Делаем крылья. Тренировка широчайших мышц спины. | Фитнес и бодибилдинг
Отличительный признак по-настоящему продвинутого атлета будет не размер бицепса или рельефный пресс. Главной мышечной группой атлета является спина. Очень широкая, мощная, плотная и бугристая спина вызывает завидный взгляд окружающих, а также любопытный интерес у женщин. V – образная форма тела появляется наряду с хорошим развитием широчайших мышц спины и мышц плечевого пояса.
Необходимо работать над техникой движения, выполнять упражнения со свободными весами (штанги, гантели), стараться чувствовать свои мышцы.
Что даёт мощная спина?!
- Мускулатура спины испытывает изометрическое напряжение во всех базовых упражнениях, этот крепкий «каркас» туловища даёт возможность повысить рабочие веса во всех упражнениях. Во время приседаний, наша спина особенно её нижний отдел, довольно сильно нагружаются. Становясь сильнее в тренировке спины, мы также становимся сильнее в тренировке ног.

- Улучшается физическое самочувствие и восприятие собственной силы. Это вызвано отсутствием ощущения слабости в спине, которая постоянно находится в тонусе в силу стабильных тренировок. Риск потянуть спину или получить межпозвоночную грыжу, резко уменьшается.

Тренировка спины на турнике
Турник является очень популярным спортивным атрибутом во всём мире. Подтягивания на турнике развивают не только спину, но также грудь, плечи и руки.
Для многих это упражнение является крайне сложным и без спортивной подготовки, подтянуться 8–12 раз не по силам. Большинство из нас, подтягиваются от 2 до 6 повторений. Для начала, нужно повысить сам результат в подтягиваниях как минимум до 10 повторений за один подход. А вместе с этим, начнут приходить мышечные ощущения.
Как увеличить число подтягиваний на турнике?!
Способ основан на достижении заветной цифры в 50 подтягиваний за один сеанс тренировок. То есть, количество повторений в каждом подходе складывается до тех пор, пока не выйдет 50. При этом, добиваясь именно увеличения результата в этом упражнении, выполнять другие упражнения для мышц спины, кроме становой тяги нельзя. Выполнять её следует второй по счёту, или на другой день. Каждый подход доводится до полного мышечного отказа.
То есть, количество повторений в каждом подходе складывается до тех пор, пока не выйдет 50. При этом, добиваясь именно увеличения результата в этом упражнении, выполнять другие упражнения для мышц спины, кроме становой тяги нельзя. Выполнять её следует второй по счёту, или на другой день. Каждый подход доводится до полного мышечного отказа.
Несколько советов по достижению мышечных ощущений при тренировке спины:
- Изучите анатомию мускулатуры человека, где находятся крепления мышц к костной структуре. Во время тренировок, мысленно сближайте эти точки друг к другу и концентрируйтесь на рабочих мышцах.
- Во время выполнения любых тяг, будь они вертикальными, горизонтальными или же в тренажёре, тяните локтями. То есть, представьте, что вес закреплён не к кистям, а к локтям. Это поможет разгрузить бицепсы и направит нагрузку на целевые мышцы спины.
- Работать нужно лопатками и плечевым поясом. В тот момент, когда мышцы полностью сокращены и движение находится в конечной точке, лопатки должны быть сближены, а плечи повёрнуты назад. И соответственно, в начальной точке, когда мышцы растянуты, лопатки должны быть разведены, а плечи повёрнуты вперёд или вверх.
Увеличив показатели в подтягиваниях, можно приступать к дальнейшим упражнениям, включая их в свою обычную программу тренировок.
Упражнения.
Ниже приведены упражнения, которые нагружают не только широчайшие мышцы спины, но и все остальные. Достичь наилучшего и более быстрого прогресса поможет их совмещение. Мышцам спины сложно, а то и невозможно развиваться по отдельности. Комплексная тренировка спины в целом станет идеальным вариантом. Но делать акцент на турнике и тягах в наклоне всё же стоит.
