблюдение за тем, что геном человека содержит по крайней мере два гена, родственных erbA на разных хромосомах, повышает эту возможность (Dayton et al. , 1984; Spurr et al. , 1984; Raines et al. , 1985; Weinberger и др. , 1986). Значительный прогресс был достигнут в области действия гормонов щитовидной железы с момента первоначальной идентификации ядерных рецепторов гормонов щитовидной железы в печени и почках (Oppenheimer et al ., 1972) и в культивируемых клетках (Samuels, Tsai, 1973). Будущие достижения в этой области потребуют клонирования мРНК рецепторов тироидных гормонов из различных тканей и клеток для определения их структуры и функции. Кроме того, выделяются и секвенируются другие гены ответа на гормоны щитовидной железы. Идентификация элементов ответа на гормоны щитовидной железы других генов и регуляторных белков, которые опосредуют их экспрессию, должна обеспечить всестороннее представление о детальных механизмах, участвующих в положительной и отрицательной регуляции генов, реагирующих на гормоны щитовидной железы, в различных клетках и тканях.
Возрастные изменения выработки гормонов: MedlinePlus Medical Encyclopedia
Эндокринная система состоит из органов и тканей, вырабатывающих гормоны. Гормоны — это природные химические вещества, которые производятся в одном месте, попадают в кровоток, а затем используются другими органами и системами-мишенями.
Гормоны контролируют органы-мишени. Некоторые системы органов имеют свои собственные системы внутреннего контроля наряду с гормонами или вместо них.
С возрастом естественным образом происходят изменения в способах управления системами организма.Некоторые ткани-мишени становятся менее чувствительными к их контролирующему гормону. Также может измениться количество вырабатываемых гормонов.
Уровень некоторых гормонов в крови повышается, некоторых понижается, а некоторых не изменяется. Гормоны также расщепляются (метаболизируются) медленнее.
Многие органы, вырабатывающие гормоны, контролируются другими гормонами. Старение также меняет этот процесс. Например, эндокринная ткань может вырабатывать меньше гормона, чем в более молодом возрасте, или же она может вырабатывать такое же количество с меньшей скоростью.
ИЗМЕНЕНИЯ СТАРЕНИЯ
Гипоталамус расположен в головном мозге. Он производит гормоны, которые контролируют другие структуры эндокринной системы, включая гипофиз. Количество этих регулирующих гормонов остается примерно таким же, но реакция эндокринных органов может меняться с возрастом.
Гипофиз расположен чуть ниже (передний гипофиз) или в (задний гипофиз) головного мозга. Эта железа достигает максимального размера в среднем возрасте, а затем постепенно становится меньше.Он состоит из двух частей:
В задней (задней) части хранятся гормоны, вырабатываемые гипоталамусом.
Передняя (передняя) часть вырабатывает гормоны, влияющие на рост, щитовидную железу (ТТГ), кору надпочечников, яичники, яички и грудь.
Щитовидная железа расположена в области шеи. Он производит гормоны, которые помогают контролировать обмен веществ. С возрастом щитовидная железа может стать бугристой (узловатой). Со временем метаболизм замедляется, начиная примерно с 20 лет. Поскольку гормоны щитовидной железы производятся и расщепляются (метаболизируются) с одинаковой скоростью, тесты функции щитовидной железы чаще всего остаются нормальными.У некоторых людей может повышаться уровень гормонов щитовидной железы, что приводит к повышенному риску смерти от сердечно-сосудистых заболеваний.
Паращитовидные железы — это четыре крошечные железы, расположенные вокруг щитовидной железы. Гормон паращитовидной железы влияет на уровень кальция и фосфата, которые влияют на прочность костей. Уровень паратироидного гормона повышается с возрастом, что может способствовать развитию остеопороза.
Инсулин вырабатывается поджелудочной железой. Он помогает сахару (глюкозе) попасть из крови внутрь клеток, где он может быть использован для получения энергии.
Средний уровень глюкозы натощак повышается на 6–14 миллиграммов на децилитр (мг / дл) каждые 10 лет после 50 лет, поскольку клетки становятся менее чувствительными к действию инсулина. Когда уровень достигает 126 мг / дл или выше, считается, что человек страдает диабетом.
Надпочечники расположены чуть выше почек. Кора надпочечников, поверхностный слой, вырабатывает гормоны альдостерон, кортизол и дегидроэпиандростерон.
Альдостерон регулирует баланс жидкости и электролитов.
Кортизол — гормон «стрессовой реакции». Он влияет на расщепление глюкозы, белков и жиров, а также обладает противовоспалительным и противоаллергическим действием.
Высвобождение альдостерона уменьшается с возрастом. Это снижение может способствовать головокружению и падению артериального давления при внезапной смене положения тела (ортостатическая гипотензия). Высвобождение кортизола также снижается с возрастом, но уровень этого гормона в крови остается примерно таким же. Также снижается уровень дегидроэпиандростерона.Воздействие этой капли на организм неясно.
Яичники и семенники выполняют две функции. Они производят репродуктивные клетки (яйцеклетки и сперму). Они также производят половые гормоны, которые контролируют вторичные половые признаки, такие как грудь и волосы на лице.
С возрастом у мужчин часто снижается уровень тестостерона.
У женщин после менопаузы снижается уровень эстрадиола и других гормонов эстрогена.
ВЛИЯНИЕ ИЗМЕНЕНИЙ
В целом, некоторые гормоны снижаются, некоторые не меняются, а некоторые увеличиваются с возрастом.Гормоны, которые обычно снижаются, включают:
У женщин уровни эстрогена и пролактина часто значительно снижаются.
Гормоны, которые чаще всего остаются неизменными или лишь незначительно снижаются, включают:
Кортизол
Адреналин
Инсулин
Гормоны щитовидной железы Т3 и Т4
Уровни тестостерона обычно постепенно снижаются с возрастом у мужчин.
Гормоны, которые могут увеличиваться, включают:
СВЯЗАННЫЕ ТЕМЫ
новых регуляторов витамина D и передачи сигналов эстрогена
Стероиды.Авторская рукопись; доступно в PMC 2012 1 марта.
Опубликован в окончательной редакции как:
PMCID: PMC3042887
NIHMSID: NIHMS267406
Отделение ортопедической хирургии и Института молекулярной биологии, Школа медицины Дэвида Геффена в Калифорнийском университете в Лос-Анджелесе, 615 Charles E Янг Драйв Саут, Лос-Анджелес, Калифорния
, США. Автор, ответственный за переписку: Томас С. Лисс, доктор философии, отделение ортопедической хирургии, Медицинская школа Дэвида Геффена, UCLA, 615 Charles E. Young Drive, Южный Лос-Анджелес, Калифорния, США Тел .: 310 206 9356 Факс: 310 825 5409 ude.alcu.tendem@essilt Окончательная отредактированная версия этой статьи издателем доступна на сайте Steroids. См. другие статьи в PMC, в которых цитируется опубликованная статья.
Abstract
Исследования приматов Нового Света, устойчивых к витамину D, и их человеческих гомологов в качестве моделей естественной и патологической нечувствительности к действию стеролов / стероидов открыли семейство новых внутриклеточных белков, регулирующих витамин D и эстроген, участвующих в действии гормонов. Белки, известные как «белки, связывающие элементы ответа на витамин D или эстроген», действуют как мощные трансдоминантные регуляторы, действующие на цис , для ингибирования связывания стероидных рецепторов с элементами ответа ДНК и отвечают за устойчивость к витамину D и эстрогену.Этот набор интеракторов принадлежит к семейству гетерогенных ядерных рибонуклеопротеидов (hnRNP) ранее известных белков, взаимодействующих с пре-мРНК. Этот обзор дает новое понимание механизма, с помощью которого эти новые регуляторы передачи сигналов и метаболизма могут действовать, регулируя ответы на витамин D и эстроген. Кроме того, в обзоре также описаны другие молекулы, которые, как известно, влияют на передачу сигналов ядерного рецептора через взаимодействие с элементами гормонального ответа.
I. Введение
Ответы клеток-мишеней на стероидные / стероловые гормоны зависят от множества факторов, включая экспрессию и регуляцию родственных ядерных рецепторов (1-3), связанных с ними дополнительных белков (4) и пререцепторную регуляцию собственно гормональные лиганды (5-6).В предыдущих исследованиях приматов Нового Света (NWP) и приматов Старого Света (OWP) мы идентифицировали новый класс белков, участвующих в регуляции передачи сигналов витамина D и эстрогена на уровне транскрипции генов (7). ЧПП обычно демонстрируют резистентность органа-мишени к стероидным гормонам, таким как эстрогены, отчасти из-за пищевой и экологической экологии ЧПП (т.е. пищевого воздействия высоких уровней растительных фитоэстрогенов и повышенных уровней витамина D в результате обычного воздействия УФ-В. свет).Предыдущая работа нашей группы показала, что резистентность к стероидным / стероловым гормонам в NWPs включает сверхэкспрессию белков, связывающих элемент ответа (REBiP), которые конкурируют с ядерными рецепторами за связывание с цис- регуляторными элементами в целевой ДНК (8-9). В OWP белок, связывающий элемент ответа на витамин D (VDRE-BP), занимает элементы ответа на витамин D гена-мишени (VDRE) в отсутствие значительных уровней гормонального 1,25-дигидроксивитамина D 3 [1,25 (OH) 2 D], биологически активная форма гормона.Однако, когда уровни 1,25 (OH) 2 D увеличиваются, лигандированный рецептор витамина D (VDR) затем может вытеснять VDRE-BP и индуцировать транскрипцию целевого гена. В NWP избыточное количество VDRE-BP означает, что гораздо более высокие уровни 1,25 (OH) 2 D необходимы для индукции передачи сигналов VDR (10). У людей нормальный ответ на 1,25 (OH) 2 D также включает конкуренцию между VDR и VDRE-BP (11), а избыточная экспрессия VDRE-BP была описана у пациента с наследственным рахитом, устойчивым к витамину D (HVDRR). ) (11-12).Эти исследования также показали, что человеческий VDRE-BP идентичен гетерогенному ядерному рибонуклеопротеину (hnRNP) C1 / C2 (11), члену семейства hnRNP белков, взаимодействующих с РНК.
Наконец, NWPs также включают новые регуляторные особенности одного из рецепторов эстрогена (ERα), участвующие в резистентности к гормонам, которая включает оба белка, связывающего элемент ответа на эстроген (ERE-BP) (13-16). В этом обзоре кратко рассматриваются молекулярные механизмы, участвующие в резистентности к гормонам, и обсуждается возможное значение различных режимов для физиологического действия рецепторов витамина D и гормона эстрогена.
II. Суперсемейство ядерных рецепторов рецепторов стероидных гормонов и активация генов
Ядерные рецепторы (NR) составляют класс факторов транскрипции и сигнальных молекул у позвоночных. Члены NR включают рецепторы гидрофобных молекул, таких как стероидные / стероловые гормоны (например, эстрогены, глюкокортикоиды, прогестерон, минералокортикоиды, андрогены, витамин D 3 , экдизон, оксистерины и желчные кислоты), ретиноевые кислоты (все — транс и 9. — цис изоформ), гормоны щитовидной железы, жирные кислоты, лейкотриены и простагландины (17-18).NR опосредуют транскрипцию генов-мишеней, связываясь с различными элементами ответа и формируя комплексы, состоящие из ко-регуляторных белков, чтобы влиять на ремоделирование хроматина и эпигенетические модификации, часто в местах, удаленных от сайта начала транскрипции. NR для всех классов стероидных гормонов могут контролировать транскрипцию генов либо путем активации факторов транскрипции, либо в целом действуя как факторы транскрипции. Классически NR функционируют в три ключевых этапа: репрессия, дерепрессия и активация транскрипции (18-19).Репрессия включает привлечение корепрессорного комплекса с гистондеацетилазной активностью (HDAC). То есть эукариотическая транскрипция нарушается репрессивным хроматиновым окружением регуляторных областей генов. Дерепрессия происходит после связывания лиганда, которое диссоциирует этот «репрессированный» комплекс и привлекает первый комплекс соактиватора с активностью гистонацетилтрансферазы (HAT), что приводит к деконденсации хроматина, которая считается необходимой, но недостаточной для активации целевого гена.На третьем этапе комплекс HAT диссоциирует и образуется второй комплекс соактиватора (например, TRAP / DRIP / GRIP / ARC), который способен устанавливать ассоциации с базальным аппаратом транскрипции и, таким образом, приводит к активации транскрипции мишени. ген. Следует также отметить, что этот механизм не является общим, так как некоторые NR могут действовать как активаторы независимо от лиганда, тогда как другие не могут взаимодействовать с промотором целевого гена в отсутствие лиганда, не говоря уже о разнообразии сопутствующих лигандов. регуляторные комплексы.
Стероидные / стероловые рецепторы представляют собой эволюционно консервативные лиганд-зависимые и -независимые факторы транскрипции, которые принадлежат к разнообразному суперсемейству белков NR (17-18). Эти рецепторы можно найти на плазматической мембране, в цитозоле, а также в ядре клеток-мишеней. Липофильные гормоны, пересекающие клеточную мембрану, для стероидных рецепторов основаны на различных химических структурах стероидного ядра, хотя некоторые рецепторы также способны связывать пищевые липиды.Рецепторы, которые связываются со стероловыми лигандами у позвоночных, являются основным предметом этого обзора и включают: 1] PXR, который связывает прегненолон; 2] LXR, распознающий 22-гидроксихолестерин; 3] FXR, который связывает желчные кислоты, такие как хенодезоксихолевая кислота; 4] CAR, который связывается с метаболитами андрогенов, такими как андростенол; и 5] VDR, рецептор почечного гормона 1,25 (OH) 2 D, который стимулирует абсорбцию кальция в кишечнике и ремоделирование костей ().
Таблица 1
Суперсемейство рецепторов витамина D и стероидных эстрогенов
Подсемейство
Группа
Имя
Лиганд
Функция
OMIM 1
Resistances
Рецептор тироидного гормона
Рецептор витамина D
Рецептор витамина D (VDR) ( VDR )
-Витамин D (1,25 (OH) 2 D) -Литохолин кислота
— Минеральный транспорт и метаболизм — Иммунный ответ — Прогрессирование рака — Децидуализация — Кишечное всасывание — Развитие скелета — Развитие волос
601769
— Наследственный витамин D-зависимый -паразит 2-го типа — Остеопороз — Недостаток витамина D — Рак предстательной железы / груди — Алопеция — Острый лимфоцит ic Лейкоз
(53, 86-95)
Рецептор эстрогена
Рецептор эстрогена
Рецептор эстрогена-α, β (ERα, β) ( ESR1,2 )
— Эстрогены
— Эстральный цикл -Сексуальное развитие -Уменьшение мышечной массы -Увеличение жировых отложений -Уменьшение резорбции костной ткани, увеличение костеобразования -Коагуляция -Липидный метаболизм -Рак -Функция легких
NR имеют общую каноническую структурную организацию.Структурные домены, отвечающие за связывание лиганда, связывание ДНК и ко-регулятора, димеризацию рецептора и трансактивацию, являются высококонсервативными во всем семействе (20). N-концевая область (A / B-домен) очень вариабельна и включает по крайней мере одну конститутивно активную область трансактивации (AF-1) и несколько автономных доменов трансактивации (AD) (21). Наиболее консервативной областью является ДНК-связывающий домен (DBD, C-домен), который, в частности, содержит два домена «цинковых пальцев» и P-бокс, короткий мотив, ответственный за специфичность связывания ДНК на целевых последовательностях, обычно содержащих шесть пар оснований. Мотив, подобный AGGTCA, участвует в гомо- или гетеродимеризации ядерных рецепторов.Шарнирная область (домен D) считается гибким доменом, который соединяет DBD с умеренно консервативным в последовательности связывающим лигандом доменом (домен E) и влияет на внутриклеточный трафик и субклеточное распределение.
Способность связанных с лигандом стериновых рецепторов, таких как VDR, трансактивировать стерин-чувствительный ген зависит от присутствия AF1- и AF2-взаимодействующих «мостиковых» нуклеопротеидов, а также ко-регуляторов, которые имеют ремоделирующие хроматин и другие ферментативные мероприятия (22) ().Коактиватор может усиливать инициацию транскрипции за счет стабилизации сборки холофермента РНК-полимеразы, обеспечивая более быстрый клиренс промотора. Коактиваторы могут контролировать многие другие подэтапы транскрипции, включая удлинение, сплайсинг РНК, а также терминацию и деградацию комплекса соактиватора. Известные коактиваторы NR принадлежат к нескольким семействам. Коактиваторы ядерных рецепторов 1-3 ( NCOA1-3 ) представляют собой ко-регуляторные белки транскрипции, которые несут в себе несколько доменов, взаимодействующих с ядерными рецепторами, и внутреннюю гистонацетилтрансферазную активность.NCOA привлекаются к участкам промотора ДНК с помощью ядерных рецепторов, активируемых лигандом, и, в свою очередь, ацетилируют гистоны. Семейство p160 и активатор стероидного рецептора коактиватора рибопротеина (SRA) включают членов, активность которых ограничена ядерными рецепторами, а факторы транскрипции, как известно, взаимодействуют с членом семейства факторов транскрипции факторов транскрипции (TEF) (23) и ковалентно модифицируют гистоновые хвосты (24). Семейство коактиваторов CREB-связывающего белка (CBP) / p300 и связанного с CBP / p300 PCAF также имеют решающее значение для других систем передачи сигнала, включая протеинкиназу A-cAMP-CREB, фактор роста-cfos / cjun. фактор роста / цитокин Jak-STAT и пути цитокин-NFκB.Из-за их широких функций CBP и p300 также называются «коинтеграторами». Наконец, необходимы другие посттрансляционные модификации, такие как фосфорилирование серина, чтобы сделать стерол / стероидные рецепторы транскрипционно активными.
Стероидный рецептор индукции транскрипции гена: модель многоступенчатого контроля активности VDR на геномном и посттрансляционном уровнях
1α, 25-дигидроксивитамин D 3 (витамин D 3 ) — гормон, который играет роль решающая роль в регуляции / метаболизме кальция и фосфора в тонком кишечнике, почках и костях.Кроме того, он также участвует в иммунной функции, подавлении опухолей, регуляции роста и секреции паратироидного гормона. Выполняя эндокринную роль, свободные (т.е. несвязанные) стероиды могут проникать в цитоплазму клетки и взаимодействовать со своим рецептором. Для некоторых типов клеток витамин D 3 может самосинтезироваться в цитоплазме. В этом процессе белки теплового шока диссоциируют, и активированный комплекс рецептор-лиганд перемещается в ядро. VDR, рецептор витамина D 3 , существует в виде гетеродимера с RXR.Витамин D 3 VDR-RXR, связанный с , вместе с другими белками-коактиваторами и ко-репрессорами, опосредует регуляцию транскрипции ряда генов в ядре. Важно отметить, что VDRE-BP также опосредуют транскрипцию VDR-чувствительных генов, участвующих в регуляции гормонов, костей и роста. Следовательно, любое нарушение работы витамина или его рецептора будет иметь последствия для ряда ключевых физиологических процессов. Более того, на пострецепторном уровне дефектный коактиватор или чрезмерная ко-репрессорная активность могут привести к резистентности к ядерным гормонам, влияющей на множественные ядерные гормональные рецепторные системы.Данные были проанализированы и подтверждены (например, цитаты) с помощью анализа путей изобретательности (Ingenuity® Systems, www.ingenuity.com)
III. Устойчивость к гормонам
Иногда гормоны не могут проявлять свои специфические реакции в определенных тканях из-за устойчивости к определенному гормону. В большинстве случаев резистентность к гормонам вызывается мутациями в рецепторах гормонов, влияющих на критические домены, участвующие в связывании гормонов, димеризации и трансактивации рецепторов, или функциональными изменениями взаимодействующих рецепторов гормонов, таких как сывороточные связывающие белки, для нарушения различных сигнальных путей.Синдромы гормональной резистентности были исследованы в соответствии с типами рецепторов, чтобы проиллюстрировать разнообразие гормональной резистентности или нарушений. Эти типы рецепторов включают семь трансмембранных рецепторов, связанных с G-белком (24-26), рецепторы тирозинкиназ (27), семейство рецепторов цитокинов (28) и группу рецепторов серинкиназы трансформирующего фактора роста (TGFβ) (29). Кроме того, мутации G-белка (например, GNAS1) вызывают наследственную остеодистрофию Олбрайта (30), которая приводит к внутримембранозным оссификациям, постепенно развивающимся из дермы.
Как и следовало ожидать, различные синдромы резистентности затрагивают членов суперсемейства NR, включая резистентность к андрогенам, витамину D 3 , гормону щитовидной железы, глюкокортикоидам и эстрогену (). Многие NR имеют несколько изоформ. Например, и ER, и Х-рецептор печени (LXR) кодируются генами α и β (). Следовательно, мутации, которые устраняют одну изоформу рецептора, могут не полностью устранить действие гормона или даже могут вызвать усиление функции немутантной изоформы.Кроме того, мутации в орфанных NR, таких как DAX-1 ( NR0B1 ), приводят к нарушению развития желез, тем самым косвенно вызывая резистентность к гормонам, что приводит как к Х-сцепленной врожденной гипоплазии надпочечников, так и к гипогонадотропному гипогонадизму (31). В DAX-1 отсутствует нормальный ДНК-связывающий домен, содержащийся в других NR, и он действует как доминантно-негативный регулятор транскрипции других NR, включая стероидогенный фактор 1. Этот белок также функционирует как ген против семенников, действуя антагонистически по отношению к фактору транскрипции SRY. (Определяющая пол область Y) (32).
В других случаях резистентности к гормонам вариации обработки могут приводить к различиям в активности гормона на уровне «пре-рецепторов». Например, эндогенное антитело к гормону может инактивировать гормон и создавать резистентность пререцепторной природы. Кроме того, аутоантитела, генерируемые против рецептора полипептидного гормона, могут действовать как агонист или как антагонист, определяя действие полипептидного лиганда, что приводит, например, к болезни Грейвса, анемии и миастении (33).Другая форма резистентности, иногда называемая «пострецепторной» резистентностью, типична для приобретенных форм резистентности к инсулину или лептину, и ее патофизиология остается недостаточно изученной (34–35). Пострецепторный дефект вызывает пропорциональное снижение действия гормона на всех уровнях гормона, включая максимально эффективные концентрации, и это называется снижением гормональной чувствительности при нормальном связывании с рецептором инсулина. Это свидетельствует о нарушении связи связывания NR с последующими этапами действия гормона.Наконец, артефактная гормональная резистентность может быть связана с несоблюдением пациентом режима приема лекарств, их рецептуры, абсорбции и метаболизма, когда усиление гормональной терапии не дает видимых эффектов.
