НЕ РАСТУТ ПЛЕЧИ? КАЧАЙТЕ ИХ ТРИСЕТАМИ! | Бодибилдинг И фитнес
НЕ РАСТУТ ПЛЕЧИ? КАЧАЙТЕ ИХ ТРИСЕТАМИ!Если вы все перепробовали, но обзавестись шаровидными дельтами так и не вышло, значит пришло время менять свой подход к их прокачке. Мне, с моими длинными руками, во всяком случае, эта методика очень помогла. И не только в плане развития дельт. Короче, если плечи не растут – бомбите их трисетами!
В чем суть такого приёма тренировки плеч ?Суть в следующем, группы мышц, состоящие из нескольких пучков есть смысл качать мини-комплексами. Наука говорит, а тренера суперзвезд, тот же Джордж Фаррах (тренер Кая Грина и Бренча Уоррена) подтверждает, что двуглавую (бицепс) лучше качать суперсетами, четырехглавую (квадрицепс), гигантскими сериями из четырех упражнений подряд, а вот трехглавую (трицепс) и дельты – трисетами.
Другими словами, проработка каждого из пучков дельтовидных мышц менее эффективна в плане набора массы, чем их совместная прокачка. Когда передняя, средняя и задние дельты трудятся в единой связке, повышается их силовой потенциал, ускоряется восстановление и главное — усиливает кровенаполнение. А с ним и синтез такого важного для набора массы гормона, как соматропин.
Когда передняя, средняя и задние дельты трудятся в единой связке, повышается их силовой потенциал, ускоряется восстановление и главное — усиливает кровенаполнение. А с ним и синтез такого важного для набора массы гормона, как соматропин.
Качать плечи трисетами – это реально круто, но при создании каждого такого мини-комплекса стоит учитывать один важный момент. На какой именно пучок дельтовидной мышцы он направлен?
ЧИТАТЬ ТАКЖЕ, УПРАЖНЕНИЕ ДЛЯ ЗАДНЕЙ ДЕЛЬТЫ, КОТОРОЕ НИКТО НЕ ДЕЛАЕТ
Ясное дело, что, выполняя три упражнения подряд без отдыха, эффективность каждого последующего будет снижаться примерно на 20-25%. То есть, если отстает в развитии задний пучок и нужно увеличить именно его, качать заднюю дельту последней будет глупо. Наоборот, с упражнения для нее и нужно выполнять всю тройку упражнений.
Как составить трисет на плечи грамотно?Принцип приоритета в составлении трисета для плеч работает, как нельзя лучше. Поэтому, первое упражнение должно быть направленно на наиболее важный пучок мышц плеч. Быть наиболее тяжелым, базовым и по возможности выполняться стоя, поскольку так можно осилить больший рабочий вес.
Поэтому, первое упражнение должно быть направленно на наиболее важный пучок мышц плеч. Быть наиболее тяжелым, базовым и по возможности выполняться стоя, поскольку так можно осилить больший рабочий вес.
Второе может быть чуть полегче, также выполняться со свободным весом, но уже сидя. А третье – быть изолированным и делаться в тренажере. Но это еще не все.
Число повторений должно увеличиваться с каждым последующим движением. Например, вот так, в первом упражнении – 8 повторений, во втором – 12 и в третьем, заключительном – 20. Зачем так париться? Объясняю: такое изменение диапазона повторов в одном комплексе позволит прокачать сразу оба вида мышечных волокон и повысить тем самым шансы на рост мышц.
Качая плечи на массу стоит работать с тренировочным напарникомКачая плечи на массу стоит работать с тренировочным напарником
Примечание: прокачка плеч таким образом – это очень тяжела работа, помощь партнера, особенно во втором упражнении будет очень кстати.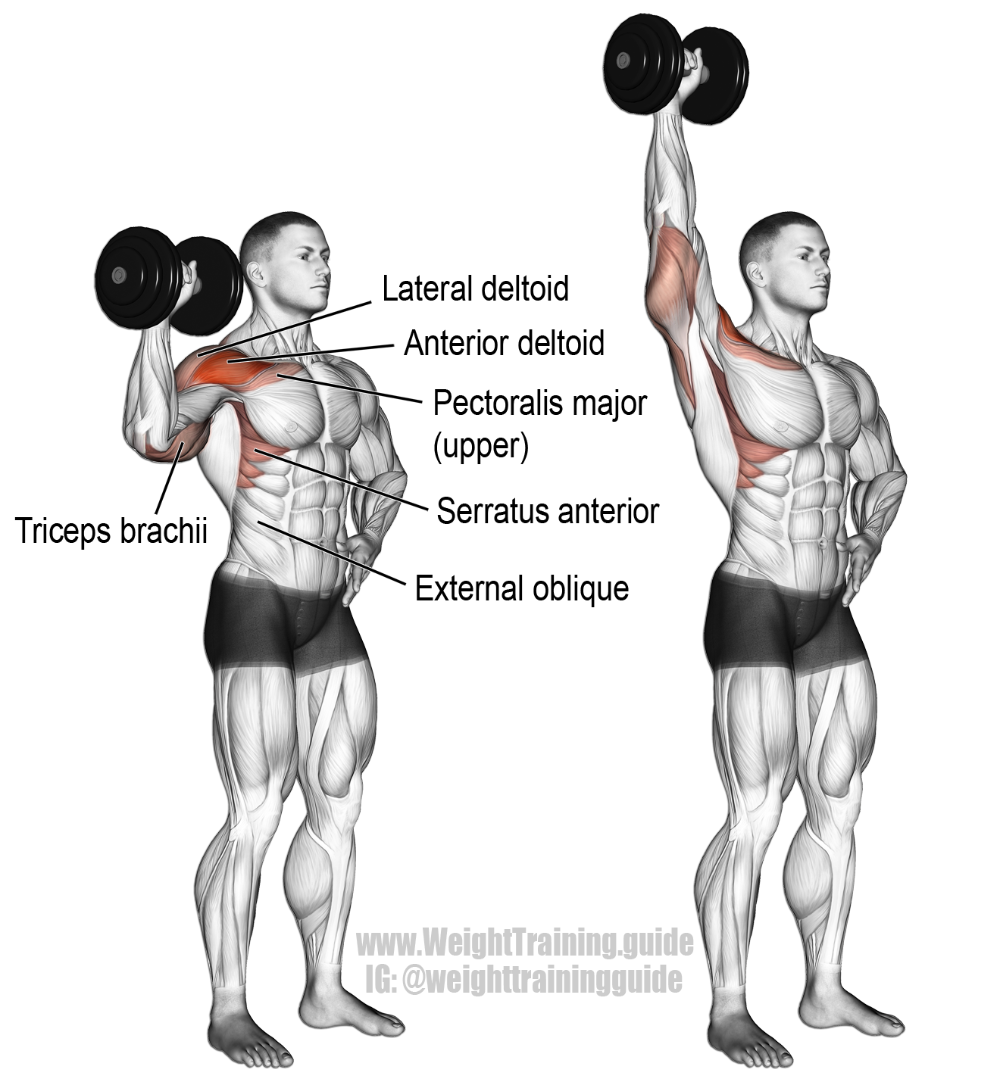 Пара форсированных повторений, выполненных с его помощью, усилят отдачу от выполнения всего трисета.
Пара форсированных повторений, выполненных с его помощью, усилят отдачу от выполнения всего трисета.
Исходя из вышеизложенного алгоритма, мини-комплекс для средней дельты может быть вот таким:
- Упражнение на среднюю дельту. Разведение с гантелями стоя – 8 повторений
- Упражнение на переднюю дельту. Подъем гантелей перед собой сидя – 12 повторений
- Упражнение на заднюю дельту. Обратные разведения в бабочке – 20 повторений.
Трисет для передней дельты таким:
- Упражнение на переднюю дельту. Подъем одной гантели перед собой стоя – 8 повторений
- Упражнение на среднюю дельту. Жим гантелей сидя – 12 повторений
- Упражнение на заднюю дельту. Скрещивания рук в кроссовере – 20 повторений.
А, трисет для заднего пучка вот таким:
- Упражнение на заднюю дельту.
 Тяга штанги в наклоне к груди – 8 повторений
Тяга штанги в наклоне к груди – 8 повторений - Упражнение на среднюю дельту. Разведение гантелей сидя – 12 повторений
- Упражнение на переднюю дельту. Махи в кроссовере двумя руками перед собой – 20 повторений
 Тяга штанги в наклоне к груди – 8 повторений
Тяга штанги в наклоне к груди – 8 повторенийТрисеты — это реальный способ накачать плечи. Хоть в зале, хоть дома
Заключение
Вариантов составления трисетов для плеч может быть великое множество, но грамотная их компоновка и соблюдение правил выполнения помогут стать обладателем широких плеч намного быстрее. Даже в условиях тренировки плеч в домашних условиях.
Программа на набор мышечной массы «Трапеции, грудь, плечи», автор: AYarmots — AtletIQ.com
Программа рассчитана на 6 тренировочных дней с получасовой загруженностью. Небольшая продолжительность тренировок будет компенсирована плотностью их расписания: рабочие будни разделены всего одним днем отдыха.
Хотя впереди тебя ждет интенсивная работа, разнообразие «поденных» комплексов упражнений не даст твоему телу уставать от нагрузок настолько, чтобы недовосстанавливаться.
Целевая аудитория пользователей
Программа подойдет тебе, если необходим тайм-аут после массированной атаки на ноги. Потрать время «отдыха» на то, чтобы развить мышцы трапеции, пекторальных и дельт.
Если ты относишься к касте «везунчиков», у которых низ тела более податлив, чем верх, в твоих интересах поскорее обратится к целевому тренингу слабых звеньев, которые и являются целью настоящего комплекса. Таким образом, ты быстро «подровняешь» мышечную композицию и уравновесишь «тяжелый низ».
Если травма не дает тебе полноценно работать с «низами», не «сачкуй» — сосредоточься на поддержании формы верхней части корпуса.
Принцип сочетаемости рабочих «единиц»
В логичности анатомического тандемы трапеций и дельт сомнений не возникает. Эти две мышечные группы являются закадычными «партнерами» по силовой работе во многих движениях.
Ну а грудные-то почему? Во-первых, грудь — это «святое», во-вторых, накачка «мелких» мышц всегда проходит продуктивнее в партнерстве с крупными группами. В нашем случае именно грудь будет играть роль «большого брата» и создавать неплохое анаболическое «подспорье»
.
К тому же, благодаря внедрению грудных в дневной комплекс, мы можем оставить передние дельты без «специализированного внимания» — они основательно задействованы в большинстве грудных движений.
Выбор упражнений
Упражненческий «компот» каждого тренировочного дня состоит из 5 разномастных упражнений, которые поочередно задействуют все твои «проблемные» зоны. Приоритет по известным причинам отдан комплексным движениям.
В программе упражнения позиционированы в логике их «тяжелости» — в авангарде идут грудные движения, далее предполагается тренировка дельт и трапеции.
Таким образом, концовка дня всегда предсказуема: качаем трапецию гантелями посредством привычных и прицеленных шрагов.

Можешь поинтересоваться, как в нашу складную картинку затесались традиционно «спинонакачательные» тяги гантелей в наклоне?
Все просто: основными ассистентами в данном упражнении выступают трапеции и задние дельты — как раз те мышцы, что интересуют нас первоочередно.
Чтобы угодить нагрузкой в трапецию, примени небольшие технические хитрости: тяни гантель к нижней части груди и максимально подводи «рабочую» лопатку к позвоночнику.
Основные фишки
Объемность подходов в упражнениях, составляющих основу комплекса (по одному на каждую мышечную группу), будет неизменна на протяжении 6-дневного микроцикла.
Повышение нагрузки в остальных упражнениях будет проходить в логике линейной схемы: то есть действуем в «лобовую» и стабильно повышаем объем сетов на несколько повторений. Такая схема работы физически не «истощительна», но выгодна по отдаче.
В изолированных упражнениях на дельты и трапеции предполагается контроль за темпом исполнения движений с акцентом на негативной фазе и пиковом сокращении мышц.

Упражнения на плечи ⋆ ЖЕЛЕЗНЫЙ СПЛИТ
Базовые упражнения
Жим штанги с груди стоя
Исходное положение:
- Возьмите штангу согнув руки к плечам, хватом шире плеч.
- Ноги так же немного шире плеч.
- Выпрямите спину.
- На выдохе выпрямляйте руки, поднимая штангу над головой.
- Не смотрите вверх при поднятии штанги, можно потерять равновесие. Смотрите прямо перед собой.
- На выдохе опускайте штангу к плечам возвращаясь в исходное положение.
Тяга штанги к подбородку
Еще одно базовое упражнение на плечи. В этом упражнении работают передняя и средняя головки дельтовидных мышц.
Исходное положение:
- Встаньте прямо, держа штангу в руках хватом уже плеч.
- На выдохе поднимите штангу к подбородку, работая плечами и поднимая локти.
- Не наклоняйтесь вперед, Держите корпус ровно, взгляд направлен прямо.
- В верхней точке локти должны быть выше плеч. Дельты и трапеция максимально напряжены.
- На вдохе медленно опускайте штангу в исходное положение.
Жим гантелей сидя
Еще одно из основных упражнений для дельтовидных мышц. За счет большей амплитуды движения позволяет больше растянуть дельты в нижней точке а так же использовать поворот кистей для всесторонней проработки мышц.
Исходное положение:
- Сядьте на лавку с упором для спины.
- Ногами упритесь в пол, спину уприте в спинку скамьи но держите прямо, прогнув поясницу.
- Возьмите гантели и согнув руки держите их возле плеч, расставив локти и развернув кисти вперед.
- На выдохе выпрямите руки, выжимая гантели.
- В верхней точке сведите гантели вместе.
- На вдохе возвращайтесь в исходное положение.
Данное упражнение можно так же выполнять стоя. Тогда появляется возможность читинговать, то есть слегка приседать и выпрямляясь ногами помогать себе выжимать гантели что позволит работать с большими весами.
Жим штанги из-за головы сидя
Это очень травмоопасное упражнение. Новичкам его выполнять не рекомендуется. При неправильном выполнении возможны растяжения и вывихи плечевого сустава. Для снижения травмоопасности это упражнение рекомендуется выполнять только в машине Смита, которая позволяет контролировать амплитуду движения.
При отсутствии машины Смита обязательно просите кого либо вас страховать.
Перед выполнением жима из-за головы обязательно сделать разминку и растяжку плечевых суставов.
Исходное положение:
- Сядьте на лавку в тренажер Смита.
- Желательно что бы был упор для спины.
- Ногами прочно упритесь в пол.
- Возьмитесь двумя руками за гриф широким хватом.
- Спину держите прямо, голову не наклоняйте.
- На выдохе опускайте гриф за голову.
- Не опускайте его слишком низко, достаточно до середины затылка.
- На выдохе выпрямляйте руки выжимая штангу в исходное положение.
Изолированные упражнения
Подъемы гантелей перед собой
Изолирующее одностороннее упражнение, направленно воздействующее на переднюю головку дельт.
Исходное положение:
- Встаньте прямо, держа гантели в руках.
- Ноги на ширине плеч.
- Спина прямая, взгляд направлен вперед.
- На выдохе поднимите вперед одну руку.
- Рука должна быть слегка согнута в локте, кисть повернута пальцами вниз.
- Подняв гантель до уровня чуть выше плеч, медленно опускайте руку в исходное положение.
- Затем выполните то же движение другой рукой.
- Не раскачивайте корпус. Совершайте подъем только силой плеча.

Махи гантелями в стороны
Изолирующее упражнение дающее нагрузку на средний пучок дельт.
Исходное положение:
- Возьмите в руки гантели.
- Встаньте прямо, руки с гантелями вдоль туловища, гантели лежат на бедрах.
- Спина прямая, ноги на ширине плеч.
- На выдохе поднимайте руки в стороны.
- Доведя гантели до уровня чуть выше плеч медленно опускайте руки в исходное положение.
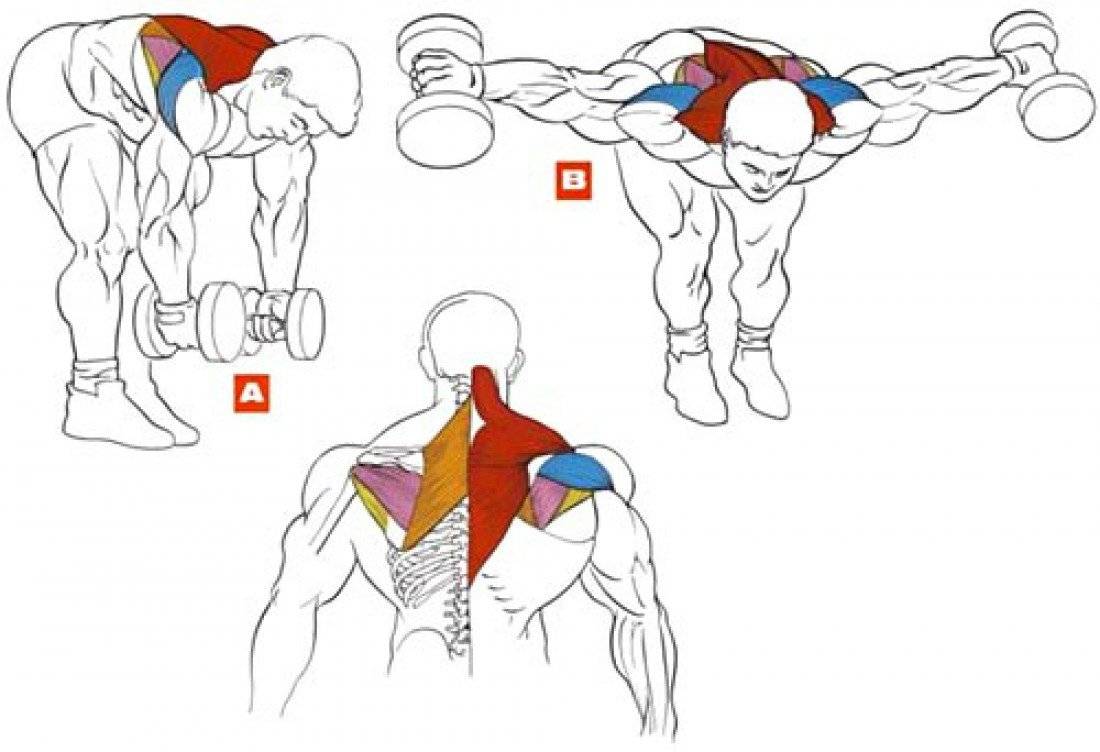
Махи гантелями назад в наклоне
Это упражнение направленно прорабатывает задние пучки дельт.
Исходное положение:
- Возьмите в руки гантели.
- Ноги на ширине плеч или чуть уже.
- Наклонитесь вперед, почти до 90 градусов.
- Спину держите прямо, прогнув ее в пояснице.
- Опустите руки с гантелями вниз, держа их сведенными вместе.
- На выдохе отводите руки назад, слегка согнув их в локтях.
- Старайтесь отвести руки как можно дальше.
- На вдохе возвращайтесь в исходное положение. Для более удобного выполнения и снятия нагрузки с поясницы можно упереться лбом в скамью.
В этой статье я хочу поделиться своим методом наращивания мышечной массы, к которому я пришёл после нескольких лет тренировок. Я перепробовал очень много различных методик, но именно этот стиль дал мне наиболее ощутимые результаты. Я не претендую на абсолютную правильность всего изложенного в этой статье, это всего лишь совет, желание поделиться своим опытом.
Мой метод основывается на разнообразии нагрузки и длительности восстановления между тренировками. Быть может он и не обеспечивает рекордные результаты в пауэрлифтинге, здесь акцент идёт в первую очередь на наращивание мышечной массы. Скажу сразу, этот метод не подходит для новичков, статья адресована не им, а тем, кто уже занимается несколько лет, и, возможно ощущает «застой» росте мышц. Все наверное слышали, что для того чтобы заставить мышцы расти, нужно разнообразие в упражнениях, мышцы быстро адаптируются и эффект от этих упражнений снижается. Мышцы не хотят расти просто так, всячески сопротивляются росту, их нужно постоянно «обманывать». Есть мнение, что для того чтобы добиться больших объёмов нужно постоянно тренироваться с большими весами, это с одной стороны верно, но не совсем, если всё время тренироваться только с максимальными весами, то мышцы «привыкнут» и к такой нагрузке, и в результате рост мышечной массы снизится. Я не претендую на абсолютную правильность всего изложенного в этой статье, это всего лишь совет, желание поделиться своим опытом.