Тяга штанги в наклоне
При правильной технике и с учётом хорошей нейромышечной связи, это упражнение превращается в невероятно мощный инструмент для развития верхней части спины.
Воздействие
Активное развитие широчайших мышц спины. Не в меньшей степени нагружены трапециевидные мышцы. Статическое напряжение испытывают мышцы разгибатели позвоночника. А также, нагружаются бицепсы.
Не в меньшей степени нагружены трапециевидные мышцы. Статическое напряжение испытывают мышцы разгибатели позвоночника. А также, нагружаются бицепсы.
Выполнение
Это упражнение можно выполнять двумя разными хватами: прямым и обратным. Прямой хват, или как его иногда называют «верхний», подразумевает расположение кистей, тыльной стороной ладони от себя. И соответственно, при обратном (нижнем) хвате, ваши тыльные стороны ладоней обращены к вам и вниз. Принцип действия упражнения при смене хвата меняется, но не существенно. При обратном хвате, за счёт возросшей амплитуды движения, нагрузка эффективно затрагивает нижний отдел широчайших мышц. Пробуйте оба варианта и выберите более подходящий для вас. Можете просто чередовать хват от одной тренировки к другой.
Можете держать штангу, как на полу, так и на удобных стойках. Возьмитесь за гриф штанги на ширине плеч. Наклонитесь вперёд, немного согнув ноги в коленях так, чтобы штанга повисла на прямых руках. Спина прямая, как и при становой тяге, но при этом, необходимо создать небольшой угол в спине, относительно пола. То есть, просто приподнять плечи выше уровня таза. Опускаться до уровня параллели спины с полом можно, но если нейромышечная связь при таком варианте хуже, то лучше выполнять упражнение слегка приподняв верх спины, как написано выше.
Спина прямая, как и при становой тяге, но при этом, необходимо создать небольшой угол в спине, относительно пола. То есть, просто приподнять плечи выше уровня таза. Опускаться до уровня параллели спины с полом можно, но если нейромышечная связь при таком варианте хуже, то лучше выполнять упражнение слегка приподняв верх спины, как написано выше.
Ведите гриф штанги как можно ближе к ногам, почти касаясь коленей. Так вы снимете неприятную изометрическую нагрузку с поясницы. Тянуть необходимо книзу живота, пока гриф его не коснется. Для качественной связи мозга с мышцами, когда вы только обучаетесь этому упражнению, работая с малыми весами, задерживайте штангу в верхней точке движения и дополнительно напрягайте мускулатуру.
Становая тяга
В плане рабочих весов, это самое мощное упражнение. Освоив и доведя технику упражнения до идеала, рабочие веса тут же начнут расти. Хороший результат в становой тяге позволяет раскрывать потенциал и в других упражнениях, особенно в приседаниях. Это упражнение придаст спине невероятную мощь, как в силовом понимании, так и в визуальном.
Это упражнение придаст спине невероятную мощь, как в силовом понимании, так и в визуальном.
Воздействие
В этом упражнении активно работают разгибатели позвоночника, так называемые мышечные «столбы», расположенные вдоль позвоночника. Также, в верхней точке усиленно подключается трапециевидная мышца. В меньшей степени работают широчайшие мышцы спины. Так же стоит упомянуть о немалой работе ягодичных мышц и мышц задней поверхности бедра. Становая тяга развивает толщину и плотность спины.