IV. Взаимодействие элементов гормонального ответа (HRE)
ДНК содержит конформационные и топологические сигнатуры, которые реализованы в хроматине и которые контролируют доступность для цис--действующих элементов (25-26). NR-индуцированные нуклеосомные изменения влияют на опосредованное рецептором рекрутирование факторов ремоделирования хроматина как часть сигнальных реакций стероидных гормонов.Вообще говоря, эти взаимодействия могут физиологически модулироваться факторами, зависящими от контекста клетки и промотора, для облегчения специфической и оптимальной реакции экспрессии генов на гормональную стимуляцию.
Уникальный способ регуляции, который мы постулировали в 1980-х годах на основе наблюдений, сделанных на NWP в отношении передачи сигналов стероидных гормонов (27-28), включает функциональный антагонизм между действующими рецепторами гормонов и другими не нацеливающими рецепторами гормонов или нерецепторные цис--действующие взаимодействующие с транскрипцией в обозначенных элементах гормонального ответа (HRE).Этот способ регуляции был расширен и охватывает не только VDRE и ERE, но также RORαRE, LXRE, (приманки) GRE, ARE, стероидогенные консенсусные полусайты и неклассические составные HRE (). Кроме того, в дополнение к открытиям нашей группы, специфический регуляторный контроль VDRE и ERE с помощью белков, связывающих «нерецепторные» элементы ответа, был подтвержден участием других не связанных компонентов, таких как повсеместный регулятор транскрипции YY1 (т. Е. Ying Yang 1, GLI-Kruppel класса белков цинковых пальцев) (29) и фактор прикрепления каркаса B (30), соответственно.Эти данные предоставляют дополнительные доказательства сложной регуляции стероидных гормонов и их рецепторов, нацеленных на VDRE и ERE в различных клеточных системах.
Таблица 2
Взаимодействующие элементы гормонального ответа
Связывающий ядерный рецептор
Связывающий белок / соединение / нкРНК
Элемент ответа
Регулируемый ген (ы)
Эффекты фактора связывания 9018 (s)
RORα, β, γ
Rev-ErbA beta (NR1D2)
RORαRE
HRE-репортерный ген
конкурентный репрессор функции RORα
(36)
(36)
β
рецептор тиреоидного гормона β1
LXRE
белок, связывающий регуляторный элемент стерола 1c
подавление трансактивации LXRα
(37)
902 AGTC
FXR 902ogenic ароматаза
базальный фактор, который конкурирует с FXR, который отрицательно действует на промотор
В остеобластах функциональный ген остеокальцина (ОС), содержащий VDRE, является транскрипционно активным в зрелом пост-рецепторе клетки в начале минерализации матрикса с образованием кости.Guo et al. (29) стремились понять витамин D-зависимую модуляцию транскрипции гена ОС с точки зрения лучшего определения взаимосвязей между VDR и фактором транскрипции YY1, связанным с нестероидными гормонами. YY1 участвует либо в репрессии, либо в активации разнообразного числа промоторов посредством рекрутирования гистондеацетилаз или ацетилтрансфераз, таким образом участвуя в модификации гистонов в VDRE или около них, чтобы способствовать ремоделированию хроматина (31). Авторы обнаружили, что многофункциональный регулятор YY1 подавляет нормальную 1,25 (OH) 2 D-индуцированную трансактивацию гена ОС, конкурируя за гетеродимеры VDR-RXR за связывание VDRE (т.е.е. очень похож на белки hnRNPC; см. ниже), а также препятствовали межбелковому взаимодействию между ДНК-связанным VDR-RXR и фактором инициации транскрипции TFIIB. Кроме того, авторы показали, что YY1 присутствует на конститутивных уровнях в пролиферирующих остеобластах, когда уровни гетеродимера VDR-RXR были низкими, возможный механизм, с помощью которого YY1 связывается с OC-VDRE, чтобы предотвратить взаимодействие гормонального рецептора с ДНК для блокирования преждевременной витамин D-зависимой трансактивации. зарезервировано для зрелых клеток.Предыдущие сообщения предполагают, что белки hnRNPC и YY1 являются ключевыми функциональными модуляторами чувствительности к витамину D в генах OC и CYP24A1 . Наши недавние неопубликованные данные свидетельствуют о том, что белки hnRNPC могут модулировать действия VDR нескольких других генов, связанных с обменом костных клеток и гомеостазом (). И YY1, и hnRNPC, по-видимому, являются компонентами регуляторных переключателей, которые контролируют костно-специфические гены. В обоих случаях выяснение молекулярных механизмов, опосредующих регуляцию стероидными гормонами развития скелета, поможет прояснить основы регуляторных нарушений, связанных с патологиями скелета, такими как наследственный рахит, устойчивый к витамину D (HVDRR).
Хотя есть и другие примеры конкуренции между NR и нерецепторными факторами транскрипции за связывание ДНК, пока нет сообщений о прямых взаимодействиях HRE, которые нарушают передачу сигналов нативного гормона для стероидных ядерных рецепторов PXR, CAR, ERR, MR и PR. . Имеются сообщения, описывающие класс составных элементов гормонального ответа (CHRE), в которых NR функционирует среди других регуляторов и усиливает или подавляет транскрипцию (32). То, что эта регуляция CHRE требует комбинированных действий рецептора-мишени вместе с одним или несколькими нерецепторными (перекрывающимися) регуляторами, специфичными для последовательности, обеспечивает другой уровень сложности (33).Такие локусы могут обозначать гормонально-зависимое усиление или подавление транскрипции и в отсутствие соответствующего нерецепторного фактора вообще не могут обеспечить гормональную регуляцию, несмотря на присутствие функционального рецептора. Это отличается от простых, более распространенных консенсусных HRE, в которых ядерный рецептор функционирует независимо от других регуляторов для усиления транскрипции (34), предполагая, что домены активации рецептора расположены близко к промоторам. В случае составного элемента plfG (т.е. GRE-AP-1), участие частичной последовательности AP-1 и субъединичного состава AP-1 (то есть гомодимеров c-Jun по сравнению с гетеродимерами c-Jun / c-Fos) определяет природу гормонального ответа, опосредованного GR (35). Вместе эти находки подразумевают, что позиционирование последовательности и тройные комплексы рецептора и AP-1 в plfG определяют направление и величину последующего гормонального ответа. Таким образом, последовательность plfG представляет собой четко очерченный составной элемент, который опосредует функциональные взаимодействия регуляторов, специфичных для последовательности, из двух хорошо охарактеризованных семейств факторов.
Другой класс белков, которые могут действовать как конкуренты за взаимодействие NR-ДНК, — это известные нецелевые (орфанные) ядерные рецепторы, которые обладают внутренней способностью регулировать экспрессию определенных генных сетей посредством конкуренции между эндогенными нативными рецепторами за один и тот же сайт узнавания. (36). Например, Rev-ErbA beta (NR1D2) распознает HRE со специфичностью, аналогичной RORα, и не активирует только транскрипцию, а скорее действует как конкурентный репрессор функции RORα (36).На аналогичной ноте существует перекрестная связь между элементами ответа T3 и LXR, которые имеют сходные конфигурации, для соответствующих рецепторов тироидных гормонов (TR) и LXRα, влияющих на пути передачи сигнала (37). В частности, основная изоформа TR в печени, TR-β1, связывается непосредственно с LXRE, расположенными в промоторе белка 1c, связывающего регуляторный элемент стерола, и взаимодействует с корепрессорами, важными для нелигандованного TR-опосредованного подавления LXRα-трансактивации. регулируют липидный обмен.
Недавно фарнезоидный рецептор X (FXR), важный для гомеостаза желчных кислот, был идентифицирован как отрицательный модулятор фермента андроген-эстроген-превращающей ароматазы в клетках рака груди человека (38). Механически в другом отчете исследования мутагенеза, сдвига электрофоретической подвижности и анализ иммунопреципитации хроматина (ChIP) с использованием опухолевых клеток Лейдига показывают, что FXR способен конкурировать со стероидогенным фактором 1 (SF-1), ключевым регулятором репродукции, в связывании с общая последовательность, присутствующая в промоторной области ароматазы, отрицательно влияющая на ее активность (39).Аналогичным образом белок теплового шока 27 (hsp27) усиливает как рост, так и лекарственную устойчивость клеток рака молочной железы и является плохим прогностическим фактором для некоторых подгрупп пациентов с раком молочной железы. Промотор hsp27 содержит элемент несовершенного ответа на эстроген (ERE), который разделен спейсером из 13 пар оснований, который содержит TATA-бокс, с которым связывается фактор прикрепления каркаса B (SAF-B) (30). Факторы прикрепления каркаса представляют собой подмножество белков ядерного матрикса (NMP), которые взаимодействуют с областями прикрепления матрикса.Избыточная экспрессия SAF-B приводит к дозозависимому снижению активности промотора hsp27, что позволяет предположить, что этот неядерный рецепторный белок является медиатором развития рака молочной железы на уровне связывания элемента гормонального ответа.
Некодирующие РНК (нкРНК) представляют собой функциональные молекулы РНК, которые не транслируются в белок. Специфический для остановки роста 5 (GAS5) широко распространен в клетках, рост которых был остановлен из-за нехватки сыворотки или питательных веществ. Интересно, что нкРНК GAS5 обладает способностью связываться с ДНК-связывающим доменом глюкокортикоидных рецепторов (GR), действуя как элемент ответа на GR (GRE), таким образом конкурируя с промоторными ДНК GRE за связывание с GR, чтобы опосредовать несколько нижестоящих ответных гены, такие как ингибитор апоптоза 2 (40).Известно, что глюкокортикоиды влияют на транскрипцию генов и обладают разнообразной активностью в отношении иммунного ответа, роста клеток, расхода энергии и выживания, в результате чего присутствие нкРНК GAS5-приманки может модулировать считывание транскрипции доступного GR.
Наконец, еще один интерактор, который может нарушить взаимодействие NR с HRE, включает низкомолекулярные полиамиды, предназначенные для конкуренции с NR за связывание с HRE (41-42). Полиамиды избирательно влияют на ДНК-зависимый путь, вмешиваясь в интерфейс белок-ДНК.Эти олигомеры могут встречаться как в естественных, так и в искусственных условиях, обеспечивая специфичность последовательности за счет параллельного спаривания гетероциклических аминокислот с малой бороздкой ДНК. Например, ДНК-связывающий полиамид нацелился на консенсусный GRE гена глюкокортикоид-индуцированной застежки-молнии (GILZ), чтобы заблокировать его экспрессию, а также на другие известные гены-мишени GR, оцениваемые с помощью анализа экспрессии в масштабе всего генома (41). Кроме того, недавно было показано, что тот же класс полиамидных соединений регулирует рак простаты путем связывания и подавления согласованного элемента андрогенного ответа (ARE) и гена простатоспецифического антигена в физиологическом контексте для ингибирования активности AR (42).Вкратце, хорошо известно, что многие пестициды способны ингибировать или активировать системы NR, чтобы влиять на уровни половых гормонов, чтобы изменить развитие или экспрессию мужской и женской репродуктивной системы (43), и, основываясь на недавних открытиях, это также может быть применимы к легко используемым промышленным полиамидным смолам, упомянутым выше.
Как описано выше, в литературе есть несколько указаний на взаимодействие между соответствующими HRE и сигнальными молекулами. Предмет взаимодействия HRE не является обобщенным: некоторые рецепторы связаны с HRE даже в отсутствие гормона, иногда функционируя для подавления экспрессии генов (44).Захватывающее наблюдение, что NRs семя формирует комплексы на HRE и включает ко-регуляторные белки и РНК, одновременно сильно возбудило и еще больше усложнило наше понимание действия гормонов (45). Применение мощных инструментов для определения местоположения сайтов связывания NR по всему геному с помощью таких методов, как ChIP с последующим глубоким секвенированием (ChIP-seq), показало, что рецепторы связываются с хроматином по всему геному, часто далеко от предполагаемых сайтов начала транскрипции, и могут быть обнаружены в межгенных областях, интронах и дистальных энхансерах (46).Эти результаты открывают новые возможности для потенциальных будущих исследований и понимания сложных механизмов и конкретных целевых регионов, с помощью которых взаимодействующие HRE влияют на реакцию стероидных гормонов.
V. Корегуляторы приматов Нового Света и не только. / стерол », резистентный к этим гормонам (47-48).В начале-середине 1980-х годов мы исследовали вспышку рахитической болезни костей в колонии NWP emperor tamarin (
Saguinas imperator ) в зоопарке Лос-Анджелеса (49-50). Анализ эпидермальных клеток, выделенных у этих животных, показал, что НПВ были защищены от потенциальных неблагоприятных эффектов длительного воздействия высоких уровней циркулирующих гормонов, таких как 1,25 (ОН) 2 D и 17β-эстрадиол (E 2 ). как следствие повышенной клеточной экспрессии белков, которые связываются с промоторами гена-мишени VDR / ERα, VDRE и ERE (10-12, 15).Избыток этих белков, называемых VDRE- и ERE-связывающими белками (VDRE-BP и ERE-BP, соответственно), означал, что требовались гораздо более высокие уровни 1,25 (OH) 2 D и E 2 для замещают связанные с хроматином VDRE-BP и ERE-BP, чтобы способствовать передаче сигналов рецептора. Очистка, идентификация и характеристика так называемых VDRE-BP и ERE-BP как двух отдельных членов суперсемейства hnRNP, hnRNP C1 / C2 и hnRNP C-подобных белков, соответственно, были выполнены в свое время (10, 15 ).Последующее молекулярное клонирование и сверхэкспрессия этих белков и идентификация их человеческих гомологов показали, что они являются 1] доминантно-отрицательными ингибиторами 1,25 (OH) 2 D-VDR- и E 2 -ERα-направленной транскрипции (51) и 2] взаимодействуют с другой категорией внутриклеточных белков-шаперонов в семействе белков расширенного теплового шока (hsp), которые связывают как лиганды, так и другие регуляторные белки для точной настройки контроля транскрипции над 1,25 (OH) 2 D- и E 2 -управляемые гены у нечеловеческих и человеческих приматов (16, 52).Мы назвали последний дуэт связанных с hsp витамином D и эстроген-связывающими белками как внутриклеточный витамин D-связывающий белок (конститутивно экспрессируемый hsc70) и внутриклеточный эстроген-связывающий белок (hsp27), соответственно. В оставшейся части обзора подробно описаны наблюдения, которые привели к открытиям белков, связывающих HRE, и белков, связывающих внутриклеточные гормоны, а также последующая работа, которая включила эти новые белки в сигнальные пути витамина D и эстрогена.
VI. Связывающий белок элемента ответа витамина D (VDRE-BP) и устойчивость к витамину D
Эндокринные прогормоны витамин D 3 , синтезируемый в эпидермисе в ответ на УФ-излучение, и диетический, синтезируемый растениями витамин D 2 (эргокальциферол) лишены какой-либо биологической активности (53). Вместо этого гормональная активность витамина D обусловлена в первую очередь дигидроксилированным метаболитом 1,25 (OH) 2 D, секостеролом, геномный механизм действия которого является общим для других стероидных гормонов, а именно опосредуется связыванием с VDR ( 53).В печени фермент витамин D 25-гидроксилаза катализирует начальное гидроксилирование витамина D на углероде 25 с образованием 25-гидроксивитамина D 2 или D 3 (25OHD), основной циркулирующей формы витамина D. Другой фермент цитохрома P450 25-гидроксивитамин D-1α-гидроксилаза (1α-гидроксилаза), продукт гена CYP27b1 , катализирует дальнейшее гидроксилирование 25OHD в активный гормональный 1,25 (OH) 2 D. Эта форма витамин D может затем связываться с VDR и образовывать комплекс с нелигандированным ретиноидным рецептором X (RXR), чтобы трансактивировать 1,25 (OH) 2 D-зависимых генов через VDRE, тем самым обеспечивая регулирование физиологических событий, таких как кальциевый гомеостаз, клеточная дифференцировка и пролиферация (54).
Нарушение регуляции метаболизма и / или действия витамина D может привести к нарушению минерализации костей в результате кишечной мальабсорбции кальция, гипокальциемии, вторичного гиперпаратиреоза, увеличения почечного клиренса фосфора и гипофосфатемии (). Формирование эндохондральной кости — это двухэтапный процесс: сначала формируется матрица хондроцитов (зачаток хряща), в которой остеобласты затем дифференцируются с образованием кости (55-56). Помимо эндокринной роли витамина D, хондроциты и остеобласты участвуют в аутокринных и паракринных петлях метаболизма витамина D, поскольку они способны превращать 25OHD в 1,25 (OH) 2 D, что позволяет предположить, что витамин D играет прямую роль в регулируют их деятельность и формирование эндохондральной кости (57-59) ().
Витамин D-зависимый рахит (VDDR) генетически неоднороден в зависимости от нарушений метаболизма или действия витамина D (60). VDDR типа 1A (OMIM 264700) возникает из-за ферментативного дефекта синтеза активного 1,25 (OH) 2 D, вызванного мутациями в гене CYP27B1 . VDDR1B (OMIM 600081) представляет собой форму рахита из-за мутаций в гене, кодирующем 25-гидроксилазу витамина D ( CYP2R1 ), еще один фермент, необходимый для синтеза активного витамина D 3 .Наследственный рахит, устойчивый к витамину D (HVDRR), также известный как витамин D-зависимый рахит 2 типа, является аутосомно-рецессивным заболеванием, которое вызвано невосприимчивостью органов-мишеней к активному витамину D, а в одном случае (VDDR2A) — мутациями в VDR (OMIM 277440) (61-62). В качестве альтернативы, VDDR2B (OMIM 600785) представляет собой необычную форму HVDRR, при которой невосприимчивость органа-мишени к активному витамину D происходит из-за нарушения регуляции hnRNP C1 / C2 (то есть VDRE-BP), который доминирует отрицательным образом влияет на функцию VDR. -ДНК взаимодействия (11-12, 63).В обоих случаях расстройства характеризуются тяжелым рахитом, гипокальциемией, вторичным гиперпаратиреозом, эпизодической алопецией, повышенными уровнями 1,25 (OH) 2 D и резистентностью к лечению с помощью 2,25 (OH) D (63-64 ).
hnRNPs представляют собой семейство повсеместно экспрессируемых ядерных ДНК- и (hn) РНК-связывающих белков, которые разделяют широкий спектр клеточных функций (65-67). По крайней мере, 20 из этих белков, обозначенных от A до U, в большом количестве присутствуют в ядре и обнаруживаются в ассоциации с белками с образованием частиц hnRNP.Члены семейства hnRNP и большое количество сопутствующих изоформ служат для опосредования процессинга пре-мРНК и сплайсинга, упаковки, экспорта и стабильности мРНК, а также ремоделирования хроматина посредством ремоделирующих комплексов, связанных с областью контроля локуса, рекомбинации и удлинения теломер (65). Функциональная роль белков hnRNP влияет на клинические и биологические исходы вирусных инфекций (68-69), спинальной мышечной атрофии (70), миотонической дистрофии (71), болезни Альцгеймера (72), туморогенеза и устойчивости к гормонам (11-12, 14). , 65).
У позвоночных hnRNPC принадлежит к подсемейству hnRNPs как основной компонент 40S hnRNP частиц; Было описано несколько вариантов транскриптов, кодирующих по крайней мере две разные изоформы (C1 и C2) (http://ensembl.org). Белок hnRNPC действует как тетрамер [(C1) 3 C2], и две изоформы представляют собой антигенно близкородственные фосфопротеины, которые прочно связываются с РНК in vitro (73-74). Белки hnRNPC делятся на отдельные домены: 1) N-концевой высококонсервативный РНК-связывающий домен (RBD), также называемый мотивом распознавания РНК (RRM), 2) центральный вариабельный домен и 3) C-концевой вспомогательный домен. домен (67).Возможно, что эти отдельные регионы сами по себе могут иметь уникальную структуру и функцию. Также возможно, что эти области могут функционировать совместно через взаимодействие или физическую связь между отдельными регионами.
HnRNP C1 / C2, как известно, играет плейотропную роль в регулировании ряда биологических активностей, от классических функций, описанных выше, до прямого ингибирования апоптоза путем взаимодействия с множеством белков (75), хотя большая часть механизма остается неизвестной.Несмотря на их классическое взаимодействие с ассоциированной с хроматином одноцепочечной РНК, несколько исследований показали, что hnRNPs являются плюрипотентными белками. В частности, они могут также функционировать как трансрегуляторные факторы посредством взаимодействия с двухцепочечной ДНК (33), что ставит вопрос о том, могут ли они также быть частью нормального механизма регуляции генов, опосредованного VDR. ChIP-исследования нормальных 1,25 (OH) 2 D-чувствительных клеток показали, что это действительно так, поскольку человеческая hnRNP C1 / C2 занимает VDRE главного гена-мишени витамина D CYP24A1 (1,25 -дигидроксивитамин D 3 24-гидроксилаза), участвующий в катаболизме метаболитов, до обработки 1,25 (OH) 2 D, а затем вытесняемый с промотора лигандным комплексом VDR-RXR (11).Эта реципрокная взаимосвязь между VDR и VDRE-BP происходит в зависимости от времени циклически после воздействия 1,25 (OH) 2 D, что позволяет предположить, что hnRNP C1 / C2 играет ключевую роль в пространственно-временной организации рецептора. -элемент ответа комплексный. Важно отметить, что hnRNP C1 / C2, как было показано, регулирует действия 1,25 (OH) 2 D у пациента с формой HVDRR из-за его клеточной сверхэкспрессии, что приводит к фенотипу, аналогичному УФB-ограниченным NWP, исследованным ранее (11- 12, 63).В этом природном эксперименте было показано, что избыток hnRNP C1 / C2 мешает связыванию VDR с VDRE, расположенным в проксимальной области промотора CYP24A1 . Впоследствии, в ходе исследований in vitro сверхэкспрессии с использованием репортерной системы транскрипции, было показано, что hnRNP C1 / C2 ингибирует трансактивацию витамина D, что является дополнительным доказательством такого механизма. Chen et al. (12) предположили, что устойчивость к витамину D у этого пациента была аналогична устойчивости, описанной в NWP, в которых аномальная экспрессия HRE-связывающего белка вызывала устойчивость клеток-мишеней к 1,25 (OH) 2 D (76).