Мой метод основывается на разнообразии нагрузки и длительности восстановления между тренировками. Быть может он и не обеспечивает рекордные результаты в пауэрлифтинге, здесь акцент идёт в первую очередь на наращивание мышечной массы. Скажу сразу, этот метод не подходит для новичков, статья адресована не им, а тем, кто уже занимается несколько лет, и, возможно ощущает «застой» росте мышц. Все наверное слышали, что для того чтобы заставить мышцы расти, нужно разнообразие в упражнениях, мышцы быстро адаптируются и эффект от этих упражнений снижается. Мышцы не хотят расти просто так, всячески сопротивляются росту, их нужно постоянно «обманывать». Есть мнение, что для того чтобы добиться больших объёмов нужно постоянно тренироваться с большими весами, это с одной стороны верно, но не совсем, если всё время тренироваться только с максимальными весами, то мышцы «привыкнут» и к такой нагрузке, и в результате рост мышечной массы снизится. При работе с небольшими весами, я всегда полностью выкладываюсь в каждом подходе, последний повтор выполняю изо всех сил. Отдых между подходами меньше минуты, в то время как при работе с большими весами я отдыхаю иногда по пять минут. Я уверен, что работа с лёгкими весами не менее важна, мышцы иногда от лёгких весов болят даже больше чем при тяжёлых нагрузках, особенно это ощутимо на следующий день после тренировки, а боль в мышцах всегда говорит о их росте. Боль в мышцах на следующий день – это показатель эффективности тренировки. Нужно всегда стремиться к этой боли, я не побоюсь этого слова «ловить кайф» от неё. Ещё один показатель эффективности это желание тренироваться, должен всегда присутствовать тренировочный «аппетит», если его нет – то значит вы что то делаете неправильно, чаще всего желание тренироваться теряется при слишком частых тренировках, когда мышцы не успевают восстановиться, тренироваться в таком состоянии бесполезно – не будет никакого роста мышц, вы только помучаетесь в спортзале. Более того, я считаю, что нет смысла тренироваться даже когда испытываешь незначительную, едва уловимую боль, например в дельтовидных мышцах, а тренировать сегодня нужно ноги. В этом случае лучше пропустить тренировку. Я вообще не имею никакого понятия о том, когда мне в следующий раз идти на тренировку, сколько раз в неделю я тренирую одну группу мышц, я иду в зал только когда полностью восстановился от предыдущей тренировки. Мир не обрушится, если дать себе лишний день отдыха, самой большой ошибкой я считаю тренировку по определённым дням недели, например понедельник-среда-пятница, бывает так что в какой-то день вы загрузили мышцы больше чем обычно, и не успели восстановиться к следующей тренировке, в этом случает ее нужно отложить, понадобиться ещё один день – значит нужно дать ещё один день на восстановление. Отдых-это часть тренировки, мышцы растут именно во время отдыха. А бывает что вроде бы и восстановился, но как то не тянет на тренировку, в этом случае думаю нужно прислушаться к своим внутренним ощущениям и разобраться что является причиной тому – если это элементарная лень, то нужно заставить себя тренироваться, но может быть у вас сегодня просто пониженный тонус, тогда тренировку нужно отложить. Наш организм не подчиняется календарю, поэтому нельзя идти в зал просто потому что сегодня по расписанию тренировка, идти нужно тогда, когда вы чувствуете потребность к тренировке, мышцы сами «просят железа». Многие качаются по схеме понедельник-среда-пятница, или вообще каждый день, я сам долгое время качался и через день, и каждый день, и могу сказать, что это рано или поздно ведёт к перетренированности, потом приходится отдыхать две или три недели. Я думаю что лучше нормально восстанавливаться и качаться постоянно. Хотя, конечно, нельзя дать общую рекомендацию, сколько нужно отдыхать, у каждого свой организм, возможно есть люди, которые способны тренироваться через день или даже каждый день, и успевают восстанавливаться. Нужно уметь слушать свой организм, понимать, что ему необходимо сегодня – отдых или тренировка. Некоторые качаются вдвоём со своим другом. Нужно ли так заниматься спорный вопрос, с одной стороны друг может «подстегнуть» вас, создать дополнительный стимул к тренировке, когда, например, вам лень идти заниматься, друг может подстраховать во время жима лёжа, допустим. Но с другой стороны, большой недостаток тренировок с приятелем, заключается в том, что у каждого организм восстанавливается по разному, возможно вы с приятелем и тренируетесь по одной схеме, что само по себе неправильно – у каждого должна быть своя схема, свои упражнения, но допустим, вы тренируетесь одинаково, а друг ваш возможно восстанавливается быстрее, и когда по расписанию уже тренировка, ваш приятель может быть уже восстановился, и будет звать вас в спортзал, а вы ещё не восстановились, и вам придётся тренироваться в таком состоянии, а это неправильно. А насчёт того чтобы подстраховать во время жима лёжа, то всегда можно попросить кого-нибудь, ведь так? Поэтому я лично не сторонник тренировок с приятелем, по крайней мере, он не заставит меня пойти в спортзал когда я ещё не восстановился. Я стараюсь тренироваться так, чтобы каждая следующая тренировка каждой отдельно взятой группы мышц была не похожа на предыдущую, т.е если я, например, сегодня качал дельты с тяжёлыми весами, то в следующий раз когда буду качать дельты я обязательно буду тренироваться с небольшими весами, если же я проведу две тренировки дельт с тяжёлыми весами, то ощущаю, что тренировочный эффект снижается, т.е я уже не испытывают такой боли в мышцах после тренировки. По этой причине я не думаю, что есть смысл в тренироваться по схеме – несколько недель работы с тяжёлыми весами, потом с лёгкими, так делают профессиональные культуристы в период «сушки», но речь не о них. Обычному же «рядовому качку» тренироваться по такой схеме я не вижу смысла, потому что мышцы очень быстро ко всему привыкают, разнообразие – вот лучший способ заставить мышцы расти. Я сейчас говорю именно о тренировках на массу, а не на увеличение силовых показателей. Некоторые пытаются добиться разнообразия в нагрузке тем, что выполняют на каждой тренировке все имеющиеся на данную группу мышц упражнения, я не считаю такой подход верным по той причине, что так мышцы адаптируются ко всем упражнениям сразу, и на следующей аналогичной тренировке данной группы мышц их уже нечем будет «удивлять». Хотя время от времени и такая «встряска» думаю не повредит. Вместо того чтобы выполнять сразу все упражнения на определённую группу мышц я делаю всего одно, максимум – два упражнения, и это не противоречит принципу разнообразия. Приведу пример того, как я качаю мышцы груди: на одной тренировке я качаю грудь жимом лёжа на горизонтальной скамье, затем разводка с гантелями на той же скамье. На следующей тренировке мышц груди я делаю только отжимания на брусьях с отягощением, на следующей жим гантелей лёжа на горизонтальной скамье и разводка рук с гантелями, на следующей жим горизонтальный на тренажёре, затем на следующей делаю жим лёжа на наклонной скамье, разводку на той же скамье, на следующей одни только кроссоверы, потом на следующей качаю грудь простыми отжиманиями от пола. Всё, цикл закончен, таким образом, учитывая, что я очень долго восстанавливаюсь, у меня между двумя аналогичными тренировками мышц груди проходит где-то около трёх месяцев, естественно, за такой период времени мышцы успевают полностью «забыть» то, что три месяца назад я качал грудь жимом лёжа на горизонтальной, и вновь реагируют на эту тренировку так, как будто я впервые делаю это упражнение. Три месяца может показаться слишком большим интервалом, я поясню почему у меня так получается: у меня в сплите все жимовые движения на одной тренировке, все тяговые на другой, т.е я качаю дельты грудь и трицепс на одной тренировке, а спину и бицепс на другой. Ноги я качаю отдельно. Я считаю, что после двух-трёх лет базовых тренировок нужно тренироваться с учётом своих сильных и слабых сторон, у меня, например, грудь всегда была сильной стороной, а дельты-слабой. По этой причине я дельты всегда качаю в первую очередь, по принципу приоритета, затем грудь, потом трицепс, который у меня развит достаточно неплохо. Из-за того, что грудь у меня развита лучше чем остальные группы, я качаю её через раз, т.е на одной тренировке дельты-грудь-трицепс, на следующей жимовой тренировке дельты-трицепс. Такой подход даёт мне возможность больше «бомбить» дельты. Дельты предпочитаю качать с гантелями, иногда с гирей, считаю, что штанга для дельт не очень подходит, хотя иногда для разнообразия работаю и со штангой. Насчёт бицепса разговор отдельный. Это тоже моя слабая сторона, бицепс это вообще трудно поддающаяся тренировке группа мышц, он моментально адаптируется к нагрузкам, и очень трудно тренироваться так, чтобы ощущать боль в бицепсах после каждой тренировки. Я нашёл выход из положения: на одной тренировки бицепс я качаю с небольшими весами, каждый подход предельный, т.е я не в состоянии выполнить больше ни одного повторения. В следующий раз я качаю бицепс с большими весами, а на следующей тренировке я вместо бицепса усиленно качаю широчайшие спины, т.к это не моя слабая сторона, на предыдущих тренировках я им уделял меньше внимания, или даже вообще пропускал тренировку верха спины, качая только бицепс и выполняя становую тягу. Так вот, в этот раз я качаю только крылья или подтягиваниями на турнике с отягощением, или на тренажёре «гребля», таким образом на бицепс всё равно идёт косвенная нагрузка, и к следующей тренировке бицепса он уже успевает «отвыкнуть» от прямой нагрузки. Я снова начинаю цикл на бицепс и вновь ощущаю результат в виде боли в бицепсах на следующий день после тренировки. Ноги я качаю приседаниями со штангой на одной тренировке, на следующей тренировке ног я качаю их тренажёре для жима ногами. Кроме того, для разнообразия в качестве альтернативной нагрузки на ноги время от времени я делаю пробежки на короткие дистанции, пару раз выполняя ускорения метров 40-50. Многие качки не любят бег, считается, что он сжигает «массу». Это в корне неверно: конечно, если бегать через день марафонскую дистанцию в 40 километров, то «масса» наверняка упадёт, но бег на 5 километров с ускорением раз в две-три недели не только не повредит, но и способствует наращиванию мышечной массы. Во время тренировок с отягощением мышцы накачиваются кровью, которая поставляет в мышцы кислород и питательные вещества, что способствует их росту, но это накачка другого рода, при тренировке с железом накачивается всегда только определённая часть тела, в то время как при беге кислородом насыщается весь организм сразу. Получается как бы «общая накачка», и таким образом вы убиваете сразу двух зайцев: даёте хорошую встряску организму в виде необычной нагрузки, что всегда стимулирует рост массы, плюс прекрасный способ прокачать ноги, неожиданная нагрузка на квадрицепсы, что особенно приятно, не на задницу, как от приседаний. Вы когда-нибудь обращали внимания на спринтеров? Такое ощущение, что многие из них могут прямо с беговой дорожки идти на соревнования по бодибилдингу. В общем то у меня ноги развиты лучше всего, они опережают в развитии верх, но иногда когда я хочу «разбомбить» ноги я делаю одно из двух: или сначала иду в спортзал и забиваю ноги жимом на тренажёре с весом позволяющим сделать 20-30 повторов, затем быстро одеваюсь и на «убитых» ногах делаю пробежку до упада, или сначала делаю пробежку, прибегаю в зал, и «забиваю» ноги на том же тренажёре. В таких случаях я делаю всего два подхода, во втором, последнем я выкладываюсь полностью, так что не могу больше выжать ни раза, затем вываливаюсь из тренажёра, и делаю просто обыкновенные приседания вообще без веса, несколько приседаний и я ноги полностью «умирают», я буквально падаю. При этом мне как-то до фонаря недоумённые вопросы качков из спортзала по поводу скоротечности моей тренировки. Тренировка – это всего лишь импульс для роста мышц, и не стоит задерживаться в спортзале после того как вы уже всё сделали, не нужно смотреть на тех, кто качается часами, для роста массы – чем короче тренировка, тем лучше. Разумеется, при условии, что за это время вы успеваете дать мышцам необходимую нагрузку. Я делаю 4-5 подходов на группу мышц не считая разминочного. Таким образом тренировка у меня длится 40 минут максимум. Думаю, что оптимальное количество подходов – не более 6. Хочу сказать несколько слов о тренажёрах. Считается, что наращивать мышечную массу нужно только со штангой и гантелями, в целом это верно, 90 процентов тренировки должны занимать упражнения со свободными отягощениями, но полностью отказываться от тренажёров всё таки думаю не стоит, я считаю что есть незаменимые для роста массы тренажёры, такие как тренажёр гребля для спины, трицепс стоя, жим ногами. Я иногда полностью качаю какую-нибудь группу мышц на тренажёре, когда чувствую, что необходимо какое-то разнообразие. Упражнения на разных людей действуют по-разному, то что нравится одному, может быть не понравится другому, а нельзя добиться успехов в том, что не нравится, всё нужно делать с желанием, отсутствие желания делать то или иное упражнение – верный признак того, что данное упражнение вам не подходит, и нужно качать эту группу мышц другим упражнением. Конечно плохо, если нелюбимым упражнением является становая тяга, или приседания, потому что это – базовые упражнения, от них никуда не денешься если хочешь нарастить массу. Ну и в заключении скажу несколько слов о питании. Я не буду писать о спортивном питании, на эту тему можно найти гору информации, хочу сказать лишь о том, что не стоит делать из еды культа, переедать, как делают многие качки, я сам раньше так делал, ел через силу, думал, что это помогает нарастить мускулы. В результате я прибавлял в весе, но за счёт жира, причём ладно бы он рос на руках, груди, плечах, так ведь нет, он растёт всегда там, где не нужно – на животе и на заднице. Переедание не способствует наращиванию мышечной массы, нужно есть столько, сколько необходимо организму, нет смысла давиться, всё лишнее всё равно уйдёт в жир. Организм сам подскажет сколько ему нужно, вы наверное замечали, когда мышцы болят после тренировки, аппетит возрастает. Ну, само собой, в пище должно быть достаточно белков, витаминов и калорий. Вот пожалуй и всё, что я хотел сказать. Буду рад если кто-то извлечёт для себя пользу от этой информации. При работе с небольшими весами, я всегда полностью выкладываюсь в каждом подходе, последний повтор выполняю изо всех сил. Отдых между подходами меньше минуты, в то время как при работе с большими весами я отдыхаю иногда по пять минут. Я уверен, что работа с лёгкими весами не менее важна, мышцы иногда от лёгких весов болят даже больше чем при тяжёлых нагрузках, особенно это ощутимо на следующий день после тренировки, а боль в мышцах всегда говорит о их росте. Боль в мышцах на следующий день – это показатель эффективности тренировки. Нужно всегда стремиться к этой боли, я не побоюсь этого слова «ловить кайф» от неё. Ещё один показатель эффективности это желание тренироваться, должен всегда присутствовать тренировочный «аппетит», если его нет – то значит вы что то делаете неправильно, чаще всего желание тренироваться теряется при слишком частых тренировках, когда мышцы не успевают восстановиться, тренироваться в таком состоянии бесполезно – не будет никакого роста мышц, вы только помучаетесь в спортзале. Более того, я считаю, что нет смысла тренироваться даже когда испытываешь незначительную, едва уловимую боль, например в дельтовидных мышцах, а тренировать сегодня нужно ноги. В этом случае лучше пропустить тренировку. Я вообще не имею никакого понятия о том, когда мне в следующий раз идти на тренировку, сколько раз в неделю я тренирую одну группу мышц, я иду в зал только когда полностью восстановился от предыдущей тренировки. Мир не обрушится, если дать себе лишний день отдыха, самой большой ошибкой я считаю тренировку по определённым дням недели, например понедельник-среда-пятница, бывает так что в какой-то день вы загрузили мышцы больше чем обычно, и не успели восстановиться к следующей тренировке, в этом случает ее нужно отложить, понадобиться ещё один день – значит нужно дать ещё один день на восстановление. Отдых-это часть тренировки, мышцы растут именно во время отдыха. А бывает что вроде бы и восстановился, но как то не тянет на тренировку, в этом случае думаю нужно прислушаться к своим внутренним ощущениям и разобраться что является причиной тому – если это элементарная лень, то нужно заставить себя тренироваться, но может быть у вас сегодня просто пониженный тонус, тогда тренировку нужно отложить. Наш организм не подчиняется календарю, поэтому нельзя идти в зал просто потому что сегодня по расписанию тренировка, идти нужно тогда, когда вы чувствуете потребность к тренировке, мышцы сами «просят железа». Многие качаются по схеме понедельник-среда-пятница, или вообще каждый день, я сам долгое время качался и через день, и каждый день, и могу сказать, что это рано или поздно ведёт к перетренированности, потом приходится отдыхать две или три недели. Я думаю что лучше нормально восстанавливаться и качаться постоянно. Хотя, конечно, нельзя дать общую рекомендацию, сколько нужно отдыхать, у каждого свой организм, возможно есть люди, которые способны тренироваться через день или даже каждый день, и успевают восстанавливаться. Нужно уметь слушать свой организм, понимать, что ему необходимо сегодня – отдых или тренировка. Некоторые качаются вдвоём со своим другом. Нужно ли так заниматься спорный вопрос, с одной стороны друг может «подстегнуть» вас, создать дополнительный стимул к тренировке, когда, например, вам лень идти заниматься, друг может подстраховать во время жима лёжа, допустим. Но с другой стороны, большой недостаток тренировок с приятелем, заключается в том, что у каждого организм восстанавливается по разному, возможно вы с приятелем и тренируетесь по одной схеме, что само по себе неправильно – у каждого должна быть своя схема, свои упражнения, но допустим, вы тренируетесь одинаково, а друг ваш возможно восстанавливается быстрее, и когда по расписанию уже тренировка, ваш приятель может быть уже восстановился, и будет звать вас в спортзал, а вы ещё не восстановились, и вам придётся тренироваться в таком состоянии, а это неправильно. А насчёт того чтобы подстраховать во время жима лёжа, то всегда можно попросить кого-нибудь, ведь так? Поэтому я лично не сторонник тренировок с приятелем, по крайней мере, он не заставит меня пойти в спортзал когда я ещё не восстановился. Я стараюсь тренироваться так, чтобы каждая следующая тренировка каждой отдельно взятой группы мышц была не похожа на предыдущую, т.е если я, например, сегодня качал дельты с тяжёлыми весами, то в следующий раз когда буду качать дельты я обязательно буду тренироваться с небольшими весами, если же я проведу две тренировки дельт с тяжёлыми весами, то ощущаю, что тренировочный эффект снижается, т.е я уже не испытывают такой боли в мышцах после тренировки. По этой причине я не думаю, что есть смысл в тренироваться по схеме – несколько недель работы с тяжёлыми весами, потом с лёгкими, так делают профессиональные культуристы в период «сушки», но речь не о них. Обычному же «рядовому качку» тренироваться по такой схеме я не вижу смысла, потому что мышцы очень быстро ко всему привыкают, разнообразие – вот лучший способ заставить мышцы расти. Я сейчас говорю именно о тренировках на массу, а не на увеличение силовых показателей. Некоторые пытаются добиться разнообразия в нагрузке тем, что выполняют на каждой тренировке все имеющиеся на данную группу мышц упражнения, я не считаю такой подход верным по той причине, что так мышцы адаптируются ко всем упражнениям сразу, и на следующей аналогичной тренировке данной группы мышц их уже нечем будет «удивлять». Хотя время от времени и такая «встряска» думаю не повредит. Вместо того чтобы выполнять сразу все упражнения на определённую группу мышц я делаю всего одно, максимум – два упражнения, и это не противоречит принципу разнообразия. Приведу пример того, как я качаю мышцы груди: на одной тренировке я качаю грудь жимом лёжа на горизонтальной скамье, затем разводка с гантелями на той же скамье. На следующей тренировке мышц груди я делаю только отжимания на брусьях с отягощением, на следующей жим гантелей лёжа на горизонтальной скамье и разводка рук с гантелями, на следующей жим горизонтальный на тренажёре, затем на следующей делаю жим лёжа на наклонной скамье, разводку на той же скамье, на следующей одни только кроссоверы, потом на следующей качаю грудь простыми отжиманиями от пола. Всё, цикл закончен, таким образом, учитывая, что я очень долго восстанавливаюсь, у меня между двумя аналогичными тренировками мышц груди проходит где-то около трёх месяцев, естественно, за такой период времени мышцы успевают полностью «забыть» то, что три месяца назад я качал грудь жимом лёжа на горизонтальной, и вновь реагируют на эту тренировку так, как будто я впервые делаю это упражнение. Три месяца может показаться слишком большим интервалом, я поясню почему у меня так получается: у меня в сплите все жимовые движения на одной тренировке, все тяговые на другой, т.е я качаю дельты грудь и трицепс на одной тренировке, а спину и бицепс на другой. Ноги я качаю отдельно. Я считаю, что после двух-трёх лет базовых тренировок нужно тренироваться с учётом своих сильных и слабых сторон, у меня, например, грудь всегда была сильной стороной, а дельты-слабой. По этой причине я дельты всегда качаю в первую очередь, по принципу приоритета, затем грудь, потом трицепс, который у меня развит достаточно неплохо. Из-за того, что грудь у меня развита лучше чем остальные группы, я качаю её через раз, т.е на одной тренировке дельты-грудь-трицепс, на следующей жимовой тренировке дельты-трицепс. Такой подход даёт мне возможность больше «бомбить» дельты. Дельты предпочитаю качать с гантелями, иногда с гирей, считаю, что штанга для дельт не очень подходит, хотя иногда для разнообразия работаю и со штангой. Насчёт бицепса разговор отдельный. Это тоже моя слабая сторона, бицепс это вообще трудно поддающаяся тренировке группа мышц, он моментально адаптируется к нагрузкам, и очень трудно тренироваться так, чтобы ощущать боль в бицепсах после каждой тренировки. Я нашёл выход из положения: на одной тренировки бицепс я качаю с небольшими весами, каждый подход предельный, т.е я не в состоянии выполнить больше ни одного повторения. В следующий раз я качаю бицепс с большими весами, а на следующей тренировке я вместо бицепса усиленно качаю широчайшие спины, т.к это не моя слабая сторона, на предыдущих тренировках я им уделял меньше внимания, или даже вообще пропускал тренировку верха спины, качая только бицепс и выполняя становую тягу. Так вот, в этот раз я качаю только крылья или подтягиваниями на турнике с отягощением, или на тренажёре «гребля», таким образом на бицепс всё равно идёт косвенная нагрузка, и к следующей тренировке бицепса он уже успевает «отвыкнуть» от прямой нагрузки. Я снова начинаю цикл на бицепс и вновь ощущаю результат в виде боли в бицепсах на следующий день после тренировки. Ноги я качаю приседаниями со штангой на одной тренировке, на следующей тренировке ног я качаю их тренажёре для жима ногами. Кроме того, для разнообразия в качестве альтернативной нагрузки на ноги время от времени я делаю пробежки на короткие дистанции, пару раз выполняя ускорения метров 40-50. Многие качки не любят бег, считается, что он сжигает «массу». Это в корне неверно: конечно, если бегать через день марафонскую дистанцию в 40 километров, то «масса» наверняка упадёт, но бег на 5 километров с ускорением раз в две-три недели не только не повредит, но и способствует наращиванию мышечной массы. Во время тренировок с отягощением мышцы накачиваются кровью, которая поставляет в мышцы кислород и питательные вещества, что способствует их росту, но это накачка другого рода, при тренировке с железом накачивается всегда только определённая часть тела, в то время как при беге кислородом насыщается весь организм сразу. Получается как бы «общая накачка», и таким образом вы убиваете сразу двух зайцев: даёте хорошую встряску организму в виде необычной нагрузки, что всегда стимулирует рост массы, плюс прекрасный способ прокачать ноги, неожиданная нагрузка на квадрицепсы, что особенно приятно, не на задницу, как от приседаний. Вы когда-нибудь обращали внимания на спринтеров? Такое ощущение, что многие из них могут прямо с беговой дорожки идти на соревнования по бодибилдингу. В общем то у меня ноги развиты лучше всего, они опережают в развитии верх, но иногда когда я хочу «разбомбить» ноги я делаю одно из двух: или сначала иду в спортзал и забиваю ноги жимом на тренажёре с весом позволяющим сделать 20-30 повторов, затем быстро одеваюсь и на «убитых» ногах делаю пробежку до упада, или сначала делаю пробежку, прибегаю в зал, и «забиваю» ноги на том же тренажёре. В таких случаях я делаю всего два подхода, во втором, последнем я выкладываюсь полностью, так что не могу больше выжать ни раза, затем вываливаюсь из тренажёра, и делаю просто обыкновенные приседания вообще без веса, несколько приседаний и я ноги полностью «умирают», я буквально падаю. При этом мне как-то до фонаря недоумённые вопросы качков из спортзала по поводу скоротечности моей тренировки. Тренировка – это всего лишь импульс для роста мышц, и не стоит задерживаться в спортзале после того как вы уже всё сделали, не нужно смотреть на тех, кто качается часами, для роста массы – чем короче тренировка, тем лучше. Разумеется, при условии, что за это время вы успеваете дать мышцам необходимую нагрузку. Я делаю 4-5 подходов на группу мышц не считая разминочного. Таким образом тренировка у меня длится 40 минут максимум. Думаю, что оптимальное количество подходов – не более 6. Хочу сказать несколько слов о тренажёрах. Считается, что наращивать мышечную массу нужно только со штангой и гантелями, в целом это верно, 90 процентов тренировки должны занимать упражнения со свободными отягощениями, но полностью отказываться от тренажёров всё таки думаю не стоит, я считаю что есть незаменимые для роста массы тренажёры, такие как тренажёр гребля для спины, трицепс стоя, жим ногами. Я иногда полностью качаю какую-нибудь группу мышц на тренажёре, когда чувствую, что необходимо какое-то разнообразие. Упражнения на разных людей действуют по-разному, то что нравится одному, может быть не понравится другому, а нельзя добиться успехов в том, что не нравится, всё нужно делать с желанием, отсутствие желания делать то или иное упражнение – верный признак того, что данное упражнение вам не подходит, и нужно качать эту группу мышц другим упражнением. Конечно плохо, если нелюбимым упражнением является становая тяга, или приседания, потому что это – базовые упражнения, от них никуда не денешься если хочешь нарастить массу. Ну и в заключении скажу несколько слов о питании. Я не буду писать о спортивном питании, на эту тему можно найти гору информации, хочу сказать лишь о том, что не стоит делать из еды культа, переедать, как делают многие качки, я сам раньше так делал, ел через силу, думал, что это помогает нарастить мускулы. В результате я прибавлял в весе, но за счёт жира, причём ладно бы он рос на руках, груди, плечах, так ведь нет, он растёт всегда там, где не нужно – на животе и на заднице. Переедание не способствует наращиванию мышечной массы, нужно есть столько, сколько необходимо организму, нет смысла давиться, всё лишнее всё равно уйдёт в жир. Организм сам подскажет сколько ему нужно, вы наверное замечали, когда мышцы болят после тренировки, аппетит возрастает. Ну, само собой, в пище должно быть достаточно белков, витаминов и калорий. Вот пожалуй и всё, что я хотел сказать. Буду рад если кто-то извлечёт для себя пользу от этой информации. |
Качаем плечи — самые эффективные упражнения : Анаболические стероиды
Я не буду вам говорить за чем нужно качать плечи и что именно широкие и мощные плечи показатель атлетической фигуры, подчёркивающие её мощь. Хочу лишь отметить, что у многих продвинутых бодибилдеров хорошо развиты грудь и мощная спина, впечатляющие бицепсы и узкая талия, а вот плечи, просто чёрт знает что. Плечи могут быть и широкими, но без мощных бугристых дельтовидных мышц, да ещё на фоне всего остального, они выглядят как женские.
Даже у тех атлетов, у которых имеется что-то похожее на передние дельты плеч, как правило напрочь отсутствуют задние дельты. А ведь именно задняя часть дельтовидных мышц и создаёт образ мощных и накачанных плеч. Итак, довольно разговоров, качаем плечи, но не абы как, а самыми эффективными упражнениями. Плечи, их дельтоиды, сравнительно маленькие мышцы и участвуют практически при любой тренировочной работе. Качаете вы руки — плечи нагружаются, качаете грудь, жмёте лёжа или качаете спину — везде часть нагрузки приходится на плечи. Поэтому плечи, дельтовидные мышцы, очень легко перегрузить и как следствие, отсутствие всякого прогресса и даже уменьшение массы дельтовидных. В силу этого, примите, как аксиому: если качаем плечи на массу, то используем только 4 самых эффективных базовых упражнения. Забудьте о всяких тягах на дельтовидные и разводках (кроме задней), как о страшном сне. Только тяжёлые жимовые упражнения строят по-настоящему мощные богатырские плечи. Все остальные упражнения не имеют к росту массы этой группы ни какого отношения.
Качать плечи необходимо отдельно. То есть качаем плечи на специально отведённых для них тренировках. Вы можете сочетать в эти дни работу и над другими мышцами, если они не требуют жимовых упражнений, но плечи качаем первыми. В этом вся соль.
Лучший вариант — качать плечи 2 раза в неделю, но если у вас большой объём жимовых упражнений на другие группы мышц (грудь и трицепсы), то можно и один раз в неделю, если вы тренируетесь 3 раза в неделю. Если у вас есть возможность тренироваться 4-6 раз в неделю, то лучше проводить специализацию на дельтовидные мышцы и качать плечи 2 раза в неделю.
Качаем плечи так:
Тренировка 1:1) Жим штанги от груди, сидя на скамье с вертикальной спинкой. Хват средний, чуть шире плеч. Спина опирается на спинку скамьи (обязательное условие). 3 рабочих подхода по 8 раз с максимальной интенсивностью. Восьмой раз вы должны выжимать аш с криком и борьбой за каждый миллиметр на мёртвой точке. Очень важен грамотный напарник, который вовремя увидит, что штанга уже не сдвинется с мёртвой точки и чуть-чуть даст лёгкий импульс, дабы вы смогли завершить подход с ещё большей, запредельной интенсивностью. Опускать вниз штангу нужно после этого максимально медленно, на сколько хватит остатка сил. По такому принципу качаем плечи во всех жимовых упражнениях, это называется максимальной запредельной интенсивностью. В следующий раз, когда будем делать это упражнение, обязательно попробуйте увеличить вес снаряда на 2,5 кг. Отдыхаем между подходами минуты 3. Обязательна хорошая разминка. Плечи легко травмировать!
Жим гантелей сидя на той же скамье. Принципы те же. 3Х8-10
Жим сидя на скамье в машине Смитта, штанга на плечах, за головой, хват широкий, 3х8-10. Выложите все оставшиеся силы.
Разводка гантелей в наклоне, на заднии дельты. В исходном положении гантели перед собой, почти касаются друг друга, хват тыльной стороной ладоней вперёд. Разводим гантели максимально встороны-вверх, руки в локтях под 9о градусов. 4 подхода по 8-10 раз, возвращаем в ип медленно.
Тренировка 2: 1) Жим штанги из-за головы, сидя на всё той же скамье с опорой на спинку. Хват широкий, 3х8, принципы те же.
То же самое упражнение, которым качали плечи на первой тренировке под №2.
Жим штанги с груди, сидя на скамье в машине Смитта 3х8-10
Разводка гантелей в наклоне, для задних дельтоидов 4х8-10.
Качаем широкие плечи | SUPER-MENS.RU
Все люди делятся на две категории. Одним всё по плечу, другим всё по …
Мне кажется, гораздо лучше относится к первой, а жизненную позицию подтвердить шириной этого самого плеча.
Как устроены наши плечи (дельтовидные мышцы)
Буквально пару слов о строении дельтовидных мышц, если кто не догадался, это и есть те самые плечи. Дельтовидная мышца состоит из пучков: передний, средний и задний, на каждый из них есть упражнения, есть комплексы, в которых задействованы все группы.
{module 276}Чтобы ваши плечи стали широкими, нужно прорабатывать каждый из пучков. Теоретически вроде всё ясно, посмотрим, как воплотить мечту в жизнь.
Как сделать красивые плечи
Каждый день мы что-то поднимаем, опускаем, работает наша мышечная система. Так устроено тело, что из трёх пучков, мы больше задействуем средний, соответственно, он развит больше. Когда посетитель тренажерного зала работает над дельтой, то часто происходит так, что он ограничивается одним-двумя упражнениями. Однако если это вполне приемлемо, на первой стадии тренировок, то потом требует обязательной корректировки.