Выполнение
Вначале и в конце упражнения, штанга должна находиться на полу, ставить её на стойки не нужно. Желательно отрывать штангу именно от пола. Наклонитесь и возьмите гриф на ширине плеч. Поднимите голову и смотрите прямо вперёд перед собой. Выпрямите спину от шеи до копчика, словно вашу спину плотно прижали к ровной доске. Такое положение спины необходимо держать и контролировать постоянно. В начальной точке, когда вы готовитесь оторвать вес от пола, не стесняйтесь «оттопыривать» попу назад, так вы займёте правильное исходное положение.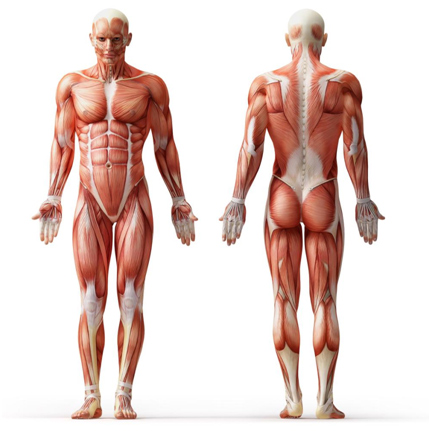 А теперь, одновременно работая в коленных и тазобедренных суставах, то есть, выпрямляя их, начните поднимать штангу. Если во время подъёма штанга сильно задевает колени, это значит, что вы выпрямляете туловище, быстрее, чем ноги, или недостаточно наклоняетесь вперёд. В идеале, во время этого упражнения, вы подаётесь вперёд, а штанга будет немного касаться ваших коленей во время подъёма. В конечной точке, важно сократить мышцы спины, подав плечи немного назад, прижав, друг к другу лопатки. Но не путайте это с прогибом поясничного отдела, вам просто необходимо сомкнуть друг к другу лопатки. Так вы значительно усилите воздействие на трапециевидные мышцы.
А теперь, одновременно работая в коленных и тазобедренных суставах, то есть, выпрямляя их, начните поднимать штангу. Если во время подъёма штанга сильно задевает колени, это значит, что вы выпрямляете туловище, быстрее, чем ноги, или недостаточно наклоняетесь вперёд. В идеале, во время этого упражнения, вы подаётесь вперёд, а штанга будет немного касаться ваших коленей во время подъёма. В конечной точке, важно сократить мышцы спины, подав плечи немного назад, прижав, друг к другу лопатки. Но не путайте это с прогибом поясничного отдела, вам просто необходимо сомкнуть друг к другу лопатки. Так вы значительно усилите воздействие на трапециевидные мышцы.
Поначалу, нужно отрабатывать технику движения с малым весом (около 50кг). Отработав технику, можно добавлять веса.
Гиперэкстензия
Гиперэкстензия для прямых мышц спины является «младшим братом» становой тяги. Поскольку принцип воздействия на мускулатуру почти идентичен, разница лишь в том, что в гиперэкстензиях отсутствует необходимость применять огромные веса. Но это не значит, что наращивать рабочие веса в этом упражнении не нужно, просто они малы, в основном это 20–40 килограмм для атлета среднего уровня. Можно применять маленький гриф, закинув его за шею, так и блин, обняв и прижав его к груди. Упражнение целенаправленно развивает силу и мощь всего мышечного «каркаса» спины. Особенно популярно это упражнение среди травмировавшихся атлетов так, как позволяет довольно быстро «встать на ноги». А весь секрет в том, что упражнение прорабатывает мелкие мышцы вокруг всего позвоночного столба, что сложно сделать в других упражнениях.
Но это не значит, что наращивать рабочие веса в этом упражнении не нужно, просто они малы, в основном это 20–40 килограмм для атлета среднего уровня. Можно применять маленький гриф, закинув его за шею, так и блин, обняв и прижав его к груди. Упражнение целенаправленно развивает силу и мощь всего мышечного «каркаса» спины. Особенно популярно это упражнение среди травмировавшихся атлетов так, как позволяет довольно быстро «встать на ноги». А весь секрет в том, что упражнение прорабатывает мелкие мышцы вокруг всего позвоночного столба, что сложно сделать в других упражнениях.
Воздействие
Исключительная работа разгибателями спины. Именно гиперэкстензиями можно как следует проработать все мышцы, которые держат позвоночник. Широчайшие мышцы практически не задействованы, а также минимизирована работа трапециевидных мышц. Также в упражнении задействованы ягодичные мышцы и мышцы задней поверхности бедра.
Выполнение
Существует множество стоек для этого упражнения, и бывают они под разными углами по отношению конечной точки движения к полу. То есть, когда вы находитесь параллельно или под небольшим углом. Считается, что выполняя гиперэкстензии в параллельной стойке, нагрузка в верхней точке движения чуть выше, чем в упражнении в стойке под углом. Но кардинальной разницы в этом нет, тем более, если в вашем зале только один вид стойки для гиперэкстензий.