VII. Связывающий белок эстрогенового ответа (ERE-BP) и устойчивость к эстрогену
NWP проявляют устойчивость к другим стероидным гормонам в дополнение к витамину D, в частности, подавляя нормальную транскрипцию, направленную на эстрадиол-ER-ERE (9). В результате исследования, аналогичные тем, которые используются для анализа устойчивости к витамину D у NWP, были выполнены для оценки передачи сигналов ядерного ER. В этом случае были подвергнуты скринингу концентрированные ядерные экстракты из устойчивых к эстрогену клеток NWP, и C-подобный белок hnRNP был идентифицирован как ERE-BP (15).Функциональность ERE-BP была продемонстрирована в исследованиях как in vitro , так и in vivo (13, 51). В случае последнего были созданы линии трансгенных мышей с различной степенью сверхэкспрессии ERE-BP, которые подтвердили, что тканеспецифическая сверхэкспрессия ERE-BP приводит к устойчивости к эстрогену in vivo (51). Учитывая фундаментальную роль эстрогена в репродукции, было высказано предположение, что глобальная сверхэкспрессия гена устойчивости к эстрогену может иметь летальные последствия.Все потомство, рожденное от матерей дикого типа и матерей с «низким» ERE-BP, было жизнеспособным, тогда как треть всех детенышей, рожденных от матерей с «промежуточным» ERE-BP, умерли до отъема. Примечательно, что 100% потомков, рожденных от матерей с «высоким» ERE-BP, умерли к 7 дню послеродового периода (51). Дальнейший анализ показал, что смертность была вызвана голоданием в результате уменьшения потребления грудного молока матерями с избыточной экспрессией ERE-BP; у детенышей, рожденных от матерей с высоким уровнем экспрессии ERE-BP, наблюдалось обратное увеличение массы тела, если они были воспитаны матерью дикого типа.
Гистологический анализ молочных желез девственных или кормящих самок мышей показал, что сверхэкспрессия ERE-BP приводит к уменьшению количества протоков и ветвей молочных желез. Было высказано предположение, что повышенная экспрессия ERE-BP приводит к нечувствительности E 2 и нарушению развития груди. Параллельные исследования, проведенные in vitro , показали, что сверхэкспрессия ERE-BP резко притупляет ответы на E 2 , хотя при использовании E 2 можно было индуцировать значительную передачу сигналов ERα-ERE в сверхэкспрессирующих ERE-BP клетках. в больших количествах способствует развитию груди и лактации (51).
На основании вышеописанного наблюдения, что E 2 может спасти ERE-BP-опосредованное подавление экспрессии ERα-управляемого гена in vitro , были проведены дальнейшие исследования с использованием трансгенных мышей ERE-BP, в которых животным давали медленную — высвобождение подкожно гранул E 2 для дифференциального повышения уровней E 2 в сыворотке (76). Полный и гистологический анализ ткани груди показал, что воздействие более высоких дополнительных доз E 2 «спасало» нарушенное развитие молочной железы у мышей с избыточной экспрессией ERE-BP.Спасение развития молочных желез у девственных нелактирующих самок мышей, а также у кормящих матерей было достигнуто с помощью E 2 , что указывает на то, что доминантно-негативный эффект ERE-BP активен на разных стадиях функции груди. Существенно и неожиданно, что эти исследования показали, что тамоксифен был так же эффективен, как и E 2 , в противодействии ингибирующим эффектам ERE-BP, предполагая, что селективный модулятор рецептора эстрогена (SERM) способен оказывать эффекты на действие эстрогена, отличные от его действия. эффекты на ERα или его вспомогательные, ко-регуляторные белки.Дальнейшее прояснение этого наблюдения было получено в экспериментах, проведенных in vitro , в которых способность ERα и ERE-BP взаимодействовать с хроматином оценивалась с помощью ChIP-анализа клеток молочной железы MCF-7 дикого типа и сверхэкспрессирующих ERE-BP. обработанные E 2 или тамоксифеном (76). Как и в случае hnRNP C1 / C2 VDRE-BP (11), клетки, сверхэкспрессирующие ERE-BP, демонстрировали нарушение регуляции пространственно-временных паттернов занятости ERE; как E 2 , так и тамоксифен были способны корректировать эту дисрегуляцию и способствовать связыванию ERα с геном pS2 ERE.Эти данные подтвердили обратимый характер физического взаимодействия между С-подобными белками hnRNP и регуляторными элементами рецептора цис .
VIII. Заключение
Со временем механизмы, лежащие в основе контроля действия и метаболизма гормонов в клетках-мишенях, продолжают расширяться, иногда в неожиданных направлениях. Открытие связанных с hnRNPC белков, связывающих витамин D и эстрогеновый ответный элемент, у приматов и людей является таким достижением.Потенциал этих многофункциональных связывающих нуклеиновые кислоты белков изменять гормонально-направленную транскрипцию генов в дополнение к контролю посттранскрипционной судьбы мРНК, транскрибируемых из этих генов, указывает на то, что традиционные взгляды на различные ядерные машины (т.е. транскриптосомы, сплайсосомы и т. Д.) работа независимо друг от друга может потребовать доработки. Дальнейшие исследования будут направлены на выяснение механизмов, с помощью которых VDRE-BP и ERE-BP связываются с HRE предполагаемых промоторов генов, чтобы направлять транскрипцию генов для критических генов-мишеней VDR и ER.Подобные механизмы также, по-видимому, являются центральными для передачи сигналов другими стероидными гормонами, обеспечивая дополнительную поддержку этих новых ко-фасилитаторов и комодуляторов в качестве ключевых компонентов передачи сигналов ядерных рецепторов.
IX. Выражение признательности
Эта работа была поддержана грантом NIH RO1AR37399-23 в JSA
X. Сокращения
NWP
Примат Нового Света
OWP
BP
OWP
BP 9070 Примат 9018 Старого Света 9018 Белок, связывающий элемент ответа на витамин D
ERE-BP
Белок, связывающий элемент ответа на эстроген
hnRNP
гетерогенный ядерный рибонуклеопротеин
Foot HVDRR
902 902 наследственный витамин D-резистентный 902 Заявление издателя: Это PDF-файл неотредактированной рукописи, принятой к публикации.В качестве услуги для наших клиентов мы предоставляем эту раннюю версию рукописи. Рукопись будет подвергнута копированию, верстке и рассмотрению полученного доказательства, прежде чем она будет опубликована в окончательной форме для цитирования. Обратите внимание, что во время производственного процесса могут быть обнаружены ошибки, которые могут повлиять на содержание, и все юридические оговорки, относящиеся к журналу, имеют отношение.
XI. Ссылки
1. Alarid ET. Жизни и времена ядерных рецепторов. Мол Эндокринол. 2006. 20 (9): 1972–1981.[PubMed] [Google Scholar] 3. Новак Н., Хайнцель Т. Ядерные рецепторы: обзор и классификация. Препарат Curr предназначен для лечения аллергии на воспаление. 2004. 3 (4): 335–346. [PubMed] [Google Scholar] 4. Такур М.К., Параманик В. Роль корегуляторов стероидных гормонов в здоровье и болезнях. Horm Res. 2009. 71 (4): 194–200. [PubMed] [Google Scholar] 5. Пеннинг TM, Джин И, Ризнер Т.Л., Бауман ДР. Пререцепторная регуляция рецептора андрогенов. Mol Cell Endocrinol. 2008. 281 (1-2): 1–8. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Томлинсон Дж. В. и др.11бета-гидроксистероиддегидрогеназа типа 1: тканеспецифический регулятор глюкокортикоидного ответа. Endocr Rev.2004; 25 (5): 831–866. [PubMed] [Google Scholar] 7. Адамс Дж. С. и др. Белки, связывающие элемент ответа, и внутриклеточные белки, связывающие витамин D: новые регуляторы перемещения, действия и метаболизма витамина D. J Стероид Biochem Mol Biol. 2004; 89-90 (1-5): 461–465. В переводе с англ. на англ. [PubMed] [Google Scholar] 8. Arbelle JE, et al. Ингибирование образования комплекса рецептор витамина D-рецептор ретиноида X-витамин D-ответный элемент ядерными экстрактами устойчивых к витамину D клеток приматов Нового Света.Эндокринология. 1996. 137 (2): 786–789. [PubMed] [Google Scholar] 9. Чен Х, Арбель Дж. Э., Гакад М. А., Аллегретто Е. А., Адамс Дж. С.. Клетки приматов Нового Света, устойчивые к витамину D и гонадным стероидам, экспрессируют внутриклеточный белок, который конкурирует с рецептором эстрогена за связывание с элементом ответа эстрогена. J Clin Invest. 1997. 99 (4): 669–675. [Бесплатная статья PMC] [PubMed] [Google Scholar] 10. Чен Х, Ху Б., Аллегретто Э.А., Адамс Дж. С.. Белок, связывающий элемент ответа на витамин D. Новый доминантно-негативный регулятор трансактивации витамина D.J Biol Chem. 2000. 275 (45): 35557–35564. [PubMed] [Google Scholar] 11. Чен Х, Хьюисон М, Адамс Дж. С. Функциональная характеристика гетерогенного ядерного рибонуклеарного белка C1 / C2 при резистентности к витамину D: новый белок, связывающий элемент ответа. J Biol Chem. 2006. 281 (51): 39114–39120. [PubMed] [Google Scholar] 12. Чен Х., Хьюисон М., Ху Б., Адамс Дж. С.. Связывание гетерогенного ядерного рибонуклеопротеина (hnRNP) с элементами гормонального ответа: причина устойчивости к витамину D. Proc Natl Acad Sci U S. A. 2003; 100 (10): 6109–6114.В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 13. Чен Х, Клеменс Т.Л., Хьюисон М., Адамс Дж. С.. Эстрадиол и тамоксифен опосредуют устранение доминантно-негативных эффектов белка, связывающего элемент эстрогенового ответа, in vivo и in vitro. Эндокринология. 2009. 150 (5): 2429–2435. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 14. Чен Х, Хьюисон М, Адамс Дж. С. Контроль трансактивации гена, направленной на эстрадиол, с помощью внутриклеточного белка, связывающего эстроген, и белка, связывающего элемент ответа на эстроген.Мол Эндокринол. 2008. 22 (3): 559–569. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Чен Х, Ху Б., Гакад М.А., Адамс Дж. С.. Клонирование и экспрессия нового связывающего элемент эстрогенового ответа доминантно-отрицательно действующего белка в семействе гетерогенных ядерных рибонуклеопротеинов. J Biol Chem. 1998. 273 (47): 31352–31357. В переводе с англ. на англ. [PubMed] [Google Scholar] 16. Чен Х и др. Очистка и характеристика нового внутриклеточного связывающего 17-бета-эстрадиол белка в устойчивых к эстрогену клетках приматов Нового Света.J Clin Endocrinol Metab. 2003. 88 (1): 501–504. В переводе с англ. на англ. [PubMed] [Google Scholar] 17. Робинсон-Рехави М., Эскрива Гарсия Х., Лауде В. Суперсемейство ядерных рецепторов. J Cell Sci. 2003. 116 (Pt 4): 585–586. В переводе с англ. на англ. [PubMed] [Google Scholar] 18. Ордонез-Моран П., Муньос А. Ядерные рецепторы: сходятся геномные и негеномные эффекты. Клеточный цикл. 2009. 8 (11): 1675–1680. В переводе с англ. на англ. [PubMed] [Google Scholar] 19. Gronemeyer H, Gustafsson JA, Laudet V. Принципы модуляции надсемейства ядерных рецепторов.Nat Rev Drug Discov. 2004. 3 (11): 950–964. В переводе с англ. на англ. [PubMed] [Google Scholar] 20. Whitfield GK, Jurutka PW, Haussler CA, Haussler MR. Рецепторы стероидных гормонов: эволюция, лиганды и молекулярные основы биологической функции. J Cell Biochem. 1999; 32-33 (Дополнение): 110–122. В переводе с англ. на англ. [PubMed] [Google Scholar] 21. Чен Т. Открытие лекарств с ядерными рецепторами. Curr Opin Chem Biol. 2008. 12 (4): 418–426. В переводе с англ. на англ. [PubMed] [Google Scholar] 22. Freedman LP. Повышение сложности коактивации передачи сигналов ядерного рецептора.Клетка. 1999. 97 (1): 5–8. В переводе с англ. на англ. [PubMed] [Google Scholar] 23. Xu J, Li Q. Обзор функций семейства коактиваторов стероидных рецепторов p160 in vivo. Мол Эндокринол. 2003. 17 (9): 1681–1692. В переводе с англ. на англ. [PubMed] [Google Scholar] 24. Heery DM, Kalkhoven E, Hoare S, Parker MG. Сигнатурный мотив в коактиваторах транскрипции опосредует связывание с ядерными рецепторами. Природа. 1997. 387 (6634): 733–736. В переводе с англ. на англ. [PubMed] [Google Scholar] 25. Вегель Э, Шоу П.Активация и дезактивация генов связаны с изменениями трехмерной структуры хроматина. Хромосома. 2005. 114 (5): 331–337. В переводе с англ. на англ. [PubMed] [Google Scholar] 26. Диллон Н. Регуляция генов и крупномасштабная организация хроматина в ядре. Chromosome Res. 2006. 14 (1): 117–126. В переводе с англ. на англ. [PubMed] [Google Scholar] 27. Адамс Дж. С., Гакад М. А., Бейкер А. Дж., Кхеун Дж., Грубый Р. К.. Снижение интернализации и действия 1,25-дигидроксивитамина D3 в дермальных фибробластах, культивируемых от приматов Нового Света.Эндокринология. 1985. 116 (6): 2523–2527. В переводе с англ. на англ. [PubMed] [Google Scholar] 28. Адамс Дж. С. и др. Новые регуляторы действия и метаболизма витамина D: уроки, полученные в зоопарке Лос-Анджелеса. J Cell Biochem. 2003. 88 (2): 308–314. В переводе с англ. на англ. [PubMed] [Google Scholar] 29. Гуо Б. и др. YY1 регулирует трансактивацию, опосредованную рецептором витамина D / рецептором ретиноида X, гена остеокальцина, чувствительного к витамину D. Proc Natl Acad Sci U S. A. 1997; 94 (1): 121–126. В переводе с англ.на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Oesterreich S, et al. Новый белок ядерного матрикса HET связывается с промотором HSP27 и влияет на его активность в клетках рака молочной железы человека. J Cell Biochem. 1997. 67 (2): 275–286. В переводе с англ. на англ. [PubMed] [Google Scholar] 31. Castellano G, et al. Участие фактора транскрипции Инь Ян 1 в развитии и прогрессировании рака. Клеточный цикл. 2009. 8 (9): 1367–1372. В переводе с англ. на англ. [PubMed] [Google Scholar] 32. Шахтер Дж. Н., Ямамото КР.Основная область AP-1 определяет активность рецептора глюкокортикоидов в составном элементе ответа. Genes Dev. 1992. 6 (12B): 2491–2501. В переводе с англ. на англ. [PubMed] [Google Scholar] 33. Шахтер Дж. Н., Ямамото КР. Регулирующие перекрестные помехи в составных ответных элементах. Trends Biochem Sci. 1991. 16 (11): 423–426. В переводе с англ. на англ. [PubMed] [Google Scholar] 34. Беато М. Генная регуляция стероидными гормонами. Клетка. 1989. 56 (3): 335–344. В переводе с англ. на англ. [PubMed] [Google Scholar] 35. Diamond MI, Miner JN, Yoshinaga SK, Yamamoto KR.Взаимодействия факторов транскрипции: селекторы положительной или отрицательной регуляции от одного элемента ДНК. Наука. 1990. 249 (4974): 1266–1272. В переводе с англ. на англ. [PubMed] [Google Scholar] 36. Retnakaran R, Flock G, Giguere V. Идентификация RVR, нового орфанного ядерного рецептора, который действует как негативный регулятор транскрипции. Мол Эндокринол. 1994. 8 (9): 1234–1244. В переводе с англ. на англ. [PubMed] [Google Scholar] 37. Кавай К. и др. Нелигандированный рецептор-бета1 гормона щитовидной железы подавляет альфа / оксистерин-зависимую трансактивацию рецептора Х печени.Эндокринология. 2004. 145 (12): 5515–5524. В переводе с англ. на англ. [PubMed] [Google Scholar] 38. Swales KE, et al. Рецептор фарнезоида X экспрессируется при раке груди и регулирует апоптоз и экспрессию ароматазы. Cancer Res. 2006. 66 (20): 10120–10126. В переводе с англ. на англ. [PubMed] [Google Scholar] 39. Каталано С. и др. Рецептор Фарнезоида X через связывание с элементом, чувствительным к стероидогенному фактору 1, подавляет экспрессию ароматазы в опухолевых клетках Лейдига. J Biol Chem. 2010. 285 (8): 5581–5593.В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 40. Кино Т., Хёрт Д.Э., Ичидзё Т., Надер Н., Хрусос Г.П. Некодирующая РНК gas5 является репрессором глюкокортикоидного рецептора, связанным с остановкой роста и голоданием. Sci Signal. 2010; 3 (107): ra8. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41. Музикар К.А., Николс Н.Г., Дерван ПБ. Репрессия зависимой от связывания ДНК экспрессии гена, опосредованной рецептором глюкокортикоидов. Proc Natl Acad Sci U S. A. 2009; 106 (39): 16598–16603.В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 42. Никольс Н.Г., Дерван ПБ. Подавление экспрессии гена, опосредованной рецептором андрогена, с помощью полиамида, связывающего ДНК с последовательностью. Proc Natl Acad Sci U S. A. 2007; 104 (25): 10418–10423. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Вингаард А.М., Хнида С., Брейнхольт В., Ларсен Дж. Скрининг отобранных пестицидов на предмет ингибирования активности ароматазы CYP19 in vitro. Toxicol In Vitro. 2000. 14 (3): 227–234.В переводе с англ. на англ. [PubMed] [Google Scholar] 44. Кининис М, Краус ВЛ. Общий взгляд на регуляцию транскрипции ядерными рецепторами: экспрессия генов, локализация факторов и анализ последовательности ДНК. Сигнал рецепта Nucl. 2008; 6: e005. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Розенфельд М.Г., Луняк В.В., Стекло СК. Датчики и сигналы: коактиватор / корепрессор / эпигенетический код для интеграции сигнально-зависимых программ транскрипционного ответа. Genes Dev. 2006. 20 (11): 1405–1428.В переводе с англ. на англ. [PubMed] [Google Scholar] 46. Pike JW, et al. Новые регуляторные парадигмы для контроля экспрессии генов 1,25-дигидроксивитамином D3. J Стероид Biochem Mol Biol. 2010. 121 (1-2): 130–135. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Chrousos GP, et al. Приматы нового мира как животные модели устойчивости к глюкокортикоидам. Adv Exp Med Biol. 1986; 196: 129–144. В переводе с англ. на англ. [PubMed] [Google Scholar] 48. Chrousos GP. Новый «новый» синдром в новом мире: обусловлена ли множественная пострецепторная резистентность к стероидным гормонам дефектом корегулятора? J Clin Endocrinol Metab.1999. 84 (12): 4450–4453. В переводе с англ. на англ. [PubMed] [Google Scholar] 49. Адамс Дж. С., Гакад М. А., Бейкер А. Дж., Гонсалес Б., Грубый Р. К.. Концентрация 1,25-дигидроксивитамина D3 в сыворотке крови у Platyrrhini и Catarrhini: филогенетическая оценка. Am J Primatol. 1985; 19: 219–224. [Google Scholar] 50. Адамс Дж. С., Гакад Массачусетс. Фенотипическое разнообразие взаимодействия клеточного 1,25-дигидроксивитамина D3-рецептора у разных родов приматов Нового Света. J Clin Endocrinol Metab. 1988. 66 (1): 224–229. В переводе с англ.на англ. [PubMed] [Google Scholar] 51. Чен Х и др. Создание устойчивости к эстрогену in vivo за счет сверхэкспрессии трансгенной структуры гетерогенного ядерного белка, связывающего элемент ответа на эстроген, связанный с рибонуклеопротеином. Эндокринология. 2005. 146 (10): 4266–4273. [PubMed] [Google Scholar] 52. Wu S, et al. Внутриклеточные белки, связывающие витамин D: новые средства, способствующие трансактивации витамина D. Мол Эндокринол. 2000. 14 (9): 1387–1397. В переводе с англ. на англ. [PubMed] [Google Scholar] 53. Bouillon R, et al.Витамин D и здоровье человека: уроки мышей с нулевым рецептором витамина D. Endocr Rev.2008; 29 (6): 726–776. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 54. Takeyama K, et al. 25-гидроксивитамин D3 1альфа-гидроксилаза и синтез витамина D. Наука. 1997. 277 (5333): 1827–1830. В переводе с англ. на англ. [PubMed] [Google Scholar] 55. Карсенти Г. Сложности биологии скелета. Природа. 2003. 423 (6937): 316–318. В переводе с англ. на англ. [PubMed] [Google Scholar] 56. Kronenberg HM.Регуляция развития пластинки роста. Природа. 2003. 423 (6937): 332–336. В переводе с англ. на англ. [PubMed] [Google Scholar] 57. Андерсон PH, Аткинс GJ. Скелет как внутрикринный орган метаболизма витамина D. Мол Аспекты Мед. 2008. 29 (6): 397–406. В переводе с англ. на англ. [PubMed] [Google Scholar] 58. ван Дриэль М., Полс Х.А., ван Леувен Дж. П. Дифференциация и контроль остеобластов с помощью метаболитов витамина D и витамина D. Curr Pharm Des. 2004. 10 (21): 2535–2555. В переводе с англ. на англ.[PubMed] [Google Scholar] 59. van Driel M, et al. Доказательства ауто / паракринного действия витамина D в костях: экспрессия и активность альфа-гидроксилазы в костных клетках человека. FASEB J. 2006; 20 (13): 2417–2419. В переводе с англ. на англ. [PubMed] [Google Scholar] 60. Bouillon R, et al. Устойчивость к витамину D. Лучшая практика Res Clin Endocrinol Metab. 2006. 20 (4): 627–645. В переводе с англ. на англ. [PubMed] [Google Scholar] 61. Фельдман Д., Маллой П.Дж. Наследственный рахит, устойчивый к 1,25-дигидроксивитамину D: молекулярная основа и значение роли 1,25 (OH) 2D3 в нормальной физиологии.Mol Cell Endocrinol. 1990. 72 (3): C57–62. В переводе с англ. на англ. [PubMed] [Google Scholar] 62. Маллой П.Дж., Фельдман Д. Генетические нарушения и дефекты действия витамина D. Endocrinol Metab Clin North Am. 2010. 39 (2): 333–346. В переводе с англ. содержание (на англ. яз.) [Бесплатная статья PMC] [PubMed] [Google Scholar] 63. Хьюисон М. и др. Устойчивость тканей к 1,25-дигидроксивитамину D без мутации гена рецептора витамина D. Clin Endocrinol (Oxf) 1993; 39 (6): 663–670. В переводе с англ. на англ.[PubMed] [Google Scholar] 64. Brooks MH и др. Витамин-D-зависимый рахит II типа. Устойчивость органов-мишеней к 1,25-дигидроксивитамину D. N Engl J Med. 1978; 298 (18): 996–999. В переводе с англ. на англ. [PubMed] [Google Scholar] 65. Кречич AM, Swanson MS. Комплексы гяРНП: состав, строение и функции. Curr Opin Cell Biol. 1999. 11 (3): 363–371. В переводе с англ. на англ. [PubMed] [Google Scholar] 66. Карпентер Б. и др. Роль гетерогенных ядерных рибонуклеопротеидов в развитии и прогрессировании опухоли.Biochim Biophys Acta. 2006; 1765 (2): 85–100. В переводе с англ. на англ. [PubMed] [Google Scholar] 67. Дрейфус Г., Матунис М.