{module 277}Все забывают про заднюю дельту
Самая распространенная ошибка, которую можно наблюдать, это отсутствие нагрузки на заднюю дельту. Не знаю почему, но часто о ней просто забывают. Впрочем, это и не удивительно, когда каждый третий занимающийся в качалке уверен, что «плечо, это единая мышца». С этого пучка и начну, пойдем, так сказать, с заду, на перед.
Задний пучок придаёт плечу правильную красивую округлую форму.
Вообще широкие плечи, это результат долгой кропотливой работы. Они выглядят привлекательно и эстетично, но набрать за счет работы с дельтой массу невозможно, это маленькие мышцы.
Упражнение здесь простое: махи гантелями по бокам, но есть одно условие.
Один парень в тренажерке называет это упражнение:
— Бодаем скамейку!
Так оно и есть, туловище должно быть в наклоне, практически параллельно полу, естественно в такой позиции нужна дополнительная точка опоры – упираемся головой и поехали.
Нам нужны широкие плечи
Визуальное расширение плеч, это в большей степени задача средней дельты. Для неё есть масса упражнений на выбор, это вам и тяга штанги к подбородку и разведение рук с гантелями по бокам с прямым корпусом и жим из-за головы. Разумеется, задействованы и другие пучки, но основная работа идет всё же на средний.
Часто, именно для плеч предлагают комбинированное упражнение.
Сидя или стоя вы делаете жим штанги из-за головы, при этом один раз опускаете её на грудь, другой за голову, и так делаете упражнение.
Среди плюсов называют работу всех пучков.
Не в восторге от такого решения.
1. Пресловутый задний пучок задействован минимально.
2. Во время упражнения вы как эквилибрист балансируете то в одну, то в другую сторону, не забывайте, что у вас позвоночник находится под нагрузкой и это ёрзание взад вперед может печально кончится.
3. То, что напрягает лично меня, вы всё время меняете траекторию и вынуждены проносить над головой немалый вес. Не способствует технике безопасности.
4. Так как вы комбинируете нагрузку, то обязательно будете делать одно из связанных вместе упражнений менее качественно. Здесь всё просто, одни мышцы сильнее, другие слабее, вы уже не в состоянии сделать жим от груди (в качестве примера), а вот из-за головы, сделали бы еще пару повторений, но так вы чередуете движения, этого не произойдет. Жим с груди не пошел и вы остановились. Проще разделить эти упражнения и делать их по отдельности, хотя, на вкус и цвет, как говорится.
Как проработать передний пучок дельт
Для целевой работы подойдут махи гантелью перед собой. Причем вы можете делать упражнение каждой рукой по отдельности, а можете взять гантель двумя руками. Попробуйте и так и так, почувствуйте, как удобно именно вам.
Маленький совет по технике. Сделайте один подход так, чтобы видеть себя сбоку. Спина должна быть идеально прямая, а движение совершаться только руками. Часто в погоне за весом, гантель начинают, раскачивать всем корпусом или гнут спину «помогая» себе, это не наш метод!
Не бойтесь нагружать плечи
Как я уже сказал, дельтовидная мышца достаточно маленькая, соответственно быстро восстанавливается и для роста требует увеличение количества подходов, по сравнению с крупными мышцами. По плану тренировки, можно комбинировать плечи с ногами или спиной. Т.е. с той группой мышц, где идет более мощная нагрузка на весь организм.
Что делать новичкам
Все эти изыски должны вас заинтересовать не раньше, чем через полгода тренировок, а то и позже. Если вы делаете первые шаги, прочитали, запомнили и пошли делать программу для начинающих. Знания полезные, но пригодятся только в будущем.
Живите широко, с широкой душой, широкими плечами и широкой улыбкой! Удачных тренировок!
avtor: тренер по бодибилдингу Максим Кандратюк, для сайта super-mens.ru
Малое количество повторений Качаемся без сушкиУпражнения на плечи с гантелями. ТОП лучших.
Приведенные в статье 7 программ тренировок на плечи помогут вам разобраться какими упражнениями можно накачать дельты. Каждый комплекс упражнений на плечи поможет прокачать весь плечевой пояс и проработать все пучки дельтовидных мышц и отдельно, средний, передний и задний.
- Как качать плечи
- Как накачать большие плечи: тренировка на массу Программа тренировок плеч на массу
- Тренировка плеч на рельеф
- Тренировочная программа
- Комплекс упражнений на средние дельты
- Комплекс упражнений на задние дельты
- Комплекс упражнений на передние дельты
Как качать плечи
Не существует 2 людей, которые бы тренировались совершенно одинаково и накачали бы большие плечи . Каждый атлет выполняет различную последовательность упражнений, количество подходов, использует разные веса и длительность периодов отдыха. Эта статья поможет разобраться, как накачать плечи в тренажерном зале или в домашних условиях каждому.
Индивидуальность – неотъемлемое свойство человека, и в этом нет ничего плохого, но это накладываете ограничения на способы как накачать дельтовидные мышцы отдельному человеку. Некоторые принципы и подходы делают тренировок лучше, особенно когда дело касается вопроса как быстро накачать плечи. Поэтому я составил 7 действительно эффективных комплексов упражнений на плечи , каждый из которых расскажет как качать плечи на рельеф, ширину и массу.
Обратите внимание на то, что порядок упражнений, веса, количество повторений и объем можно изменять, чтобы подобрать лучший именно для вас способ, как накачать дельтовидные мышцы. Как только вы выберете подходящую для себя тренировку, следуйте ей в течение 4-8 недель, а затем вернитесь к вашей обычной или же попробуйте другую из этого списка.
Примечания:
- Ниже основном речь пойдет о том, как накачать плечи в тренажерном зале, но некоторые из программ хорошо подойдут для тренировок дома.
- Приведенные комплексы упражнений не включают в себя разминку. Разминаясь, выполняйте столько повторений, сколько вам нужно, но никогда не достигайте мышечного отказа.
- Выберите вес, который позволит вам достичь мышечного отказа в предписанном количестве повторений. Это будет правильная прокачка плеч, чтобы вовлечь в работу максимальное количество волокон мышц дельт и добиться мышечного роста.
- Если вы тренируетесь с партнером, сделайте несколько форсированных повторений в самом тяжелом подходе в жимах над головой. Если нет, то в последнем подходе каждого упражнения выполните дропсет, снижая вес примерно на 25% при достижении мышечного отказа. Всего его нужно достичь 2 раза.
Как накачать большие плечи: тренировка на массу
Цель: наращивание всех пучков дельт
Самый эффективный метод как накачать плечи в тренажерном зале — это работать с большими весами, но нужно правильно подойти к тренировкам, чтобы не получить травм. Для этого необходимо хорошо разминаться и соблюдать технику выполнения упражнений.
Чтобы нарастить массу плечей, всегда начинайте тренировку с самых сложных упражнений (в данном случае жимы над головой), которые позволяют поднять больший вес. Затем выполняйте односуставные упражнения на каждый из трех пучков дельтовидных мышц: передний, средний и задний. Это позволит создать основу для работы по наращиванию мышечной массы, если вы будете сохранять общий объем тренировок.
Когда мы качаем плечи можно усложнить тренировки несколькими способами. В жиме над головой начните с использования гантелей, которые, как известно, сложнее балансировать и которые позволяют двигаться в большем диапазоне движения, чем штанга. Вы также будете выполнять обратную пирамиду, поскольку она дает возможность сделать больше подходов с мышечным отказом. В первых 2-х подходах вы будете использовать довольно тяжелый вес в низком диапазоне повторений (6), чтобы нарастить силу. По мере накопления усталости в последующих подходах снижайте вес примерно на 5 кг. Последние 2-х самых тяжелых подхода выполняйте со страхующим партнером, чтобы вы могли сохранять технику.
Из-за того что передние дельты проделывают много работы на тренировках груди, а средние принимают на себя всю тяжесть веса при выполнении жима над головой, зачастую задние дельты остаются маленькими и слабыми. В этой тренировке прокачка плеч происходит в условие, когда у вас много сил в запасе. В то же время не бойтесь менять порядок односуставных упражнений, основываясь на своих слабых местах. Если вы считаете, что все пучки дельт у вас развиты пропорционально, просто выполняйте такие упражнения в разном порядке на каждой тренировке.
Программа тренировок плеч на массу
- Жим гантелей над головой — 4 подхода из 6,6,8,10 повторений (2 минуты отдыха)
- Тяга штанги к подбородку — 3 подхода из 8,8,10 повторений (2 минуты отдыха)
- Разведение гантелей в стороны сидя в наклоне — 3 подхода из 8,10,12 повторений (1 минута отдыха)
- Разведение гантелей в стороны стоя — 3 подхода из 8,10,12 повторений (1 минута отдыха)
- Подъем штанги над головой на прямых руках — 3 подхода из 8,10,12 повторений (1 минута отдыха)
Наклонные отжимания
Подняв ноги на скамью, стол или стул, вы сможете продолжать работать с плечами, но с большей легкостью. Чем выше расположены ваши ноги, тем лучше будут работать мышцы плеча.
Как выполнять:
- Примите исходное положение для отжиманий и поднимите ноги на скамью или стул.
- Поставьте ладони на пол чуть шире плеч. Держите корпус и руки прямыми.
- Медленно опускайтесь на пол, ноги при этом остаются поднятыми.
- Удержите положение в нижней точке в течение секунды, и снова оттолкнитесь вверх. Повторите.
- Начните с трех подходов, выполняя как можно большее число повторений. Старайтесь увеличивать количество повторений каждую неделю.
Как накачать рельефные плечи
Цель: дефиниция дельт
Здесь вы узнаете хороший способ как накачать дельты с прорисовкой отдельных волокон. Подъем легких весов в большом количестве повторений уже не считается наилучшим способом достичь дефиниции дельт. Прежде всего, эта тренировка на плечи в зале направлена на стимулирование роста мышц (умеренный вес в умеренном диапазоне повторений). Для увеличения количества сожженных калорий во время и после тренировки (эффект послетренировочного потребления кислорода) используется большой объем в сочетании с суперсетами. Здесь вы будете двигаться быстрее, и ощущать настоящее жжение в мышцах. Теперь вы знаете как накачать мышцы, чтобы они были не только рельефными, а и появилась так называемая «сечка».
Тренировка плеч на рельеф
- Армейский жим сидя — 4 подхода из 8-12 повторений (2 минуты отдыха)
- Разведение гантелей в стороны стоя — Суперсет: 3 подхода из 10-12 повторений (без отдыха)
- Разведение гантелей в стороны сидя в наклоне — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
- Подъем руки перед собой в кроссовере — 3 подхода из 10-12 повторений (без отдыха)
- Тяга к подбородку в кроссовере — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
- Разведение рук в тренажере «бабочка» — 3 подхода из 10-12 повторений (без отдыха)
- Разведение рук в стороны с эспандером — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
Как быстро накачать плечи
Цель: освоение техники и посторонние прочных основ для дальнейших тренировок
Этот комплекс состоит из жима над головой и односуставных упражнений для каждого пучка дельт. Начните с работы в тренажере, чтобы научиться движениям, прежде чем перейти к свободным весам, которые лучше всего сказываются на наращивании мышечной массы.
Начните с небольшой нагрузки и сосредоточьтесь на надлежащей технике. Добавляйте вес лишь тогда, когда сможете полностью контролировать движения.
Тренировочная программа
- Жим над головой в тренажере — 3 подхода из 12 повторений (60-90 секунд для отдыха)
- Разведение гантелей в стороны стоя — 3 подхода из 12 повторений (60-90 секунд для отдыха)
- Подъем руки перед собой в кроссовере — 3 подхода из 12 повторений (60-90 секунд для отдыха)
- Разведение рук в тренажере «бабочка» — 3 подхода из 12 повторений (60-90 секунд для отдыха)
Жим гантелей от плеч
Это упражнение нацелено в основном на плечи, но задействует также трицепсы и мышцы верхней части спины. Это одно из лучших упражнений для развития верхней части тела, так как оно задействует множество мышц, работающих вместе. Чем больше вовлеченных мышц, тем больший вес вы сможете осилить, а это напрямую связано с ростом мышц.
Как выполнять:
- Выберите вес, подходящий для выполнения необходимого количества повторений. Поднимите гантели и опустите их на уровень плеч (или чуть выше). Ладони направлены вперед.
- Поднимите гантели над головой и на вытянутых руках соедините их. Затем опустите гантели на уровень плеч.
- Выполняйте движения в полной амплитуде — вытягивайте руки вверх и опускайте вниз. Повторите, чтобы выполнить необходимое количество повторений и подходов.
- Для начала мы рекомендуем выполнять 3 подхода по 8-12 повторений.
Как накачать средние дельты
Цель: наращивание средних дельтовидных мышц Чтобы плечи стали шире, необходимо развивать средние пучки дельтовидных мышц. Это также позволит талии визуально казаться уже и начать широкий плечевой пояс. Упор в данной программе, естественно, делается на упражнения для средних дельт.
Выполнять вы их будете в начале тренировки, когда энергия находится на самом высоком уровне. Вы можете чередовать эту программу с более сбалансированной тренировкой дельт (например, на массу) в ходе своего недельного сплита.
Комплекс упражнений на средние дельты
- Жим штанги над головой сидя — 4 подхода из 8 повторений (2 минуты для отдыха)
- Тяга штанги к подбородку — 3 подхода из 8-10 повторений (60-90 секунд для отдыха)
- Разведение гантели в сторону одной рукой — 3 подхода из 10 повторений (60-90 секунд для отдыха)
- Разведение гантелей в стороны стоя — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
Разведение гантелей в стороны
Разведение гантелей в стороны прорабатывает латеральную головку дельты. Эта мышца находится на внешней стороне плеча и, будучи полностью развитой, придает плечам трехмерный вид и делает вашу фигуру шире. Упражнение прорабатывает плечо, благодаря тяге гантелей.
Как выполнять:
- Встаньте ровно и возьмите в каждую руку гантели. Руки опущены вниз по бокам, ладони смотрят внутрь.
- Медленно поднимайте гантели вбок, пока руки не окажутся параллельно полу. Задержитесь на секунду в этом положении и медленно опустите руки в исходное положение.
- Держите корпус напряженным и не качайте бедрами, чтобы не сместить вес.
- Постарайтесь выполнить от 2 до 3 подходов по 12-15 повторений.
Как накачать задние дельты
Цель: наращивание задних пучков дельтовидных мышц
Задние пучки дельтовидных мышц зачастую отстают не только у новичков, но и продвинутых бодибилдеров. Проще говоря, они не получают столько стимуляции, сколько передние и средние дельты, которые вовлечены в упражнения для груди и жимы от плеч.
Чтобы развить задние дельты, выполняйте этот комплекс на протяжении 4-8 недель или чередуйте его с более сбалансированной тренировкой плеч.
Комплекс упражнений на задние дельты
- Жим из-за головы — 4 подхода из 8-10 повторений (2 минуты для отдыха)
- Разведение гантелей в стороны сидя в наклоне — 4 подхода из 8 повторений (60-90 секунд для отдыха)
- Разведение рук в кроссовере — 3 подхода из 10 повторений (60-90 секунд для отдыха)
- Разведение рук в тренажере «бабочка» — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
Разведение гантелей в наклоне
Это упражнение прорабатывает задний пучок дельтовидной мышцы. Развитые задние пучки дельт придают плечам полностью развитый округлый вид. Обычно из всех мышц плеча сложнее всего накачать именно эту мышцу. Она прорабатывается при тяговом движении с отягощением, но когда тело при этом не выпрямлено, а наклонено вперед.
Как выполнять:
- Встаньте и наклоните туловище вперед к полу. Держите нижнюю часть спины прямо, чтобы туловище было практически параллельно полу. Держите голову в нейтральном положении и не тяните шею вверх.
- Возьмите в руки гантели. Руки опущены перед собой, ладони обращены друг к другу. Убедитесь, что локти расслаблены и слегка согнуты.
- Задействуйте задние дельты и потяните локти вверх, разведя руки в стороны. Гантели должны оказаться наверху, почти параллельно полу, ладони смотрят вниз.
- Избегайте раскачивания туловища. Держите корпус напряженным на протяжении всего движения.
- Постарайтесь выполнить от 2 до 3 подходов по 12-15 повторений.
Заключение
Чтобы качественно проработать плечи, вам не нужно иметь под рукой какие-то модные снаряды или ходить в тренажерный зал. Если вы включите эти упражнения в свои обычные тренировки, то уже в течение нескольких недель увидите заметные улучшения.
Просто помните о правильной технике и выполняйте полный диапазон движений, чтобы убедиться, что мышцы плеча работают и развиваются правильно. Как только вы усвоите эти упражнения, начинайте усложнять их, постепенно увеличивая количество повторений.
Статьи на нашем сайте представлены только в просветительских и информационных целях. Мы не рекомендуем использовать материалы статей в качестве медицинских рекомендаций. Если вы решили принимать биодобавки или внести основательные изменения в свой рацион, предварительно проконсультируйтесь со специалистом.
Перевод: Фарида Сеидова
Тренируетесь дома? Тогда вам также будет интересна эта статья:
Тренировки
Как накачать передние дельты
Цель: наращивание передних пучков дельтовидных мышц
Если вы часто тренируете грудь, то у вас, вероятно, уже хорошо развиты передние дельты. Ведь они задействуются во всех жимовых упражнениях, особенно при выполнении их в наклоне. Однако относительно слабые передние дельтовидные мышцы могут сдерживать процесс наращивания грудных мышц. Данная тренировка призвана исправить такую ситуацию.
Между тренировкой груди и плеч должно проходить как минимум 48 часов, чтобы мышцы могли полностью восстановиться.
Комплекс упражнений на передние дельты
- Жим над головой сидя — 4 подхода из 8-12 повторений (2 минуты для отдыха)
- Жим Арнольда — 4 подхода из 8-10 повторений (2 минуты для отдыха)
- Подъем гантелей перед собой — 3 подхода из 10 повторений (60-90 секунд для отдыха)
- Подъем руки перед собой в кроссовере — 3 подхода из 10-12 повторений (60-90 секунд для отдыха)
Подъем гантелей перед собой
Это упражнение прежде всего укрепляет дельтовидные мышцы, но также прорабатывает и мышцы верхней части груди (грудные). Выполнять его следует, если эта часть плеча у вас недостаточно развита. При выполнении жима передняя дельта будет работать интенсивно, поэтому будьте осторожны — не перетренируйте эту часть тела.
Как выполнять:
- Встаньте прямо, руки опущены, гантели лежат сбоку на бедрах, ладони смотрят внутрь.
- Поднимайте руки по очереди перед собой так, чтобы гантель находилась параллельно полу, чуть выше линии плеч. Ваша рука должна быть почти прямой, а ладони направлены к полу. Локоть не должен напрягаться. Другая рука вытянута вниз.
- Задержитесь на секунду в верхней точке движения и медленно опустите гантель вниз. Затем повторите движение с другой рукой.
- Постарайтесь выполнить от 2 до 3 подходов по 10-15 повторений с каждой рукой.
Что делать, если не растут плечи
Цель: предварительное утомление дельт
Трицепсы иногда могут выступать ограничивающим фактором в тренировке плеч, особенно в жимах. Если эти мышцы будут всегда «сдавать», прежде чем вы как следует проработаете плечи, вы никогда не сможете довести дельты до мышечного отказа и накачать плечи в многосуставных упражнениях. Практика предварительного утомления призвана исправить такую ситуацию. Это лучший способ как правильно накачать отстающую мышцу. Здесь вы сначала утомляете дельты с помощью односуставных упражнений, а затем выполняете жим над головой, когда трицепсы полны сил. Таким образом, дельты должны достигать отказа прежде, чем это сделает трицепс.
В начале тренировки не подавайтесь соблазну перейти на тяжелые веса, поскольку это даст дополнительную нагрузку на локтевые суставы. Также, если к тому моменту, когда вы подойдете к многосуставному упражнению, вы будете чувствовать сильную усталость, выполните его в тренажере. Так будет немного безопаснее.
Тренировочная программа
- Тяга нижнего блока в сторону одной рукой — 4 подхода из 8-10 повторений (60-90 секунд для отдыха)
- Подъем штанги перед собой на вытянутых руках — 3 подхода из 10 повторений (60-90 секунд для отдыха)
- Разведение рук в тренажере «бабочка» — 3 подхода из 10 повторений (60-90 секунд для отдыха)
- Жим над головой в тренажере — 3 подхода из 8-10 повторений (2 минуты для отдыха)
- Тяга штанги к подбородку — 3 подхода из 10-12 повторений (2 минуты для отдыха)
Источник: https://www.bodybuilding.com/content/shoulder-workouts-for-men-the-7-best-routines-for-bigger-delts.html
границ | Морфодинамика и стратиграфическая архитектура прибрежных дельт шельфа, подверженных постоянным и динамическим воздействиям окружающей среды: лабораторное исследование
Введение
Системы маршрутизации наносов, которые транспортируют отложения от источников к стокам, состоят из связанных сегментов, внутри которых доминирующие морфодинамические процессы остаются довольно постоянными (Allen, 2008; Sømme et al., 2009b; Allen, 2017). Соединение этих сегментов происходит на важных границах земной поверхности, что коренным образом меняет метод, скорость и изменчивость переноса наносов.Здесь я исследую взаимосвязь сегментов переноса наносов, которые происходят на береговой линии, с особым акцентом на дельтах и окружающей их системной среде. В частности, я исследую, как величина и период циклов уровня моря влияют на извлечение массы и осаждение через переход дельта-склон.
Влияние уровня моря или изменения относительного уровня моря (RSL) в регионах со значительным вертикальным движением земной поверхности из-за тектоники, на перенос наносов и стратиграфию — не новый вопрос.Он изучается со времен Гилберта (1890) и активизировался с появлением и изучением сейсмических исследований, проводимых при разведке углеводородов, что послужило стимулом для разработки методов стратиграфии последовательностей и интерпретаций (Vail et al., 1977; Van Wagoner et al. , 1990; Catuneanu et al., 2009). В то время как в основном качественные, наблюдательные и концептуальные подходы, лежащие в основе стратиграфии последовательностей, продолжают учить нас стратиграфии континентальных окраин, подходы, отстаиваемые источником для движения опускания, требуют количественных данных для развития теории, полученных из полевых наблюдений, а также численных и физических экспериментов.
В современных условиях высокого стояния ледника многие дельты отделены от края шельфа широкими континентальными шельфами (Sweet and Blum, 2016). В этих условиях перенос наносов в глубоководные морские глубины, особенно крупных отложений, ограничен. Условия ледникового покрова относятся к периодам времени в истории Земли, когда увеличение и уменьшение размеров ледяных щитов континентального масштаба приводит к колебаниям уровня моря большой амплитуды (≥ 100 м), в то время как парниковый эффект относится к периодам без ледников континентального масштаба и, таким образом, к малым величинам Миланковича. принудительные циклы RSL из-за теплового расширения и сжатия воды.Эффективный перенос наносов в глубоководные районы в настоящее время ограничен реками, которые связаны с подводными каньонами или фьордами (Xu et al., 2004; Vendettuoli et al., 2019). Однако анализ стратиграфии континентальной окраины, выявленной в сейсмических данных обнажений и отражений, показывает, что дельты берегов шельфа эффективно питали склоновые системы во время недавних ледниковых низинных условий, а также в парниковые периоды (Porębski and Steel, 2003; Straub et al., 2012; Sylvester et al. др., 2012). Здесь я представляю подробный набор измерений, полученных в результате физического лабораторного эксперимента по роли RSL в разделении отложений между преимущественно наземными и наземными отложениями.преимущественно морские обстановки в дельтах краев шельфа. В частности, я сосредотачиваюсь на (A) характеристиках цикла уровня моря, необходимых для создания стратиграфических признаков, которые могут быть четко связаны с изменением уровня моря в палео, и (B) времени и величине доставки наносов в регионы за пределами края шельфа.
Мотивацией для данного экспериментального исследования является набор результатов численных экспериментов, в которых исследуется влияние вариаций RSL на распространение наносов по краю шельфа во время ледникового противостояния.тепличные условия Земли (Harris et al., 2016, 2018). Используемая в этих исследованиях численная модель, Dionisos, моделирует перенос наносов в масштабе бассейна длины и геологическом масштабе времени, основанном на диффузионном переносе отложений (Granjeon, 1999). Этот диффузионный алгоритм адекватно воспроизводит модели осадконакопления в длинных пространственных и временных масштабах. Однако на более коротких отрезках времени и в более коротких масштабах морфодиамические процессы, возникающие в результате сочетания топографии, потока и переноса наносов, приводят к богатому стохастическому поведению, которое плохо описывается диффузией (Kim et al., 2014; Паола, 2016; Hajek and Straub, 2017). Отчасти эта стохастичность возникает из-за внутренних процессов в системах маршрутизации наносов, например миграции дюн и каналов и отрывов, которые обычно называют аутогенными процессами.
Даже с диффузным описанием переноса наносов эти модели делают несколько прогнозов, которые бросают вызов общепринятым представлениям о роли изменения RSL в доставке наносов на континентальный склон и за его пределы из-за возникающей автогенной динамики модели.Эта динамика является результатом поперечных градиентов осаждения, которые способствуют случайному образованию каналов и компенсационному наложению лепестков. В частности, я выделяю следующие результаты: (1) Harris et al. (2016) обнаружили тривиально небольшую разницу в кумулятивной доставке глубоководного песка между моделью эволюции континентальной окраины с эвстатической историей, полученной из полевой компиляции, и второй моделью без эвстатических вариаций. В то время как пиковая доставка песка в глубоководные районы действительно происходила во время коротких периодов падения уровня моря в модели переменной устойчивости, аналогичные эпизоды доставки песка происходят в модели постоянной устойчивости, когда транспортные системы иногда достигают и временно блокируются на краю шельфа.(2) Доставка песка в глубоководные районы плохо коррелирует со скоростью или амплитудой падения уровня моря, которое сохраняется независимо от коэффициентов диффузии, используемых в моделях (Harris et al., 2018).
Воодушевленный этими численными наблюдениями, я провел физический эксперимент, чтобы изучить связь дельт с континентальным склоном. В частности, я сосредотачиваюсь на системах с реками, порождающими гиперпикнические потоки вниз по течению от береговой линии. Хотя схема диффузии, используемая в Dionisos, вероятно, производит приглушенную аутогенную динамику в масштабах времени роста и отказа дельта-доли, физические эксперименты известны самоорганизацией и сложным стохастическим поведением (Paola et al., 2009).
Эта работа также мотивирована и основана на теории, которая предсказывает пороговые характеристики циклов уровня моря, необходимые для получения геоморфических и стратиграфических сигналов, которые можно уверенно отличить от аутогенных процессов и продуктов (Li et al., 2016; Yu et al., 2017) . Эта теория была недавно проверена в серии дельтовых экспериментов, но в этих экспериментах отсутствовал уклон вниз по течению от береговой линии и набор батиметрических данных высокого разрешения, оба из которых присутствуют в этом исследовании.