Если вы новичок, то дополнительное отягощение не нужно, достаточно веса собственного тела. Подрегулируйте вылет опоры под тазовую область так, чтобы верхний край опоры был на уровне вашего пояса. Есть один нюанс, некоторые стойки для гиперэкстензий очень неудобны и доставляют дискомфорт, а иногда и боль. В таком случае, подкладывайте что-нибудь мягкое под бёдра.
Тренажёры для тренировки мышц спины
В крупных элитных залах количество тренажёров огромное. Это очень хорошо, постоянное разнообразие тренинга хорошо влияет на прогресс в физическом развитии. Но, работая со свободными весами (штанги, гантели), создаётся более мощный импульс к развитию, нежели работая с сопротивлением в тренажёрах.
Как часто тренировать спину?
Вначале пути, начинать нужно с подтягиваний и параллельно оттачивать технику выполнения становой тяги с весом не более 50-ти кг. Далее, можно увеличить арсенал упражнений тягой штанги в наклоне. И так далее, пробовать можно все упражнения для спины. Позже можно выделить самые эффективные из них. Для восстановления 1−2 дней достаточно. Ничего сложного, боли прошли, можно тренироваться.
Что же касается других мышечных групп, то можно совмещать тренировку спины с грудью или с руками. Если тренировать руки и спину в один день, то начинать нужно со спины. Так и с другими группами мышц. Тренировки всегда должны начинаться с крупных мышечных групп.
Вывод:
Необходимо изучать упражнения, начинать с подтягиваний и становой тяги. Повышать силовые показатели в базовых упражнениях. Нарастить мышцы без грамотного питания, также как и построить дом без кирпичей – невозможно. Нужно употреблять достаточное количество белка для роста мышц, качественные углеводы для энергии и полезные жиры для развития организма в целом.
Нужно употреблять достаточное количество белка для роста мышц, качественные углеводы для энергии и полезные жиры для развития организма в целом.
На крыльях ночи коленками назад
Нежити и упыри, «полумышь-полуптица», «полуптица-полумышь»… Какими только эпитетами не награждали летучих мышей за длительную историю их знакомства с человеком! А меж тем, эти удивительные животные относятся к тому же классу, что и мы с вами, к классу млекопитающих. Если присмотреться повнимательнее, можно заметить множество общих черт, указывающих на родственные отношения рукокрылых. Тело их покрыто мехом, а на груди у самок расположены соски, позволяющие вскармливать детенышей молоком. Вообще-то, сидящую летучую мышь со сложенными крыльями неспециалист легко может принять за полевку или землеройку: этакий же меховой шарик, с небольшими глазками на пушистой мордочке и торчащими вверх ушами, да еще и нередко оглашающий окружающее пространство тонким «мышиным» писком. Но уже если зверек расправит крыло – сомнений ни у кого не останется. Именно большие кожистые крылья, позволяющие этим зверькам, весом от нескольких грамм до целого килограмма, подниматься в воздух, являются уникальной визитной карточкой всех рукокрылых (многие из которых вовсе не похожи на мышей).
Впервые настоящие крылья появились у млекопитающих примерно 60 – 70 миллионов лет назад. Первые ископаемые остатки рукокрылых известны из нижнего эоцена. Однако они проливают не много света на происхождение этих удивительных животных – ведь крыло самого примитивного рукокрылого – Onychonycteris (nycteris переводится с древнегреческого как «дитя ночи», а onycho в переводе с латыни означает «коготь») – очень похоже на крыло современных рукокрылых. Свое название «когтистый потомок ночи» получил за то, что все пальцы его крыла увенчаны рудиментами когтей, что отличает его от современных рукокрылых, у которых на третьем, четвертом и пятом пальцах когти никогда не встречаются.
Именно большие кожистые крылья, позволяющие этим зверькам, весом от нескольких грамм до целого килограмма, подниматься в воздух, являются уникальной визитной карточкой всех рукокрылых (многие из которых вовсе не похожи на мышей).