Дж., Пинол-Рома С, Бурд К.Г. белки hnRNP и биогенез мРНК. Анну Рев Биохим. 1993; 62: 289–321. В переводе с англ. на англ. [PubMed] [Google Scholar] 68. Ким К.С., Сеол СК, Сон ОК, Пак Дж.Х., Чан СК. РНК-связывающий белок hnRNP A1 и каркасный белок септин 6 способствуют репликации вируса гепатита С. J Virol. 2007. 81 (8): 3852–3865. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 69.Бруннер Дж. Э., Эртель К. Дж., Розович Дж. М., Семлер Б. Л.. Задержка кинетики синтеза РНК полиовируса в линии клеток человека с пониженными уровнями белков hnRNP C. Вирусология. 2010. 400 (2): 240–247. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 70. Россолл В. и др. Специфическое взаимодействие Smn, продукта гена, определяющего мышечную атрофию позвоночника, с hnRNP-R и gry-rbp / hnRNP-Q: роль Smn в процессинге РНК в моторных аксонах? Hum Mol Genet. 2002. 11 (1): 93–105. В переводе с англ.на англ. [PubMed] [Google Scholar] 71. Nezu Y, Kino Y, Sasagawa N, Nishino I, Ishiura S. Экспрессия транскриптов мРНК MBNL и CELF в мышцах с миотонической дистрофией. Нервно-мышечное расстройство. 2007. 17 (4): 306–312. В переводе с англ. на англ. [PubMed] [Google Scholar] 72. Ли Е.К. и др. hnRNP C способствует трансляции APP, конкурируя с FMRP за рекрутирование мРНК APP в P-тельца. Nat Struct Mol Biol. 2010. 17 (6): 732–739. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 73. Накагава Т.Ю., Суонсон М.С., Уолд Б.Дж., Дрейфус Г.Молекулярное клонирование кДНК ядерных рибонуклеопротеиновых частиц C белков: консервативное семейство генов. Proc Natl Acad Sci U S. A. 1986; 83 (7): 2007–2011. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 74. Merrill BM, Barnett SF, LeStourgeon WM, Williams KR. Различия в первичной структуре белков C1 и C2 ядерных рибонуклеопротеиновых частиц HeLa 40S. Nucleic Acids Res. 1989. 17 (21): 8441–8449. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 75.Ромеро Ф. и др. Grb2 и его апоптотическая изоформа Grb3-3 связываются с гетерогенным ядерным рибонуклеопротеином C, и эти взаимодействия модулируются поли (U) РНК. J Biol Chem. 1998. 273 (13): 7776–7781. В переводе с англ. на англ. [PubMed] [Google Scholar] 76. Адамс JSCH, Лиссе Т.С., Чун Р.Ф., Хьюисон М. Рецепторно-независимая резистентность к витамину D у нечеловеческих и человеческих приматов. В: MF H, редактор. Питание и здоровье: витамин D. 1-е изд. Springer Science; 2010. С. 715–728. [Google Scholar] 77. Gacad MA, Chen H, Arbelle JE, LeBon T, Adams JS.Функциональная характеристика и очистка внутриклеточного витамин D-связывающего белка в устойчивых к витамину D клетках приматов нового мира. Гомология аминокислотной последовательности с белками семейства hsp-70. J Biol Chem. 1997. 272 (13): 8433–8440. В переводе с англ. на англ. [PubMed] [Google Scholar] 78. Gacad MA, Adams JS. Белки семейства теплового шока-70 специфически связывают 25-гидроксивитамин D3 и 17-бета-эстрадиол. J Clin Endocrinol Metab. 1998. 83 (4): 1264–1267. В переводе с англ. на англ. [PubMed] [Google Scholar] 79.Gacad MA, Adams JS. Эндогенная блокада связывания 1,25-дигидроксивитамина D-рецептора в клетках приматов Нового Света. J Clin Invest. 1991. 87 (3): 996–1001. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 80. Hartl FU. Молекулярные шапероны в сворачивании клеточных белков. Природа. 1996. 381 (6583): 571–579. В переводе с англ. на англ. [PubMed] [Google Scholar] 81. Лутц В., Коно К., Кумар Р. Роль белка теплового шока 70 в функции рецептора витамина D. Biochem Biophys Res Commun. 2001. 282 (5): 1211–1219.В переводе с англ. на англ. [PubMed] [Google Scholar] 82. Wu S, et al. Регулирование синтеза 1,25-дигидроксивитамина d внутриклеточным белком-1, связывающим витамин d. Эндокринология. 2002; 143 (10): 4135. В переводе с англ. на англ. [PubMed] [Google Scholar] 83. Чун Р., Гакад М.А., Хьюисон М., Адамс Дж. С.. Аденозин-5′-трифосфат-зависимое связывание стерола витамина D с шаперонами белка теплового шока-70. Эндокринология. 2005. 146 (12): 5540–5544. В переводе с англ. на англ. [PubMed] [Google Scholar] 84. Каббидж М., Дикман МБ.Белки BAG: повсеместное семейство регуляторов шаперонов. Cell Mol Life Sci. 2008. 65 (9): 1390–1402. В переводе с англ. на англ. [PubMed] [Google Scholar] 85. Чун Р.Ф., Гакад М., Нгуен Л., Хьюисон М., Адамс Дж. С.. Ко-шаперонное усиление трансактивации, опосредованной рецептором витамина D: роль Bcl2-ассоциированного атаногена-1 как внутриклеточного связывающего белка для 1,25-дигидроксивитамина D3. J Mol Endocrinol. 2007. 39 (2): 81–89. В переводе с англ. на англ. [PubMed] [Google Scholar] 86. Тагами Т., Лутц WH, Кумар Р., Джеймсон Дж. Л..Взаимодействие рецептора витамина D с корепрессорами и коактиваторами ядерных рецепторов. Biochem Biophys Res Commun. 1998. 253 (2): 358–363. В переводе с англ. на англ. [PubMed] [Google Scholar] 87. Норман А.В. Миниобзор: рецептор витамина D: новые назначения уже занятому рецептору. Эндокринология. 2006. 147 (12): 5542–5548. В переводе с англ. на англ. [PubMed] [Google Scholar] 88. Malloy PJ, et al. Наследственный рахит, устойчивый к витамину D, вызванный новой мутацией рецептора витамина D, которая приводит к снижению сродства к гормонам и гипореактивности клеток.J Clin Invest. 1997. 99 (2): 297–304. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 89. Arcidiacono MV, et al. Активация EGFR увеличивает гиперплазию паращитовидных желез и резистентность к кальцитриолу при заболевании почек. J Am Soc Nephrol. 2008. 19 (2): 310–320. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 90. Fountas L и др. Вклад полиморфизма гена рецептора витамина D в остеопороз и семейный остеопороз. Osteoporos Int. 1999. 10 (5): 392–398.В переводе с англ. на англ. [PubMed] [Google Scholar] 91. Зинеб Р., Жор Б., Одиллия В., Марта Р. Четкая тканеспецифическая регуляция рецептора витамина D в кишечнике, почках и коже с помощью диетического кальция и витамина D. Эндокринология. 1998. 139 (4): 1844–1852. В переводе с англ. на англ. [PubMed] [Google Scholar] 92. Чен Л. и др. Генетические варианты рецептора витамина d связаны с распространенным раком простаты на момент постановки диагноза: результаты исследования простаты на предмет выявления рака и лечения, а также систематический обзор.Биомаркеры эпидемиологии рака Пред. 2009. 18 (11): 2874–2881. В переводе с англ. на англ. [PubMed] [Google Scholar] 93. Гай М. и др. Полиморфизм гена рецептора витамина D и риск рака груди. Clin Cancer Res. 2004. 10 (16): 5472–5481. В переводе с англ. на англ. [PubMed] [Google Scholar] 94. Накадзима С. и др. Снижение концентрации рецепторов 1,25-дигидроксивитамина D3 в периферических мононуклеарных клетках пациентов с Х-сцепленным гипофосфатемическим рахитом: эффект приема фосфатов. Костяной шахтер.1990. 10 (3): 201–209. В переводе с англ. на англ. [PubMed] [Google Scholar] 95. Rocha JC и др. Фармакогенетика исходов у детей с острым лимфобластным лейкозом. Кровь. 2005. 105 (12): 4752–4758. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 96. Макдоннелл Д.П., Норрис Д.Д. Связи и регуляция рецептора эстрогена человека. Наука. 2002. 296 (5573): 1642–1644. В переводе с англ. на англ. [PubMed] [Google Scholar] 97. Alcazar LP, Аракаки PA, Годой-Сантос А, Сантос М.Полиморфизм рецепторов эстрогенов и его связь с патологическим процессом. Am J Med Sci. 2010. 340 (2): 128–132. В переводе с англ. на англ. [PubMed] [Google Scholar] 98. Alquobaili F, et al. Зависимые от рецептора эстрогена профили экспрессии генома в клетках рака груди в ответ на жирные кислоты. J Carcinog. 2010; 8: 17. В переводе с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar] 99. Лебо А. и др. Амплификация гена рецептора эстрогена (ESR1) часто встречается при карциноме эндометрия и ее предшественниках.J Pathol. 2008. 216 (2): 151–157. В переводе с англ. на англ. [PubMed] [Google Scholar] 100. Asada H, et al. Возможная связь между гипометилированием гена рецептора эстрогена-альфа и образованием миомы матки. Мол Хум Репрод. 2008. 14 (9): 539–545. В переводе с англ. на англ. [PubMed] [Google Scholar] 101. Лу Х и др. Связь полиморфизма гена рецептора эстрогена-альфа с ишемической болезнью сердца у пациентов с семейной гиперхолестеринемией. Артериосклер Thromb Vasc Biol. 2002. 22 (5): 817–823.В переводе с англ. на англ. [PubMed] [Google Scholar] 102. Лиссе Т.С. и др. Нацеливание на ген с помощью связывающего белка элемента ответа витамина D выявляет роль витамина D в передаче сигналов mTOR остеобластов. FASEB J. 2010 Перевод с англ. на англ. [Бесплатная статья PMC] [PubMed] [Google Scholar]
гормонально-зависимых генов в сигнальных путях SHH и WNT / β-катенина влияют на закрытие уретры и рост фаллоса † | Биология размножения
Аннотация
Экологические эндокринные разрушители (EED), которые влияют на активность андрогенов или эстрогенов, могут нарушать регуляцию генов во время развития фаллоса, вызывая гипоспадию или маскулинизированный клитор.Мы лечили развивающихся таммарных валлаби мужского пола эстрогеном, а женщин — андрогеном с 20-40-го дня после родов (пп) во время окна чувствительности импринтинга андрогенов. Эстроген подавлял удлинение фаллоса, но не влиял на закрытие уретры и не подавлял в значительной степени синтез андрогенов в яичках. Лечение андрогенами у женщин не способствовало удлинению фаллоса, но инициировало закрытие уретры. Фаллосы собирали для секвенирования транскриптома на 50-й день, когда они впервые становятся половой диморфией, чтобы изучить изменения в двух сигнальных путях, sonic hedgehog ( SHH ) и семействе сайтов интеграции MMTV бескрылого типа ( WNT ) / β-катенин.МРНК SHH и β-катенин преимущественно экспрессировались в уретральном эпителии таммарного фаллоса, как и у человеческих млекопитающих. Лечение эстрогенами и кастрация мужчин вызывали повышенную регуляцию SHH , в то время как лечение андрогенами снижало регуляцию SHH . Эти эффекты кажутся прямыми, поскольку мы обнаружили предполагаемые сайты связывания рецептора эстрогена α (ERα) и рецептора андрогена (AR) около SHH. WNT5A , как и SHH , подавлялся андрогеном, тогда как WNT4 усиливался в женских фаллосах после лечения андрогеном.После лечения эстрогеном, WIF1 и WNT7A были подавлены в мужских фаллосах. После кастрации был активирован WNT9A . Эти результаты предполагают, что пути SHH и WNT регулируются как эстрогеном, так и андрогеном, чтобы направлять пролиферацию и удлинение фаллоса во время дифференцировки. Их реакция на экзогенные гормоны делает эти гены потенциальными мишенями для EED в этиологии аномального развития фаллоса, включая гипоспадию.
Введение
Развитие фаллоса — это андроген-зависимый процесс [1–6], контролируемый множеством молекулярных сигнальных путей, включая пути sonic hedgehog (SHH) и WNT / β-catenin. Потеря Shh [7, 8] или β-катенина [9] в развивающемся фаллосе предотвращает закрытие уретры, что приводит к гипоспадии. Гипоспадия — все более частый порок развития, от которого страдает примерно 1/125 живорожденных мальчиков в развитых странах [10].Увеличение частоты на 2% каждый год не может быть объяснено мутациями и, следовательно, должно иметь экологическое происхождение [11–13]. Эндокринные разрушители окружающей среды (EED), соединения, которые препятствуют или индуцируют активность андрогенов или эстрогенов, такие как инсектициды, гербициды и многие пластмассы, вероятно, ответственны за рост числа случаев гипоспадии [14–21]. Воздействие флутамида, ингибитора рецепторов андрогенов (AR), феминизирует фаллос и вызывает гипоспадию у мышей [19, 20, 22], крыс [21, 23] и таммарного валлаби [24].У женщин маскулинизация женского фаллоса и увеличенный клитор могут быть вызваны андрогенными EED [2, 5, 25–27]. В некоторых элегантных экспериментах Zheng et al. установили важность баланса между активностью AR и рецептора эстрогена α (ERα) для поддержания нормального развития полового члена [20]. Нарушение этого баланса либо обработкой ингибиторами AR (флутамида) и ERα (фулвестрантом), либо условным нокаутом Ihh вызывает микропенис через сигнальные пути hedgehog и WNT [20].Чтобы понять онтологию развития внешних гениталий и то, как EEDs потенциально влияют на развитие фаллоса, мы изучили влияние экзогенного андрогена и эстрогена на два взаимодействующих сигнальных пути, необходимых для развития полового члена, а именно пути SHH и WNT / β-catenin.
У мышей Shh является двойным регуляторным морфогеном, контролирующим рост и формирование паттерна генитального бугорка (GT) и образование уретры [28–31]. Он экспрессируется в клоакальной мембране ранних эмбрионов до протрузии GT и, по-видимому, важен для регуляции эпителиально-мезенхимальных взаимодействий между эпителием уретральной пластинки и прилегающей мезенхимой [28-31].SHH и его эффектор GLI2 влияют на дифференцировку мезенхимы, прилегающих к эпителию уретры, и способствуют реакции на андрогены [29, 32, 33]. Оба важны для поддержания пролиферации и выживания клеток [29, 31]. Белки семейства GLI участвуют в передаче сигналов SHH [33, 34], а фактор транскрипции Zinc Finger 2 (GLI2) семейства GLI опосредует передачу сигналов hedgehog и регулирует дифференцировку мезенхимы GT. Транскрипция генов Gli регулируется двумя мембранными рецепторами, Patched 1 (PTCh2) [35, 36] и Smoothened (SMO) [35, 37].В отсутствие передачи сигналов hedgehog PTCh2 ингибирует SMO, что дополнительно инактивирует факторы транскрипции GLI [38]. Patched 2 (PTCh3), продукт Ptch3 , паралог Ptch2 , также действует, подавляя передачу сигналов SHH во время раннего эмбрионального развития [39].
Несколько генов семейства WNT, включая Wnt3, Wnt4, Wnt5a, Wnt7a и Wnt9b , экспрессируются в мезенхиме и эктодерме GT [8, 9, 30, 40]. У мышей β-катенин в основном находится в энтодермальном эпителии недифференцированного фаллоса [41].Мезенхимальный β-catenin и эктодермальный β-catenin способствуют пролиферации клеток и поддерживают клеточную целостность во время роста GT [42]. Условный нокаут β-катенина вызывает тяжелую гипоспадию [9]. Фактор 1 ингибирования WNT (WIF1), секретируемый белок, регулирует апоптоз энтодермальных клеток во время разделения клоаки путем ингибирования пути передачи сигналов WNT / β-catenin [43–45]. WIF1 ингибирует сигнальный путь WNT / β-catenin путем прямого связывания с молекулами WNT и впоследствии вызывает деградацию β-catenin [43].Нарушение регуляции Wif1 вызывает неспособность клоакальной перегородки и индуцирует гипоспадийный фенотип, сходный с наблюдаемым у мутантов Shh -null [45], указывая на то, что Wif1 также участвует в развитии фаллоса.
Во время роста GT у мышей SHH, расположенный выше передачи сигналов WNT / β-catenin, регулирует пролиферацию клеток, контролируя активность WNT / β-catenin [41]. Вырост фаллоса и уровень активности передачи сигналов WNT / β-catenin зависят от дозы сигнала SHH [41].Потеря передачи сигналов SHH останавливает пролиферацию или индуцирует апоптоз [31], снижает экспрессию гена Wnt и снижает активность передачи сигналов WNT / β-catenin [41]. Кроме того, присутствие андрогена поддерживает сигнальную активность WNT / β-катенин. Сам β-катенин может взаимодействовать с AR для усиления AR-опосредованной активации генов-мишеней, тем самым маскулинизируя мужской фаллос [9].
Хотя роль передачи сигналов SHH и WNT в развитии фаллоса хорошо изучена у мышей, обычно с помощью нокаута генов [30, 46], гормональная регуляция этих генов в фаллосе во время раннего развития плода все еще остается неясной.Недоступность зародышей мышей в утробе матери затрудняет экспериментальные гормональные манипуляции. У сумчатых половая дифференциация фаллоса происходит постнатально, поэтому эти млекопитающие предоставляют уникальную возможность для изучения гормональной регуляции экспрессии генов. Стероиды можно вводить непосредственно молодым во время длительной фазы постнатальной дифференцировки фаллоса и закрытия уретры без осложняющих эффектов материнской физиологии и плацентарного переноса. Это первое исследование, в котором стероидные гормоны вводятся непосредственно развивающимся молодым людям с индифферентными фаллосами для изучения гормональной реакции ключевых генов фаллоса.
Развитие фаллоса у таммарных валлаби сходно с развитием фаллоса у других млекопитающих, но половая дифференциация происходит после рождения. Начало роста GT очевидно за 2 дня до рождения [47], но последующая дифференцировка происходит постнатально. С рождения до 50-го дня послеродового периода (пп) не существует очевидных морфологических различий между мужским и женским фаллосом [27]. После 50-го дня расстояние между проходом уретры и анусом у самцов становится больше, чем у самок [27, 47, 48].К 60-м суткам GT самцов удлиняется и становится длиннее, чем у самок [5]. Кроме того, уретральная борозда в половом члене начинает закрываться вдоль вентральной средней линии, поэтому отверстие уретры постепенно перемещается от проксимального к дистальному [27]. К 150 дню pp закрытие уретры завершается, и проход уретры находится в головке полового члена. В женском фаллосе уретральная борозда не срастается [5, 27]. У мужчин также имеется заметный трубчатый дивертикул, консервативная структура, сливающаяся с уретрой, которая также встречается у козла [49] и пениса человека [50], но у таммарских самок он остается твердым шнуром [5, 27].
Как у мышей и людей, развитие таммарного мужского фаллоса андроген-зависимо [51]. Однако самые ранние половые диморфические различия возникают в то время, когда уровни циркулирующих андрогенов одинаковы у мужчин и женщин [5, 27], что позволяет предположить, что гормонально-ответные пути по-разному активируются до половой дифференциации фаллоса. Это гормоночувствительное окно (окно импринтинга андрогена) [5] было первым из нескольких, описанных в настоящее время у млекопитающих, и возникает между 25 и 30 днями pp [52].Гормональное воздействие на этой ранней фазе развития имеет решающее значение для инициирования более позднего отрастания фаллоса и начала закрытия уретры [5]. Изменение уровней гормонов во время этого окна чувствительности способствует гипоспадии или развитию фаллоса с измененным полом на более поздних стадиях [5, 51].
У таммаров концентрация тестостерона в семенниках увеличивается со 2-го дня до 40-го дня [53]. После 40-го дня концентрация тестостерона в яичках быстро падает до 50-го дня [53, 54].Однако основным секретируемым андрогеном в развивающихся таммарах является не тестостерон, а 5α-андростан-3α, 17β-диол (адиол), который обратимо превращается в дигидротестостерон (ДГТ) 3α-гидроксистероиддегидрогеназой в органах-мишенях по альтернативному пути синтеза стероидов. [55]. Имеются убедительные доказательства того, что адиол ответственен за вирилизацию таммарного мочеполового тракта, продуцируемую альтернативным путем [55, 56], и что уровни адиола сексуально диморфны во время вирилизации урогенитального синуса [56].
мРНК SHH имеет такую же локализацию в раннем пренатальном таммарном фаллосе, что и у мышей [29, 30]. Выявляется в эпителии пластинки уретры за 1 день до рождения [52]. Экспрессия SHH остается низкой во время андроген-зависимого окна чувствительности (25-30 п.н.), которое мы определили, когда происходит импринтинг андрогенов, но повышается примерно на 50-й день, когда тестикулярные андрогены уменьшаются и начинается половая дифференцировка фаллоса [52] . У молодых самцов, кастрированных на 29 п.п., SHH временно повышается на 50 п.п. и, аналогично, он снижается в женских фаллосах после лечения мощным андроген-адиолом [52].
Сумчатые были отделены от млекопитающих более 160 миллионов лет [57]. Механизмы, которые сохраняются между сумчатыми и эвтерианами, вероятно, являются ключевыми компонентами развития фаллоса. В этом исследовании мы исследовали влияние гормональных манипуляций на морфологию и экспрессию генов в развивающемся фаллосе таммарного молодняка. Мы лечили женщин адиолом (основным циркулирующим андрогеном при закрытии уретры), а мужчин — эстрогеном или кастрацией во время окна импринтинга андрогенов.Мы выполнили RNA-Seq и гибридизацию in situ для определения изменений транскрипции и локализации мРНК ключевых генов в сигнальных путях SHH и WNT / β-catenin.
Материалы и методы
Животные
валлаби таммар ( Macropus eugenii ) происхождения с острова Кенгуру (Южная Австралия) содержались в нашей гнездовой колонии в Мельбурне. Самок наблюдали ежедневно от 25 до 30 дней после извлечения детенышей из мешочка для проверки новорожденных детенышей [58].Пол детенышей определяли по наличию зачатков мошонки или молочных желез [59]. Для молодых людей с неизвестным днем рождения возраст оценивался по измерениям длины головы и веса по опубликованным кривым роста [60]. Все ткани были собраны в соответствии с соответствующими разрешениями, а эксперименты одобрены комитетами по этике экспериментов на животных Мельбурнского университета в соответствии с рекомендациями Национального совета по здравоохранению и медицинским исследованиям Австралии (2013 и 2014).