Материалы и методы
Чтобы изучить влияние RSL на разделение отложений между преимущественно наземной и морской средами, я провел физический лабораторный эксперимент в Глубоководном бассейне, расположенном в Лаборатории динамики и стратиграфии отложений Тулейнского университета. Бассейн имеет длину 6 м, ширину 4 м и глубину 2,2 м (рис. 1А). Уровень моря контролируется плотиной, которая гидравлически сообщается с бассейном. Плотина находится на управляемой компьютером вертикальной направляющей, которая позволяет контролировать высоту уровня моря в субмиллиметровом масштабе.Повышение уровня моря отслеживалось и регистрировалось датчиком один раз в минуту, чтобы убедиться, что он соответствует целевым отметкам.
Рисунок 1. Принципиальная схема экспериментальной установки и условия эксперимента. (A) Вид снизу объекта и исходная топографическая форма с отмеченными важными размерами бассейна. Солевую смесь перекачивали из резервуара-резервуара в резервуар с постоянным напором перед смешиванием с осадком в воронке и затем поступлением в резервуар. (B) Характерное перспективное фото экспериментальной поверхности с окрашенным входным потоком для визуализации. (C) Временные ряды уровня моря за вычетом долгосрочного дрейфа, обусловленного псевдо-проседанием и привязанного к уровню моря в начале этапа 1, при этом время топографических сканирований отмечено красными закрашенными кружками.
Первоначальный рельеф бассейна характеризовался падением плоской полки длиной 1,2 м, которая соединялась с участком длиной 3,8 м с уклоном 10%. Первоначальный рельеф не менялся по направлению удара.Хотя более крутой, чем типичные континентальные окраины, 10% -ный наклон был реализован для сопоставления с другими экспериментальными исследованиями, в которых использовалось искаженное масштабирование Фруда, чтобы показать, что увеличение наклона для лабораторных систем помогает лучше воспроизводить процессы в масштабе поля (Cantelli et al., 2011).
Входной поток в бассейн был спроектирован таким образом, чтобы облегчить перенос наносов в глубоководные районы моря гиперпикническими потоками. Поток вводился через резервуар с постоянным напором в виде смеси воды и растворенной соли, что придавало ему 2% -ную избыточную плотность по сравнению с пресной водой бассейна океана.Чтобы сохранить контраст плотности между входящим потоком и океаном, была реализована система для извлечения плотного потока в основании бассейна и замены его пресной водой сверху со скоростью, равной 11-кратному поступлению солевого раствора. Соленость нашей реки и океана может показаться нелогичной, учитывая, что дельты, построенные на стыке суши и океана, имеют пресную воду в своих реках, которые впадают в соленые океаны. Однако этот выбор был сделан для того, чтобы способствовать гиперпикнальному погружению потоков в море и, таким образом, способствовать переносу наносов на континентальный склон.Подобно предыдущим исследованиям, введенную соль можно рассматривать как компонент смыва системы, который не входит в область исследования (Straub et al., 2008; Hamilton et al., 2017). Каждые 3 часа эксперимент приостанавливался, и в бассейн циркулировали пресная вода до тех пор, пока соленость не перестанет быть обнаружена. Входной гидрограф также был разработан с учетом наводнений для облегчения откачки наносов в глубоководные районы. Базовый расход для эксперимента составил 1,54 × 10 -4 м 3 / с. Затопление было вдвое больше, чем базовый поток, и происходило непрерывно в течение 6 минут в течение каждого рабочего часа.Отношение времени базового стока до паводка было установлено таким образом, чтобы имитировать сезонные паводки в дельтах крупных рек. Продолжительность цикла потока у основания затопления устанавливается так, чтобы большое количество циклов происходило за время, необходимое для увеличения, в среднем, глубины одного канала повсюду на вершине дельты, что, как было показано, является важной временной шкалой автогенного воздействия. активности (Wang et al., 2011).
Подача наносов в бассейн контролировалась через компьютерный интерфейс. В условиях базового потока скорость подачи осадка составляла 3.53 × 10 -4 кг / с, и отношение расхода осадка к воде оставалось постоянным для основного потока и условий заводнения при объемном соотношении 1: 1000. Входящая смесь осадка была разработана для имитации более ранних экспериментальных работ (Hoyal and Sheets, 2009; Li et al., 2016) с диаметром частиц от 1 до 1000 мкм со средним значением 67 мкм и преимущественно белым кварцем. Четверть наиболее грубых 23,5% отложений была окрашена в красный цвет, чтобы облегчить визуализацию стратиграфической архитектуры.Небольшое количество коммерчески доступного полимера (New Drill Plus, распространяемого Baker Hughes Inc.) было добавлено к смеси осадка для повышения сцепления осадка, что способствует образованию каналов в экспериментах. Я использовал эту смесь отложений, поскольку более ранние экспериментальные исследования показали, что ее мелкие зерна транспортируются во взвешенном состоянии по самоорганизованным глубоким и относительно стабильным каналам, что привело к перекачке значительных объемов отложений в море (Straub et al., 2015). Комбинация входящего потока солевого раствора, приемного бассейна пресной воды и взвешенных наносов привела к постоянному погружению гиперпикнальных потоков в устьях рек, которые пересекали склон бассейна в виде мутных нижних потоков (рис. 1B).
Эксперимент включал 120-часовой этап наращивания, на котором уровень моря поддерживался постоянным. Основная фаза эксперимента последовала, когда я установил постоянную скорость подъема уровня моря (rSL¯ = 0,25 мм / ч), чтобы имитировать постоянную долгосрочную скорость опускания. Это псевдо-проседание привело к скорости образования наземных аккомодаций, которая соответствовала среднему проценту объемной скорости поступления наносов, захваченных в земной среде, определенному в ходе более ранней экспериментальной работы (Straub et al., 2015). Таким образом, долгосрочное повышение уровня моря вызвало долгосрочную скорость осаждения на вершине дельты, rD¯, которая соответствовала rSL¯. Это позволило сохранить постоянное среднее положение береговой линии с наложенными вариациями из-за автогенных процессов и циклов уровня моря. Основная фаза эксперимента включала три стадии: (1) отсутствие циклов уровня моря, (2) циклы уровня моря с малой амплитудой и коротким периодом и (3) цикл с большой амплитудой и долгим периодом уровня моря (рис. 1C). Этап 1 длился 108 часов и дал стратиграфию с глубиной чуть более двух каналов по всей вершине дельты (максимальная глубина аутогенных каналов составляла ~ 12 мм).
Вслед за Ли и др. (2016), я масштабирую величину и период циклов уровня моря на стадиях 2 и 3 до критических автогенных пространственных и временных масштабов. Величина циклов уровня моря, M RSL , (т.е. разница в высоте от пика цикла до впадины) сравнивается с максимальной глубиной аутогенного канала, H c :
H * = MRSLHC (1)
В то время как период цикла уровня моря, T RSL , сравнивается со средним временем, необходимым для нанесения стратиграфии на глубину одного канала по всей вершине дельты, которая использует долгосрочную скорость отложений на вершине дельты, rD ¯, как:
TC = HCrD¯ (Ван и др., 2011) (2)
Что позволяет:
Т * = TRSLTC (3)
Этап 2 включал четыре последовательных цикла уровня моря, характеризуемых значениями H * = 0,5 и T * = 0,5, и, таким образом, были небольшими по сравнению с аутогенными шкалами. Этап 3 включал один цикл измерения уровня моря, характеризуемый значениями H ∗ = 2 и T ∗ = 2, и, таким образом, был большим по сравнению с аутогенными шкалами.В идеале я бы провел большее количество циклов измерения уровня моря для всех этапов, чтобы можно было исследовать статистическую изменчивость форсингов. Однако результаты аналогичной серии экспериментов по реакции дельт на воздействие уровня моря без наклона системы вниз указывают на быстрое приспособление дельт к их параметрам воздействия на цикл (Yu et al., 2017).
Стоит отметить, что T c не совпадает с временной шкалой отрыва, T A , как первоначально определено Mohrig et al.(2000). T A определяется как глубина каналов системы, деленная на внутриканальную, локальную скорость разложения, измеренную за один цикл отрыва. Скорость разложения, используемая в формулировке T c , является долгосрочной или геологической скоростью (т. Е. Измеряется в достаточно длительных временных масштабах, чтобы скорости сохранялись во времени; Jerolmack and Sadler, 2007). Обычно T c на несколько порядков больше, чем T A .Таким образом, T c не относится к средней периодичности какого-либо одного поверхностного процесса, а скорее оценивает время, необходимое для равномерного распространения отложений по бассейну в результате полного набора аутогенных процессов (Wang et al., 2011).
Входной ток окрашивали дважды в час синим пищевым красителем для облегчения визуализации морфодинамики. Это выполнялось в течение 1 минуты каждого затопления, а затем 30 минут спустя в течение 1 минуты в условиях базового потока. Аэрофотоснимки экспериментальной поверхности собирались в самом конце каждого импульса красителя.Карты всей экспериментальной поверхности собирались каждые 3 часа работы с помощью лазера смещения дальнего действия 1 кГц, подключенного к регистратору данных и размещенного на измерительной тележке, способной перемещаться во всех трех декартовых направлениях. Последовательные топографические карты были собраны на общей сетке с шагом сетки по горизонтали 5 мм в направлениях вниз и между бассейнами. Это позволило получить карты с вертикальным разрешением 0,25 мм. Для стратиграфического анализа топографические карты использовались для построения синтетической стратиграфии путем суммирования всех топографических снимков и вырезания на предмет эрозии.После завершения экспериментального запуска месторождение было рассечено по трем трансектам, ориентированным в основном по наклону, и снято с помощью цифровой камеры для стратиграфической характеристики.
Результаты
Общая системная динамика
Я начинаю с общего описания динамики системы перед количественным анализом распределения масс от источника к поглотителю. Псевдопадение было включено в начале этапа 1 с одним крупным речным руслом, расположенным на левом берегу бассейна.Этот канал и его дельта-выступ значительно выступали за излом шельфа и оставались фиксированными в течение примерно 70 часов до первого крупномасштабного отрыва лепестка (рис. 2, 3). Эта шкала времени стабильности значительно больше, чем шкала времени компенсации для этапа, и больше, чем любое время между отрывами в предыдущем дельтовом эксперименте, в котором использовалась установка, очень похожая на обсуждаемую здесь, но без связанной системы наклона (Straub et al. др., 2015; Ли и др., 2017). Это может указывать на повышенную стабильность наземных каналов, когда их концы достигают края шельфа, как было количественно определено в предыдущих экспериментах (Kim et al., 2013). За этим последовало несколько событий отрыва меньшего масштаба. По мере реорганизации системы длительное псевдо-проседание привело к затоплению старых лопастей дельты и созданию среды на шельфе. Я характеризую шельф в нашем эксперименте как низко наклонную (S = 0,05 м / м) морскую среду, которая была построена в наземных условиях, но позже затонула из-за местной истории RSL.
Рис. 2. Верхние изображения трех экспериментальных стадий.Изображения отображают транспортную систему в ключевые периоды времени в циклах уровня моря (этапы 2 и 3) или в ключевые периоды в цикле автогенных каналов (этап 1). Показаны фотографии основных условий потока и последующего паводка. Изображения были сделаны после 1 полной минуты высвобождения красителя в резервуар. Основные отметки на границах изображений имеют интервал 1 м, а на верхнем левом фото указано контрольное местоположение с расстояниями от проксимальной стенки бассейна ( x ) и от правой стенки бассейна реки ( y ).
Рисунок 3. Топографические карты трех этапов эксперимента. Карты отображают транспортную систему в ключевые периоды в циклах уровня моря (этапы 2 и 3) или в ключевые периоды в цикле автогенного русла (этап 1). Расстояние между линиями сетки на топографических картах составляет 0,5 м, а на верхней левой карте указано опорное местоположение с расстояниями от проксимальной стенки бассейна ( x ) и от правой стенки бассейна реки ( y ).
Во время этапа 1, входящий поток и наносы направлялись к береговой линии через смесь канализированного и выходящего за пределы берега потока.Крупнозернистые отложения в основном были приурочены к каналам, в то время как поток через берег привел к значительным мелкозернистым отложениям на вершине дельты. В отложениях у береговой линии преобладали крупнозернистые лопасти дельты. Осадки, которые не выходили за границы дельты в мутных нижних стоках, были почти исключительно мелкозернистыми. Эти нижние потоки были пластинчатыми и не имели обнаруживаемых каналов. Нижние потоки проходили по всей длине бассейна и погружались в дальний ров бассейна, но в этом месте не было большого количества взвешенных наносов.Единственное существенное различие между базовыми условиями и условиями паводкового потока заключалось в том, что количество наземного потока, выходящего за пределы берега, было заметно выше во время паводков.
Небольшие по величине и короткие циклы RSL этапа 2 привели к реорганизации канальных сетей, большая часть из которых произошла в условиях высоких нагрузок, когда наземная система уменьшилась в размерах, а полочная система расширилась. Однако так было не всегда. Между вторым и третьим циклами RSL образовался единственный канал в центре бассейна во время повышения уровня моря.Этот канал оставался на месте во время высокого эстакады, перекачивая большие объемы наносов в море, и продолжал оставаться открытым в последующем нижнем стояке. Как и на этапе 1, значительная часть входящего потока покинула наземные каналы в сторону берега как во время базового, так и во время паводкового потока в течение всего цикла уровня моря на этапе 2 (Рисунки 2, 3).
Единый цикл RSL стадии 3 дал четкие геоморфологические отклики (Рисунки 2, 3). Первоначальное повышение уровня моря, а затем повышение уровня моря привели к быстрому нарушению береговой линии и широкомасштабному расширению экспериментального шельфа.Поскольку наша экспериментальная установка не включала волн, на шельфе во время трансгрессии не образовывалась крупномасштабная эрозионная поверхность оврага. Во время высокого подъема система дельта-вершина имела высокую боковую подвижность и относительно небольшую канализацию. Во время падения уровня моря произошло образование русел, которое в конечном итоге превратилось в единый эрозионный коридор на правой стороне бассейна. Этот канал был достаточно глубоким и широким, чтобы сдерживать весь поток в условиях основного и паводкового потока. Таким образом, этот канал действовал как конвейерная лента, транспортирующая весь входящий поток и отложения к морю, а наземный берег испытывал недостаток наносов.Хотя размер шельфа уменьшился в условиях падения и низкого уровня грунта, он не исчез полностью, так как перенос наносов и их отложение в море было сужено по бокам до правого берега бассейна.
Стратиграфическая архитектура
Динамика поверхности, обсуждавшаяся выше, привела к стратиграфическому комплексу с отложениями в верхней части дельты, которые в среднем составляли примерно 0,07 м в толщину, с предельными уровнями дельты шельфа более 0,15 м и с отложениями на склонах, которые сужались примерно от 0.Мощность от 03 м до 0,005 м. Панели сохранившихся пластов были получены путем вертикального разреза окончательной залежи вдоль трех преимущественно наклонных панелей. Я использовал расположение транспортной системы на каждом этапе эксперимента при выборе расположения измерительных панелей. К ним относятся: (1) панель вниз по реке с левой стороны бассейна, которая выровнена с наиболее долгоживущим аутогенным руслом на этапе 1, (2) панель в центре бассейна, которая захватила пласты, отложившиеся в течение нескольких этапов. 2 цикла, и (3) панель вниз по реке на правой стороне бассейна, которая была выровнена с доминирующим путем потока во время этапа 3 (Рисунок 4).
Рисунок 4. Изображения сохранившейся физической стратиграфии трех экспериментальных этапов. Основное направление транспортировки — слева направо. Окрашивание в синий цвет возникает из-за пищевого красителя, используемого для окрашивания входящего потока, и в основном присутствует на тонкозернистых слоях. На панели «Вставка» указано расположение разделов, которые отображаются с четырехкратным увеличением по вертикали.
Отложения в верхней части дельты на центральном разрезе и на левом разрезе реки были схожи по своей природе и включали прослои красного песка и белые мелкозернистые пласты с редкими косыми врезами русла одинаковой глубины.Пласты в верхней части дельты вдоль правого разреза реки включали большое эрозионное заполнение русла со стадии 3. Наиболее крупные отложения на каждом разрезе были обнаружены в выступах кромки шельфа с мелкозернистыми пластами, образующими пласты нисходящего склона.
Чтобы дополнить фациальную информацию, содержащуюся в изображениях физической стратиграфии, я строю тома синтетической стратиграфии, складывая ЦМР топографии, которые обрезаны для учета эрозии в топографических временных рядах (рис. 5).Поскольку я знаю время выполнения, связанное с каждой картой и соответствующим уровнем моря, я могу рисовать панели синтетических слоев по положению в цикле RSL. Полный пространственный охват бассейна позволяет мне извлекать эти поперечные сечения по любому желаемому пути. Поскольку уровень моря не менялся на этапе 1, я сосредотачиваюсь на сравнении этапов 2 и 3. Я извлекаю разрезы синтетической стратиграфии вдоль тех же разрезов, что и наша физическая стратиграфия. Слои наклонной панели, которая проходила по середине бассейна, показывает, что стратиграфия верхней части дельты была построена как в условиях высокого, так и в условиях низкого стояния RSL во время стадии 2 и стадии 3, но с предпочтением отложений в условиях высокого стояния.По крайней мере, вдоль этой трансекты значительная часть прогнозных пластов также откладывалась в условиях высокого стояния, но с перекрытием на прогнозных пластах, отложенных во время последующего низкого стояния.
Рис. 5. Панели синтетической стратиграфии с отложениями, нарисованными как функция времени отложений в пределах цикла уровня моря. Страты этапа 1 отображаются без раскраски, но с сохраненными временными горизонтами в виде сплошных черных линий. Толстые зеленые линии обозначают разрыв между слоями стадии 2 и 3.Вставьте детали карты местоположения каждого разреза; цветная полоса показывает, как синтетический стратиграфический цвет соответствует времени отложения.
Синтетические пласты, проанализированные вниз по правой стороне бассейна реки, следуя по пути доминирующего эрозионного русла стадии 3, показывают, что строительство началось во время высокого уровня моря, но его рост ускорился, когда уровень моря упал до низкого уровня. Сохранившиеся пласты кровли дельты вдоль этой трансекты образовались в условиях низкого уровня, поскольку более ранние отложения кровли дельты были в значительной степени эродированы и переносились вниз.
Я также извлек панели пластов, ориентированных по простиранию. Здесь я сравниваю три панели: одна ориентирована на полпути вниз по вершине дельты, одна около начального разрыва склона шельфа, а третья — у носка доминирующих выступов дельты края шельфа. Я вижу общее уменьшение сохранившихся пластов, отложившихся в условиях высокого стояния, и увеличение сохранившихся пластов низкого стояния по мере продвижения от источника к опусканию.
Количественная оценка взаимосвязи между RSL и массовым извлечением
Основная цель этого исследования — количественная оценка условий, необходимых для того, чтобы циклы RSL влияли на характер отложений связанных систем дельты и склонов на краю шельфа.Принято считать, что циклы уровня моря влияют на место осаждения, при этом осаждение перемещается к бассейну по мере понижения уровня моря и наоборот, с повышением уровня моря (Van Wagoner et al., 1990; Catuneanu et al., 2009). Я исследую это в экспериментальных слоях, строя набор модифицированных диаграмм Уиллера (Wheeler, 1964). Сначала я построил их для отдельных участков падения и простирания. Падение происходит от биссектрисы бассейна (рис. 6А), а простирание — от места расположения края шельфа в начале эксперимента (рис. 6В).Я генерирую матрицу, заполненную сохраненными скоростями отложений из последовательных пар топографических карт. Я отмечаю, что это сохраненные скорости отложений и, таким образом, они построены после того, как произошло топографическое отсечение, чтобы учесть эрозию после образования отложений. Это делается для каждого участка ниже по течению на участке падения и каждого участка поперечного потока на участке простирания, при этом расстояние между точками измерения равно 5-миллиметровой топографической сетке. Чтобы облегчить сравнение нашего эксперимента с полевыми системами, я нормализовал скорость осаждения на наложенную длительную скорость псевдоседания:
D * = D (x или y) r¯ (4)
Где D (x или y) — скорость осаждения в данном месте нисходящего потока (x) или поперечного потока (y).
Рис. 6. Модифицированные диаграммы Уиллера , на которых подробно показано, как средняя скорость осаждения изменяется в зависимости от расстояния от источника, бокового положения в бассейне и времени работы. (A) Диаграмма Уиллера, полученная из одного разреза по падению вниз по средней линии бассейна (разрез A – C на Рисунке 5). (B) Диаграмма Уиллера, полученная по разрезу одиночного простирания, расположенному на краю шельфа в начале эксперимента (разрез G – H на Рисунке 5). (C) Диаграмма Уиллера, ориентированная на среднее падение, построенная на основе среднего значения всех разрезов падения на топографических картах. (D) Диаграмма Уиллера с ориентировкой по простиранию, построенная на основе средних значений всех разрезов по простиранию на топографических картах. Белые точки на диаграммах Уиллера представляют собой стратиграфические перерывы. Черные пунктирные линии отмечают разрывы стадий, а экспериментальная история уровня моря показана для облегчения анализа пространственно-временной информации.
На этих двумерных разрезах я наблюдаю крайнюю изменчивость скорости отложений в пространстве и времени с большим количеством стратиграфических перерывов. Измеренные значения D ∗ достигают пика при значениях, превышающих 20 по прогнозам дельты, но сложно связать динамику отложений с наложенной историей уровня моря при падении или простирании.
В попытке уменьшить «осадочный шум», связанный с просмотром разрезов с одним падением или простиранием, я создал второй набор модифицированных диаграмм Уиллера. Для этого была сгенерирована диаграмма Уиллера с ориентировкой по наклону путем усреднения всех строк с ориентацией по наклону в наборе топографических данных ( n = 531) (Рисунок 6C). Тот же процесс был использован для построения диаграммы, ориентированной по среднему простиранию путем усреднения всех колонок, ориентированных по простиранию в наборе топографических данных ( n = 923) (рис. 6D).
Я наблюдаю D ∗ значений в диапазоне от 0 до 3 на диаграммах Уиллера, усредненных по бассейну, с самыми высокими значениями по прогнозам дельты (Рисунок 6). Если посмотреть на участок падения, то на стадии 1 значения пика D ∗ сначала перемещаются в сторону бассейна, пока не произойдет быстрый переход к источнику во время первого крупного отрыва. За этим следуют более мелкие циклы проградации, а затем ретроградация значений пика D ∗ . Я не наблюдаю четкого отпечатка малой амплитуды и короткопериодных циклов второй ступени на диаграмме Уиллера.По сравнению с короткопериодными циклами уровня моря наблюдается гораздо более продолжительный период автогенной проградации, а затем ретроградный цикл. Однако высокая величина и длительный цикл уровня моря на этапе 3 вызвали четкую реакцию в месте пиковых скоростей осаждения. Первоначальный подъем уровня моря к условиям высокого стояния привел к значению пика D ∗ в направлении источника отложений, в то время как падение уровня моря во время этого цикла привело к сдвигу в сторону бассейна D ∗ .Стадия 3 также была единственной стадией со стратиграфическим перерывом по всей ширине проксимальной системы осадконакопления. Этот перерыв произошел во время падающей части цикла стадии 3, когда земная система закрепилась за счет эрозии и вытолкнула отложения в море.
Усредненное по бассейну простирание снова показывает аналогичный характер на первом и втором этапах с зонами отложений, смещенными вбок для компенсации топографических понижений. Стадия 3, однако, характеризуется отложениями, которые остаются на правой стороне бассейна реки, и стратиграфическим перерывом, который охватывает большую часть реки с левой стороны бассейна во время падения уровня моря.Этот стратиграфический перерыв в значительной степени связан с бездействием, а не с эрозией, поскольку транспортная система в это время была ограничена правым берегом реки.
Модифицированная диаграмма Уиллера, представленная выше, помогает визуализировать движение пиков депоцентров в эксперименте, но меня также интересует описание кумулятивного извлечения массы для осаждения как функции расстояния от источника отложений и того, как это изменяется (1) между стадиями. и (2) в течение отдельных циклов уровня моря.Это может быть полезно для определения того, способствуют ли циклические характеристики уровня моря перекачке наносов в более отдаленные места и совпадает ли движение накопленного источника к опусканию депоцентра с наложенными циклами уровня моря. Для этого я построил кривые извлечения массы в зависимости от расстояния ниже по потоку для каждой экспериментальной стадии. Я использую определение массового извлечения, данное Паола и Мартин (2012), как общий поток наносов, потерянный в результате осаждения на интересующем расстоянии x от источника, нормированный на поток наносов на входе в бассейн, Q S 0 :
χ (x) = 1QS0∫0xB (x) D¯ (x) dx (5)
Где B (x) — ширина бассейна в местоположении x .Наглядным примером этого преобразования является χ (x) = 0,5, где половина наносов, поступающих в бассейн, откладывается выше по течению от местоположения x , а половина наносов все еще находится в потоке. χ (x) = 0,5 также представляет собой среднюю точку осадконакопления бассейна (Паола и Мартин, 2012).
Я начинаю анализ экстракции массы с построения кривых χ (x) для каждой стадии (рис. 7A). Это делается с помощью карты изопахит, созданной путем отличия топографии в конце каждого экспериментального этапа от топографии в начале этапа.Форма кривых χ (x) аналогична для трех стадий с начальным медленным ростом χ (x) на расстояниях от источника отложений, которые обычно были средами в верхней части дельты. Затем наблюдается быстрый рост χ (x) между 1 и 2 м от входа в бассейн, который, как правило, представлял собой выступ дельты и верхний склон. Наконец, χ (x) медленно растет на 2–4 м от источника, который всегда был средой морского склона. Используя базу данных DEM, я также вычисляю среднее расстояние от входа в бассейн до береговой линии для каждого этапа и размещаю его на кривых χ (x) (рис. 7).Это позволяет мне измерить массовое распределение между преимущественно наземными и преимущественно морскими условиями на каждой стадии. Я обнаружил, что на стадии 1 19% входной массы было отложено в земном, в то время как на обеих стадиях 2 и 3 было отложено 32% входной массы в земном. Самая большая разница в кривых χ (x) трех стадий — это медленный начальный рост χ (x) на стадии 1 по сравнению со стадиями 2 и 3. Расстояние от входа до средней береговой линии почти одинаковое. для всех стадий, но наносов, извлекаемых у береговой линии, меньше на стадии 1 по сравнению с стадиями 2 и 3.Это, вероятно, связано с долгоживущим и относительно стабильным каналом, который присутствовал на большей части стадии 1, который эффективно перекачивал отложения в море.