Впервые настоящие крылья появились у млекопитающих примерно 60 – 70 миллионов лет назад. Первые ископаемые остатки рукокрылых известны из нижнего эоцена. Однако они проливают не много света на происхождение этих удивительных животных – ведь крыло самого примитивного рукокрылого – Onychonycteris (nycteris переводится с древнегреческого как «дитя ночи», а onycho в переводе с латыни означает «коготь») – очень похоже на крыло современных рукокрылых. Свое название «когтистый потомок ночи» получил за то, что все пальцы его крыла увенчаны рудиментами когтей, что отличает его от современных рукокрылых, у которых на третьем, четвертом и пятом пальцах когти никогда не встречаются.
Летательная перепонка рукокрылых практически лишена волосяного покрова. Она образована двумя слоями кожи, между которыми лежат кровеносные сосуды и нервы, а также тончайшие пучки мышц. Эти мышцы позволяют рукокрылым всегда держать свою перепонку натянутой – даже при сложенном крыле. Благодаря этому, она не рвется и не повреждается даже в самых тесных убежищах.
Однако в отличие от сравнительно жесткого перьевого покрова птиц, тонкой эластичной перепонке рукокрылых требуется прочный управляемый каркас, который не только поддерживает форму крыла, но и меняет его конфигурацию в зависимости от потребностей животного. Самой примечательной частью этого каркаса являются удлиненные кости кисти (пясти и пальцев). Но не нужно забывать, что в состав крыла входят не только все кости передней, но и кости задней конечности. Задние лапы рукокрылых играют очень важную роль в поддержании края перепонки и нередко участвуют и в летательных движениях. Так что в отличие от птиц, крыло рукокрылых образовано не только руками, но и ногами!
 Перепонка, натянутая между передней и задней конечностью (самая большая по площади часть крыла, производящая в полете наибольшую подъемную силу), подразделяется на плагиопатагиум (участок между телом и пятым пальцем) и хиропатагиум (перепонка между пальцами передней конечности). Перепонка между задними лапами, обычно соединенная с хвостом, называется уропатагиум. У многих видов хвост отсутствует, и тогда правая и левая части перепонки соединяются напрямую.
Перепонка, натянутая между передней и задней конечностью (самая большая по площади часть крыла, производящая в полете наибольшую подъемную силу), подразделяется на плагиопатагиум (участок между телом и пятым пальцем) и хиропатагиум (перепонка между пальцами передней конечности). Перепонка между задними лапами, обычно соединенная с хвостом, называется уропатагиум. У многих видов хвост отсутствует, и тогда правая и левая части перепонки соединяются напрямую.
Интересно, что скелетный каркас крыла рукокрылых ни содержит никаких «новых» костей. Все его составляющие характерны и для других млекопитающих, например нас с вами. Однако образующие у других млекопитающих ладонь кости пясти, которые обычно скованы кожей и связками и практически неподвижны друг относительно друга, у рукокрылых сильно удлинены и очень подвижны. Именно они, вместе с длинными фалангами пальцев составляют скелетную основу хиропатагиума. И хотя обычно каждый такой элемент принято называть пальцем, это не совсем верно, так как помимо собственно пальца в него включена и пястная кость.
Первый палец однако в перепонку не включен, он так и остался у рукокрылых коротким и используется в основном для лазания. А вот второй палец у большинства видов утрачен – сохранилась лишь пястная кость, которая вместе с тесно притянутым к ней перепонкой третьим пальцем укрепляет переднюю кромку крыла.
Задние ноги также сохраняют «стандартный» для млекопитающих набор костей. Основным их отличием является то, что коленки обращены на спину. Видимо, такое положение «навязано» задним лапам летательной перепонкой. Дело в том, что предки рукокрылых, по всей вероятности, были внешне похожи на современных летяг – имели перепонку для планирования по бокам тела. А как мы можем видеть на различных представителях планирующих млекопитающих, такая перепонка всегда развивается между обращенными друг к другу сторонами передних и задних. И, соответственно, полноценно натянуть ее можно, лишь «вывернув» заднюю ногу коленкой вбок или на спину.