Фаллосы измеряли штангенциркулем на 150 пп.Фаллосы вывертывались осторожным нажатием на сторону GT и измерялись от кончика головки до нижней части стержня, как описано ранее [5]. Молодняк из мешочков, обработанных стероидами, умерщвляли на 50 п.п. дня или 150 п.п. для морфологического исследования и анализа экспрессии генов. Фаллосы либо мгновенно замораживали, либо хранили при -80 ° C для более поздних РНК-Seq (n = 5), либо фиксировали в 4% параформальдегиде, заливали парафином и разрезали на 5-мкм срезы для гистологического анализа (n = 3). и разрезать на срезы 7 мкм для сечения гибридизации in situ (n = 3).Образцы фаллоса для морфологического анализа (n = 3) отбирали у самцов и самок на 20-й день и 90-й день. Молодых самцов с кошачьей сумкой (n = 5) кастрировали на 25-й день после пикапа, как описано ранее [61–63], или перорально вводили эстроген каждый день между 20-м и 40-м днем (n = 5), как описано ранее [51, 64]. . Молодым самкам (n = 5) вводили андростандиол (адиол) с 20-го дня до 40-го дня, как описано ранее [52]. Фаллосы собирали на 50-й день (n = 5) для RNA-Seq (см. Ниже).Вторую обработанную группу (n = 5) оставили до 150 п.п. до дня забора тканей. Семенники из полученного эстрогеном мужского мешочка молодых (n = 5) и нормальных самцов из сумки молодых (n = 5) собирали для измерения концентрации тестостерона в яичках и сравнения морфологии.
Экстракция РНК и получение кДНК
Перед извлечением РНК из образцов замороженные образцы гомогенизировали в реагенте TRIzol (каталожный № 15596026, ThermoFisher Scientific). РНК выделяли из развивающихся фаллосов с помощью набора RNeasy Mini (Cat # 74804, QIAgen) в соответствии с рекомендациями производителя.РНК рассчитывали с помощью флуорометра Qubit 2.0 (ThermoFisher Scientific). Все образцы РНК обрабатывали ферментом, не содержащим ДНК (каталожный № 1906, Ambion), для предотвращения загрязнения геномной ДНК. Тотальную РНК (200 нг) использовали для синтеза матрицы кДНК с использованием набора для синтеза кДНК с высокой точностью транскрипции (№ по каталогу 05081963001, Invitrogen).
Секция мРНК гибридизация in situ
Фаллосы, залитые парафином, были разрезаны на 7 мкм. Праймеры для зондов были созданы с помощью онлайн-программы Primer 3 (http: // primer3.ut.ee/) ( SHH : вперед: 5′-AGCAGTTTATCCCCAACGTG-3 ′, обратный: 5′-GCGGTTGATGAGAATGGTG-3 ′). Дигоксин использовали для мечения зондов (каталожный номер 11277073910, Roche). Промоторы T7 или SP6 использовали для транскрипции смысловых или антисмысловых зондов in vitro с помощью полимеразы T7 / SP6 (каталожный номер P1460, Promega). Предварительную гибридизацию проводили в течение 2 ч при 42 ° C, а гибридизацию — в течение 16–18 ч при 42 ° C. Затем предметные стекла промывали и инкубировали с анти-дигоксигенином-AP (разведение 1: 300) (№ по каталогу 110
910, Roche) в течение 16–18 ч при 4 ° C.NBT / BCIP (разведение 1:50) использовали для проявления окраски (№ по каталогу 11681451001, Roche). Для подсчета окрашивания использовали раствор Nuclear Fast Red (№ по каталогу N3020, Sigma Aldrich). Отрицательные контроли (сенсорный зонд) были сделаны для каждого возраста и пола для гибридизации in situ (дополнительный рисунок 1 и данные не показаны).
Рисунок 1.
Морфология нормальных фаллосов на 150 п.о. дня (n = 3) и фаллосов, обработанных гормонами на 150 п.п. (n = 3). Длины фаллосов между группами (n = 3–5) сравнивали с тестом Стьюдента t .A: лечение адиолом, CC: кавернозное тело, D: дивертикул, E2: лечение эстрогенами, U: уретра. * P <0,05, ** P <0,01, *** P <0,001. Масштабная линейка 500 мкм. Панель ошибок: SEM.
Рисунок 1.
Морфология нормальных фаллосов на 150 п.о. (n = 3) и обработанных гормонами фаллосов на 150 п.п. (n = 3). Длины фаллосов между группами (n = 3–5) сравнивали с тестом Стьюдента t . A: лечение адиолом, CC: кавернозное тело, D: дивертикул, E2: лечение эстрогенами, U: уретра.* P <0,05, ** P <0,01, *** P <0,001. Масштабная линейка 500 мкм. Панель ошибок: SEM.
Иммунофлуоресценция
Фаллосы, залитые парафином, были разрезаны на 5 мкм. Срезы депарафинизировали гистоленом и регидратировали с помощью серии разбавлений этанолом от 100% до воды. Извлечение антигена проводили путем нагревания при 30% мощности микроволн (1000 Вт) в течение 20 мин. Первичное антитело, антитело к β-катенину (кролик, каталожный номер sc-7199, Санта-Крус, Австралия, разведение 1: 100) или антитело SHH (кролик, каталожный номер ab19897, Abcam, США, разведение 1: 100) (дополнительная таблица S1) добавляли на срез и инкубировали при 4 ° C в течение ночи.Вторичное антитело (ослиное антитело Alexa Fluor 555 против кролика, 1: 1000, 2 мг / мл, № по каталогу A-31572, Invitrogen) получали при комнатной температуре в течение 1 часа. DAPI (1: 10 000 5 мг / мл, № по каталогу D3571, Invitrogen) использовали в качестве контрастного красителя для ядер. Для монтажа использовалась флуоресцентная монтажная среда (№ по каталогу S302380, Dako). Края покровных стекол закрывали лаком для ногтей и хранили при 4 ° C в темноте. Отрицательные контроли, которые инкубировали с фракцией кроличьего иммуноглобулина IgG (№ по каталогу X0936, Dako), были приготовлены для каждого возраста и пола для срезов антител (дополнительные рисунки S1 и S2).
Экстракция стероидов и конкурентный ИФА
Семенники гомогенизировали в 400 мкл ацетонитрила (№ по каталогу 270717, Sigma). Супернатант отделяли центрифугированием при 10 160 × g в течение 10 мин при 4 ° C. В супернатант добавляли восемьсот микролитров гексана (№ по каталогу 24577.323, BDH Prolabo) и энергично перемешивали в течение 5 мин. Верхний слой переносили в другую чистую пробирку, и жидкость упаривали с помощью концентратора SpeedVac (Quantum Scientific, Biostrategy, США).Высушенный образец растворяли в 100 мкл этанола, а затем в 900 мкл 1% бычьего сывороточного альбумина в фосфатно-солевом буфере (PBS, 137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na 2 HPO 4 , 1,8 мМ KH 2 PO 4 ).
Мини-набор тестостерона ISWE (каталожный номер ISWE001, Arbor Assays, США) был использован для измерения концентрации тестостерона в нормальных яичках 50 п.п. (n = 5) и обработанных эстрогеном 50 п.п. таммар. Плоскодонные 96-луночные планшеты Nunc MaxiSorp (№ по каталогу 44–2404-21 Thermo-Fisher) покрывали козьими антителами против кроличьего IgG (2 мкг / мл, № по каталогу E0432, Dako) при 4 ° C в течение ночи.В каждую лунку добавляли сто микролитров антитела тестостерона ISWE 1: 100 и инкубировали при комнатной температуре в темноте в течение ночи. Планшеты блокировали на 30 мин при комнатной температуре 3% BSA в PBS с 0,05% Tween. После блокирования в планшеты добавляли 50 мкл разбавленных образцов, контролей и стандартов и в планшеты добавляли 100 мкл тестостерона, конъюгированного с ISWE HRP. Планшеты инкубировали при комнатной температуре в течение 3 ч в темноте. Цветную реакцию проводили, добавляя 100 мкл раствора TMB (№ по каталогу 002023, Invitrogen) в течение 30 мин.Для остановки реакции добавляли сто микролитров 1 н. Соляной кислоты. Измерение проводили с помощью считывающего устройства для микропланшетов при длине переплетения 620 нм. Все этапы выполнялись на шейкере, за исключением покрытия и блокировки пластины. Планшеты промывали трижды PBST после каждого шага, за исключением блокировки. Для расчета мы использовали подгонку логистической кривой с пятью параметрами, восстановление спайка составляет 95,90% ± 2,43%, степень согласия> 0,999. Данные выражены в нг тестостерона на мг яичка.
Прогноз промоутера
AR- и ERα-связывающих сайтов в промоторной области SHH у мыши были обнаружены с использованием базы данных по регуляции транскрипции генов (GTRD) [65]. Последовательности вверх и вниз по течению SHH в таммаре были извлечены из генома таммар валлаби 3.0 (Паск, А.Дж. Ренфри, личное сообщение М.Б.). Мы использовали рецепторный элемент андрогена мыши (ARE) и элемент рецептора эстрогена (ERE) для прогнозирования сайтов связывания AR и ERα в близлежащих областях tammar SHH .Прогнозирование проводилось программой онлайн-сканирования JASPAR [66].
Сборка и анализ RNA-Seq
Образцы
РНК были дополнительно протестированы с помощью TapeStation (Agilent). Соответствующие количества РНК подвергали мультиплексному анализу индексированной РНК-Seq с использованием набора TruSeq (Illumina) и анализатора HiSeq2500 (Illumina). Приблизительно 10–14 × 10 6 прочтений (100 п.н.) было получено из каждого образца после Q.V. > 30 фильтров. Средний балл по качеству составил ∼39. Данные RNA-Seq были собраны с помощью конвейеров Trimmomatic [67] и Cufflinks-Cuffdiff [68] и проанализированы с помощью пакета CummeRbund R [69].Средняя скорость картирования для всех образцов была выше 80%, а качество было хорошим для анализа дифференциальной экспрессии. Геном таммар валлаби 3.0 (Паск, А.Дж. Ренфри, М.Б. личное сообщение) использовался в качестве руководства для сборки транскриптома. Аннотация была получена путем взлома базы данных белков UNIPROT [70]. Дифференциально экспрессируемые гены были извлечены с помощью пакета CummeRbund R и базовых сценариев R [71]. Гены, представляющие интерес в каждом сигнальном пути, были отобраны путем сравнения наших дифференциально экспрессируемых генов из RNA-Seq с генами из базы данных GeneCards (http: // www.genecards.org/). FPKM (количество фрагментов на килобазу модели экзона на миллион отображенных считываний) — это нормализованная оценка экспрессии гена на основе данных RNA-Seq. FPKM рассчитываются по количеству считываний, картированных с каждой конкретной последовательностью гена, с учетом длины гена и глубины секвенирования [72].
Статистика
Данные были проанализированы с использованием R версии 3.12 [71] и пакета nplr (для ELISA) [73]. Длину фаллоса (n = 3-5 повторов) между разными группами лечения и контрольными группами сравнивали с помощью критерия Стьюдента t .Пятипараметрическая логистическая регрессия использовалась для подбора стандартной кривой для результатов ELISA. Тест Стьюдента t использовали для сравнения значимости различий в концентрации тестостерона в яичках между группами и для расчета значимости различий в экспрессии генов между группами. Все планки ошибок представляют стандартную ошибку среднего (SEM)
Результаты
Закрытие уретры, регулируемое андрогенами и эстрогенами, и рост фаллоса в таммаре
У мужчин на 150-й день уретры уретра закрыта, и дивертикул имеет просвет, но у женщин на 150-й день уретра остается открытой, а дивертикул представляет собой сплошной шнур из эпителиальных клеток (рис. 1).Мужские фаллосы были значительно длиннее, чем женские (рис. 1). Обработка эстрогеном во время окна импринтинга андрогенов (день 20-40 п.н.) значительно ингибировала удлинение фаллоса у мужчин (рис. 1), но не подавляла закрытие уретры (рис. 1). Напротив, кратковременное воздействие андрогенов во время окна импринтинга андрогенов у женщин не вызывает удлинения фаллоса (рис. 1), но вызывает закрытие уретры (рис. 1). Мы также сравнили концентрацию тестостерона в яичках и морфологию яичек между нормальными семенниками 50 п.п. и обработанными эстрогеном 50 п.п. яичек.Как и ожидалось, наблюдалась вариабельность концентраций [74, 75], но не было ни значительных различий в концентрации тестостерона в яичках, ни каких-либо морфологических различий между этими двумя группами на 50-й день, через 10 дней после прекращения лечения эстрогенами (рис. 2).
Рисунок 2.
Концентрация тестостерона в яичках (n = 5) и морфология семенников (n = 5) в контрольных и эстроген-обработанных семенниках на 50-й день. E2: лечение эстрогеном, GC: зародышевые клетки, SC: клетки Сертоли, ST: семенные канальцы.Шкала 100 мкм. Панель ошибок: SEM.
Рисунок 2.
Концентрация тестостерона в яичках (n = 5) и морфология семенников (n = 5) в контрольных и эстроген-обработанных семенниках на 50 дней. E2: лечение эстрогеном, GC: зародышевые клетки, SC: клетки Сертоли, ST: семенные канальцы. Шкала 100 мкм. Панель ошибок: SEM.
Экспрессия гена β-катенина не зависела от андрогенов и локализовалась в эпителии уретры
Не было сексуально диморфной экспрессии мРНК β-катенина между нормальными мужскими фаллосами и нормальными женскими фаллосами на 50-й день (фиг. 3).На экспрессию β-катенина не влияло лечение адиолом, лечение эстрогеном или кастрация. Белковая локализация β-катенина преимущественно обнаруживалась в клеточных мембранах, но не в ядре внешнего эпителия и эпителия уретры с 20-го дня до 90-го дня у обоих полов (дополнительный рисунок S3). β-катенин слабо обнаруживался в кавернозных телах (дополнительный рисунок S3).
Рисунок 3.
Экспрессия WIF1, WNT4, WNT11, WNT5A, WNT7A, WNT9A и β-катенина в контроле (n = 5) и обработанных фаллосах (n = 5) на 50 п.о.A: обработка адиолом, E2: обработка эстрогеном, FPKM: количество фрагментов на килобазу транскрипта на миллион отображенных считываний. * P <0,05 ** P <0,01. Панель ошибок: SEM.
Рисунок 3.
Экспрессия WIF1, WNT4, WNT11, WNT5A, WNT7A, WNT9A и β-катенина в контроле (n = 5) и обработанных фаллосах (n = 5) на 50 п.о. : лечение адиолом, E2: лечение эстрогеном, FPKM: количество фрагментов на килобазу транскрипта на миллион отображенных считываний. * P <0.05 ** P <0,01. Панель ошибок: SEM.
Гены WIF1 и WNT гормонально реагировали во время дифференцировки фаллоса
В отличие от β-катенина, другие гены пути WNT были затронуты лечением андрогенами или эстрогенами. Экспрессия WIF1 была значительно выше у самцов, чем у самок, на 50-й день (фиг. 3). Обработка эстрогеном снижала экспрессию WIF1 до уровня, аналогичного тому, который наблюдается в женских фаллосах (рис. 3).Обработка адиолом не влияла на экспрессию WIF1 (фиг. 3). Экспрессия WNT4 и WNT Family Member 11 ( WNT11) была значительно выше у нормальных самцов, чем у нормальных самок на 50-й день (фиг. 3). Интересно, что на WNT11 не влияли лечение адиолом, эстрогенами или кастрация. Обработка адиолом у женщин повышала экспрессию WNT4 до уровня, аналогичного тому, который наблюдается в мужских фаллосах (рис. 3).Экспрессия WNT Family Member 5A ( WNT5A) была значительно выше у нормальных женщин, чем у мужчин (рис. 3). Обработка адиолом у женщин снижала экспрессию WNT5A до уровня, аналогичного тому, который наблюдается в мужских фаллосах (рис. 3). После кастрации экспрессия WNT5A увеличилась до аналогичного уровня, наблюдаемого в женских фаллосах (рис. 3). Экспрессия WNT Family Member 7A ( WNT7A) и WNT Family Member ( WNT9A) не различалась между полами (Рисунок 3).Однако экспрессия WNT7A подавлялась, когда самцов лечили эстрогеном, тогда как WNT9A подавлялась после кастрации (рис. 3).
Консервативное распределение
SHH в эпителии уретры таммара Белок
SHH был обнаружен в основном в эпителии уретры фаллосов плода мужского и женского пола на 25-й день pp (рис. 4). Как мРНК SHH, так и белок также были обнаружены в коже и эпителии уретры в фаллосах мужчин и женщин на 20 п.н. (рис. 4).Распределение мРНК SHH в фаллосах на 90 п.п. аналогично распределению мРНК в фаллосах 20 п.п. (рис. 4). Интересно, что мРНК SHH также была обнаружена в мезенхимальных клетках, окружающих дивертикул и кавернозные тела, и сохранялась в дивертикуле и эпителии уретры после закрытия уретры у мужчин на 90-е сутки и в открытой уретре у женщин (Рисунок 4).
Рисунок 4.
Распределение SHH как в мужских (n = 3), так и в женских фаллосах (n = 3) на 20-й день и 90-й день.D: дивертикул, E: эпителий, M: мезенхима, U: уретра, UE: эпителий уретры. Масштабная линейка 500 мкм.
Рисунок 4.
Распределение SHH как в мужских (n = 3), так и в женских фаллосах (n = 3) на 20-й день и 90-й день. D: дивертикул, E: эпителий, M: мезенхима, U : уретра, УЭ: эпителий уретры. Масштабная линейка 500 мкм.
SHH и PTCh3 гормонально реагировали во время дифференцировки фаллоса
Паттерны экспрессии SHH и PTCh3 были изменены на 50-й день, когда фаллосы начали быть сексуально диморфными, хотя другие нижестоящие члены этого пути изменились мало (см. Ниже). SHH мРНК была значительно выше в женских фаллосах, чем у мужчин. Обработка эстрогеном мужчин и кастрация усиливали экспрессию SHH в мужских фаллосах, тогда как обработка адиолом подавляла экспрессию SHH в женских фаллосах (фиг. 5).
Рисунок 5.
Экспрессия SHH, GLI1, GLI2, PTCh2, PTCh3 и SMO в нормальных (n = 5) и обработанных фаллосах (n = 5) на 50-й день с. A: обработка адиолом, E2 : лечение эстрогеном, FPKM: количество фрагментов на килобаз транскрипта на миллион отображенных считываний.* P <0,05 ** P <0,01. Панель ошибок: SEM.
Рисунок 5.
Экспрессия SHH, GLI1, GLI2, PTCh2, PTCh3 и SMO в нормальных (n = 5) и обработанных фаллосах (n = 5) на 50-й день с. A: лечение адиолом, E2: лечение эстрогеном, FPKM: количество фрагментов на килобазу транскрипта на миллион отображенных считываний. * P <0,05 ** P <0,01. Панель ошибок: SEM.
Последовательности белков AR и ERα высоко консервативны между мышами и таммаром (дополнительный рисунок S4).ДНК-связывающий домен AR и ERα между двумя видами имеет 100% идентичность в аминокислотных последовательностях (дополнительный рисунок S4). Следовательно, можно предсказать сайты связывания AR и ERα в промоторе SHH в таммаре с мышами ARE и ERE. У мышей в промоторе Shh были идентифицированы множественные сайты связывания AR и ERα, и, аналогичным образом, множественные предсказанные сайты связывания AR и ERα были обнаружены в промоторе SHH в таммаре (рис. 5).
Экспрессия PTCh3 была значительно выше в фаллосах самок 50 п.п. и фаллосах кастрированных самцов на 50 п.п. по сравнению с фаллосами самцов 50 п.н. Однако не было резких изменений в экспрессии нижестоящих генов-мишеней ( GLI1, GLI2, GLI3, PTCh2 и SMO ) после лечения стероидными гормонами (рис. 5).
Обсуждение
В этом исследовании изучалась гормональная регуляция сигнальных путей SHH и WNT / β-catenin во время развития таммарного фаллоса.После лечения женщин андрогенами или мужчин эстрогенами во время окна импринтинга андрогенов или удаления андрогенов у мужчин посредством кастрации наблюдалась задержка роста фаллоса и нарушение закрытия уретры, что приводило к гипоспадии. Это сопровождалось значительными изменениями экспрессии генов SHH и WNT . Повышающая и понижающая регуляция генов этих двух путей после лечения андрогенами и эстрогенами предполагает, что они являются мишенями стероидных гормонов, влияющих на нормальную дифференцировку фаллоса.Присутствие консервативных сайтов связывания AR и ER в промоторных областях этих генов у сумчатых, мышей и людей предполагает, что гормональная регуляция является консервативной особенностью этих критических генов развития фаллоса.
Консервативная локализация мРНК
SHH и белка β-катенина в таммарном фаллосе
SHH мРНК была в основном локализована в эпителии уретры, особенно в фаллосах плода, подобно мышам [29, 30], крысам [76], змеям [77], аллигаторам [78] и черепахам [79].На более поздних стадиях развития он также был обнаружен в кожном эпителии диафиза полового члена таммара. Поскольку дифференциация таммарного фаллоса происходит после рождения, в отличие от здоровых млекопитающих, эпидермис фаллоса намного толще, и эпителиальные клетки быстро размножаются, заменяя кератиновый слой для защиты, пока детеныш находится в сумке. Это постнатальное развитие таммарного фаллоса может быть причиной разницы в локализации SHH в таммарном фаллосе по сравнению с другими млекопитающими, которые были исследованы только на эмбриональной и неонатальной стадиях.Интересно, что мРНК SHH также была обнаружена в кавернозных телах и в мезенхиме вокруг дивертикула во время его формирования, указывая тем самым, что SHH может участвовать в нескольких аспектах развития фаллоса таммара.
Хотя на экспрессию β-катенина не оказало явного влияния какое-либо лечение, крайне важно поддерживать целостность эпителиальных клеток фаллоса во время его дифференциального роста и развития у самцов и самок соответственно, как у мышей [42, 80] и у человека [81].В таммаре β-catenin был широко обнаружен в мембранах эпителиальных клеток фаллоса и эпителия уретры, что позволяет предположить, что он играет такую же консервативную роль в таммаре, что и у мышей [42]. Интересно, что мы не наблюдали какого-либо окрашивания β-catenin в ядре, предполагая, что канонический путь может не играть критической роли на стадиях, которые мы наблюдали в таммаре.