Рис. 7. Зависимость между расстоянием от источника и массовым извлечением для экспериментального месторождения. (A) χ (x) кривая для отложений навалочного яруса с указанием положения средней береговой линии для каждого этапа. (B) χ (x) кривых для каждого 3-часового окна между топографическими сканированиями на этапе 1, жирная сплошная серая линия — это средний тренд этапа. (C, D) χ (x) кривых для трехчасовых окон между топографическими сканированиями на этапе 2 и 3, соответственно. Цвет кривой соответствует времени окна выборки относительно цикла уровня моря, а сплошные серые линии показывают средние тенденции стадии.
Затем я вычисляю кривые χ (x) для каждого последовательного набора ЦМР, следуя процессу отсечения для пост-осадочной эрозии. Это служит двум целям: (1) охарактеризовать степень вариабельности в месте добычи массы из-за как аутогенной, так и аллогенной реорганизации сетей каналов и (2) определить, приводит ли положение в цикле уровня моря к предсказуемым изменениям массы. тенденция добычи.Что касается точки 2, за вычетом аутогенной динамики, я мог бы ожидать, что уровень моря будет коррелировать с проксимально-дистальной тенденцией извлечения массы: более низкий уровень моря может привести к проксимальному шунтированию и более дистальному извлечению массы по сравнению со средним значением χ (x) кривая цикла уровня моря.
Сравнение стадий 1 и 2 предполагает аналогичную степень изменчивости кривых χ (x) вокруг среднего тренда (Рисунки 7B, C). χ иногда превышает значение 1 на этих кривых из-за эрозии и повторного отложения ранее отложенных отложений, что может привести к большему объему отложений, отложенных и сохраненных в течение данного трехчасового периода, чем поступление в бассейн в то же время.Для кривых χ (x) стадии 3 (рис. 7D) наблюдается больший разброс среднего значения, что указывает на отпечаток воздействия, превышающий стохастическую изменчивость.
Положение в пределах цикла уровня моря не вызывает предсказуемых сдвигов в кривых χ (x) для короткопериодных циклов стадии 2 с малой величиной. В то время как некоторые периоды низкого уровня моря соответствуют сдвигу в сторону бассейна тенденции извлечения массы. , относительно среднего тренда стадии, другие периоды низкого уровня моря связаны с усиленным извлечением проксимальной массы по сравнению со средней тенденцией (Рисунок 7C).Напротив, четкий и предсказуемый ответ χ (x) наблюдается на стадии 3 с низким RSL, связанным с удалением более дистальных масс, и наоборот (рис. 7D).
Ранее при описании синтетических стратиграфических панелей я отмечал общую тенденцию сокращения сохранившихся пластов, отложившихся в условиях высокого стояния, и увеличения сохранившихся пластов низкого стояния при перемещении от истока к опусканию. Моя цель здесь — дать количественную оценку этой тенденции. Это делается с помощью расчета параметра α D , впервые представленного Yu et al.(2017), который дает количественную оценку того, когда в среднем происходит осаждение во время цикла уровня моря, и как это изменяется в зависимости от места извлечения массы. Этот показатель строится как:
αD = 1N∑i = 1N (Di¯ (χ) r¯ηSL, iMRSL / 2) (6)
Где D¯i (χ) — средняя толщина пластов, отложившихся в течение часа эксперимента i в месте добычи массы χ, η SL, i — уровень моря в течение часа запуска i и N — общее количество проанализированных часов работы.Для пластов, отложенных в течение данного цикла RSL, α D теоретически может принимать любое значение от -1 до 1. В описательных терминах значение -1 будет представлять месторождение в интересующем месте добычи, которое было построено только когда уровень моря был на пике цикла, тогда как значение 1 будет представлять отложения, образовавшиеся только тогда, когда уровень моря был на пике цикла.
Поскольку на стадии 1 не было циклов уровня моря, α D остается постоянным значением 0 для всех местоположений χ.На стадиях 2 и 3 α D представляет собой положительное значение, выходящее за пределы береговой линии средней стадии, что указывает на предпочтение высокостоящих отложений (рис. 8). На обоих этапах это предпочтение увеличивается с удалением от источника до достижения пиковых значений. Однако пик в α D значительно выше на стадии 3 по сравнению со стадией 2, что указывает на быстрое осаждение в проксимальных местах в условиях высокого уровня моря для циклов на уровне моря большой величины. В более удаленных местах извлечения массы эта тенденция меняется на противоположную.Между местоположениями χ 0,6–0,8 я наблюдаю отрицательные значения α D на обеих стадиях, причем тенденция достигает более низких значений на стадии 3 по сравнению со стадией 2. Это указывает на предпочтение быстрого осаждения в море во время низких условий, увеличивается с увеличением величина цикла уровня моря. Ближе к концу транспортной системы α D начинает подниматься обратно к нулю, что указывает на потерю предпочтения отложения в зависимости от местоположения в цикле уровня моря.Однако следует отметить, что в этих чрезвычайно удаленных местах толщина отложений резко падает, что, вероятно, затрудняет идентификацию этого сигнала в системах масштаба поля.
Рис. 8. Данные, определяющие среднее положение в пределах цикла уровня моря, при котором отложения осаждаются, как функция расстояния от входа в бассейн. Символы указывают положение средней береговой линии для каждого этапа.
Обсуждение
Я начинаю с общего резюме моих экспериментальных результатов и сравнения с наблюдениями из численных экспериментов Харриса и др.(2016, 2018). Для этого сравнения я произвольно масштабирую экспериментальные этапы до полевых условий, используя возникающие экспериментальные автогенные шкалы. Здесь я делаю приблизительный апскейлинг до масштабной системы реки Миссисипи, предполагая сходство морфодинамической реакции между экспериментальной (обозначенной как e ) и прототипной (обозначенной как p ) системами, которые имеют безразмерные соотношения, которые сравнивают аутогенную и аллогенную шкалы. Я подчеркиваю, что, как и все стратиграфические эксперименты, этот эксперимент не является моделированием какой-либо одной конкретной системы, поскольку сложно сопоставить многие важные безразмерные числа, которые описывают поля потока и переноса наносов при сравнении лабораторных и полевых систем.Однако некоторые возникающие геоморфологические масштабы и масштабная инвариантность многих процессов позволяют сравнить некоторую динамику с полевыми системами (Paola et al., 2009). Таким образом, это сравнение дает приблизительное представление о временных масштабах и откликах, которые следует ожидать для систем в масштабе поля.
Я начинаю с апскейлинга экспериментальных циклов уровня моря:
MRSL, eHC, e = H * = MRSL, pHC, p (7)
Учитывая, что глубокие аутогенные участки реки Миссисипи в ее затоне имеют глубину около 50 м (Nittrouer et al., 2012), шкала 2-го и 3-го циклов уровня моря составляет 25 и 100 м соответственно.
Затем я увеличиваю продолжительность или период экспериментальных циклов уровня моря, используя следующее соотношение:
TRSL, eTC, e = T * = TRSL, pTC, p (8)
Используя уравнение 2 и значение rD¯ для реки Миссисипи, полученное по биостратиграфическим датам (rD¯ = 0,26 м / тыс. Лет) (Straub et al., 2009), этапы 2 и 3 циклов уровня моря масштабируются до 96 и 384 тыс. Лет соответственно .
Однако приведенное выше масштабирование справедливо только для аутогенных чешуек дельты реки Миссисипи. Если мы применим уравнения 7, 8 для сравнения наших стадий 2 и 3 с дельтой реки Рейн, которая имеет значения H c и rD¯, равные 7 м и 1,2 мм / год соответственно (Zagwijn, 1989; Hijma et al. , 2009), мы получаем существенно иное масштабирование. Для дельты Рейна мои циклы уровня моря 2 и 3 масштабируются до 3,5 и 14 м с продолжительностью 2,9 и 11,7 тыс. Лет соответственно. Сравнение систем дельты реки Миссисипи и Рейн позволяет предположить следующее.Во-первых, не все речные дельты одинаково реагируют на определенный цикл изменения уровня моря. Более крупные системы с их более крупными аутогенными масштабами будут менее чувствительны к циклам уровня моря по сравнению с их более мелкими соседними системами. Этот момент был подчеркнут в ряде недавних исследований (Li et al., 2016; Yu et al., 2017; Trower et al., 2018). Во-вторых, эти результаты предполагают, что следует проявлять осторожность при универсальном применении одного набора правил к дельтам состояния ледника, а другого — к дельтам состояния теплицы.Например, отклик более крупных дельт с автогенными масштабами, подобными циклам уровня моря ледникового покрова, может быть аналогичен отклику более мелких систем на высокочастотный и низкомасштабный цикл уровня моря в парниковом масштабе, как те, которые предложены для позднего мела и более раннего палеогена (Тиберт и Leckie, 2004; Miller, 2009). В-третьих, хотя некоторые большие системы в условиях ледника могут вести себя как меньшие системы в условиях теплицы, в среднем автогенные масштабы дельт будут больше по сравнению с масштабами навязанных циклов уровня моря в условиях теплицы и, следовательно, в среднем менее чувствительны к ним. циклы.Это будет способствовать более непрерывной связи между наземными условиями и условиями на склоне в это время, подобно тому, что было предложено на основе исследований численного моделирования (Sømme et al., 2009a).
Привязка массового распределения вдоль источника к профилям стока к циклам уровня моря
В наших экспериментах соотношение между извлечением массы и физическим расстоянием от входа в бассейн было почти идентичным на трех экспериментальных стадиях (рис. 7A). Стадия, которая перекачивала в море больше всего наносов, была той, на которой отсутствовали какие-либо циклы уровня моря, однако эта разница составила всего 13%.Это увеличение закачки наносов в море на стадии 1 может быть связано с долгоживущим и относительно стабильным руслом, присутствующим более чем на половине этой стадии. В этом канале образовалась дельта-лопасть, которая выступала далеко за пределы начального разрыва склона шельфа. Это согласуется с предыдущим экспериментом, в котором наблюдалась стабилизация каналов, которые могут перекачивать свои отложения в глубокие воды (Kim et al., 2013). Хотя отрывы все еще происходят после стыковки с краем шельфа, считается, что их частота снижается, что способствует оттоку наносов в море.
Сходство среднего тренда χ ( x ) для стадий 2 и 3 поразительно и поддерживает идею о том, что разделение наносов вдоль разреза истока и погружения в значительной степени обусловлено долгосрочным трендом продукции аккомодации бассейна. Под долгосрочными я имею в виду временные масштабы, важные для возникновения оседания бассейна, которые превышают большинство циклических колебаний уровня Милинковича. В то время как тенденции извлечения валовой массы для отдельных стадий схожи (рис. 7A), мой анализ среднего положения в пределах цикла уровня моря, в течение которого были построены сохранившиеся пласты (рис. 8), предполагает предпочтение земных пластов, построенных во время высоких выступов, и морских пластов. во время невысоких платформ.Это предпочтение приглушается в циклах с низкой величиной и коротким периодом стадии 2 по сравнению с циклом большой величины и с длинным периодом стадии 3, но все же присутствует.
Важно отметить, что предпочтение наземных отложений во время высоких стояков и морских отложений во время низких, очевидное в нашем анализе α D , является усредненным по времени результатом. В частности, сигнал, присутствующий на этапе 2 депонирования, является средним результатом четырех циклов на этом этапе.Без точного контроля времени этот сигнал будет трудно выделить из стратиграфической архитектуры, построенной в маргинальных морских условиях, вызванных малой величиной и короткими циклами уровня моря. Например, экспериментальная транспортная система, испытывающая циклы уровня моря в масштабе 2 стадии, не всегда давала предсказуемые продукты аллогенного воздействия. Изменчивость краткосрочных тенденций извлечения массы на стадии 2 была аналогична изменчивости кривых извлечения массы на автогенной стадии 1 (Рисунки 7B, C), которые предполагают, что их стратиграфическая структура также была аналогичной.Кроме того, низкие условия на этапе 2 не всегда вынуждали транспортную систему перемещаться по бассейну. Напротив, большой и долгопериодный цикл уровня моря на этапе 3 произвел предсказуемое движение депоцентра с положением в циклах уровня моря, и изменения в месте добычи массы в условиях низкого (или высокого) уровня превышали автогенную стохастическую изменчивость. в депоцентре (рис. 7D).
Теперь я возвращаюсь к наблюдениям из численных экспериментов, подробно описанным в серии статей Харриса и др.(2016, 2018). Точное сравнение численных экспериментов Harris et al. (2016, 2018) в наш физический эксперимент невозможно из-за экспериментальных параметров масштабирования. Кроме того, глубины каналов в численных экспериментах Harris et al. неизвестны, что делает оценку H ∗ и T ∗ для их численных экспериментов неточной. Однако многие из смоделированных ими циклов уровня моря имеют величину менее 30 м и происходят на протяжении менее 100 тыс. Лет в области с вынуждающими условиями, мотивированными пассивной окраиной северо-востока Соединенных Штатов.Это может указывать на значения H ∗ и T ∗ , близкие к 1.
Физические экспериментальные результаты подтверждают открытие Harris et al. (2016, 2018), что существует лишь небольшая разница в доставке наносов между системами с постоянным и динамическим уровнем моря. Для систем, испытывающих циклы уровня моря, которые являются небольшими по сравнению с их аутогенными масштабами, время доставки наносов в море действительно имеет предпочтение для условий низкого уровня.Подобно результатам Harris et al. (2016), я отмечаю, что этот ответ может быть переменным и из-за аутогенных процессов иногда противоположен тем, которые предсказываются чисто аллогенными моделями. Например, канал, образовавшийся между вторым и третьим циклами уровня моря на стадии 2, мог удерживать доставку наносов в море на высоком уровне даже при повышении уровня моря. Этот канал был похож на некоторые из автогенных каналов 1-й стадии, и скорость его роста позволяла ему продолжать расширяться даже при длительном нарушении береговой линии остальной части дельты.Однако реакция на циклы уровня моря, которые велики по сравнению с автогенными шкалами системы, кажется предсказуемой и согласуется с давней теорией (Van Wagoner et al., 1990). Большие падения уровня моря коррелируют с выраженным движением депоцентра в направлении бассейна и увеличением поступления наносов в море. Для системы шкалы Миссисипи это будет означать циклы уровня моря с амплитудами более 50 м и / или периодами более 200 тыс. Лет. Однако, как упоминалось ранее, эти масштабы будут варьироваться в зависимости от размера системы, поскольку автогенные временные и пространственные масштабы обычно коррелируют с размером системы (Hajek and Straub, 2017).Таким образом, меньшие системы будут легче производить обнаруживаемые реакции при доставке наносов в море по сравнению с более крупными системами, подверженными воздействию того же цикла уровня моря (Li et al., 2016).
Закачка осадка, прошедшая экспериментальные прогнозы дельты
Ключевым моментом, который следует выделить в описанной здесь системе, является сложность откачки наносов за пределы дельта-прогнозов в физических экспериментах. Экспериментальный план был разработан таким образом, чтобы способствовать откачке наносов за границы дельты на краю шельфа в условия континентального склона.Эти условия включали частые наводнения и входящий поток, который имел избыточную плотность по сравнению с принимающим бассейном. Даже при этих проектных параметрах ~ 80% входящего осадка хранилось в вершине дельты и на прогнозе. Для сравнения, подробный сейсмический анализ дельты на краю шельфа и связанных подводных каналов в северо-восточной части Мексиканского залива обнаружил разделение объемов шельфа и склона 50:50 (Sylvester et al., 2012). Очевидно, что необходима дополнительная работа для определения экспериментального пространства параметров, необходимого для обеспечения потока больших объемов наносов в море.
Несмотря на то, что поток наносов за пределы прогнозируемой дельты не такой высокий, как в некоторых системах, я все же могу сравнить полученные изопахи уклона на трех стадиях. Этапы 1 и 2 имеют одинаковые отложения и объемы откосов (рис. 9A, B), которые были распределены по всей ширине платформы. Для сравнения, большой по величине и длительный цикл уровня моря на стадии 3 привел к образованию отложений на склоне, которые были более изолированными в поперечном направлении, но с более толстыми отложениями в отдаленных местах по сравнению с другими стадиями (Рисунок 9C).
Рис. 9. Карты, показывающие тенденции толщины отложений на экспериментальном склоне для трех экспериментальных стадий. Пределы шкалы цветов установлены для выделения рисунков отложений на откосах с наложением контуров 2 мм.
Заключение
Ключевые результаты, которые напрямую связаны с развитием теории процессов для систем маршрутизации донных отложений, являются двоякими:
(1) Результаты физического эксперимента на связанных системах дельты и склонов на краю шельфа предполагают, что масштаб циклов уровня моря не сильно влияет на перекачку наносов в море, если усреднить доставку наносов за полный цикл.Эта доставка наносов в море аналогична системам без циклов уровня моря, которые из-за длинных автогенных масштабов, когда дельты стыкованы с краем шельфа, действительно могут иметь устойчивую скорость доставки глубоководных отложений в течение длительных периодов времени.
(2) Хотя величина и период циклов уровня моря не влияют на долгосрочную доставку наносов в море (т.е. скорость, усредненную за весь цикл RSL), эти параметры действительно влияют на время доставки глубинных морских наносов. Циклы уровня моря со шкалами, превышающими автогенные динамические шкалы, дают предсказуемые реакции во время цикла уровня моря с максимальной доставкой во время предельно низких уровней.Напротив, циклы уровня моря со шкалой меньше, чем автогенная шкала, вызывают различные реакции. В среднем морские отложения доставляются во время низких стоянок для небольших циклов, но эта реакция иногда может быть перевернута, с высокой морской доставкой во время высоких стоянок. Переход между предсказуемой и стохастической реакцией может составлять от 50 м до 200 тыс. Лет для крупных речных систем.
Доступность данных
Наборы данных, созданные для этого исследования, доступны по запросу соответствующему автору.
Авторские взносы
Автор подтверждает, что является единственным соавтором этой работы, и одобрил ее к публикации.
Финансирование
Это исследование было частично поддержано Национальным научным фондом (EAR-1424312).
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Я благодарю бывших и нынешних членов Лаборатории динамики и стратиграфии отложений Тулейна, в частности Анджали Фернандеса и Тушара Бишноя, за помощь в создании экспериментальных начальных условий и за помощь в вскрытии окончательного месторождения.Наконец, я благодарю редакторов Д.Х. и Джули Фосдик за обработку заявки, а также рецензентов JB и KM за обзоры, которые помогли повысить ясность рукописи.
Список литературы
Аллен, П. А. (2008). «Временные масштабы тектонических ландшафтов и их системы маршрутизации отложений», в «Эволюция ландшафта: денудация, климат и тектоника в различных временных и пространственных масштабах» Галлагер , К., Джонс и Уэйнрайт, Дж. Лондон: Геологическое общество Лондона
Аллен, П.А. (2017). Системы маршрутизации отложений: судьба отложений от источника к раковине , Кембридж: Издательство Кембриджского университета.
Google Scholar
Кантелли А., Пирмез К., Джонсон С. и Паркер Г. (2011). Морфодинамическая и стратиграфическая эволюция самоканальных подводных вееров, создаваемых мутными течениями. J. Sediment. Res. 81, 233–247. DOI: 10.2110 / jsr.2011.20
CrossRef Полный текст | Google Scholar
Катуняну О., Абреу В., Bhattacharya, J. P., Blum, M. D., Dalrymple, R. W., Eriksson, P. G., et al. (2009). К стандартизации стратиграфии последовательностей. Науки о Земле. Ред. 92, 1–33.
Google Scholar
Гилберт, Г. К. (1890). Озеро Бонневиль, Монография Геологической службы США 1 . Вашингтон, округ Колумбия, Типография правительства США
Гранджон, Д. (1999). Концепции и применение трехмерной множественной литологии, диффузионной модели в стратиграфическом моделировании .Франция: специальные публикации SEPM
Хайек, Э.А., и Штрауб, К.М. (2017). Аутогенная седиментация в обломочной стратиграфии. Ann. Преподобный «Планета Земля». Sci. 45, 681–709. DOI: 10.1146 / annurev-earth-063016-015935
CrossRef Полный текст | Google Scholar
Гамильтон, П., Гайо, Г., Стром, К., Феделе, Дж., И Хойал, Д. (2017). связь гидравлических свойств в сверхкритических подводных распределительных каналах с геометрией осадочных лепестков. Гамильтон и др.связь гидравлических свойств в подводных каналах с геометрией осадочных лопастей. J. Sediment. Res. 87, 935–950. DOI: 10.2110 / jsr.2017.53
CrossRef Полный текст | Google Scholar
Харрис А. Д., Ково Дж. А., Мадоф А. С., Сан Т., Сильвестр З. и Гранджон Д. (2016). Трехмерное численное моделирование эвстатического контроля над распределением песка на окраинах континентов А. d. Харрис и др. численное моделирование эвстатического контроля над распределением песков континентальной окраины. J. Sediment. Res. 86, 1434–1443. DOI: 10.2110 / jsr.2016.85
CrossRef Полный текст | Google Scholar
Харрис А. Д., Баумгарднер С. Э., Сан Т. и Гранджон Д. (2018). Плохая связь между уровнем моря и доставкой глубоководного песка. Осадок. Геол. 370, 42–51. DOI: 10.1016 / j.sedgeo.2018.04.002
CrossRef Полный текст | Google Scholar
Хиджма, М. П., Коэн, К., Хоффманн, Г., Ван дер Спек, А. Дж., И Стаутхамер, Э. (2009).От долины реки до устья: эволюция устья Рейна в период от раннего до среднего голоцена (западные Нидерланды, дельта Рейн-Маас). Нидерланды J. Geosci. 88, 13–53. DOI: 10.1017 / s0016774600000986
CrossRef Полный текст | Google Scholar
Хойал, Д. К. Дж., И Шитс, Б. А. (2009). Морфодинамическая эволюция экспериментальных когезионных дельт. J. Geophys. Res. Прибой Земли. 114: F02009
Google Scholar
Джеролмак, Д. Дж., и Сэдлер, П. (2007). Кратковременность и стойкость записей отложений на окраинах континентов. J. Geophys. Res. Прибой Земли. 112: F03S13.
Google Scholar
Ким В., Петтер А., Штрауб К. и Мориг Д. (2014). «Изучение реакции аутогенного процесса на аллогенное воздействие: экспериментальная геоморфология и стратиграфия»: От осадочных систем к осадочным сукцессиям на норвежской континентальной окраине , изд. Дж. П. Вонхэм А. В. Мартиниус, Хобокен: John Wiley & Sons, Ltd, 127–138.DOI: 10.1002 / 9781118920435.ch5
CrossRef Полный текст | Google Scholar
Ким Ю., Ким В., Чеонг Д., Муто Т. и Пайлс Д. Р. (2013). Подача крупнозернистого осадка в глубоководный вентилятор через перепускной канал дельты у кромки шельфа: эксперименты с резервуарами. J. Geophys. Res. Прибой Земли. 118, 2279–2291. DOI: 10.1002 / 2013jf002813
CrossRef Полный текст | Google Scholar
Ли, К., Ю., Л., Штрауб, К. М. (2016). Пороги хранения относительных сигналов уровня моря в стратиграфической записи. Геология 44, 179–182. DOI: 10.1130 / g37484.1
CrossRef Полный текст | Google Scholar
Ли, К., Мэтью Бенсон, В., Харлан, М., Робишо, П., Ша, X., Сюй, К. и др. (2017). Влияние сплоченности наносов на морфодинамику и стратиграфию дельт в масштабах времени заполнения бассейна. Дж. Геофи. Res. Прибой Земли. 122, 1808–1826. DOI: 10.1002 / 2017jf004216
CrossRef Полный текст | Google Scholar
Мохриг, Д., Хеллер, П. Л., Паола, К.и Лайонс, В. Дж. (2000). Интерпретация процесса отрыва из древних аллювиальных отложений: Гуадалоп-Матаррания (северная Испания) и образование уазач (западный Колорадо). Геол. Soc. Являюсь. Бык. 112, 1787–1803.
Google Scholar
Nittrouer, J. A., Shaw, J., Lamb, M. P., and Mohrig, D. (2012). Пространственные и временные тренды скорости потока воды и перенос донных отложений в нижнем течении реки Миссисипи. Геол. Soc. Являюсь. Бык. 124, 400–414. DOI: 10.1130 / b30497.1
CrossRef Полный текст | Google Scholar
Паола К., Штрауб К. М., Мориг Д. и Рейнхардт Л. (2009). «Необоснованная эффективность» стратиграфических и геоморфологических экспериментов. Науки о Земле. Ред. 97, 1–43. DOI: 10.1016 / j.earscirev.2009.05.003
CrossRef Полный текст | Google Scholar
Паола, К. (2016). Собственный разум: последние достижения в автогенной динамике рек и дельт. SEPM Спец. Publ. 106, 5–17.DOI: 10.2110 / sepmsp.106.04
CrossRef Полный текст | Google Scholar
Порембски, С. Дж., И Стил, Р. Дж. (2003). Дельты окраин шельфа: их стратиграфическое значение и связь с глубоководными песками. Науки о Земле. Ред. 62, 283–326. DOI: 10.1016 / s0012-8252 (02) 00161-7
CrossRef Полный текст | Google Scholar
Сёмме, Т. О., Хелланд-Хансен, В., и Гранджон, Д. (2009a). Влияние вариаций эвстатической амплитуды на морфологию шельфа, распространение наносов и стратиграфическую интерпретацию последовательностей: ледниковые системы против тепличных систем. Геология 37, 587–590. DOI: 10.1130 / g25511a.1
CrossRef Полный текст | Google Scholar
Сёмме, Т. О., Хелланд-Хансен, В., Мартинсен, О., и Турмонд, Дж. (2009b). Взаимосвязь между морфологическими и седиментологическими параметрами в системах «источник-поглотитель»: основа для прогнозирования полуколичественных характеристик в подземных системах. Basin Res. 21, 361–387. DOI: 10.1111 / j.1365-2117.2009.00397.x
CrossRef Полный текст | Google Scholar
Штрауб, К.М., Мохриг, Д., МакЭлрой, Б., Баттлс, Дж., И Пирмез, К. (2008). Взаимодействие между мутными течениями и топографией в расширяющихся извилистых подводных каналах: лабораторное исследование. Геол. Soc. Являюсь. Бык. 120, 368–385. DOI: 10.1130 / b25983.1
CrossRef Полный текст | Google Scholar
Штрауб, К. М., Паола, К., Мориг, Д., Волински, М. А., и Джордж, Т. (2009). Компенсационная укладка русловидных осадочных отложений. J. Sediment. Res. 79, 673–688.DOI: 10.2110 / jsr.2009.070
CrossRef Полный текст | Google Scholar
Штрауб К. М., Мохриг Д. и Пирмез К. (2012). Архитектура агградационной сети подводных каналов притока на континентальном склоне у берега Брунея-Даруссалама, в г. Применение принципов сейсмической геоморфологии к системам континентального склона и основания склона: тематические исследования по аналогам морского и подводного дна eds BE, Prather ME , Дептак Д., Мориг Б., ван Хорн и Р. Б., Винн Талса: Общество осадочной геологии
Штрауб, К. М., Ли, К., Бенсон, В. М. (2015). влияние сплоченности наносов на динамику береговой линии дельты и объемное удержание наносов: лабораторное исследование. Geophys. Res. Lett. 42, 9808–9815. DOI: 10.1002 / 2015gl066131
CrossRef Полный текст | Google Scholar
Свит, М. Л., и Блюм, М. Д. (2016). Связи между речной и мелководной морской средой и подводными каньонами: последствия для переноса наносов в глубокие воды. J. Sediment. Res. 86, 1147–1162. DOI: 10.2110 / jsr.2016.64
CrossRef Полный текст | Google Scholar
Сильвестр, З., Дептак, М., Пратер, Б., Пирмез, К., О’Бирн, К., Мохриг, Д. и др. (2012). Сейсмическая стратиграфия дельты края шельфа и связанных подводных каналов в северо-восточной части Мексиканского залива: применение принципов сейсмической геоморфологии к системам с континентальным склоном и подножием склона: тематические исследования по аналогам морского дна и морского дна. SEPM Спец.Publ. 99, 31–59. DOI: 10.2110 / pec.12.99.0031
CrossRef Полный текст | Google Scholar
Тиберт, Н. Э., и Леки, Р. М. (2004). Циклы уровня моря в устьях с высоким разрешением из позднего мела: ограничения амплитуды с использованием агглютинированных фораминифер. J Foram. Res. 34, 130–143. DOI: 10.2113 / 0340130
CrossRef Полный текст | Google Scholar
Троуэр, Э. Дж., Ганти, В., Фишер, В. В., и Лэмб, М. П. (2018). Эрозионные поверхности в верхних меловых песчаниках из ливневых отложений (Юта, США): границы разреза или аутогенный размыв от гидродинамики подпора ?. Геология 46, 707–710. DOI: 10.1130 / g40273.1
CrossRef Полный текст | Google Scholar
Вейл, П. Р., Митчам, Р. М., и Томпсон, С., III. (1977). «Сейсмическая стратиграфия и глобальные изменения уровня моря: Часть 3. Относительные изменения уровня моря от прибрежного перекрытия», в Сейсмическая стратиграфия — Приложения к разведке углеводородов ed C.E., Пэйтон Талса AAPG Memoir 63–82.
Ван Вагонер, Дж. К., Митчам, Р. М., Кэмпион, К. М., и Рахманян, В.Д. (1990). Последовательная стратиграфия кремнисто-обломочного материала в каротажных диаграммах, кернах и обнажениях: концепции высокоразрешающей корреляции времени и фаций : Талса: Американская ассоциация геологов-нефтяников:
Google Scholar
Vendettuoli, D., Clare, M., Clarke, J.H., Vellinga, A., Hizzet, J., Hage, S., et al. (2019). Ежедневные батиметрические исследования показывают, как устроена стратиграфия, и какова ее крайняя неполнота в подводных каналах. Земля. Планета. Sci. Lett. 515, 231–247.DOI: 10.1016 / j.epsl.2019.03.033
CrossRef Полный текст | Google Scholar
Ван, Ю., Штрауб, К. М., и Хайек, Э. А. (2011). Компенсационное накопление в зависимости от масштаба: оценка автогенных временных масштабов в канальных осадочных отложениях: Geology 39, 811–814. DOI: 10.1130 / g32068.1
CrossRef Полный текст
Уиллер, Х. Э. (1964). Базовый уровень, поверхность литосферы и временная стратиграфия. Геол. Soc. Являюсь. Бык. 75, 599–610.
Google Scholar
Сюй, Дж.П., Нобель, М.А., и Розенфельд, Л.К. (2004). Измерения на месте скоростной структуры при мутных течениях. Geophys. Res. Lett. 31: L09311
Google Scholar
Ю. Л., Ли К. и Штрауб К. М. (2017). Масштабирование реакции дельт на циклы относительного уровня моря по автогенным пространственным и временным шкалам: лабораторное исследование. J. Sediment. Res. 87, 817–837. DOI: 10.2110 / jsr.2017.46
CrossRef Полный текст | Google Scholar
Zagwijn, W.Х. (1989). Нидерланды в третичном и четвертичном периодах. история развития прибрежных низменностей. Геол. En Mijnbouw 68, 107–120. DOI: 10.1007 / 978-94-017-1064-0_6
CrossRef Полный текст | Google Scholar
Масс-спектрометр изотопного отношения Delta V ™
Вы используете нашу серию Delta V? Нам интересно услышать ваше мнение, и мы будем признательны, если вы заполните этот опрос в пакете программного обеспечения Thermo Scientific ™ Isodat Software Suite. Пожалуйста, пройдите по этой ссылке.
Нужна гибкость для выполнения всех задач масс-спектрометрии изотопного состава (IRMS) в одном анализаторе? Системы IRMS Thermo Scientific ™ Delta V ™ сочетают выдающуюся чувствительность с превосходной линейностью и стабильностью для решения таких разнообразных задач, как анализ δ 13 C ПАУ в почве, 15 N / 14 мониторинг N в производных хлорофилла и обнаружение примесей меда. Системы Delta V могут быть оснащены широким спектром устройств и входов для подготовки проб, включая предварительные концентраторы, элементные анализаторы, интерфейсы ГХ, интерфейсы ЖХ и входы непрерывного потока, чтобы ваша приборная система идеально подходила для вашего применения.
Thermo Scientific ™ Delta V ™ IRMS обеспечивает выдающуюся чувствительность до 800 молекул на ион CO 2 в режиме двойного входа и 1100 молекул на ион в режиме непрерывного потока. Оптоволоконная передача сигнала устраняет контуры заземления и другие источники электронного шума. Можно сконфигурировать до 10 коллекторов в широком диапазоне, поддерживая все приложения МС с соотношением изотопов в диапазоне масс до m / z 96, при этом диапазон одновременных масс составляет до ± 25%. Система также предлагает возможность он-лайн H 2 .
Thermo Scientific ™ Delta V ™ IRMS совместим с широким спектром систем подготовки проб и впускных систем, таких как предварительные концентраторы, элементные анализаторы, интерфейсы ГХ, интерфейсы ЖХ и впускные отверстия для непрерывного потока.
Delta V доступен в двух вариантах: Delta V Advantage и Delta V Plus. Delta V Advantage IRMS может использоваться во всех стандартных приложениях, включая анализ H 2 при нагрузке газом-носителем гелием. Delta V Plus IRMS предоставляет дополнительные функции для более широкого круга приложений.Существует возможность полной модернизации системы Delta V Advantage до системы Delta V Plus, если аналитические потребности увеличиваются.
Отправляясь в плавание по извилистым водным путям дельты Калифорнии
Доктор Питер Мойл, почетный профессор Калифорнийского университета, факультет дикой природы, рыб и биологии охраны природы Дэвиса, изучал дельту на протяжении десятилетий. Доктор Мойл сосредоточил свои исследования на находящейся под угрозой исчезновения корюшки Дельты и других рыб Суисунского болота, района, который, по его словам, «больше всего напоминает первоначальную Дельту».Он не сомневается в том, что, каким бы ни был путь вперед, серьезные перемены неизбежны.
«Дельта — это совсем другая система, чем она была 150 лет назад, даже 50 лет назад. Он будет постоянно меняться », — сказал он. «То, с чем мы сейчас живем, — это временное состояние, и людям нужно выяснить, как они хотят, чтобы система выглядела на самом деле».
Есть бесконечные возможности того, как это могло бы выглядеть, от попыток максимально сохранить статус-кво до своего рода экологической рекультивации открытых водных путей и болот.Все хотят спасти Дельту, но какая версия Дельты заслуживает спасения? И кому лучше всего подходит эта Дельта?
Плыть в дельту реки — мечта по ветру; отплытие — это утомительное занятие с наветренной стороны. На лето мы арендовали слип в гавани Совы-Харбор на острове Твитчелл (который, по словам доктора Мойла, в ближайшие десятилетия может оказаться под водой). Ослепительно жарким июльским вечером пятницы, после того, как мы поехали на выходные на воду, мы сидели в кабине лодки, пока солнце садилось и поднимался ветер, а небо светилось оранжевым светом; В тот день было 110 градусов тепла, а на следующий обещал быть еще жарче.Мы наблюдали, как пара ласточек нервничает из-за нашей близости к их гнезду, ненадежно построенному под одной из солнечных панелей нашей лодки. Птицы, казалось, спорили о том, как лучше действовать.
«Какое опасное место для гнезда», — подумали мы, обсуждая вероятность того, что из их яиц вылупятся яйца, прежде чем мы отплыли, надеясь, что они выживают, несмотря на сомнительный выбор дома.
Когда мы вернулись через пару недель, температура упала, гнездо опустело, ласточек не было.Мы осторожно прошли по узкому каналу, избегая неглубоких участков и морской травы, миновали давно заброшенные, полузатонувшие лодки, окруженные агрессивным водным гиацинтом, а затем и мы.
Подписывайтесь на New York Times Travel в Instagram , Twitter и Facebook . А подпишитесь на нашу еженедельную рассылку Travel Dispatch , чтобы получать советы экспертов о том, как путешествовать умнее и вдохновлять на следующий отпуск.Мечтаете о будущем отпуске или просто о путешествии в кресле? Ознакомьтесь с нашим списком 52 мест на 2021 год .
Масс-спектрометрия в исследованиях химии белковых тиолов и передачи сигналов: возможности и предостережения
Abstract
Масс-спектрометрия (МС) стала мощным и широко используемым инструментом в исследовании химии, биохимии и биологии белковых тиолов. Очень ранние биохимические исследования метаболических ферментов выявили широкий спектр профилей реактивности, которые отличают цистеиновые тиолы с функциями катализа и стабильности белка от других остатков цистеина в белках.Разработка методов масс-спектрометрии для анализа белков с использованием ионизации электрораспылением (ESI) или матричной лазерной десорбции / ионизации (MALDI) в сочетании с появлением масс-анализаторов высокого разрешения сыграли важную роль в продвижении исследований модификаций тиолов, как в отдельных случаях. белки и в клеточном контексте. В этой статье рассматривается аппаратура МС и методы анализа, используемые при исследовании тиолов и их реакционной способности по отношению к ряду малых биомолекул. Отобранное количество исследований подробно описано, чтобы выделить преимущества, обеспечиваемые технологиями МС, а также оговорки, связанные с этими анализами.
1. Введение
Масс-спектрометрия (МС) — важная аналитическая технология, которая применяется практически во всех областях наук о жизни и за ее пределами. Хотя основные физические и химические принципы МС были известны с 19 — века [1], эта технология применялась в биологических науках только в течение последних нескольких десятилетий после разработки устройств, которые позволяют переносить полярные молекулы, такие как белки, нуклеиновые кислоты и метаболиты из жидкого и твердого состояний в газовую фазу в процессе ионизации.Этот скачок в инженерии был совершен Джоном Фенном (США) и Коичи Танака (Япония), получившими Нобелевскую премию по химии в 2002 г. за разработки в области ионизации электрораспылением (ESI) и мягкой лазерной десорбции (SLD) соответственно. ESI сегодня повсеместно используется в исследованиях биомолекул, а развитие SLD облегчило более позднее применение ранее известной матричной лазерной десорбции / ионизации (MALDI) для анализа белков, которая также широко используется по сей день [2, 3].
В последующие годы усилия бесчисленных исследователей расширили области применения МС до секвенирования белков, идентификации метаболитов, подтверждения последовательностей генов, изучения того, как нуклеиновые кислоты и белки модифицируются после их биосинтеза, обнаружения не ковалентные белковые комплексы [4] и даже измерение интактных микроорганизмов, таких как вирусы [5].Полезность МС для секвенирования и идентификации белков была установлена в 1993 году с первым отчетом о массовом фингерпринте пептидов, а относительная количественная оценка белков и пептидов на основе МС была достигнута несколько лет спустя с использованием аффинных меток, кодированных изотопами (ICAT) [ 6, 7]. Начиная с разработки ICAT, который усиливает реактивность биотин-йодацетиловых реагентов с цистеиновыми тиолами, мечение изотопов стало отличительной чертой количественной оценки с помощью MS и лежит в основе таких подходов, как мечение стабильных изотопов аминокислотами в культуре клеток (SILAC) [ 8], тандемные массовые метки (TMT) [9] и изобарические метки для относительной и абсолютной количественной оценки (iTRAQ) [10], среди прочего.Сегодня анализ белков и пептидов с помощью рассеянного склероза предоставляет обширную информацию, позволяющую исследовать структуру, реактивность и функцию белков. Более того, использование MS в области протеомики привело к идентификации новых белков, количественной оценке экспрессии белка и посттрансляционных модификаций (PTM), выяснению компонентов в сложных белковых ансамблях и проверке геномных последовательностей (т. Е. Протеогеномике). ).
Динамическая посттрансляционная модификация (PTM) белков особенно важна для регуляции биологических процессов и привела к созданию специализированных областей протеомики, ориентированных на идентификацию и количественную оценку селективных PTM, таких как фосфорилирование, гликозилирование, метилирование и другие.Модификации тиолов цистеина также все чаще признаются в качестве критических регуляторных переключателей, контролирующих функцию белка. Однако методы исследования Цистиоловых ПТМ в отдельных рекомбинантных белках и во всем клеточном контексте менее надежны, чем для других ПТМ, и отмечены существенными оговорками, которые следует учитывать для обеспечения точных результатов. Исследования тиоловых ПТМ затруднены из-за лабильного характера модифицированных цистеинов, часто требующего дериватизации, чтобы обеспечить стабильную, детектируемую часть и предотвратить последующие реакции, в том числе в виде артефактов во время лизиса клеток и обработки образцов.Для решения этих экспериментальных проблем требуются тщательные параллельные контрольные эксперименты, которые отмечены в этом обзоре.
Тиоловый протеом является мишенью для многочисленных разнообразных окислительных модификаций, происходящих из взаимодействий остатков Cys с химически активными формами азота, кислорода и серы, что приводит к более чем 15 модификациям (). Многие окисленные продукты Cys обратимы и / или впоследствии реагируют с другими производными цистеина с образованием более термодинамически стабильных продуктов.Некоторые модификации обычно считаются необратимыми, однако для некоторых известны механизмы репарации, катализируемые ферментами [11, 12]. Конкретным примером, более подробно описанным далее в этом обзоре, является восстановление пероксиредоксина (Prx) сульфиновой кислоты до активной тиоловой формы, катализируемое сульфиредоксином (Srx) в присутствии АТФ, Mg 2+ и тиоредоксина / система тиоредоксинредуктазы (Trx / TrxR). Продолжительность жизни обратимых тиоловых модификаций контролируется локальной структурой белка и микроокружением, в котором находится модифицированный Cys.Факторы, влияющие на микроокружение, включают 1) вариации уровней окислителя и антиоксиданта в окружающей области и 2) близлежащую конформацию и внутреннюю химию (нуклеофильность, кислотность, основность) соседних аминокислот. Вместе эти физические и химические среды влияют на химическую стабильность модифицированного Cys, что требует тщательной оптимизации подготовки образцов перед анализом МС, детали которого обсуждаются в следующих разделах.
Таблица 1
Степени окисления и дельта-массы продуктов окисления цистеина
2.Основы МС для исследования белков и пептидов
В этом разделе представлен краткий обзор принципов МС и основных компонентов приборов МС. Современная аппаратура МС характеризуется такими параметрами, как простота использования, универсальность, точность измерения массы, чувствительность и простота автоматизации. Компоненты масс-спектрометров будут кратко обсуждены в свете подходов на основе МС, наиболее часто используемых для изучения тиоловых модификаций в белках и пептидах, которые кратко описаны в.За дополнительными подробностями читатель может обратиться к другим статьям по этой теме ([13] и др.).
Подготовка и отделение образцов
Для анализа белков и пептидов часто требуются чистые образцы, не содержащие детергентов и солей, которые мешают ионизации. В частности, соли Na + и K + обеспечивают сильное ионное спаривание в режиме положительных ионов и могут приводить к фракционированию сигнала и подавлению ионов. По этим причинам образцы часто обессоливают перед МС-анализом или готовят в летучих буферах, таких как ацетат аммония и бикарбонат аммония, которые легко теряются в виде аммиака, уксусной кислоты, диоксида углерода и воды в процессе десольватации [18].Анализ отдельных биомолекул обычно выполняется путем прямой инфузии после обессоливания, но для более сложных смесей требуется разделение по одному или нескольким параметрам перед анализом МС. Методы препаративного масштаба для фракционирования образцов включают ВЭЖХ и гель-электрофорез и часто требуют последующих этапов обработки для подготовки образцов для МС. В качестве альтернативы аналитическое разделение выполняется с использованием модулей, подключенных к MS; Таким образом используются ВЭЖХ (и связанные с ней инструменты наноЖХ) и капиллярный электрофорез (КЭ).Подходы ЖХ, используемые в приложениях протеомики, включают разделение на основе дифференциальной полярности (например, жидкостная хроматография с обращенной фазой (RPLC) [14] и жидкостная хроматография гидрофильного взаимодействия (HILIC) [15, 16]) или по заряду (ионный обмен). Последние достижения в области ЖХ позволяют проводить двухмерное разделение, часто с прямым ионным обменом с последующей RPLC. Кроме того, разделение ионов на основе их подвижности в газовой фазе стало полезным аналитическим инструментом [Ion Mobility MS, IMS-MS] (недавние подробные обзоры см. В [17, 18]).В IMS-MS ионы мигрируют через однородное электрическое поле к детектору, где ионы с большей площадью поверхности подвергаются большему количеству вызывающих трение столкновений с инертным газом по сравнению с ионами меньшего размера, замедляющими их движение. Экспериментальные данные IMS сравниваются с известными архитектурами белков и вместе с данными MS предоставляют ценную структурную информацию [19].
Источники ионизации
ESI / NSI VS MALDI
Приложения протеомики чаще всего полагаются на источники ионов MALDI и ESI, которые обеспечивают более «мягкие» средства ионизации по сравнению с методами электронной ионизации или химической ионизации.Используя MALDI, ионы вводятся в масс-спектрометр посредством индуцированной УФ-лазером ионизации матрицы, содержащей образец, что облегчает ионизацию образца. Источники MALDI часто генерируют +1 заряженные ионы (обозначаемые как [M + H] + или [M + Na] + ), что для белков и пептидов обычно приводит к очень большим отношениям m / z. Совсем недавно MALDI был применен для пространственного анализа образцов ткани в методе, называемом MALDI imaging MS [20], в котором срезы тканей обрабатываются матрицей MALDI и облучаются в небольших конкретных областях, представляющих интерес, с получением масс-спектра для каждого набора. x, y-координат на срезе ткани.Поскольку подробное обсуждение MALDI-визуализации МС выходит за рамки этого обзора, читатель может обратиться к недавней статье об этом подходе [21].
Источники ESI обеспечивают более прямую ионизацию, когда раствор аналита вводится в источник через капилляр из плавленого кварца или нержавеющей стали, через который подается напряжение. Образуется туман заряженных капель, из которого испаряется растворитель, уменьшая размер капель, что приводит к взрыву капель и появлению множества небольших газообразных ионов [22].В отличие от MALDI, ESI обеспечивает ионы с более высокими зарядовыми состояниями, что приводит к более сложным масс-спектрам, но снижает отношения m / z, что обеспечивает совместимость с более широким спектром анализаторов MS. Ионизация наноспреем (NSI, также сокращенно nanoESI) была позже разработана по принципу ESI, но использует уменьшенные скорости потока (нл / мин) и напряжение распыления, что требует меньшего количества образца без потери чувствительности. По характеру более низкой скорости аналита и уменьшенного диаметра капилляров капли, производимые NSI, значительно меньше, чем капли, произведенные ESI: 1–2 мкм в диаметре для ESI и <200 нм в диаметре для NSI [18].NSI требует использования приборов наноЖХ, которые разделяют аналиты на колонках, обычно сделанных из плавленого кварца с внутренним диаметром 50–150 мкм, со слоем длиной 10–250 мм, заполненным требуемой средой для разделения целевых аналитов и с использованием скорости потока ниже 1 мкл / мин. обычно от 200 до 400 нл / мин.
Масс-анализаторы
Масс-анализаторы, обычно используемые в приложениях протеомики, включают квадруполи, времяпролетные (TOF), ионные ловушки и новые анализаторы орбитальной ловушки. Относительная производительность масс-анализаторов зависит от таких параметров, как скорость сканирования, точность измерения массы, разрешение, диапазон сбора данных и возможности тандемного анализа.Времяпролетные масс-анализаторы измеряют время, необходимое для прохождения ионов от источника до детектора, и предлагают высокий верхний предел обнаружения (~ 10 000 м / з), что делает их особенно хорошо подходящими для исследования интактных белков. особенно с источниками MALDI. Получение тандемных масс-спектров (МС / МС) с использованием анализатора TOF требует использования квадрупольного или дополнительного фильтра масс TOF (Q-TOF, TOF-TOF). Напротив, ионные ловушки используют магнитное и электрическое поля для захвата и выброса ионов за счет возрастания отношения m / z.По сравнению с тройными квадруполями ионные ловушки могут быть менее чувствительными для количественной оценки, но они меньше, дешевле и также служат в качестве коллизионных ячеек для тандемной фрагментации МС [22]. Кроме того, ионные ловушки при использовании в тандемной МС ограничены тем, что их нижний предел обнаружения составляет одну треть отношения m / z иона-предшественника (например, выделение и фрагментация иона с m / z 600 дает много дочерних ионов; наблюдаются только те, что> 200 m / z). Орбитальная ловушка, разработанная за последние 15 лет, представляет собой импульсный масс-анализатор, который использует статические электрические поля для улавливания и анализа ионов, предлагая повышенную чувствительность и разрешение наряду с высокой точностью определения массы [23].
Поскольку методы ионизации MALDI, ESI и NSI недостаточны для фрагментации большинства молекул, получение соотношений m / z для всех ионов, попадающих в детектор, обеспечивает спектр MS 1 (ионов-предшественников), из которого высокое разрешение масса, молекулярная формула и, в некоторых случаях, относительное количественное определение. Информация, касающаяся пептидной последовательности, выяснения структуры и разрешения сайта модификаций, требует тандемного анализа MS, в котором ион-предшественник выделяется и фрагментируется с образованием многочисленных фрагментных (или дочерних) ионов.Изоляция ионов-предшественников осуществляется квадруполями или ионными ловушками, а фрагментация чаще всего вызывается диссоциацией, индуцированной столкновениями (CID), столкновительной диссоциацией с более высокой энергией (HCD) или диссоциацией с переносом электрона (ETD) () [24–26] . Выбор метода фрагментации в конечном итоге зависит от типа присутствующих модификаций, включая PTM и изотопные метки для количественного определения, а анализ образцов обычно включает использование нескольких различных методов фрагментации для получения дополнительных спектров MS 2 .
Пептидные связи, расщепленные фрагментацией MS 2 . A : Диссоциация, вызванная Коллисоном, фрагментирует амидную связь, образуя b- и y-ионы. Также нацелены на многие PTM (не показаны). B : Диссоциация с переносом электрона нацелена на связь амино-α-углерод, образуя c- и z-ионы, и оставляя PTM нетронутыми.
3. Структурные и кинетические исследования тиоловых белков с помощью MS
В этом разделе описаны различные применения МС (в первую очередь ESI-TOF MS, но также и другие) для исследования химии и кинетики, лежащих в основе образования, реактивности и восстановления оксоформы тиоловых белков с использованием интактные тиолсодержащие белки как модельные системы.Также обсуждаются методы МС для идентификации структурных изменений, вызванных окислительными модификациями тиолов в белках. Мы используем термин «оксоформа» для обозначения тиоловых модификаций, приводящих к увеличению степени окисления серы по сравнению с цистеиновым тиолом (-2). Кроме того, обсуждаемые модификации будут сокращены в формате Cys-SR, где «S» представляет собой цистеиновую серу, а «R» представляет собой добавленный фрагмент.
Общие соображения и модельные системы для исследования химии тиоловых белков
Исследования интактных белков с использованием ESI-MS предоставляют важную и подробную информацию, но имеют несколько важных технических требований, хотя не все они относятся к исследованиям белка цистеина.Во-первых, в среднем для приемлемого отношения сигнал / шум необходимо минимум 20-40 мкг чистого белка. Это значение изменяется в зависимости от массы и уникальной эффективности ионизации каждого белка, но это значительно больше материала, чем требуется для вестерн-блоттинга (<1 мкг чистого белка). Тем не менее, более высокий уровень химической информации можно получить с помощью МС по сравнению с Вестерн-блоттингом или путем разделения белка с помощью нативного или денатурирующего гелевого анализа. Как указывалось ранее, образцы не должны содержать солей и детергентов, что требует, чтобы белок (белки) или реакционная смесь выдерживали летучие буферы и / или слабокислый pH (4–5), используемый для облегчения ионизации в режиме положительных ионов.Для целей количественной оценки необходимо учитывать эффективность ионизации всех частиц, образующихся в ходе реакции, так что изменение содержания конкретного иона может быть связано с его относительной концентрацией в растворе. Такая стандартизация выполняется путем анализа известных количеств каждого соответствующего вида в чистом виде, когда это возможно, или путем подгонки данных реакции к теоретическим кинетическим моделям. Важным предостережением, которое следует учитывать, является возможность протекания газофазных реакций во время МС-анализа, например, известных для равновесия цистеинсульфеновая кислота / сульфенамид и окисления остатков метионина, среди прочего [27, 28].Эти технические требования не помешали ESI-MS быть мощным инструментом для идентификации новых видов белков и измерения скорости белковых реакций.