Присутствие андрогена в окне импринтинга инициировало закрытие уретры у женщин, но не удлинение фаллоса
Обработка самок адиолом с 20-го по 40-й день в течение окна импринтинга вызвала полное закрытие уретры на 150-й день, но длина фаллоса не увеличивалась (рис. 6).В отличие от закрытия уретры, рост фаллоса требует длительного воздействия андрогенов [5]. У молодых самок, которым пересажены неонатальные семенники вскоре после рождения, наблюдается рост фаллоса по мужскому типу и закрытие уретры при обследовании на 300-й день [51], а в 3 года — мужские фаллосы нормального размера [61]. Напротив, кастрация самцов до 25 п.н. приводит к высокофеминизированным фаллосам с открытой уретрой [5, 51]. Очевидно, что андроген является доминирующим стероидом, контролирующим закрытие уретры и удлинение фаллоса во время развития таммарного фаллоса.Концентрация андрогенов в яичках снижается к дню на 40–45 п.п. у нормальных мужчин [53], и нет значительной разницы в концентрациях тестостерона в плазме между мужчинами и женщинами в любое время вплоть до 70-80 п.п. [53, 54], когда мужские фаллосы начинают удлиняться быстрее, чем женские фаллосы [5, 54]. Однако плазменный адиол сексуально диморфен между 20 и 40 днями, когда начинается развитие простаты [56], и является гормоном, ответственным за импринтинг андрогенов в это время [5]. На молекулярном уровне возможно, что закрытие уретры и удлинение фаллоса контролируются двумя разными наборами генов, которые имеют разную андрогенную чувствительность.
Рисунок 6.
Краткое описание эффектов лечения адиолом, эстрогеном и кастрацией на морфологию фаллоса и экспрессию генов. A: анус, E2: лечение эстрогенами, ES: эпителиальный шов, GT: генитальный бугорок, U: уретра, UE: эпителий уретры, UM: уретральный проход, TS: поперечный разрез; проксимальный у основания фаллоса. Зеленая стрелка: усиление регуляции. Красная стрелка: подавление.
Рисунок 6.
Краткое изложение эффектов лечения адиолом, эстрогеном и кастрацией на морфологию фаллоса и экспрессию генов.A: анус, E2: лечение эстрогенами, ES: эпителиальный шов, GT: генитальный бугорок, U: уретра, UE: эпителий уретры, UM: уретральный проход, TS: поперечный разрез; проксимальный у основания фаллоса. Зеленая стрелка: усиление регуляции. Красная стрелка: подавление.
Андроген контролирует развитие фаллоса и регулирует
SHH и несколько генов WNT
В таммаре SHH остается низким у мужчин до 50 п.п., когда тестикулярный андроген высокий, но временно увеличивается между 50 и 60 п.п. [52], когда уровень тестостерона в яичках низкий [53], прежде чем снова падает.Удаляя семенники или давая андроген самкам, мы наблюдали увеличение или уменьшение экспрессии SHH , соответственно (рис. 5), что дополнительно подтверждает предположение о том, что SHH негативно регулируется андрогеном. Аналогичным образом, экспрессия SHH также снижается в линиях клеток предстательной железы человека после культивирования с DHT, и эффект отменяется в присутствии бикалутамида, антиандрогена [82]. PTCh3 транскриптов, как и SHH , были выше у женщин на 50-й день и увеличились после кастрации (рис. 5). Уровни SHH резко, но временно повышаются в таммарных мужских фаллосах на 50-й день, когда фаллос впервые становится сексуально диморфным [52], так что это может быть переключатель, который контролирует время роста фаллоса и закрытия уретры. Напротив, нижние гены SHH ( GLI1, GLI2, GLI3, PCTh2 и SMO ) не подвергались значительному влиянию гормонального лечения. У мышей транскрипция Ptch2 и Gli1 запускается только после активации сигнального пути SHH [83, 84], поэтому мы могли пропустить транскрипционные изменения, которые происходят после 50 п.н.
WNT5A показал паттерн экспрессии, аналогичный паттерну экспрессии SHH. WNT5A также ингибировался обработкой андрогенами, но увеличивался после удаления семенников (рис. 6). Может быть взаимодействие между андрогеном, SHH , и WNT5A , управляющее маскулинизацией фаллоса, как это происходит у мышей с нокаутом [9, 41]. WIF1 подавляет действие белков WNT, и у мышей характер экспрессии каждого гена Wnt во время развития фаллоса различен [41, 45].Следовательно, возможно, что разные Wnts реагируют на андрогены иначе, чем гены hedgehog. Точно так же в таммаре WNT4 был активирован обработкой андрогенов у женщин, тогда как WNT9A был активирован только после кастрации (рис. 6). WNTA11 был более высоко экспрессирован в нормальных мужских фаллосах, чем в женских фаллосах на 50-й день, но на экспрессию не влияла обработка андрогенов у женщин или кастрация у мужчин. Эти результаты предполагают, что андрогены контролируют пролиферацию и выживаемость клеток посредством регулирования путей передачи сигналов SHH и WNT во время развития таммарного фаллоса, как это происходит у мышей [9, 41].
Эстроген подавляет удлинение фаллоса, но не синтез андрогенов
У мужчин, получавших эстроген во время окна импринтинга, пенисы были меньше, чем у контрольных мужчин. Поскольку для поддержания удлинения фаллоса таммара требуется постоянный андроген [5], возможно, что лечение эстрогенами у мужчин нарушило нормальный синтез андрогенов, тем самым подавляя удлинение фаллоса. Подобные результаты наблюдались также у людей [85], крыс [86] и мышей [87]. Однако воздействие эстрогена на самцов таммаров между 20 и 40 днями pp не повлияло на синтез тестикулярного тестостерона на 50 pp, а также не изменило морфологию яичек.Однако, помимо экзогенного эстрогена, у этих мужчин были неповрежденные семенники, поэтому и эстроген, и андроген присутствовали в обработанном эстрогеном мужском фаллосе. Это вызвало неоднозначный фенотип гениталий микропениса с полностью закрытой уретрой. Таким образом, могут быть прямые эффекты эстрогена, отличные от эффектов андрогенов на развитие фаллоса.
Влияние эстрогенов на сигнальные пути SHH и WNT
Лечение
эстрогеном у мужчин усиливало экспрессию SHH , но подавляло экспрессию WNT7A и WIF1 , что отличается от действия андрогена на эти гены.Эстроген не влиял на WNT4, WNT5A или WNT9A . В клетках рака желудка человека, не продуцирующей андрогены клеточной линии, эстроген усиливал экспрессию SHH и индуцировал клеточную пролиферацию в ERα-положительных клетках. Обработка фулвестрантом (ICI 182,780), ингибитором ERα, предотвращала индуцированную эстрогеном экспрессию SHH , предполагая, что эстроген увеличивает экспрессию SHH за счет активации передачи сигналов ERα [88]. Похожая картина наблюдалась также в клеточных линиях рака груди человека [89].Мы обнаружили ERα-связывающие сайты возле промотора SHH в таммаре, используя базу данных GTRD [65]. ERα является одним из регуляторов активации hedgehog при раке груди и желудка [88, 89]. Следовательно, вполне вероятно, что эстроген оказывает консервативное и прямое влияние на экспрессию SHH, WIF1 и WNT7A . Эти данные также показывают, что окно импринтинга андрогенов — это период, когда гены фаллоса чувствительны не только к андрогенам, но и к эстрогенам.
Выводы
Это исследование подтверждает, что андроген может вызывать закрытие уретры по мужскому типу у женщин, в то время как эстроген может подавлять удлинение фаллоса у мужчин, предполагая, что каждое морфологическое изменение чувствительно к разному набору генов.Эстроген не влиял на закрытие уретры, когда семенники оставались in situ у обработанных мужчин. Очевидно, что существует устойчивое взаимодействие как андрогенных, так и эстрогенных стероидных гормонов с генами в сигнальных путях SHH и WNT, которые контролируют закрытие уретры и рост фаллоса, демонстрируя, что эти гены являются потенциальными мишенями для возрастающих уровней EED в окружающей среде.
Дополнительные данные
Дополнительный рисунок S1 . Репрезентативные отрицательные контроли на 90 п.н. в мужском и женском фаллосе.(A) Отрицательный контроль для гибридизации in situ в мужском фаллосе на 90-й день п.п. (B) Отрицательный контроль для гибридизации in situ в женском фаллосе на 90-й день. (C) Отрицательный контроль для иммунофлуоресценции в мужском фаллосе на 90-й день стр. (D) Отрицательный контроль иммунофлуоресценции в женском фаллосе на 90-е сутки. D: дивертикул, E: эпителий, M: мезенхима, U: уретра, UE: эпителий уретры. Масштабная линейка 500 мкм.
Дополнительный рисунок S2 . Распределение белков SHH в мужских и женских фаллосах на 50 п.о. (вверху) и в соответствующих отрицательных контролях (внизу).E: эпителий, M: мезенхима, UE: эпителий уретры. Масштабная линейка: 500 мкм.
Дополнительный рисунок S3 . Распределение белков эпителиального маркера β-катенина на 20-й и 90-й дни фаллосов мужчин и женщин. A: анус, D: дивертикул, E: эпителий, M: мезенхима, U: уретра, UE: эпителий уретры. Dis: дистальный, Mid: средний, Pro: проксимальный. Синий: DAPI, Красный: β-катенин. Шкала шкалы: 1,0 мм.
Дополнительный рисунок S4 . Вверху: выравнивание белковых последовательностей AR и ESR1 между мышами и таммаром.В центре: мышь ARE и ERE и таммар предсказали ARE и ERE. Внизу: местоположения ARE (красный) и ERE (синий) мыши в геномной области SHH и предсказанные таммарные сайты связывания ARE и ERE на основе мышей.
Дополнительная таблица S1 . Список используемых антител.
Благодарности
Мы благодарим членов исследовательской группы Валлаби (Walgroup) за помощь при обращении с животными. Мы также благодарим членов Advanced Genomics Center, NIG, за техническую поддержку по секвенированию РНК.
Банкноты
Под редакцией доктора Мириам Хембергер, доктора философии, Институт Бабрахама
Список литературы
1.
Husmann
DA
,
Cain
MP
.
Микрофаллос: возможный размер фаллоса зависит от времени введения андрогена
.
Дж Урол
1994
;
152
(
2
):
734
—
739
. 2.
Валлийский
M
,
MacLeod
DJ
,
Walker
M
,
Smith
LB
,
Sharpe
RM
.
Критические андроген-чувствительные периоды развития полового члена и клитора крысы
.
Int J Androl
2010
;
33
:
e144
—
e152
.3.
Drea
CM
,
Weldele
ML
,
Forger
NG
,
Coscia
EM
,
Франк
LG
,
Licht
P
,9 94 Glickman SE
Андрогены и маскулинизация гениталий у пятнистой гиены (Crocuta crocuta).2. Эффекты пренатальных антиандрогенов
.
Репродукция
1998
;
113
(
1
):
117
—
127
. 4.
Baskin
LS
,
Sutherland
RS
,
DiSandro
MJ
,
Hayward
SW
,
Lipschutz
J
,
Cunha
GR
.
Влияние тестостерона на рецепторы андрогенов и рост полового члена человека
.
Дж Урол
1997
;
158
(
3
):
1113
—
1118
. 5.
Leihy
МВт
,
Shaw
G
,
Wilson
JD
,
Renfree
MB
.
Развитие полового члена начинается в сумке таммар-валлаби в молодом возрасте в период, когда 5α-андростан-3α, 17β-диол секретируется семенниками
.
Эндокринология
2004
;
145
(
7
):
3346
—
3352
.6.
Wilson
JD
,
George
FW
,
Renfree
MB
.
Эндокринная роль в половой дифференциации млекопитающих
.
Недавнее исследование Prog Horm Res
1994
;
50
:
349
—
364
.7.
Линь
C
,
Инь
Y
,
Veith
GM
,
Fisher
AV
,
Long
F
,
Ma
L
.
Временное и пространственное рассечение передачи сигналов Shh в развитии генитального бугорка
.
Девелопмент
2009
;
136
(
23
):
3959
—
3967
. 8.
Seifert
AW
,
Yamaguchi
T
,
Cohn
MJ
.
Функциональный и филогенетический анализ показывает, что Fgf8 является маркером генитальной индукции у млекопитающих, но не требуется для развития наружных половых органов
.
Девелопмент
2009
;
136
(
15
):
2643
—
2651
. 9.
Miyagawa
S
,
Satoh
Y
,
Haraguchi
R
,
Suzuki
K
,
Iguchi
T
,
Taketo
MM
,
Taketo
MM
,
Taketo
MM
,
Мацумото
T
,
Ki
Takeyama
,
Kato
S
.
Генетические взаимодействия путей андрогенов и Wnt / β-катенинов для маскулинизации наружных гениталий
.
Мол эндокринол
2009
;
23
(
6
):
871
—
880
. 10.
Sweet
RZ
,
Schrott
HG
,
Kurland
R
,
Culp
OS
.
Исследование частоты гипоспадии в Рочестере, Миннесота, 1940–1970, и сравнение возможных этиологических факторов случай-контроль.
.
Mayo Clin Proc
1974
;
49
(
1
):
52
—
58
. 11.
Паулоцци
LJ
.
Международные тенденции развития гипоспадии и крипторхизма
.
Environ Health Perspect
1999
;
107
:
297
—
302
.12.
Паулоцци
LJ
,
Эриксон
JD
,
Джексон
RJ
.
Тенденции гипоспадии в двух системах эпиднадзора в США
.
Педиатрия
1997
;
100
:
831
—
834
. 13.
Нассар
Н
,
Бауэр
С
,
Баркер
А
.
Рост распространенности гипоспадии в Западной Австралии, 1980–2000 гг.
.
Arch Dis Child
2007
;
92
:
580
—
584
. 14.
Vilela
MLB
,
Willingham
E
,
Buckley
J
,
Liu
BC
,
Agras
K
,
Shiroyanagi
Y
,
Shiroyanagi
LS
Разрушители эндокринной системы и гипоспадия: роль генистеина и фунгицида винклозолин
.
Урология
2007
;
70
:
618
—
621
0,15.
Hsieh
MH
,
Breyer
BN
,
Eisenberg
ML
,
Baskin
LS
.
Связь между гипоспадией, крипторхизмом, аногенитальной дистанцией и эндокринными нарушениями
.
Curr Urol Rep
2008
;
9
:
137
—
142
.16.
Fernandez
MF
,
Olmos
B
,
Granada
A
,
López-Espinosa
MJ
,
Molina-Molina
JM000
,9
Fernandez Cruz
M
,
Olea-Serrano
F
,
Olea
N
.
Воздействие на человека химических веществ, нарушающих работу эндокринной системы, и пренатальных факторов риска крипторхизма и гипоспадии: вложенное исследование случай – контроль
.
Environ Health Perspect
2007
;
115
:
8
—
14
. 17.
Kim
KS
,
Torres
CR
,
Yucel
S
,
Raimondo
K
,
Cunha
GR
,
Baskin
LS
.
Вызвание гипоспадии на мышиной модели путем воздействия синтетических эстрогенов на мать
.
Environ Res
2004
;
94
:
267
—
275
.18.
Willingham
E
,
Agras
K
,
Vilela
M
,
Baskin
LS
.
Лоратадин оказывает эстрогеноподобное действие и нарушает развитие полового члена у мышей
.
Дж Урол
2006
;
175
:
723
—
726
.19.
Seifert
AW
,
Harfe
BD
,
Cohn
MJ
.
Анализ клеточных клонов демонстрирует энтодермальное происхождение дистальных отделов уретры и промежности
.
Dev Biol
2008
;
318
:
143
—
152
.20.
Чжэн
Z
,
Армфилд
BA
,
Кон
MJ
.
Время разрушения рецепторов андрогенов и воздействия эстрогенов лежит в основе спектра врожденных аномалий полового члена
.
Proc Natl Acad Sci USA
2015
;
112
:
E7194
—
E7203
. 21.
Wolf
CJ
,
LeBlanc
GA
,
Ostby
JS
,
Серый
LE
Jr.
Характеристика периода чувствительности мужского полового развития плода к винклозолину
.
Toxicol Sci
2000
;
55
:
152
—
161
. 22.
Petiot
A
,
Perriton
CL
,
Dickson
C
,
Cohn
MJ
.
Развитие уретры млекопитающих контролируется Fgfr2-IIIb
.
Девелопмент
2005
;
132
:
2441
—
2450
.23.
Валлийский
M
,
Saunders
PT
,
Fisken
M
,
Scott
HM
,
Hutchison
GR
,
Smith
LB
,.
Выявление у крыс окна программирования маскулинизации репродуктивного тракта, нарушение которого приводит к гипоспадии и крипторхизму
.
J Clin Invest
2008
;
118
:
1479
—
1490
.24.
Coveney
D
,
Shaw
G
,
Hutson
JM
,
Renfree
MB
.
Влияние антиандрогенов на опускание яичек и закрытие паховой области у сумчатых таммарных валлаби (Macropus eugenii)
.
Репродукция
2002
;
124
:
865
—
874
0,25.
Sane
K
,
Pescovitz
OH
.
Клиторальный индекс: определение размера клитора у нормальных девочек и девочек с аномальным половым развитием
.
J Pediatr
1992
;
120
:
264
—
266
0,26.
Wiig
Ø
,
AE
Derocher
,
Cronin
MM
,
Skaare
JU
.
Псевдогермафродитные самки белых медведей на Шпицбергене
.
J Wildl Dis
1998
;
34
:
792
—
796
.27.
Leihy
MV
,
Shaw
G
,
Wilson
JD
,
Renfree
MB
.
Развитие уретры полового члена у таммарного валлаби
.
Sex Dev
2011
;
5
:
241
—
249
0,28.
Seifert
AW
,
Bouldin
CM
,
Choi
KS
,
Harfe
BD
,
Cohn
MJ
.
Многофазные и тканеспецифичные роли sonic hedgehog в клоакальной перегородке и развитии наружных гениталий
.
Девелопмент
2009
;
136
:
3949
—
3957
. 29.
Haraguchi
R
,
Mo
R
,
Hui
C
,
Motoyama
J
,
Makino
S
,
Shiroishi
T
,
Gaffield
Ямада
Г
.
Уникальные функции передачи сигналов Sonic hedgehog во время развития наружных гениталий
.
Девелопмент
2001
;
128
:
4241
—
4250
.30.
Perriton
CL
,
Powles
N
,
Chiang
C
,
Maconochie
MK
,
Cohn
MJ
.
Передача сигналов Sonic hedgehog из эпителия уретры контролирует развитие внешних половых органов
.
Dev Biol
2002
;
247
:
26
—
46
. 31.
Seifert
AW
,
Zheng
Z
,
Ormerod
BK
,
Cohn
MJ
.
Sonic hedgehog контролирует рост наружных половых органов, регулируя кинетику клеточного цикла
.
Nat Comms
2010
;
1
:
1
—
9
.32.
Haraguchi
R
,
Motoyama
J
,
Sasaki
H
,
Satoh
Y
,
Miyagawa
S
,
Nakagata
N
,
N
,
Ямада
Г
.
Молекулярный анализ скоординированного образования мочевого пузыря и мочеполовых органов с помощью передачи сигналов Hedgehog
.
Девелопмент
2007
;
134
:
525
—
533
0,33.
Miyagawa
S
,
Matsumaru
D
,
Murashima
A
,
Omori
A
,
Satoh
Y
,
Haraguchi
R
0 Ja
R
0
Iguchi
T
,
Nakagata
N
,
Hui
CC
,
Yamada
G
.
Роль пути sonic hedgehog-Gli2 в маскулинизации наружных гениталий
.
Эндокринология
2011
;
152
:
2894
—
2903
. 34.
Bai
CB
,
Auerbach
W
,
Lee
JS
,
Stephen
D
,
Joyner
AL
.
Gli2, но не Gli1, необходим для начальной передачи сигналов Shh и эктопической активации пути Shh
.
Девелопмент
2002
;
129
:
4753
—
4761
0,35.
Мариго
V
,
Дэви
RA
,
Zuo
Y
,
Cunningham
JM
,
Tabin
CJ
.
Биохимическим свидетельством того, что пропатчен, является рецептор Hedgehog
.
Nature
1996
;
384
:
176
—
179
0,36.
Stone
DM
,
Hynes
M
,
Armanini
M
,
Swanson
TA
.
Патченный ген супрессора опухолей кодирует кандидатный рецептор для Sonic hedgehog
.
Nature
1996
;
384
:
129
—
134
0,37.
Murone
M
,
Rosenthal
A
,
de Sauvage
FJ
.
Передача сигналов Sonic hedgehog с помощью патч-сглаженного рецепторного комплекса
.
Curr Biol
1999
;
9
:
76
—
84
.38.
Taipale
J
,
Cooper
MK
,
Maiti
T
,
Beachy
PA
.
Erratum: Patched действует каталитически, подавляя активность Smoothened
.
Nature
2002
;
418
:
892
—
896
0,39.
Alfaro
AC
,
Roberts
B
,
Kwong
L
,
Bijlsma
MF
,
Roelink
H
.
Ptch3 обеспечивает ответ Shh в Ptch2 — / — клетках
.
Девелопмент
2014
;
141
:
3331
—
3339
.40.
Chiu
HS
,
Szucsik
JC
,
Georgas
KM
,
Jones
JL
,
Rumballe
BA
,
Tang
SM
,
Tang
D
,
Tang
D
,
Tang
D
,
Льюис
AG
,
Ароноу
BJ
,
Лессард
JL
.
Сравнительный анализ экспрессии генов развития генитального бугорка обнаруживает предполагаемую аппендикулярную сеть Wnt7 для эпидермальной дифференцировки
.
Dev Biol
2010
;
344
:
1071
—
1087
.41.
Miyagawa
S
,
Moon
A
,
Haraguchi
R
,
Inoue
C
,
Harada
M
,
Nakahara
C
,
Nakahara
C
,
Matsumaru
D
,
Kaneko
T
,
Matsuo
I
.
Зависимые от дозировки сигналы hedgehog, интегрированные с передачей сигналов Wnt / β-catenin, регулируют формирование наружных гениталий как аппендикулярную программу
.
Девелопмент
2009
;
136
:
3969
—
3978
.42.
Линь
C
,
Инь
Y
,
Длинный
F
,
Ma
L
.
Тканевые потребности бета-катенина в развитии наружных половых органов
.
Девелопмент
2008
;
135
:
2815
—
2825
. 43.
Кавано
Y
,
Kypta
R
.
Секретируемые антагонисты сигнального пути Wnt
.
J Cell Sci
2003
;
116
:
2627
—
2634
. 44.
Hsieh
JC
,
Kodjabachian
L
,
Rebbert
ML
,
Rattner
A
,
Smallwood
PM
,
Samos
CH4
,
Samos
CH4
Давид
IB
,
Натанс
J
.
Новый секретируемый белок, который связывается с белками Wnt и ингибирует их активность
.