Многочисленные продукты окисления Cys можно классифицировать на основании их относительной термодинамической стабильности. Менее стабильные формы, в том числе сульфеновая кислота (Cys-SOH) и нитрозотиол (Cys-SNO), реактивны по отношению к множеству богатых электронами молекул, но также потребляются окружающими элементами, такими как кислород (Cys-SOH) и свет (Cys-SNO). ).Полустабильные формы, включая дисульфиды и тиосульфинаты, нестабильны по отношению к восстановителям и некоторым нуклеофилам, но в остальном являются долгоживущими. К наиболее термодинамически стабильным продуктам окисления Cys относятся сульфиновая и сульфоновая кислоты, необратимость которых доказана, за исключением особого случая восстановления пероксиредоксинсульфиновых кислот сульфиредоксином. Восстановительная обратимость менее стабильных оксоформ серы подразумевает роль окислительно-восстановительных переключателей, способных влиять на передачу сигналов в клетке, активируя и дезактивируя пути ферментов.Однако их прямой анализ затруднен из-за их короткого времени жизни в белках дикого типа и общих ограничений в получении значительных количеств чистых эукариотических белков. По этим причинам в последние десятилетия появился ряд модельных белковых систем, обеспечивающих средства для анализа тиолоксоформ с помощью МС и других методов. Мы кратко обсудим их вместе с некоторыми конкретными примерами нашей работы с использованием таких моделей и методов.
Для наблюдения цистиолов в менее стабильных оксоформах используется несколько общих стратегий.Во-первых, это использование белка дикого типа и дериватизирующего агента, селективного по отношению к конкретным видам, производным от Cys. Этот тип подхода был первоначально использован Эллисоном для идентификации образования цистеинсульфеновой кислоты в GAPDH посредством реакции образующегося Cys-SOH с димедоном [29]. Другими белками дикого типа, используемыми таким образом, являются папаин и альбумины сыворотки крови человека и крупного рогатого скота. Папаин, цистеиновая протеаза из папайи, содержит 8 Cys в зрелой форме, хотя 6 обнаружены в дисульфидных связях. Тиолат Cys25 в активном центре образует Cys-SOH в присутствии H 2 O 2 [30] и пертиола (Cys-SSH, также называемого персульфидом) в ответ на H 2 S [31] и имеет таким образом, был использован для исследования реакционной способности и кинетики этих видов [32, 33].Сывороточный альбумин (69 кДа), основной антиоксидантный белок в плазме, имеет только один из 35 остатков Cys, присутствующих в виде свободного тиола. Было показано, что этот Cys является редокс-чувствительным как в человеческом (HSA), так и в бычьем (BSA) белках путем окисления до соответствующих сульфеновых кислот, продуктов S, -глутатионилированных (Cys-SSG) продуктов [34, 35] и в случае БСА персульфид Cys-SSH [35].
В отличие от эукариотических белков дикого типа, которые содержат много остатков Cys и других нецистеиновых PTM, ряд групп разработали мутантные белки, часто из прокариот или низших эукариот, которые предлагают простоту снижения содержания Cys, немного или совсем нет. модификации и более низкий молекулярный вес.Мутации, в том числе расщепляющие Cys или все остатки Cys, за исключением перекисного сайта, обеспечивают метод довольно количественного получения конкретной оксоформы и исследования ее реакционной способности. Например, цистеин в активном центре метионинсульфоксидредуктазы (Msr, 23 кДа) образует Cys-SOH на пути к выработке восстановленного метионина [36, 37]. Мутации от цистеина к серину в свободном Msr (fRMsr, который восстанавливает свободный Met- ( R ) -сульфоксид) из E. coli , продуцируют единственный Cys118-содержащий мутант Cys84Ser / Cys94Ser, используемый для измерения захвата Cys-SOH с помощью химические зонды [32].Кроме того, цистеиновые мутанты фактора транскрипции OxyR (34 кДа, E. coli ), пероксиредоксина AhpC (21 кДа, S. typhimurium ) и глутатионпероксидазы (Gpx, дрожжи, 18 кДа), оставляя только пероксидный цистеины (Cys208Ser OxyR; Cys165Ser или Cys165Ala AhpC; Cys82Ser Gpx) были использованы в качестве модельных систем для исследования реактивности Cys-SOH [32, 38-40], Cys-SNO [39-42], Cys-SO 2 H [39, 40], Cys-SSH [33] и другие формы. Типичные масс-спектры ESI-TOF нескольких оксоформ цистеина wt и мутантного AhpC показаны на рис.
Типичные масс-спектры ESI-TOF для AhpC дикого типа и мутанта из S. typhimurium . A : AhpC дикого типа наблюдается как мономер (20 616 а.е.м.) и гомодимер (41 230 а.е.м.). B : мутант Cys165Ser AhpC, лишенный разрешающего Cys, позволяет легко получать множество оксоформ. R-SN относится к циклическому сульфенамиду Cys-SN, образованному в результате опосредованной внутримолекулярным амином дегидратации Cys-SOH в газовой фазе и / или в растворе. Неопубликованные данные.
Обзор методов исследования кинетики ферментов с помощью MS
Кинетические исследования реакций интактных белков с помощью ESI-MS требуют средств гашения протекающей реакции в определенные моменты времени. Химическое гашение путем добавления дестабилизатора, такого как кислота, основание или хаотропный реагент, останавливает реакцию, но может вызвать необходимость в стадии очистки образца перед анализом МС. Альтернативный подход к гашению реакции включает прохождение через смолы для исключения размера, обеспечивающие одновременное удаление низкомолекулярных компонентов и буферных солей, замену белкового раствора на летучий буфер, поддающийся ионизации и МС ().Для реакций, протекающих достаточно быстро, чтобы потребовать анализа моментов времени менее 30 секунд, был разработан ряд методов с временным разрешением (TR-MS) с использованием десольватации ESI в качестве средства гашения реакций. В этом подходе два шприца, приводимые в действие одним и тем же насосом, соединяются смесительным тройником с общим капилляром из плавленого кварца, по которому реакционный раствор поступает к источнику ESI (). В один шприц загружают интересующий белок, а в другой — субстрат. Два раствора смешиваются вместе в тройнике с переменной скоростью, чтобы инициировать реакцию, часто достаточно высокую, чтобы гарантировать турбулентное перемешивание, хотя некоторые методы были описаны с использованием ламинарного потока [43–45].Объем трубки от смесительного тройника до источника ионов и скорость потока позволяют определить время реакции до десольватации и гашения смеси; Изменения скорости потока изменяют время реакции, обеспечивая анализ реакционной смеси в различные моменты времени и всего за 7 миллисекунд [46]. Импульсный поток, остановленный поток и непрерывный поток подходы TR-MS были применены к исследованиям конформации и структуры белка [47–49], исследованиям ферментативных механизмов и идентификации промежуточных продуктов [46, 50–54] и другим кинетическим исследованиям. исследования [55, 56].Недавно десорбционный ESI (DESI) был использован в качестве источника ионизации для TR-MS, обеспечивая улучшенный уровень разрешения по времени [57]. Источник DESI отбирает образцы из струйного потока жидкости, протекающего с большей скоростью, чем это совместимо с источниками ESI, и десольватируется быстрее, чем ESI, что позволяет анализировать более ранние и более частые моменты времени, чем это было возможно ранее [57, 58]. Некоторые из этих методов были применены для исследования механизмов и кинетики белков, основанных на химии тиолов, и далее мы рассмотрим несколько тематических исследований.
Прерывистые и непрерывные методы кинетических исследований с использованием интактных белков. A : Прерывистые подходы включают аликвоты реакции гашения с использованием условий низкого pH или смолы для исключения размера с последующим анализом масс-спектрометрии каждой временной точки. B : Для непрерывного кинетического анализа с временным разрешением требуется прямая инфузия с использованием двойных шприцев с реакцией, происходящей между смесительным тройником и источником ионов (так называемая петля старения). Пошаговое изменение скорости потока позволяет наблюдать за несколькими временными точками реакции за один проход.Эти методы позволяют одновременно контролировать все ковалентные и в некоторых случаях нековалентные частицы, присутствующие в каждый момент времени реакции.
MS для исследования реакционной способности тиолоксоформ
Давний интерес к области окислительно-восстановительной биологии на основе тиолов заключается в разработке методов, которые позволяют количественно измерить модификацию тиола цистеина физиологически релевантными молекулами (например, H 2 O 2 , GSH, NO, HNO и т. Д.). Хотя некоторые из этих модифицированных частиц измеряются непосредственно с помощью МС или других методов, традиционно химические зонды использовались для селективного нацеливания и с высокой реакционной способностью различных оксоформ цистеина.Примером этого является использование 7-хлор-4-нитробензо-2-окса-1,3-диазола (хлорид NBD) для дериватизации белковых сульфеновых кислот для обнаружения с помощью УФ-видимой спектроскопии [59]. Хотя этот реагент не является селективным для модификации Cys-SOH, его реакция с Cys-SOH дает сигнал, уникальный для этой оксоформы Cys. В последние годы наша группа и другие использовали модельные тиоловые белки, такие как те, которые обсуждались и упоминались ранее в этом разделе, чтобы помочь в разработке химических зондов с селективностью в отношении определенных тиоловых оксоформ, для количественной оценки реактивности и устранения неполадок в рабочих процессах для маркировки, обогащение и обнаружение с помощью МС различных модифицированных тиолом субпротеомов, генерируемых -сульфенилированием белка S (Cys-SOH), S -сульфгидрацией (Cys-SSH), S -нитрозилированием (Cys-SNO) и S -ацилирование (например,g., S -пальмитат) в клетках.
Например, в недавней публикации мы задались вопросом, реагируют ли широко используемые тиол-блокирующие реагенты йодацетамид (IAM), N, -этилмалеимид (NEM) и метилметантиосульфонат (MMTS) с белком Cys-SOH, и если да, то какие последствия таких реакций будут заключаться в способности обнаруживать Cys-SOH в более широком контексте окислительно-восстановительного протеома с использованием химических зондов или подходов с непрямым переключением меток. Во-первых, кинетический профиль был установлен путем взаимодействия Cys165Ser AhpC-SOH с каждым из трех электрофилов.Используя анализ интактного белка на основе ESI-TOF, мы обнаружили и количественно оценили кинетику образования ковалентного аддукта (называемого Cys-SOR), образованного в результате реакций AhpC-SOH с каждым из трех электрофилов, выявив следующий профиль скорости: MMTS> NEM> IAM. Непредвиденные продукты реакции были также идентифицированы с помощью MS-анализа, такие как превращение Cys в дегидроаланин (DHA) в случае MMTS () [39]. Кроме того, мы дополнительно использовали ESI-TOF MS для исследования стабильности видов Cys-SOR в восстанавливающих условиях (т.е.е., в присутствии DTT, TCEP или аскорбата), критический шаг в подходах с переключением меток для исследований окислительно-восстановительной протеомики. Как показано на фиг.1, за восстановлением Cys-SOR до Cys-SH следовала ESI-TOF MS, показывающая, что извлечение Cys165Ser AhpC-SH зависело как от блокирующего реагента, так и от используемого восстановителя. Общее влияние этой недавно детализированной химии на результат экспериментов зависит от интересующих видов цистеина в каждом конкретном эксперименте. Например, общие подходы к окислительно-восстановительной протеомике, сфокусированные на количественной оценке глобального соотношения восстановленного и окисленного цистеина, скорее всего, не пострадают, поскольку после мечения используется стадия восстановителя.Напротив, исследованиям, направленным на мечение Cys-SOH, например с димедоном и родственными реагентами, будет препятствовать перекрестная реактивность реагентов, блокирующих тиол, в конкуренции с зондом, нацеленным на Cys-SOH. В наших исследованиях MMTS наиболее быстро реагировал с AhpC-SOH, но соответствующий аддукт также был наиболее чувствительным к восстановлению. Конечно, предполагается, что относительная стабильность таких аддуктов также зависит от белка и микроокружения. Эти результаты вызвали озабоченность по поводу косвенных методов обнаружения оксоформ тиоловых белков и указали на необходимость продолжения разработки зондов, которые химически нацелены на специфические модификации тиолов.
Мутант AhpC-SOH легко захватывает электрофилы иодацетамид (IAM), N, -этилмалеимид (NEM) и метилметантиосульфонат (MMTS), широко используемые для блокирования тиолов для получения ковалентных аддуктов Cys-SOR (точная связь неизвестна). Последующая обработка восстановителем (тиолами, фосфинами, аскорбатом) превращает Cys-SOR в Cys-SH в различной степени. [DHA: дегидроаланин.] По материалам Reisz et al. [39]
Подобные подходы, основанные на МС, были использованы для исследования реакции белка Cys-SOH с различными химическими зондами и для проверки селективности зондов по отношению к Cys-SOH по сравнению с их реакцией с Cys-SH, Cys- СНО и межмолекулярные дисульфиды.Эти химические зонды включают димедон и его производные [29, 60–62], S, -этилциклопентадионы [63], ациклические β-кетоэфиры [64] и самые последние бициклононины [40].
MS-анализ сульфенилирования белков в интактных белках
Сульфеновые кислоты (Cys-SOH) являются исходными продуктами, возникающими в результате окисления остатков Cys в белках в физиологических или стрессовых условиях. Анализ образования Cys-SOH легко доступен с помощью ESI-MS путем мониторинга Δ массы 16 Да относительно родительского белка.В большинстве случаев образование Cys-SOH на модельных белках, обсуждавшихся ранее (например, при обработке H 2 O 2 ), сопровождается более окисленными продуктами, включая сульфиновые (Cys-SO 2 H, + 32 Да) и / или сульфоновые кислоты (Cys-SO 3 H, +48 Да), полученные либо в результате окислительного стимула, во время обработки образца [65], либо в газовой фазе во время сбора данных МС [66]. Кроме того, мы и другие исследователи наблюдали в MS-анализе интактных белков, содержащих Cys-SOH, сигнал, соответствующий продукту циклического сульфенамида (Cys-SN), продукту дегидратации Cys-SOH, который известен для окислительно-восстановительных белков, таких как PTP1B. (структура показана на) [67, 68].В наших руках отношения сигналов Cys-SN к Cys-SOH, наблюдаемые с помощью интактного белка MS, широко варьируют для разных белков [32, 39, 69]. Без четкого определения того, образуется ли Cys-SN в растворе или в газовой фазе, обычно эти частицы рассматриваются вместе как Cys-SOH. Можно отличить образование Cys-SN в растворе от газовой фазы для данного белка, следуя экспериментальному плану, в котором пероксид водорода, меченный 18 O, используется для генерации Cys-S 18 OH () и используя преимущества недавно открытой реакции Cys-SOH с иодацетамидом.Понимание функциональной роли белковых сульфенамидов все еще находится в зачаточном состоянии, но было описано как возможный защитный механизм против гиперокисления Cys (например, Prxs) [69] или как петля цепи для поддержания передачи сигналов. Используя ESI MS, биохимические эффекты сульфенилирования были приписаны множеству целевых белков [38, 65, 70–72] и привели как к усилению [73], так и к потере функции [74–76].
Равновесие гидратации-дегидратации сульфеновой кислоты и сульфенамида. 18 O-мечение сульфенамида приводит к образованию 18 O-меченых форм Cys-SOR. Равновесие SOH / SN в фазе раствора будет снижаться. 18 включение O в пользу более обильного 16 O.
MS-анализ S-сульфгидратации белка в интактных белках
Термин белок S -сульфгидратация был недавно введены для определения ПТМ, возникающего на остатках Cys при их взаимодействии с газотрансмиттером сероводорода (H 2 S) (e.g., персульфид и пертиол). Однако до описания этого нового определения ПТМ персульфиды были известны как ключевые промежуточные продукты в каталитических циклах различных ферментов, включая цистеин-десульфуразу и серотрансферазу, а также как лиганды в кластерах Fe-S [77–80]. Прямой анализ модификации персульфида с помощью МС был предпринят с несколькими интактными белками [33]. Персульфиды белков характеризуют с помощью МС по появлению сигнала Δ масс +32 Да относительно исходного тиола.Несмотря на значительный сдвиг массы, необходимы контрольные эксперименты, чтобы избежать ошибочной идентификации из-за того, что другие тиоловые ПТМ, такие как цистеинсульфиновая кислота (Cys-SO 2 H), также дают чистую Δ массу +32 Да (см. масс в). Даже при широком использовании приборов с высоким разрешением различение этих двух модификаций интактных белков остается проблемой, поскольку, например, для различения этих двух модификаций в белке 20 кДа потребуется точность измерения массы <1 ppm.Контрольные эксперименты в этом случае могут включать 1) обработку восстанавливающими агентами (например, DTT или TCEP) для получения восстановленной тиоловой формы анализируемого белка и / или 2) реакцию с реагентами, блокирующими тиол, такими как IAM или NEM [33, 78 , 79]. Оба они имеют решающее значение, поскольку они позволяют специфически отличать персульфиды от сульфиновой кислоты, отмечая, что последняя не может быть восстановлена DTT, а также инертна к обработке IAM и NEM [39]. В отдельных случаях и при определенных условиях образование персульфида сопровождается полисульфинилированием белка (Cys-S- (S) n -SH), что приводит к более высокому спектральному шуму и снижению спектрального качества.Например, окисление цистеин-сертрансферазы SufE персульфидом SufS-S-SH дает частицы с масс-аддуктами +32, +64 и +96 Да, что соответствует включению одной, двух и трех серы в родительский белок [78] . Хотя такие полисульфиды могут скрывать сигналы других присутствующих частиц, они также могут служить молекулярными отпечатками активных форм серы, в отличие от окисления, опосредованного исключительно ROS, с образованием Cys-SO 2 H. Интересно, что полисульфиды и персульфиды, как полагают, сосуществуют. биологически и служат носителями сигнала H 2 S [81].С этой целью очевидно, что анализ сульфгидратации белка с помощью МС возможен; однако устранение несоответствий в идентификации продукта требует: 1) использования инструментов МС с высоким разрешением и / или включения контрольных экспериментов; 2) исключение окислителей для поддержания полисульфидной серы в восстановленном состоянии.
MS-анализ белка
S -нитрозилирование в интактных белках (также называемое белком S -нитрозирование)S -нитрозотиолы (Cys-SNO) образуются в результате радикальной реакции сульфгидрильных групп (Cys- SH) и оксида азота (NO), в результате чего масса Δ равна +29.Дополнительный путь, посредством которого образуются белковые Cys-SNO, включает перенос нитрозильной группы от других белковых SNO или, чаще, от низкомолекулярных SNO, включая свободный Cys-SNO, S, -нитрозоглутатион (GSNO) и недавно открыли HSNO к остатку Cys целевого белка в процессе, называемом транснитрозированием (или транснитрозилированием) [82, 83]. Фрагмент SNO особенно нестабилен для анализа MS и может расщепляться во время ионизации. При разложении функциональной группы SNO образуется исходный предшественник сульфгидрильного радикала и NO.Несмотря на внутреннюю нестабильность S -нитрозотиолов, за последние два десятилетия многие белковые SNO были обнаружены с помощью ESI. Например, фермент гидролаза p21ras был проанализирован с помощью ESI / тройного квадруполя, хотя полученный спектр страдал от плохого отношения сигнал / шум [84]. Совсем недавно нитрозилированный Trx1 наблюдали с помощью ESI MS после тщательной оптимизации состава растворителя и параметров ионизации ESI [85]. Получение и МС-анализ двух нитрозилированных пептидов Trx1 модели S , близких к нейтральному pH (6.8) показал снижение вызванного МС денитрозилирования в присутствии хелаторов металлов, таких как ЭДТА или неокупроин. Однако настройка как напряжения конуса, так и энергии столкновения была критичной для минимизации нежелательной фрагментации Cys-SNO. После этих открытий включение хелатирующих добавок в буфер стало обычным явлением для пробоподготовки и МС-анализа нитрозилирования белков. Другими факторами, которые затрудняют анализ интактных S -нитрозилированных белков с помощью MS, являются: 1) присутствие модифицированных дисульфидом белков, образующихся во время приготовления белковых SNO, что может затруднить количественную оценку сигнала SNO; 2) пространственное транснитрозилирование соседних остатков Cys, которое может ограничивать точную идентификацию «целевого» сайта нитрозилирования.Последнее можно преодолеть путем введения реагентов, блокирующих тиол, для алкилирования свободных сульфгидрилов после нитрозилирования, как обсуждается в разделе 4. С этой целью очевидно, что необходима тщательная оптимизация экспериментальных параметров для полного использования МС для анализа S — нитрозилированные белки, обеспечивающие фундаментальную и дополнительную технологию для анализа функциональной роли сайтов Cys-SNO чистых белков [84, 86, 87].
MS-анализ -тиолирование белка
S в интактных белках-тиолирование белка S описывает ПТМ, в котором белковые цистеины связаны с низкомолекулярными тиолами посредством дисульфидной связи.Механически S -тиолирование происходит как по ферментативным, так и неферментативным путям, обычно с участием окисленного электрофильного цистеина (например, Cys-SSR, Cys-SOH, Cys-SNO) и восстановленного нуклеофильного тиола, хотя радикальные реакции тиильный и глутатионильный радикалы также приводят к тиолированию S . S -тиолирование происходит как часть нормальной физиологии и передачи сигналов во многих организмах; однако этот PTM в первую очередь наблюдается в ответ на стресс как механизм предотвращения необратимого и непоправимого белкового окисления белков и, в других случаях, в регуляции клеточной передачи сигналов [88]. In vivo , обычными продуктами тиолирования S являются производные глутатионила (Cys-SSG) и цистеинила (Cys-SSCys) и, в меньшей степени, продукт гомоцистеинила (Hcy) [89, 90]. Высокое содержание внутриклеточного GSH, от низкого до 10 мМ, делает S -глутатионилирование преобладающим видом S -тиолирования [91]. Методы, используемые для идентификации мишеней глутатионилирования на интактных белках, включают метаболическое включение радиоактивно меченного GSH ( 35 S), химически индуцированное образование дисульфидов с биотинилированным GSSG и обнаружение антителами против S -глутатионильного фрагмента [92].В последние годы были разработаны другие нерадиоактивные методы, использующие хемоферментное образование и включение ненативных / меченых субстратов глутатиона для идентификации мишеней глутатионилирования (например, меченный биотином S -глутатионил спермидин [93] и S -глутатионилазидоаланин [94]. Эти и другие методы позволили идентифицировать более 2200 мишеней глутатионилирования, обеспечивая понимание структурных мотивов, белкового домена и онтологии генов, связанных с -глутатионилированием S [95].Хотя эти методы очень полезны для идентификации и изучения мишеней S -глутатионилирования, другие ПТМ S -тиолирования менее изучены.
Общий анализ интактного белка S -тиолирования, включая S -цистеинилирование, S -гомоцистеинилирование и S -глутатионилирование, можно провести с помощью МС. Каждая модификация дает характеристическую массу, позволяющую просто идентифицировать чистые S -тиолированные белки даже с помощью МС с низким разрешением.Характерные масс-сигналы для S -цистеинилирования, S -гомоцистеинилирования и S -глутатионилирования от родительского белка составляют + 119, +143 и + 305 Да, соответственно. Для идентификации типа тиолирования S может быть проведен контрольный эксперимент, в котором образец обрабатывают DTT с последующим анализом масс-спектрометрии для отслеживания потери массы, соответствующей Cys, Hcy и GSH [96, 97]. Недавние успехи в подходах к МС, таких как нисходящая протеомика, позволяют анализировать тиолирование S в интактных отдельных белках и простых смесях, и недавно были продемонстрированы как глутатионилирование белка S , так и -цистеинилирование S [98 , 99].Недавние отчеты выдвинули на первый план несколько соображений, которые необходимо учитывать при анализе -тиоляции S с помощью нисходящей протеомики, например, при анаэробной работе с образцами белков и использовании поглотителей тиолов [100].
MS-анализ белка
S -ацилирование в интактных белкахS -Ацилирование белковых остатков цистеина представляет собой ковалентное включение цепи жирной кислоты в сульфгидрильную группу через тиоэфирную связь, катализируемое ферментами ацилтрансферазы.Типичным примером S -ацилирования белков является S -пальмитоилирование, которое представляет собой присоединение 16-углеродной цепи насыщенной жирной кислоты (пальмитиновой кислоты), хотя также могут быть включены другие жирные кислоты, такие как стеариновая и ненасыщенная олеиновая кислота. Количество анализов, которые позволяют обнаруживать интактные S -ацилированные белки с помощью MS, ограничено; однако некоторые из них предоставили полезную информацию при разработке надежных методов глобального анализа S -ацилированного протеома.Эти исследования показали, что -ацилирование S может приводить к множественным образцам жирных кислот, что означает, что белок может быть модифицирован по данному остатку Cys различными жирными кислотами, а также что несколько остатков Cys в одном и том же белке могут содержать одну и ту же жирную кислоту или другой набор жирных кислот [101]. Кроме того, ацилирование жирных кислот также наблюдалось на серине и на N-конце белка [102–104]. Анализ ацилированных интактных белков с помощью МС, обычно с использованием MALDI-TOF [44], часто сочетается с деацилированием интересующего образца для определения состава, содержания жирной кислоты (тип и%) и аминокислоты / жирной кислоты. связность цепи (O-, N- или S-связанная).Деацилирование обычно достигается с помощью щелочного метанолиза [105, 106], аминолиза с помощью гидроксиламина [107, 108] или гидрогенолиза [109] (). Используя условия деацилирования, можно выявить состав и содержание аддукта жирной кислоты, обычно в сочетании с идентификацией жирных кислот с использованием газовой хроматографии-МС (ГХ-МС). Аминолиз широко используется из-за хемоселективности гидроксиламина в отношении тиоэфира по сравнению с другими типами связей аминокислот / жирных кислот, однако недавний отчет показывает, что гидрогенолиз обеспечивает более воспроизводимое и точное количественное определение жирных кислот [109].
Стратегии деацилирования белков: A . щелочной метанолиз; В . аминолиз; С . гидрогенизация [FA = муравьиная кислота]
MS для исследования кинетики гиперокисления 2-цистеина пероксиредоксина и восстановления сульфиредоксином
Пероксиредоксины (Prx) — это очень распространенные антиоксидантные ферменты, чей активный центр Cys циклически проходит через многочисленные состояния окисления тиола во время восстановления H 2 O 2 к воде. Многие организмы имеют ферменты Prx, от бактерий до людей, и хотя их каталитические механизмы весьма схожи, они сильно различаются по своей склонности к образованию активного центра Cys-SO 2 H, который инактивирует пероксидазу.Кинетика окисления Prx, структурные факторы, лежащие в основе гиперокисления Prx, и катализируемая сульфиредоксином (Srx) репарация ферментов Prx дрожжей и млекопитающих исследуются с использованием множества методов, включая МС [11, 12]. Механизм репарации Srx зависит от Mg 2+ и АТФ, в первую очередь с участием образования сульфинового фосфорилового эфира Prx, который захватывается тиолом активного центра Srx с образованием смешанного тиосульфината Prx-Srx. В присутствии восстановителя на основе тиола тиосульфинат разрушается, высвобождая Prx-SOH и регенерируя активный Srx [110] ().
Каталитический цикл 2-Cys пероксиредоксинов и масс-спектры ESI-TOF, показывающие различия в чувствительности к гиперокислению между ферментами Prx2 и Prx3 человека в присутствии H 2 O 2 . [Prx: пероксиредоксин; Trx: тиоредоксин; TrxR: тиоредоксинредуктаза] Адаптировано из Haynes et al . [69].
MS был применен в нескольких различных экспериментальных установках для исследования кинетики гиперокисления Prx и восстановления с помощью Srx. Например, наша группа недавно сообщила о первом применении онлайн-TR-MS для исследования биохимии тиолов в исследовании дифференциальной устойчивости Prx 2 и 3 человека к гиперокислению / инактивации [69].Prxs 2 и 3 легко детоксифицируют H 2 O 2 до воды через активный центр 2-Cys, в котором пероксидный Cys (C p ) сначала присоединяется к H 2 O 2 с образованием Cys- SOH (). Локальное разворачивание позволяет захватывать Cys-SOH разделяющимся Cys (C r ) другого мономера, что приводит к образованию связанного дисульфидной связью гомодимера Prx. Однако с каждым оборотом небольшая фракция Prx в состоянии Cys-SOH не разворачивается и подвергается дальнейшей реакции с H 2 O 2 с образованием неактивных Cys-SO 2 H (гиперокисленных) частиц.До этого исследования было хорошо задокументировано, что человеческий Prx2 (цитоплазматический) был более восприимчив к гиперокислению, чем Prx3 (митохондриальный), хотя молекулярные механизмы, лежащие в основе дифференциальной устойчивости, были неизвестны [111]. Чтобы исследовать структурные факторы, придающие внутреннюю устойчивость Prx3 к инактивации, образование и реактивность белков Cys-SOH наблюдали в состоянии нативного белка с помощью TR-MS. Ранее репарацию Cys-SO 2 H в Prx2 с помощью Srx контролировали в короткие моменты времени с помощью быстрого химического гашения с последующей масс-спектрометрией [110].Хотя быстрое химическое гашение обеспечивает низкие моменты времени в миллисекундах, оно требует сильно кислых или других жестких методов гашения, которые часто мешают обнаружению нестабильных промежуточных продуктов. В экспериментах TR-MS в непрерывном потоке эквимолярные растворы Prx и H 2 O 2 (100 мкМ), оба в летучих буферах на основе аммония (бикарбонат или ацетат), загружали в отдельные шприцы, а затем смешивали при различных условиях. скорости потока. Мониторинг реакции с помощью TR-MS в низком втором временном диапазоне позволил провести первое прямое наблюдение за Cys-SOH для Prx2 (как второстепенный вид), который потреблялся ~ 6 с.В этих условиях соответствующий Cys-SOH Prx3 не наблюдался, хотя окисленный димер присутствовал, что указывает на сильную склонность к образованию дисульфидов и очень короткое время жизни сульфеновой кислоты. Дополнительные аспекты этого исследования выявили ключевые остатки около C-концевой области, которые влияют на предрасположенность к гиперокислению, однако только с TR-MS был возможен подробный сравнительный анализ химии Cys-SOH в двух ферментах Prx.
Также в это время сообщалось об отдельном исследовании кинетики окисления Prx2 и Prx3 человека с использованием как ЖХ / МС интактного белка, так и гель-электрофореза.Каждый фермент Prx инкубировали с различными количествами субстрата H 2 O 2 , затем гасили добавлением каталазы, после чего восстановленные Prx Cys-SH блокировались NEM [112]. Поскольку гомодимер Prx SS ионизируется менее эффективно, чем мономерные частицы, полученные растворы были восстановлены с помощью DTT для восстановления димерных частиц (как SS, так и SS + SO 2 H, где один C p является дисульфидным партнером, а другой C p — сульфиновая кислота) к мономерам Prx-SH и Prx-SO 2 H.Масс-спектры, полученные для восстановленных образцов, сравнивали с масс-спектрами невосстановленных образцов для определения общих соотношений этих веществ для данной концентрации H 2 O 2 . Эти и другие исследования позволили определить константы скорости гиперокисления Prx и образования димера SS как для Prx2, так и для Prx3. Константы скорости реакций пероксидного тиола (k = 2 × 10 7 M −1 s −1 ) и образующейся сульфеновой кислоты (k = 1.2 × 10 4 M −1 с −1 ) с H 2 O 2 относительно равны для Prx2 и Prx3; резкое различие заключается в относительных константах скорости образования гомодимера, 2 с -1 для Prx2 и 20 с -1 для Prx3, что убедительно свидетельствует о том, что более высокая скорость димеризации снижает вероятность гиперокисления и инактивации Prx3 [112 ].
Таким образом, несмотря на разные экспериментальные подходы и подходы МС, использованные в этих исследованиях, результаты были согласованными, демонстрируя большую универсальность и преимущества использования МС при исследовании кинетики ферментов в целом и кинетики на основе тиолов в частности.В контексте биохимии Prx подходы MS также использовались для сравнения чувствительности ферментов Prx к инактивации пероксидом [113], для идентификации Cys PTM в мышином Prx6 [114] и для характеристики нитрования Tyr193 как активатора активности Prx2. и протектор от гиперокисления [115].
MS-анализы для выявления конформационных изменений белка, вызванных окислением
Классические подходы структурной биологии к изучению конформационных изменений белков, возникающих в результате окисления или других химических модификаций, включают использование рентгеновской кристаллографии и спектроскопии ядерного магнитного резонанса (ЯМР).Хотя эти подходы обеспечивают однозначные структурные детали и широко используются сегодня, применение МС в качестве инструмента для исследования структуры белка привлекло все большее внимание из-за его относительной простоты использования и требований к очень небольшому количеству образцов. Исследования структуры белка с использованием МС используют разнообразный набор экспериментальных подходов, хотя в этом разделе наше обсуждение будет ограничено МС ионной подвижности (МСМ-МС), водородно-дейтериевым обменом (HDX) и химическим сшиванием, все в контексте тиоловых модификаций и структурных изменений, вызванных этими модификациями в белках.
МС для подвижности ионов (IMS-MS), как обсуждалось ранее, обеспечивает удобное средство разделения ионов белка, генерируемых ESI, на основе их площади поперечного сечения столкновения (CCS). Изменения в третичной структуре белков, например, возникающие в результате образования внутримолекулярных дисульфидных связей после обработки окислителем, обнаруживаются в результате их измененного времени дрейфа в трубке для определения подвижности ионов относительно необработанных частиц (). Разделение IM до получения МС обеспечивает значения CCS для ионов, которые можно сравнивать с базами данных характеристических значений CCS для прогнозирования изменений в белковой матрице, и, что не менее важно, обеспечивает средства аналитического разделения более сложных образцов, обеспечивая более тщательное измерение m / z для соответствующих видов.Используя этот подход, остатки цистеина, участвующие в дисульфидных связях, измеряли с помощью IMS-MS в моноклональных антителах IgG2, которые, как известно, образуют белковые комплексы более высокого порядка [116]. В этом исследовании использование IMS-MS вместе с целевыми мутациями Cys в Ser пролило свет на конкретные паттерны дисульфидных связей, лежащих в основе молекулярного поведения IgG2, и предоставило средства для изучения других антител, в том числе тех, которые используются в качестве лекарственной терапии [116]. Помимо интактных белков, IMS-MS успешно использовался для картирования дисульфидных связей в пептидах.Существенной проблемой при изучении внутрипептидных дисульфидов является склонность к дисульфидному скремблированию, особенно среди пептидов с множественными внутримолекулярными дисульфидными связями. Альтернативный подход основан на расщеплении дисульфидов пептидов и окислении серы дисульфидов до цистеинсульфоновой кислоты (Cys-SO 3 H) с использованием m -хлорпербензойной кислоты (mCPBA) [117]. Это нововведение предотвратило перетасовку дисульфидов на модельных пептидах и было легко связано с MS 2 секвенирования (CID) пептидов и модифицированных цистеиновых сайтов.
Обзор стратегий исследования конформационных изменений белков с помощью масс-спектрометрии. A : Общая схема метода водородно-дейтериевого обмена (HDX), используемого для выявления гибких и / или подверженных воздействию растворителя областей третичной и четвертичной структуры белка. B : Химическое сшивание как ценное средство для исследования конформации белка в нативном и нарушенном состояниях. В этом примере близлежащие остатки цистеина поперечно сшиты гомобифункциональным малеимидом (Mal-Ph-Mal), образуя ковалентную поперечную связь, устойчивую к протеолизу, ионизации и фрагментации MS.События как внутримолекулярного, так и межмолекулярного сшивания могут происходить в зависимости от уникальной структуры исследуемого белка или белкового комплекса. C : IMS-MS позволяет разделить конформеры белка на основе третичной структуры белка.
Исследования третичной и четвертичной структуры белков также возможны с помощью ESI или MALDI MS. Поскольку структура белка более высокого порядка является функцией многих параметров, включая ковалентные и нековалентные взаимодействия, как меж-, так и внутримолекулярные, для определения структуры часто требуются более мягкие образцы и условия ионизации, которые не нарушают структурные атрибуты до сбора данных.Белковые комплексы особенно хорошо подходят для ESI и были проанализированы в ряде исследований [см. [118] для обзора по этой теме]. Кроме того, MS-анализ интактных белков оказался полезным для определения стехиометрии связывания белков и их кофакторов или субстратов, хотя точные молекулярные местоположения, где происходят взаимодействия, не могут быть легко определены с использованием таких методов [119].
Чтобы понять динамические изменения в структуре белка, которые не приводят к разнице масс (m / z), к МС был применен метод водородно-дейтериевого обмена (HDX), известный для структурных исследований на основе двумерного ЯМР.В экспериментах с HDX белки в их естественном состоянии погружаются в раствор D 2 O на контролируемое время, в течение которого обмениваемые (относительно кислые) протоны в доступных для растворителя областях будут заменены дейтронами, вызывая изменение массы (). В зависимости от индивидуального исследования, эксперименты HDX могут выполняться с использованием непрерывного мечения, при котором аликвоты белка удаляются, гасятся и анализируются с помощью МС в различные моменты времени после добавления D 2 O, или с использованием импульсного мечения.При импульсной маркировке белок инкубируют с D 2 O в течение очень короткого периода времени (10 с) после того, как индуцируют нарушение в структуре белка (например, связывание лиганда) [120]. Сайты, испытывающие наибольшую степень обмена, включают протоны на амидных атомах азота и на гетероатомах боковых цепей аминокислот (-OH, -NH 2/3 , -SH и т. Д.), Особенно в тех областях белка, которые находятся на поверхности наряду с теми, кто находится в неструктурированных или гибких регионах. После включения достаточного количества дейтерия ( 2 H) обмен гасят быстрым снижением pH (2–3) и температуры (0 ° C), денатурируя белок и резко замедляя обмен.Такое введение кислоты позволяет большинству дейтронов гетероатомов боковой цепи обмениваться обратно на протоны, оставляя амидные дейтроны, которые увеличивают массу на 1 Да за замену [119]. Эти динамические изменения в структуре, обозначенные HDX, обнаруживаются при анализе интактного белка или после переваривания кислотоустойчивой протеазой, такой как пепсин.
В контексте ферментов активного центра Cys, HDX-MS был использован для исследования структурной динамики пероксиредоксина AhpC из Salmonella typhimurium , поскольку склонность к окислительной инактивации всех ферментов Prx сильно зависит от структурной среды, окружающей активный сайт Cys [121].Кроме того, HDX предоставил средства для анализа конформационных аспектов моноклональных антител до и после конъюгации лекарств, выявляя наличие межцепочечных дисульфидных связей, которые помогают стабилизировать конъюгаты антитело-лекарство [122]. Эффекты S -нитрозилирования человеческой глутатионтрансферазы P1-1 по двум сайтам Cys были проанализированы с использованием HDX, который показал, что модификация Cys101 конформационно стабилизирует, но двойное нитрозилирование Cys47 и Cys101 приводит к нарушению структурной целостности в активный центр и соответствующая потеря ферментативной активности [123].Широкая применимость HDX для изучения многих типов белков на основе тиолов, включая те, которые участвуют в механизме образования кластеров Fe-S [124], белков-супрессоров опухолей [125], а также исследования биотерапевтических средств, таких как IgG [126], иллюстрируют неоспоримое значение этого метода в области структурной протеомики на основе МС.
Химическая маркировка — это метод, применяемый для исследования остатков, близких в трехмерном пространстве, включая те, которые участвуют в нековалентных взаимодействиях, с помощью МС.Экспериментально белки, в том числе в комплексах, реагируют с реагентом, который ковалентно связывает остатки, а затем перевариваются с использованием выбранной протеазы. MS-анализ связанных пептидов предоставляет информацию о последовательностях двух связанных пептидов вместе с присоединенными остатками (). Длина линкерных реагентов адаптирована для получения информации об остатках на определенных расстояниях в свернутом белке. Обычно сшивающие реагенты являются гомобифункциональными и включают малеимиды для нацеливания на остатки Cys [127] или сложные эфиры N -гидроксисукцинимида (NHS) для нацеливания на остатки Lys.Остатки лизина чаще становятся мишенью для исследователей, использующих подходы перекрестного связывания из-за их более высокой частоты в протеоме по сравнению с цистеином (6% для Lys; <2% для Cys) [128]. Библиотека доступных перекрестно-сшивающих реагентов в последнее время становится все более сложной и универсальной, включая реагенты, которые снабжены биотином для обогащения перекрестно-сшитых пептидов [129, 130], а также реагенты, которые имеют расщепляемые сайты, чтобы либо уменьшить сложность образца до MS. анализа или расщепления во время фрагментации CID [131, 132].MS-анализ поперечно-сшитых пептидов усложняется их повышенным m / z, менее предсказуемым паттерном фрагментации и потенциальной фрагментацией самого линкера, что требует уровней оптимизации MS наряду с усовершенствованием самих химических линкеров. В недавнем отчете подробно описывается разработка, в которой CID используется параллельно с диссоциацией с захватом электронов (ECD): CID расщепляет линкер в предсказуемом сайте для генерации репортерных ионов, в то время как ECD не вызывает расщепление линкера и вместо этого предоставляет информацию о пептидной последовательности [131 ].Кроме того, были разработаны сшивающие реагенты, нацеленные на дисульфиды, которые расщепляют только в режиме отрицательных ионов, что позволяет проводить дополнительные анализы на одном образце [133].
Помимо дисульфидного картирования, исследования перекрестного связывания основывались на присущей цистеину нуклеофильности с образованием аддуктов с гетеробифункциональными линкерами. Канцероген фуран, компонент дыма, активируется in vivo цитохромами P450 до цис -2-бутен-1,4-диал (BDA), который легко связывает Lys и окислительно-восстановительный Cys в белках, обеспечивая уникальная сигнатура для белков или тканей, подвергшихся воздействию фурана [134].Точно так же образование конъюгатов Cys-guanine-N7 (белок-ДНК) в ответ на воздействие 1,3-бутадиена, компонента выхлопных газов транспортных средств и сигаретного дыма, было идентифицировано с помощью MS в более чем 150 белках в клетках фибросаркомы человека [135 ]. Наконец, HDX и поперечное сшивание были недавно использованы в тандеме для исследования третичной структуры нецистеинсодержащего лизостафина из Staphylococcus simulans [136]. Во-первых, HDX-MS был использован для идентификации предполагаемых границ раздела доменов, установленных нековалентными взаимодействиями, затем остатки цистеина были встроены в структуру в положениях, которые, как предполагается, должны быть достаточно близкими, чтобы способствовать образованию дисульфидной связи [136].Успешное образование дисульфидных связей, по оценке MS, предоставило ключевые доказательства близости двух областей в 3-мерном пространстве, иллюстрируя, что исследования структуры белка с помощью MS достижимы без мечения аминокислот с использованием реакционноспособных соединений.
FitOn расширяет свою фитнес-платформу за счет финансирования Delta-v Capital в размере 18 миллионов долларов — TechCrunch
Выделить на тренировку 45 минут или час может быть сложно на работе, дома и в повседневной жизни. Компания FitOn, занимающаяся цифровым фитнесом и оздоровлением, была создана для 15 или 20-минутных тренировок, направленных на то, чтобы люди двигались в любое удобное для них время.
Компания завершила раунд финансирования серии B в размере 18 миллионов долларов, возглавляемый Delta-v Capital, с участием существующих инвесторов Accel, Telstra Ventures, Crosscut Ventures, Maverick Ventures и Second Avenue Partners.
Финансирование поступает, поскольку компания из Лос-Анджелеса набрала 10 миллионов участников для своего приложения, которое предлагает персонализированные фитнес- и оздоровительные программы и уникальный социальный опыт от таких тренеров, как Orangetheory Fitness, KINRGY и Zumba, и даже некоторых послов знаменитостей, таких как Габриэль Юнион, Джулианна. Хаф, Холли Берри, Джонатан Ван Несс и The Chainsmokers.
Основанное мужем и женой бывшего руководителя Fitbit Линдси Кука и основателя AllTrails Рассела Кука, приложение было запущено два года назад с миссией, которую генеральный директор Линдси Кук назвала «демократизацией цифрового фитнеса».
«Я была полностью погружена в эту категорию и работала в фитнес-компании, но обнаружила, что у меня самой не так много времени на фитнес», — сказала она. «Я был не в восторге от некоторых приложений, которые не были продуктами премиум-класса и имели множество препятствий для входа, например, дорогие велосипеды или оборудование.Я увидел изменение в демократизации фитнеса, когда увидел, что массовый рынок никому не нужен ».
Она объединилась со своим мужем, который проработал 20 лет в цифровых продуктах, чтобы создать приложение для здоровья и фитнеса, которое будет делать то, что AllTrails делает для активного отдыха, и то, что Calm делает для медитации, добавил Кук.
Приложение FitOn
FitOn — это продукт freemium, и его участники имеют доступ к сотням тренировок в области кардио, силовых тренировок, йоги, растяжки и медитации. У него также есть профессиональная модель подписки, которая включает в себя музыкальные треки поверх тренировок, подключение к устройствам для мониторинга здоровья, офлайн-загрузки, планы питания и рецепты.Он также имеет встроенную социальную платформу с функцией чата, и участники могут приглашать друзей для участия в программах и задачах.
Последний раунд финансирования дает FitOn в общей сложности 30 миллионов долларов, привлеченных на сегодняшний день, включая посевной раунд в 2018 году и серию A в 2020 году.
«FitOn предоставляет своим участникам миллионы тренировок в месяц, уделяя приоритетное внимание инновациям в продуктах, сообществу и премиальному контенту, чтобы создать одно из крупнейших сообществ здоровья и фитнеса», — сказал Дэвид Шаллер, управляющий партнер Delta-v Capital.«Мы очень рады, что FitOn присоединился к нашему портфелю новаторов в области цифрового фитнеса и велнеса, в который входят и другие лидеры в этой категории, такие как Tonal».
Начиная с серии A, рост компании ускорился и вырос на 500% в годовом исчислении за счет выручки от модели подписки. Теперь, когда количество участников превысило 10 миллионов, Кук рассчитывает достичь 25 миллионов к концу следующего года. Кроме того, участники FitOn отработали более 1 миллиарда минут тренировок, и большинство из них тренируются три или четыре раза в неделю.
Приложение получает на 285% больше загрузок, чем Peloton, на 588% больше, чем Beachbody, и постоянно входит в двадцатку лучших в категории здоровья и фитнеса и, по словам Кука, получило более 300 000 пятизвездочных отзывов.
Новое финансирование позволит компании расширить свою команду, укрепить ключевые позиции и продолжить быстрый темп выпуска новых продуктов. За последние два года компания выпустила более 78 продуктов. FitOn в основном ориентировался на Соединенные Штаты, но планирует также расти и на международном уровне.
«Для бизнеса, который создается два года назад, мы довольны доходом и имеем много возможностей для создания крупномасштабного бизнеса», — сказала Линдси Кук. «В этой культуре, направленной против диет, речь идет не о« идеальном теле », а о поощрении ухода за собой. Мы создаем бренд стиля жизни, поощряющий людей делать 10-минутные тренировки здесь и там ».
Мощность, получаемая жидкостью от насоса или вентилятора
Мощность, получаемая жидкостью
Мощность, получаемая жидкостью от насоса или вентилятора, может быть выражена как:
P = mw (1)
, где
P = мощность (Вт)
м = массовый расход (кг / с)
w = удельная работа (Нм / кг, ) Дж / кг )
Удельная работа
Удельная работа — w — может быть выражена:
w = gh (2)
где
h = напор (м)
г = ускорение свободного падения (9.81 м / с 2 )
Массовый расход
Массовый расход — м — можно выразить:
м = ρ Q (3)
где
ρ = плотность (кг / м 3 )
Q = объемный расход (м 3 / с)
Объединение (1), (2) и (3) мощность, получаемая жидкостью от насоса или вентилятора, может быть выражена как:
P = ρ Q gh (4)
С удельным весом, выраженным как:
γ = ρ г (5)
где
γ = удельный вес (Н / м 3900 36)
уравнение (4) можно изменить так, чтобы мощность, получаемая жидкостью от насоса или вентилятора, могла быть выражена как:
P = γ Q h (6)
Поскольку напор можно выразить как
h = (p 2 — p 1 ) / γ (7)
, уравнение (4) может быть изменено таким образом, чтобы мощность, полученная с помощью жидкость из насоса или вентилятора может быть выражена как:
P = Q (p 2 — p 1 ) (8)
Пример — подъем напора встроенного насоса
An Встроенный водяной насос работает при измеренном давлении 1 бар (1 10 5 Н / м 2 ) и 10 бар (10 10 5 Н / м 2 ) .Плотность воды 1000 кг / м 3 . Объемный расход измеряется как 1 10 -3 м 3 / с .
Фактический напор (водяной столб) можно рассчитать по формуле (7):
h = (p 2 — p 1 ) / γ
= (p 2 — p 1 ) / ρ g
= ((10 10 5 Н / м 2 ) — (1 10 5 Н / м 2 )) / (1000 кг / м 3 ) (9.81 м / с 2 )
= 91,7 м — водяной столб
Мощность, приобретаемая жидкостью, может быть рассчитана с помощью уравнения (4):
P = ρ Q gh
= (1000 кг / м 3 ) (1 10 -3 м 3 / с) (9,81 м / с 2 ) (91,7 м)
= 899,6 кг.м 2 / с 3 (Вт)
= 0,9 кВт
Губ.Ньюсом расширяет прокламацию о засухе
Восстановить Дельту 11 мая 2021 г.
— Тим Строшейн и Барбара Барриган-Паррилья
10 мая 2021 года губернатор Ньюсом распространил свое объявление о чрезвычайной засухе еще на 39 округов, а теперь всего 41 округ. В его последнем заявлении вырваны страницы из сборника правил, который губернатор Браун использовал в 2013 и 2014 годах.
Департамент водных ресурсов Калифорнии установит по крайней мере один, а может и больше, засоленных барьеров в Дельте.Это нарушит водные пути, создаст застойные водоемы, а с установленной концентрацией водорослей приведет к еще большему и, вероятно, более значительному вредоносному цветению водорослей (ВЦВ) в течение лета и осени. ВЦВ представляют опасность для здоровья населения из-за контакта с водой людей и собак, а также из-за выбросов загрязняющих веществ, переносимых по воздуху.
Временные рамки для барьеров солености в Дельте пока отсутствуют. Объявление Ньюсома о чрезвычайной ситуации приостанавливает действие правил экологической экспертизы, поэтому барьеры, очевидно, будут возведены без тщательной экологической экспертизы или общественного контроля.Его прокламация приостанавливает публикацию объявлений в газетах, но требует, чтобы государственные водные агентства размещали информацию о своих действиях, связанных с засухой, на своих веб-сайтах.
DWR находится на water.ca.gov; Государственный водный совет находится на сайте waterboards.ca.gov.
Прокламация Ньюсома также направлена на сохранение существующих бассейнов с холодной водой в водохранилищах, расположенных выше по течению, особенно на озерах Шаста и Оровилль. Скорее всего, это слишком поздно. К сожалению, в этих резервуарах уже очень мало воды, и их бассейны с холодной водой рассосались прошлой зимой, когда запасы были отправлены в южную Калифорнию и водохранилище Сан-Луис к югу от Дельты.
Племена индейцев Северной Калифорнии глубоко обеспокоены исчезновением лосося в этом году, поскольку такое исчезновение означало бы культурный геноцид — лосось является важным символом в их духовной жизни. Племена и их союзники провели важный День действий в прошлый вторник, 4 мая в Сакраменто, чтобы потребовать действий и защиты от администрации Ньюсома. Вымирание лосося также подорвет усилия Ньюсома по восстановлению отношений штата Калифорния с племенами калифорнийских индейцев.
Губернатор также дал указания водным ведомствам штата, включая Государственный совет по контролю за водными ресурсами и Департамент рыб и дикой природы, для облегчения передачи и обмена водоснабжения, принятия чрезвычайных положений для содействия сохранению воды, плана по ограничению прав несовершеннолетних на воду по всему штату. для защиты источников воды и оценки стандартов речного стока для защиты рыб и экосистем.
В целом, последнее заявление Ньюсома сигнализирует о том, что Бюро мелиорации США и Департамент водных ресурсов будут запрашивать временные «петиции о срочном изменении», чтобы отменить целевые показатели качества воды в Дельте.Это было сделано в сочетании с добавлением засоленного барьера False River в 2015 году, чтобы ограничить циркуляцию приливной воды в центральной дельте, что защищает свежесть воды, поступающей из реки Сакраменто на насосные станции южной части дельты.
Множество видов рыб находятся на грани исчезновения или находятся под большой угрозой исчезновения в Дельте, и решение губернатора Ньюсома состоит в том, чтобы позволить этим видам, таким как чавычи и корюшка Дельты, стать частью шестого великого массового вымирания.
Если бы губернатор Ньюсом заботился о Дельте, он бы предпринял шаги в прошлом году, когда мы знали, что мы находимся в условиях засухи, для защиты этих видов и для поддержки действий штата по сокращению вредоносного цветения водорослей, которое становится все более распространенным и ухудшается. в Дельте. Его план расходов на воду не предлагает ничего для значимого снижения вредного цветения водорослей, изучения или отслеживания — просто больше мертвых водоемов, где эти цветы могут расти.
Его заявление включает, однако, помощь промышленному сельскому хозяйству с финансированием ремонта канала Фриан-Керн, который затонул в результате чрезмерного перекачивания грунтовых вод этими производителями.