Nature
1999
;
398
:
431
—
436
. 45.
Ng
RCL
,
Matsumaru
D
,
Ho
ASH
,
Garcia-Barceló
MM
,
юаней
ZW
,
Smith
D
,
Smith
D
,,
Там
PKH
,
Ямада
G
,
Луи
VCH
.
Нарушение регуляции экспрессии Wnt ингибирующего фактора 1 (Wif1) приводит к аберрантной передаче сигналов Wnt-β-catenin и гибели клеток энтодермы клоаки, а также к аноректальным уродствам
.
дифференциал клеточной смерти
2014
;
21
:
978
—
989
.46.
Suzuki
K
,
Огино
Y
,
Murakami
R
,
Satoh
Y
,
Bachiller
D
,
Yamada
G
.
Эмбриональное развитие внешних гениталий мыши: понимание уникального способа органогенеза
.
Evol Dev
2002
;
4
:
133
—
141
0,47.
Renfree
MB
,
Короткий
RV
,
Shaw
G
.
Половая дифференциация мочеполовой системы плода и новорожденного таммарного валлаби, Macropus eugenii
.
Анат Эмбриол
1996
;
194
:
111
—
134
.48.
Батлер
CM
,
Shaw
G
,
Renfree
MB
.
Развитие полового члена и клитора у таммарного валлаби, Macropus eugenii
.
Анат Эмбриол (Берл)
1999
;
199
:
451
—
457
.49.
Омиди
А
,
Монжези
S
,
Набипур
А
.
Гипоспадия, дивертикул и агенезия диафиза полового члена козленка ( Capra hircus )
.
Comp Clin Pathol
2011
;
20
:
417
—
419
0,50.
Крафт
KH
,
Шукла
AR
,
Консервный
DA
.
Проксимальная гипоспадия
.
Sci World J
2011
;
11
:
894
—
906
. 51.
Renfree
MB
,
Chew
KY
,
Shaw
G
.
Индуцирование смены пола мочеполовой системы сумчатых
.
Дифференциация
2014
;
87
:
23
—
31
. 52.
Chew
KY
,
Pask
AJ
,
Hickford
D
,
Shaw
G
,
Renfree
MB
.
Двойная роль SHH во время развития фаллоса у сумчатых
.
Sex Dev
2014
;
8
:
166
—
177
.53.
Renfree
MB
,
Wilson
JD
,
Short
RV
,
Shaw
G
,
George
FW
.
Содержание стероидных гормонов гонад таммарного валлаби во время половой дифференциации
.
Biol Reprod
1992
;
47
:
644
—
647
. 54.
Wilson
JD
,
Джордж
FW
,
Shaw
G
,
Renfree
MB
.
Вирилизация молодых самцов таммарного валлаби, по-видимому, не опосредуется тестостероном плазмы или дигидротестостероном
.
Biol Reprod
1999
;
61
:
471
—
475
0,55.
Wilson
JD
,
Auchus
RJ
,
Leihy
МВт
,
Гурьев
OL
,
Estabrook
RW
,
Osborn
SM
,
Osborn
SM
,
Osborn
SM
Renfree
МБ
.
5α-андростан-3α, 17β-диол образуется в молодых семенниках таммарного валлаби посредством пути с участием 5α-прегнан-3α, 17α-диол-20-он в качестве ключевого промежуточного соединения
.
Эндокринология
2003
;
144
:
575
—
580
. 56.
Shaw
G
,
Renfree
MB
,
Leihy
MW
,
Shackleton
CH
,
Roitman
E
,
Wilson
JD
.
Формирование простаты у сумчатых опосредуется андрогенами яичек 5α-андростан-3α, 17β-диолом
.
Proc Natl Acad Sci USA
2000
;
97
:
12256
—
12259
. 57.
Ло
ZX
,
юаней
CX
,
Meng
QJ
,
Ji
Q
.
Юрское эвтериальное млекопитающее и расхождение сумчатых и плацентарных
.
Природа
2011
;
476
:
442
—
445
.58.
Renfree
MB
,
Fletcher
TP
,
Blanden
DR
,
Lewis
PR
,
Shaw
G
,
Gordon
K
,
Gordon
K
,,
Parer-Cook
E
,
Parer
D
.
Физиологические и поведенческие события во время рождения у макроподидных сумчатых
. В:
Grigg
GC
,
Jarman
P
,
Hume
ID
(ред.),
Кенгуру, валлаби и крысы-кенгуру
,
т. 1
.
Чиппинг Нортон, Новый Южный Уэльс
:
Суррей Битти и сыновья
;
1989
:
323
—
337
0,59.
Wai-Sum
O
,
Short
RV
,
Renfree
MB
,
Shaw
G
.
Первичный генетический контроль соматической половой дифференциации у млекопитающих
.
Nature
1988
;
331
:
716
—
717
.60.
Wood
JT
,
Carpenter
SM
,
Poole
WE
.
Доверительные интервалы возраста сумчатых, определенные по обмеру тела
.
Wildl Res
1981
;
8
:
269
—
274
0,61.
Tyndale-Biscoe
CH
,
Hinds
LA
.
Влияние незрелого семенника на половую дифференциацию таммарного валлаби, Macropus eugenii (Macropodidae: Marsupialia)
.
Reprod Fertil Dev
1989
;
1
:
243
—
254
0,62.
Renfree
MB
,
Fenelon
J
,
Wijiyanti
G
,
Wilson
JD
,
Shaw
G
.
Дифференциация протоков Вольфа по физиологическим концентрациям системно доставленных андрогенов
.
Dev Biol
2009
;
334
:
429
—
436
.63.
Whitworth
DJ
,
Shaw
G
,
Renfree
MB
.
Гонадное изменение пола развивающегося сумчатого яичника in vivo и in vitro
.
Девелопмент
1996
;
122
:
4057
—
4063
0,64.
Renfree
MB
,
Coveney
D
,
Shaw
G
.
Влияние эстрогенов на развитие самцов сумчатых животных
.
Reprod Fertil Dev
2001
;
13
:
231
—
240
0,65.
Евшин
И
,
Шарипов
R
,
Валеев
Т
,
Кел
А
,
Колпаков
Ф
.
GTRD: база данных сайтов связывания факторов транскрипции, идентифицированных с помощью экспериментов ChIP-seq
.
Nucleic Acids Res
2017
;
45
:
D61
—
D67
.66.
Khan
A
,
Fornes
O
,
Stigliani
A
,
Gheorghe
M
,
Castro-Mondragon
JA
,
van der Lee
Бесси
A
,
Chèneby
J
,
Kulkarni
SR
,
Tan
G
.
JASPAR 2018: обновление базы данных открытого доступа профилей связывания факторов транскрипции и ее веб-структуры
.
Nucleic Acids Res
2018
;
46
:
D260
—
D266
0,67.
Bolger
AM
,
Lohse
M
,
Usadel
B
.
Trimmomatic: гибкий триммер для данных последовательности Illumina
.
Биоинформатика
2014
;
30
:
2114
—
2120
0,68.
Trapnell
C
,
Williams
BA
,
Pertea
G
,
Mortazavi
A
,
Kwan
G
,
Van Baren
MJ
SL
MJ
,
,
Wold
BJ
,
Pachter
L
.
Сборка и количественная оценка транскриптов с помощью RNA-Seq выявляет неаннотированные транскрипты и переключение изоформ во время дифференцировки клеток
.
Nat Biotechnol
2010
;
28
:
511
—
515
0,69.
Гофф
L
,
Трапнелл
C
,
Келли
D
.
2012
;
2
0,70.
Apweiler
R
,
Bairoch
A
,
Wu
CH
,
Barker
WC
,
Boeckmann
B
,
Ferro
S
,
Хуанг
H
,
Лопес
R
,
Magrane
M
.
UniProt: универсальная база знаний о белках
.
Nucleic Acids Res
2004
;
32
:
115D
—
119D
.71.
Команда
RC
.
R: язык и среда для статистических вычислений
. . 2013. 72.
Trapnell
C
,
Williams
BA
,
Pertea
G
,
Mortazavi
A
,
Kwan
G
,
Van Baren
,
Ван Барен
Salzberg
SL
,
Wold
BJ
,
Pachter
L
.
Сборка и количественная оценка транскриптов с помощью RNA-Seq выявляет неаннотированные транскрипты и переключение изоформ во время дифференцировки клеток
.
Nat Biotechnol
2010
;
28
:
511
—
515
0,74.
Joseph
DR
,
O’Brien
DA
,
Sullivan
PM
,
Becchis
M
,
Tsuruta
JK
,
Petrusz
P
.
Сверхэкспрессия андроген-связывающего белка / полового гормона-связывающего глобулина у самцов трансгенных мышей: распределение тканей и фенотипические нарушения
.
Biol Reprod
1997
;
56
:
21
—
32
0,75.
Coquelin
A
,
Desjardins
C
.
Секреция лютеинизирующего гормона и тестостерона у молодых и старых самцов мышей
.
Am J Physiol
1982
;
243
:
E257
—
E263
.76.
Podlasek
CA
,
Zelner
DJ
,
Jiang
HB
,
Tang
Y
,
Houston
J
,
McKenna
KE
KE,
McKenna
.
Каскад Sonic hedgehog необходим для постнатального морфогенеза полового члена, дифференциации и гомеостаза взрослых
.
Biol Reprod
2003
;
68
:
423
—
438
.77.
Leal
F
,
Cohn
MJ
.
Развитие гемипенов в шаровом питоне змеи Python regius
.
Sex Dev
2015
;
9
:
6
—
20
0,78.
Gredler
ML
,
Seifert
AW
,
Cohn
MJ
.
Морфогенез и формирование паттерна фаллоса и клоаки американского аллигатора, alligator mississippiensis
.
Sex Dev
2015
;
9
:
53
—
67
0,79.
Ларкинс
CE
,
Кон
MJ
.
Развитие фаллоса у черепахи Trachemys scripta
.
Sex Dev
2015
;
9
:
34
—
42
0,80.
Ямагути
TP
,
Брэдли
A
,
McMahon
AP
,
Джонс
S
.
Путь Wnt5a лежит в основе роста множественных структур в эмбрионе позвоночных
.
Девелопмент
1999
;
126
:
1211
—
1223
.81.
Thie
M
,
Fuchs
P
,
Butz
S
,
Sieckmann
F
,
Hoschützky
H
,
Kemler
R
,
Kemler
R
,
Адгезивность апикальной поверхности эпителиальных клеток матки: роль целостности соединительного комплекса
.
евро J Cell Biol
1996
;
70
:
221
—
232
0,82.
Sirab
N
,
Terry
S
,
Giton
F
,
Caradec
J
,
Chimingqi
M
,
Moutereau
S
,
Moutereau
S
,
Moutereau
S
Taille
Adl
,
Kouyoumdjian
JC
,
Loric
S
.
Андрогены регулируют передачу сигналов и пролиферацию Hedgehog в андроген-зависимых клетках простаты
.
Int J Cancer
2012
;
131
:
1297
—
1306
,83.
Lee
J
,
Platt
KA
,
Censullo
P
,
i Altaba
AR
.
Gli1 является мишенью для Sonic hedgehog, который индуцирует развитие вентральной нервной трубки
.
Девелопмент
1997
;
124
:
2537
—
2552
.84.
Hooper
JE
,
Scott
MP
.
Патченный ген Drosophila кодирует предполагаемый мембранный белок, необходимый для формирования сегментарного паттерна
.
Cell
1989
;
59
:
751
—
765
0,85.
Джонс
TM
,
Фанг
VS
,
Ландау
RL
,
Розенфилд
R
.
Прямое ингибирование функции клеток Лейдига эстрадиолом
.
J Clin Endocrinol Metab
1978
;
47
:
1368
—
1373
0,86.
Akingbemi
BT
,
Sottas
CM
,
Koulova
AI
,
Klinefelter
GR
,
Hardy
MP
.
Ингибирование стероидогенеза яичек ксеноэстрогеном бисфенолом А связано со снижением секреции лютеинизирующего гормона гипофизом и снижением экспрессии гена стероидогенного фермента в клетках Лейдига крыс
.
Эндокринология
2004
;
145
:
592
—
603
0,87.
Delbes
G
,
Levacher
C
,
Duquenne
C
,
Racine
C
,
Pakarinen
P
,
Habert
R
.
Эндогенные эстрогены подавляют развитие эмбриональных клеток Лейдига мыши через рецептор эстрогена α
.
Эндокринология
2005
;
146
:
2454
—
2461
.88.
Камеда
C
,
Накамура
M
,
Танака
H
,
Ямасаки
A
,
Кубо
M
,
Танака
M
,
Танака
M
,,
Катано
М
.
Рецептор эстрогена-α участвует в регуляции пути передачи сигналов hedgehog при ERα-положительном раке желудка
.
Br J Cancer
2010
;
102
:
738
—
747
.89.
Koga
K
,
Nakamura
M
,
Nakashima
H
,
Akiyoshi
T
,
KUBO
M
,
Sato
Kuroki
N
,,
Номура
М
,
Танака
М
,
Катано
М
.
Новая связь между рецептором эстрогена α и путем hedgehog при раке груди
Механизм действия: гормоны с внутриклеточными рецепторами
Механизм действия: гормоны с внутриклеточными рецепторами
Рецепторы стероидных и тироидных гормонов расположены внутри клеток-мишеней, в цитоплазме или ядре, и функционируют как лиганд-зависимые факторы транскрипции .То есть комплекс гормон-рецептор связывается с промоторными областями чувствительных генов и стимулирует, а иногда и ингибирует транскрипцию этих генов.
Таким образом, механизм действия стероидных гормонов заключается в модуляции экспрессии генов в клетках-мишенях. Выборочно воздействуя на транскрипцию из батареи генов, изменяется концентрация этих соответствующих белков, что, несомненно, может изменить фенотип клетки.
Структура внутриклеточных рецепторов
Рецепторы стероидов и гормонов щитовидной железы являются членами большой группы («надсемейства») факторов транскрипции.В некоторых случаях в клетках экспрессируются несколько форм данного рецептора, что усложняет ответ. Все эти рецепторы состоят из единой полипептидной цепи, которая, при упрощенном анализе, состоит из трех отдельных доменов:
Аминоконец : В большинстве случаев эта область участвует в активации или стимуляции транскрипции путем взаимодействия с другими компонентами транскрипционного аппарата. Последовательность сильно варьирует среди разных рецепторов.
ДНК-связывающий домен : Аминокислоты в этой области отвечают за связывание рецептора со специфическими
последовательности ДНК.
Карбокси-конец или лиганд-связывающий домен : это область, которая связывает гормон.
В дополнение к этим трем коровым доменам, две другие важные области рецепторного белка представляют собой последовательность ядерной локализации, которая направляет белок в ядро, и домен димеризации, который отвечает за фиксацию двух рецепторов вместе в форме
способен связывать ДНК.
Связывание гормональных рецепторов и взаимодействие с ДНК
Являясь липидами, стероидные гормоны проникают в клетку путем простой диффузии через плазматическую мембрану. Гормоны щитовидной железы попадают в клетку путем облегченной диффузии. Рецепторы существуют либо в цитоплазме, либо в ядре, где они встречаются с гормоном. Когда гормон связывается с рецептором, происходит характерная серия событий:
Активация рецептора — это термин, используемый для описания конформационных изменений рецептора, вызванных связыванием гормона.Основным следствием активации является то, что рецептор становится компетентным связывать ДНК.
Активированные рецепторы связываются с «элементами гормонального ответа» , которые представляют собой короткие специфические последовательности ДНК, расположенные в промоторах гормонально-чувствительных генов. В большинстве случаев комплексы гормон-рецептор связывают ДНК парами, как показано на рисунке ниже.
Транскрипция тех генов, с которыми связан рецептор, нарушена. Чаще всего связывание рецептора стимулирует транскрипцию.Таким образом, комплекс гормон-рецептор функционирует как фактор транскрипции.
Как и следовало ожидать, существует ряд вариаций описанных выше тем в зависимости от конкретного рассматриваемого рецептора. Например, в отсутствие гормона некоторые внутриклеточные рецепторы действительно связывают свои гормональные ответные элементы слабо и подавляют транскрипцию, но при образовании комплекса с гормоном активируются и сильно стимулируют транскрипцию. Некоторые рецепторы связывают ДНК не с другим в своем роде, а с другим внутриклеточным рецептором.
В качестве конкретного примера рассмотрим глюкокортикоиды, тип стероидного гормона, который, вероятно, влияет на физиологию всех клеток в организме. На изображении справа изображена пара
рецепторы глюкокортикоидов (синий и зеленый вверху), связанные с их ответным элементом гормона ДНК (внизу). На этом изображении не видны два стероидных гормона.
Консенсусная последовательность гормонального ответного элемента в этом случае (называемого глюкокортикоидным ответным элементом ) — это GGTACANNNTGTTCT, где N — любой нуклеотид.
Клетки рака груди, взятые во время биопсии или хирургического вмешательства, будут проверены на наличие в них определенных белков, которые являются рецепторами эстрогена или прогестерона. Когда гормоны эстроген и прогестерон присоединяются к этим рецепторам, они способствуют росту рака. Раковые опухоли называются положительными по рецепторам гормонов или отрицательными по рецепторам гормонов в зависимости от наличия у них этих рецепторов (белков).Знание статуса рецепторов гормонов важно при выборе вариантов лечения. Спросите своего врача о статусе рецепторов гормонов и о том, что это значит для вас.
Что такое рецепторы эстрогена и прогестерона?
Рецепторы
— это белки в клетках или на них, которые могут присоединяться к определенным веществам в крови. Нормальные клетки груди и некоторые клетки рака груди имеют рецепторы, которые прикрепляются к гормонам эстрогену и прогестерону и зависят от этих гормонов для роста.
Клетки рака груди могут иметь один, оба или ни один из этих рецепторов.
ER-положительный: Рак молочной железы с рецепторами эстрогена называется ER-положительным (или ER +) раком.
PR-позитивный : Рак груди с рецепторами прогестерона называется PR-позитивным раком (или PR +).
Гормон рецептор-положительный : Если раковая клетка имеет один или оба рецептора выше, можно использовать термин гормон-рецептор-положительный (также называемый гормон-положительный или HR +) рак груди.
Гормон рецептор-отрицательный : Если раковая клетка не имеет ни эстрогенового, ни прогестеронового рецептора, это называется гормонально-отрицательным рецептором (также называемым гормонально-отрицательным или HR-).
Предотвращение присоединения гормонов эстрогена и прогестерона к рецепторам может помочь предотвратить рост и распространение рака. Есть препараты, которые можно использовать для этого.
Почему важно знать статус рецепторов гормонов?
Знание статуса рецепторов гормонов рака помогает врачам решить, как его лечить.Если у вашего рака есть один или оба этих рецептора гормона, можно использовать препараты гормональной терапии, чтобы либо снизить уровень эстрогена, либо остановить действие эстрогена на клетки рака груди. Этот вид лечения полезен при раке молочной железы с положительными рецепторами гормонов, но не работает с опухолями, отрицательными по рецепторам гормонов (как ER-, так и PR-отрицательными).
Все инвазивные виды рака молочной железы должны быть проверены на оба этих гормональных рецептора либо на образце биопсии, либо при удалении опухоли хирургическим путем.Примерно в 2 из 3 случаев рака молочной железы есть хотя бы один из этих рецепторов. Этот процент выше у пожилых женщин, чем у молодых женщин. DCIS также следует проверять на наличие рецепторов гормонов.
Что означают результаты теста на рецепторы гормонов?
Тест, называемый иммуногистохимией (ИГХ), чаще всего используется для определения наличия в раковых клетках рецепторов эстрогена и прогестерона. Результаты теста помогут вам и вашей команде по лечению рака принять правильное решение о лечении.
Результаты анализа покажут вам статус рецептора гормонов.Он будет говорить, что опухоль является рецептор-положительным гормоном , если по крайней мере 1% протестированных клеток имеют рецепторы эстрогена и / или прогестерона. В противном случае тест покажет, что опухоль отрицательная по рецепторам гормона .
Положительные по рецепторам гормонов (или положительные по гормонам) клетки рака молочной железы имеют рецепторы либо эстрогена (ER), либо рецептора прогестерона (PR), либо обоих. Эти виды рака груди можно лечить с помощью препаратов гормональной терапии, которые снижают уровень эстрогена или блокируют рецепторы эстрогена.Раковые опухоли, положительные по рецепторам гормонов, имеют тенденцию расти медленнее, чем рак, отрицательные по рецепторам гормонов. Женщины с раком, положительным по рецепторам гормонов, имеют тенденцию к лучшему прогнозу в краткосрочной перспективе, но эти виды рака могут иногда возвращаться через много лет после лечения.
Рак молочной железы, отрицательный по рецепторам гормонов (или отрицательный по гормонам), не имеет рецепторов эстрогена и прогестерона. При этом раке лечение гормональными препаратами не помогает. Эти виды рака имеют тенденцию расти быстрее, чем рак, положительный по рецепторам гормонов.Если они возвращаются после лечения, часто в первые несколько лет. Раковые образования, отрицательные по рецепторам гормонов, чаще встречаются у женщин, которые еще не прошли менопаузу.
Тройно-отрицательные клетки рака молочной железы не имеют рецепторов эстрогена или прогестерона, а также не производят слишком много белка под названием HER2 . Эти виды рака чаще встречаются у женщин моложе 40 лет, афроамериканок или женщин с мутацией в гене BRCA 1. Тройной отрицательный рак груди растет и распространяется быстрее, чем большинство других видов рака груди.Поскольку раковые клетки не имеют рецепторов гормонов, гормональная терапия не помогает при лечении этих видов рака. И поскольку у них не так много HER2, лекарства, нацеленные на HER2, тоже не помогают. Химиотерапия все еще может быть полезной. См. Раздел «Тройной отрицательный рак груди», чтобы узнать больше.
Тройной положительный раковые опухоли являются ER-положительными, PR-положительными и HER2-положительными. Эти виды рака можно лечить гормональными препаратами, а также лекарствами, нацеленными на HER2.
Хронический стресс подвергает опасности ваше здоровье
Хронический стресс подвергает опасности ваше здоровье
Хронический стресс может нанести ущерб вашему разуму и телу.Примите меры, чтобы контролировать свой стресс.
Персонал клиники Мэйо
Ваше тело запрограммировано так, чтобы реагировать на стресс таким образом, чтобы защитить вас от угроз со стороны хищников и других агрессоров. Такие угрозы сегодня редки, но это не значит, что в жизни нет стрессов.
Напротив, вы, вероятно, ежедневно сталкиваетесь со многими требованиями, такими как выполнение огромной рабочей нагрузки, оплата счетов и забота о своей семье. Ваше тело воспринимает эти так называемые незначительные неприятности как угрозу. В результате вам может казаться, что вас постоянно атакуют.Но ты можешь дать отпор. Вы не должны позволять стрессу контролировать свою жизнь.
Понимание естественной реакции на стресс
Когда вы сталкиваетесь с предполагаемой угрозой, например, с большой собакой, лающей на вас во время утренней прогулки, ваш гипоталамус, крошечная область в основании вашего мозга, включает систему сигнализации в вашем теле. Посредством комбинации нервных и гормональных сигналов эта система побуждает надпочечники, расположенные над почками, выделять выброс гормонов, включая адреналин и кортизол.
Адреналин увеличивает частоту сердечных сокращений, повышает кровяное давление и увеличивает запасы энергии. Кортизол, основной гормон стресса, увеличивает содержание сахара (глюкозы) в кровотоке, усиливает использование глюкозы мозгом и увеличивает доступность веществ, восстанавливающих ткани.
Кортизол также подавляет функции, которые могут быть несущественными или вредными в ситуации борьбы или бегства. Он изменяет реакцию иммунной системы и подавляет пищеварительную систему, репродуктивную систему и процессы роста.Эта сложная естественная система сигнализации также взаимодействует с областями мозга, которые контролируют настроение, мотивацию и страх.
Когда естественная реакция на стресс становится безумной
Система реакции организма на стресс обычно является самоограничивающейся. Как только предполагаемая угроза миновала, уровень гормонов возвращается к норме. Когда уровни адреналина и кортизола падают, частота сердечных сокращений и артериальное давление возвращаются к исходным уровням, а другие системы возобновляют свою обычную деятельность.
Но когда стрессоры всегда присутствуют и вы постоянно чувствуете себя атакованными, реакция «бей или беги» остается включенной.
Длительная активация системы реакции на стресс и последующее чрезмерное воздействие кортизола и других гормонов стресса могут нарушить почти все процессы в организме. Это подвергает вас повышенному риску возникновения многих проблем со здоровьем, в том числе:
Беспокойство
Депрессия
Проблемы с пищеварением
Головные боли
Напряжение и боль в мышцах
Болезнь сердца, сердечный приступ, высокое кровяное давление и инсульт
Проблемы со сном
Прибавка в весе
Нарушение памяти и концентрации
Вот почему так важно научиться здоровым способам справляться с факторами жизненного стресса.
Почему вы так реагируете на жизненные стрессоры
Ваша реакция на потенциально стрессовое событие отличается от чьей-либо другой. На вашу реакцию на факторы жизненного стресса влияют такие факторы, как:
Генетика. Гены, контролирующие реакцию на стресс, поддерживают у большинства людей довольно устойчивый эмоциональный уровень, лишь изредка настраивая тело на борьбу или бегство. Гиперактивные или недостаточные реакции на стресс могут возникать из-за небольших различий в этих генах.
Жизненный опыт. Сильные стрессовые реакции иногда можно отнести к травматическим событиям. Люди, которым пренебрегали или подвергались жестокому обращению в детстве, как правило, особенно уязвимы к стрессу. То же самое касается выживших в авиакатастрофе, военнослужащих, полицейских и пожарных, а также людей, пострадавших от насильственных преступлений.
У вас могут быть друзья, которые кажутся расслабленными почти во всем, а другие сильно реагируют на малейший стресс.Большинство людей реагируют на жизненные стрессоры где-то между этими крайностями.
Научиться правильно реагировать на стресс
Стрессовые события — это факты жизни. И вы не сможете изменить свою текущую ситуацию. Но вы можете предпринять шаги, чтобы управлять влиянием этих событий на вас.
Вы можете научиться определять, что вызывает у вас стресс, и как заботиться о себе физически и эмоционально в стрессовых ситуациях.
Стратегии управления стрессом включают:
Соблюдение здоровой диеты, регулярные физические упражнения и много сна
Практика методов релаксации, таких как йога, глубокое дыхание, массаж или медитация
Вести дневник и записывать свои мысли или то, за что вы благодарны в своей жизни
Найдите время для хобби, например, чтения, прослушивания музыки или просмотра любимого шоу или фильма
Укрепление здоровой дружбы и общение с друзьями и семьей
Обладать чувством юмора и находить способы включить юмор и смех в свою жизнь, например, смотреть смешные фильмы или просматривать сайты с анекдотами
Волонтерство в вашем сообществе
Организация и расстановка приоритетов в том, что вам нужно выполнить дома и на работе, и удаление ненужных задач
Обращение за профессиональной консультацией, которая может помочь вам разработать конкретные стратегии преодоления стресса
Избегайте нездоровых способов управления стрессом, таких как употребление алкоголя, табака, наркотиков или избыточной пищи.Если вы обеспокоены тем, что употребление этих продуктов увеличилось или изменилось из-за стресса, поговорите со своим врачом.
Награды за умение справляться со стрессом могут включать душевное спокойствие, меньше стресса и беспокойства, лучшее качество жизни, улучшение таких состояний, как высокое кровяное давление, лучший самоконтроль и сосредоточенность, а также лучшие отношения. И это может даже привести к более долгой и здоровой жизни.
Получите самые свежие советы по здоровью от клиники Мэйо.
в ваш почтовый ящик.
Зарегистрируйтесь бесплатно и будьте в курсе новостей
достижения, советы по здоровью и актуальные темы о здоровье,
например, COVID-19, плюс советы экспертов по поддержанию здоровья.
Узнайте больше о нашем использовании данных
Чтобы предоставить вам наиболее актуальную и полезную информацию и понять, какие
Информация
выгодно, мы можем объединить вашу электронную почту и информацию об использовании веб-сайта с другими
информация, которая у нас есть о вас.Если вы пациент клиники Мэйо,
это может включать защищенную медицинскую информацию (PHI). Если мы объединим эту информацию
с вашей PHI, мы будем рассматривать всю эту информацию как PHI,
и будет использовать или раскрывать эту информацию только в соответствии с нашим уведомлением о конфиденциальности
практики. Вы можете отказаться от рассылки по электронной почте.
в любое время, нажав ссылку «Отказаться от подписки» в электронном письме.
Подписывайся!
Спасибо за подписку
Наша электронная рассылка Housecall будет держать вас в курсе
на последней информации о здоровье.
Сожалеем! Наша система не работает. Пожалуйста, попробуйте еще раз.
Что-то пошло не так на нашей стороне, попробуйте еще раз.
Пожалуйста, попробуйте еще раз
8 июля 2021 г. Показать ссылки
Как стресс влияет на ваше здоровье. Американская психологическая ассоциация. https://www.apa.org/topics/stress/health. По состоянию на 19 марта 2021 г.
Стрессовое воздействие на организм.Американская психологическая ассоциация. https://www.apa.org/topics/stress/body. По состоянию на 19 марта 2021 г.
Снижение стресса: как стресс влияет на тело? Американская Ассоциация Сердца. https://www.heart.org/en/healthy-living/healthy-lifestyle/stress-management/lower-stress-how-does-stress-affect-the-body. По состоянию на 18 марта 2021 г.
Стресс и ваше здоровье. Министерство здравоохранения и социальных служб США. https://www.womenshealth.gov/mental-health/good-mental-health/stress-and-your-health.По состоянию на 18 марта 2021 г.
AskMayoExpert. Управление стрессом и устойчивость (взрослые). Клиника Майо. 2019.
Seaward BL. Основы управления стрессом. 5-е изд. Jones & Bartlett Learning; 2021.
Seaward BL. Управление стрессом: навыки самообслуживания, личная устойчивость и баланс между работой и личной жизнью в быстро меняющемся мире. 10-е изд. Jones & Bartlett Learning; 2022.

 Иногда пара лишних килограммов лучше, чем испорченное здоровье.
Иногда пара лишних килограммов лучше, чем испорченное здоровье.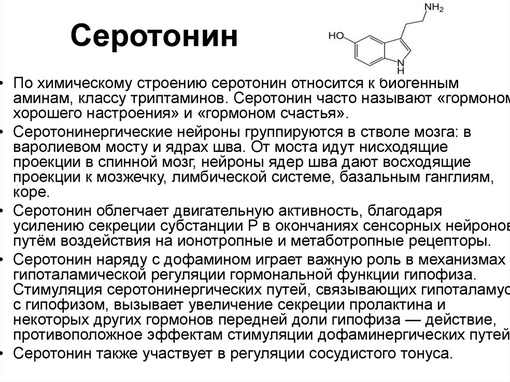 Все они вырабатываются в яичниках, отвечают за формирование фигуры по женскому типу, развитие половых органов и регулируют менструальный цикл. «Если эстроген вырабатывается в нормальном количестве, он очень положительно влияет на внешний вид женщины, – рассказывает Ирина Вяткина, врач гинеколог-эндокринолог «Клиники Марины Рябус». – Этот гормон участвует в процессе обновления клеток всего организма, в том числе и кожи. Эстроген положительно влияет и на красоту волос: придает им блеск, сохраняет молодость и здоровье. Эстрогены поднимают настроение, располагают к флирту и кокетству, препятствуют отложению холестерина в стенках сосудов, заставляют блестеть глаза, разглаживают морщинки, делают кожу эластичной и упругой, тем самым делая женщину красивой. Кроме того, эстрогены поднимают настроение и располагают к флирту и кокетству – они способствует развитию влечения у женщин к лицам мужского пола. На фоне увеличения выработки эстрогенов улучшается координация движений и тем самым походка, меняется качество речи.
Все они вырабатываются в яичниках, отвечают за формирование фигуры по женскому типу, развитие половых органов и регулируют менструальный цикл. «Если эстроген вырабатывается в нормальном количестве, он очень положительно влияет на внешний вид женщины, – рассказывает Ирина Вяткина, врач гинеколог-эндокринолог «Клиники Марины Рябус». – Этот гормон участвует в процессе обновления клеток всего организма, в том числе и кожи. Эстроген положительно влияет и на красоту волос: придает им блеск, сохраняет молодость и здоровье. Эстрогены поднимают настроение, располагают к флирту и кокетству, препятствуют отложению холестерина в стенках сосудов, заставляют блестеть глаза, разглаживают морщинки, делают кожу эластичной и упругой, тем самым делая женщину красивой. Кроме того, эстрогены поднимают настроение и располагают к флирту и кокетству – они способствует развитию влечения у женщин к лицам мужского пола. На фоне увеличения выработки эстрогенов улучшается координация движений и тем самым походка, меняется качество речи. Она становится более приятной для восприятия».
Она становится более приятной для восприятия». У небеременных женщин прогестерон также присутствует и повышается во второй фазе менструального цикла, что, увы, негативно сказывается на внешности. «Прогестерон способствует задержке жидкости в организме, повышает проницаемость сосудистой стенки, кожа становится растяжимой, повышается выделение кожного сала и появляются угревые высыпания. Может появляться одутловатость лица за счет задержки жидкости», – говорит Ирина Вяткина. Прогестерон также активно способствует запасанию жира, подготавливая нас к беременности, даже если мы ее не планируем. Поэтому во второй фазе цикла многие набирают один-два лишних килограмма. Кроме того, прогестерон снижает сопротивляемость организма инфекциям, что благотворно влияет на патогенную микрофлору кожи из-за ослабления иммунитета и зачастую приводит к образованию прыщей.
У небеременных женщин прогестерон также присутствует и повышается во второй фазе менструального цикла, что, увы, негативно сказывается на внешности. «Прогестерон способствует задержке жидкости в организме, повышает проницаемость сосудистой стенки, кожа становится растяжимой, повышается выделение кожного сала и появляются угревые высыпания. Может появляться одутловатость лица за счет задержки жидкости», – говорит Ирина Вяткина. Прогестерон также активно способствует запасанию жира, подготавливая нас к беременности, даже если мы ее не планируем. Поэтому во второй фазе цикла многие набирают один-два лишних килограмма. Кроме того, прогестерон снижает сопротивляемость организма инфекциям, что благотворно влияет на патогенную микрофлору кожи из-за ослабления иммунитета и зачастую приводит к образованию прыщей.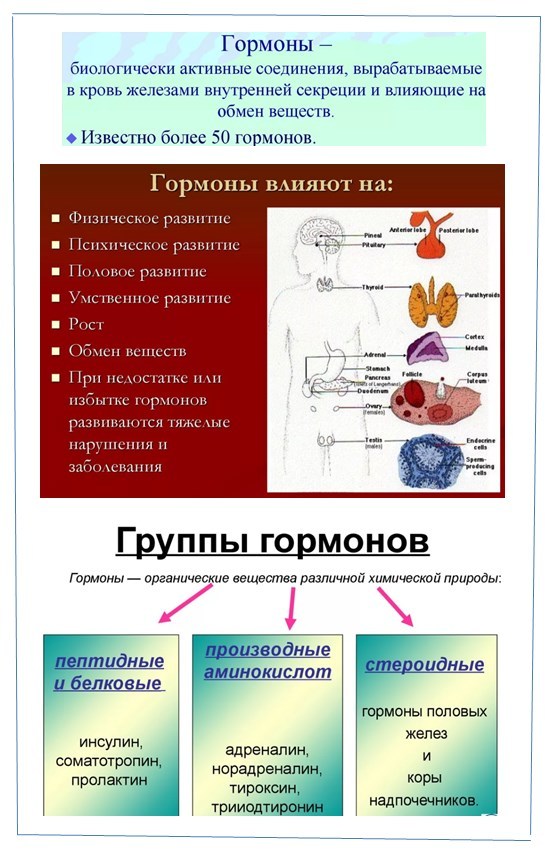 Пониженное содержание дегидроэпиандростерона негативно сказывается как на всем гормональном фоне, так и непосредственно на внешности и самочувствии. В первую очередь дефицит ДГЭА проявляется ломкостью ногтей и выпадением волос, плохим настроением и изменением продолжительности менструального цикла.
Пониженное содержание дегидроэпиандростерона негативно сказывается как на всем гормональном фоне, так и непосредственно на внешности и самочувствии. В первую очередь дефицит ДГЭА проявляется ломкостью ногтей и выпадением волос, плохим настроением и изменением продолжительности менструального цикла.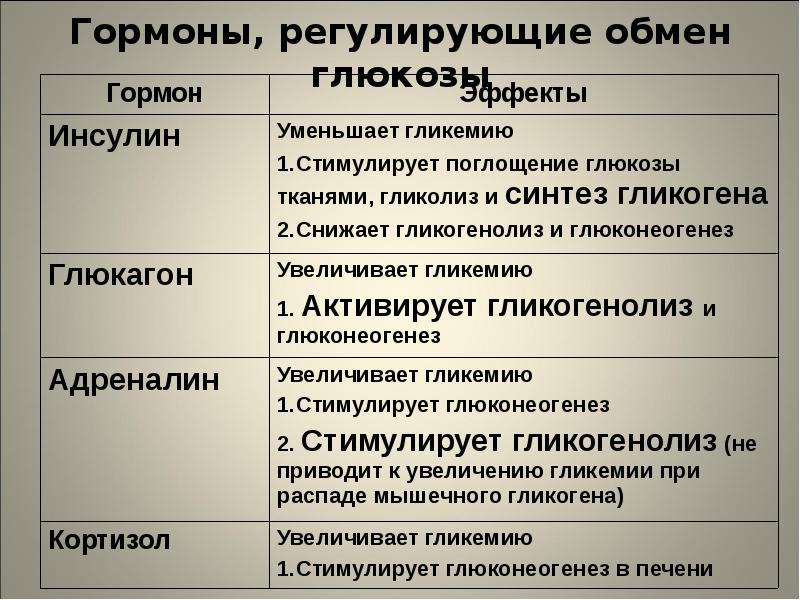 Критерием темноты может служить факт невозможности рассмотреть свою вытянутую руку. Любой свет – это враг мелатонина, в том числе циферблат электронного будильника, не говоря уже об экране смартфона, компьютера, телевизора или фонаре на улице. Уменьшение уровня мелатонина – это сигнал другим органам и железам, что настало время для отдыха. Яичники у женщин прекращают работу, сильно снижается уровень эстрогена, приходит менопауза. У мужчин значительно снижается количество гормона тестостерона». Таким образом, ночной образ жизни напрямую связан не только с ухудшением работы всего организма, но и с лишним весом и быстрым увяданием кожи.
Критерием темноты может служить факт невозможности рассмотреть свою вытянутую руку. Любой свет – это враг мелатонина, в том числе циферблат электронного будильника, не говоря уже об экране смартфона, компьютера, телевизора или фонаре на улице. Уменьшение уровня мелатонина – это сигнал другим органам и железам, что настало время для отдыха. Яичники у женщин прекращают работу, сильно снижается уровень эстрогена, приходит менопауза. У мужчин значительно снижается количество гормона тестостерона». Таким образом, ночной образ жизни напрямую связан не только с ухудшением работы всего организма, но и с лишним весом и быстрым увяданием кожи. При повышении уровня кортизола кожа истончается и становится склонной к появлению пигментации. При нормальной выработке кортизола кожа упругая, ровная и красивая», – рассказывает Ирина Вяткина. Повышенный кортизол способствует накоплению жира в области лица и живота, поэтому справедливо выражение, что стресс мешает похудеть.
При повышении уровня кортизола кожа истончается и становится склонной к появлению пигментации. При нормальной выработке кортизола кожа упругая, ровная и красивая», – рассказывает Ирина Вяткина. Повышенный кортизол способствует накоплению жира в области лица и живота, поэтому справедливо выражение, что стресс мешает похудеть. Сейчас набирает популярность тестостероновая терапия, которая улучшает качество кожи при возрастном снижении синтеза гормона. Врачи предупреждают, что самостоятельным приемом тестостерона можно сильно навредить своему здоровью – такая терапия проводится только по показаниям под наблюдением эндокринолога.
Сейчас набирает популярность тестостероновая терапия, которая улучшает качество кожи при возрастном снижении синтеза гормона. Врачи предупреждают, что самостоятельным приемом тестостерона можно сильно навредить своему здоровью – такая терапия проводится только по показаниям под наблюдением эндокринолога.

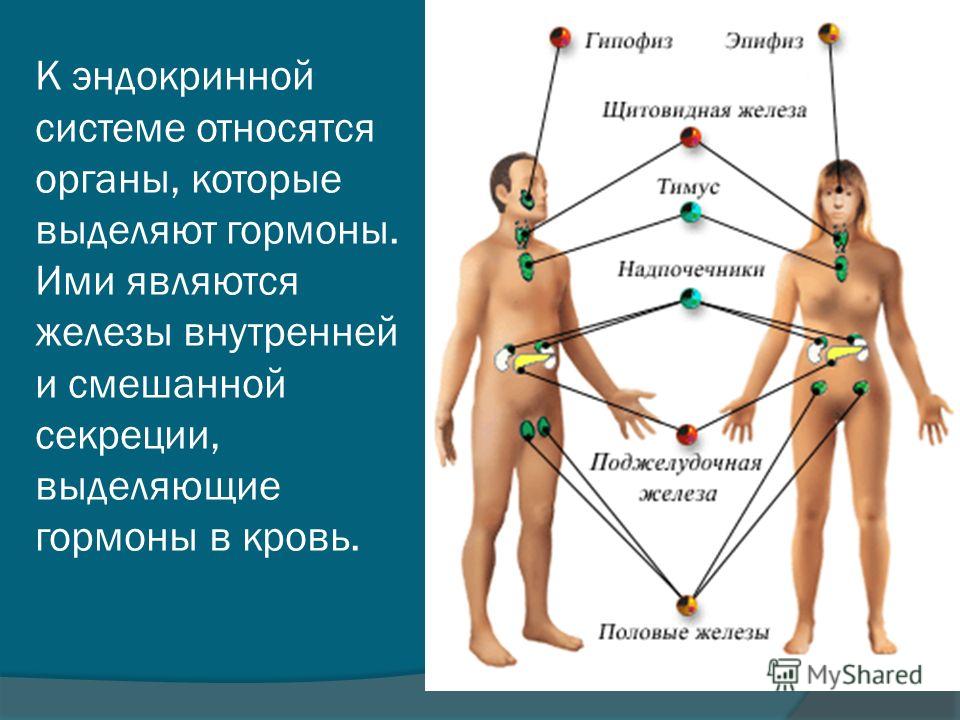 А вот желание обойтись одной косметологией, без плановой терапии, – это, как правило, трата денег и времени на процедуры, которые не сработают или сработают недостаточно хорошо. И дело не в самих процедурах, а в патологиях пациента. Так, например, если убрать лазером сосудистую сетку с лица, но не решить проблемы с избытком эстрогена или дефицитом прогестерона, из-за которых она часто возникает, – сосуды быстро вернутся. То же касается и гиперпигментации: казалось бы, бесследно удаленные лазером пигментные пятна будут возвращаться до тех пор, пока не придут хотя бы в относительный порядок стероидные гормоны и гормоны гипофиза, влияющие на клетки-меланоциты. Словом, при эндокринных проблемах, которые сегодня бывают у большого количества пациентов, сочетание косметологии с плановой терапией работает на порядок лучше, чем что-то одно.»
А вот желание обойтись одной косметологией, без плановой терапии, – это, как правило, трата денег и времени на процедуры, которые не сработают или сработают недостаточно хорошо. И дело не в самих процедурах, а в патологиях пациента. Так, например, если убрать лазером сосудистую сетку с лица, но не решить проблемы с избытком эстрогена или дефицитом прогестерона, из-за которых она часто возникает, – сосуды быстро вернутся. То же касается и гиперпигментации: казалось бы, бесследно удаленные лазером пигментные пятна будут возвращаться до тех пор, пока не придут хотя бы в относительный порядок стероидные гормоны и гормоны гипофиза, влияющие на клетки-меланоциты. Словом, при эндокринных проблемах, которые сегодня бывают у большого количества пациентов, сочетание косметологии с плановой терапией работает на порядок лучше, чем что-то одно.» Порой можно обойтись и вовсе без нее: на нарушение работы гормональной системы может влиять любое другое заболевание и даже низкий уровень витаминов и микроэлементов. В таких случаях врач будет корректировать именно первопричину.
Порой можно обойтись и вовсе без нее: на нарушение работы гормональной системы может влиять любое другое заболевание и даже низкий уровень витаминов и микроэлементов. В таких случаях врач будет корректировать именно первопричину.
 Именно этот гормон играет ключевую роль в поддержании мышечного тонуса кожи, так как он регулирует синтез коллагена, миозина, актина и других белков.
Именно этот гормон играет ключевую роль в поддержании мышечного тонуса кожи, так как он регулирует синтез коллагена, миозина, актина и других белков.  Если глаза – это зеркало души, то кожа – зеркало гормональной системы. Если гормональная система в балансе — кожа выглядит сияющей и старение замедляется, а если наоборот — старение и увядание кожи происходят очень быстро.
Если глаза – это зеркало души, то кожа – зеркало гормональной системы. Если гормональная система в балансе — кожа выглядит сияющей и старение замедляется, а если наоборот — старение и увядание кожи происходят очень быстро.