Sw ч ZJ A theoretical cluster of thoughts on the skeletal anatomy of winged people and how they might work. Its purely theoretical and hopefully gives an idea of how it might work. The scapula provides a socket allow ing for movement of the arms and wings. It provides a structured anchor socket for the wing as well as being in a position where the muscles are able to support the w ings. Being connected to a scapula socket means that there is no loss in movement hut also lessens collisions w ith the arms in flight as tin* two are joined to the same structure.
 vanaUon for how far down .he back spine and When pulling the wings down U Ule hu*ferus reaches. Smaller wings will would damage the ribs. // «¡hnrtnr rn:irh hill nuor fill I hi* mavimi nn
humerus reach is to the top of the pelvic bone.
The hones are not completely iiollow hut pneumatized, meaning that they have criss crossing struts across the inside of the hone for strength. This makes makes them less heavy hut not fragile. They’re denser which makes them more durable.
They need to be this way to deal with the stresses of flying, taking off and landing as well as supporting the muscles.
Note that not all the bones are pneumatized, only the limbs are. The spine, scapula, rib cage, skull and pelvis may have a slightly thinner layer of compact hone hut its still of the same density as the limbs, in these hones is where red marrow is. which is important for the formation of red blood cells..
Rib Barbs.
These arc present to strengthen the ribcage, overlapping so that they separate too far and become damaged
Straight on Back View.
vanaUon for how far down .he back spine and When pulling the wings down U Ule hu*ferus reaches. Smaller wings will would damage the ribs. // «¡hnrtnr rn:irh hill nuor fill I hi* mavimi nn
humerus reach is to the top of the pelvic bone.
The hones are not completely iiollow hut pneumatized, meaning that they have criss crossing struts across the inside of the hone for strength. This makes makes them less heavy hut not fragile. They’re denser which makes them more durable.
They need to be this way to deal with the stresses of flying, taking off and landing as well as supporting the muscles.
Note that not all the bones are pneumatized, only the limbs are. The spine, scapula, rib cage, skull and pelvis may have a slightly thinner layer of compact hone hut its still of the same density as the limbs, in these hones is where red marrow is. which is important for the formation of red blood cells..
Rib Barbs.
These arc present to strengthen the ribcage, overlapping so that they separate too far and become damaged
Straight on Back View. Hummingbird Dove
There isn’t just one size or shape to wings, just take a look at birds.
The length of the hones, how thin or thick they are and what they are designed to do all have an impact on shape, flight speed and loading weight.
The bones w ould be proportional to body size as they are in birds, and even though they may look liny, the flight leathers are w hat make up the length and shape.
Smaller wings are usually faster and more agile whilst larger wings are more powerful and are for long distance travel. Feathers also make a difference to flying slvle but that w ill be on a different sheet.
Starling
Smaller w ings have a much shorter humerus, hut the bones in manus arc usualy proportionally the saint4 length.
A shorter humerus usualy means that the wing can beat faster and is more powerful than those of longer proportions.
Albatross Smaller wings
also usualy mean larger chest muscles.
When the wings fold the bones wont touch because there are muscles and feathers which prevent that.
Hummingbird Dove
There isn’t just one size or shape to wings, just take a look at birds.
The length of the hones, how thin or thick they are and what they are designed to do all have an impact on shape, flight speed and loading weight.
The bones w ould be proportional to body size as they are in birds, and even though they may look liny, the flight leathers are w hat make up the length and shape.
Smaller wings are usually faster and more agile whilst larger wings are more powerful and are for long distance travel. Feathers also make a difference to flying slvle but that w ill be on a different sheet.
Starling
Smaller w ings have a much shorter humerus, hut the bones in manus arc usualy proportionally the saint4 length.
A shorter humerus usualy means that the wing can beat faster and is more powerful than those of longer proportions.
Albatross Smaller wings
also usualy mean larger chest muscles.
When the wings fold the bones wont touch because there are muscles and feathers which prevent that.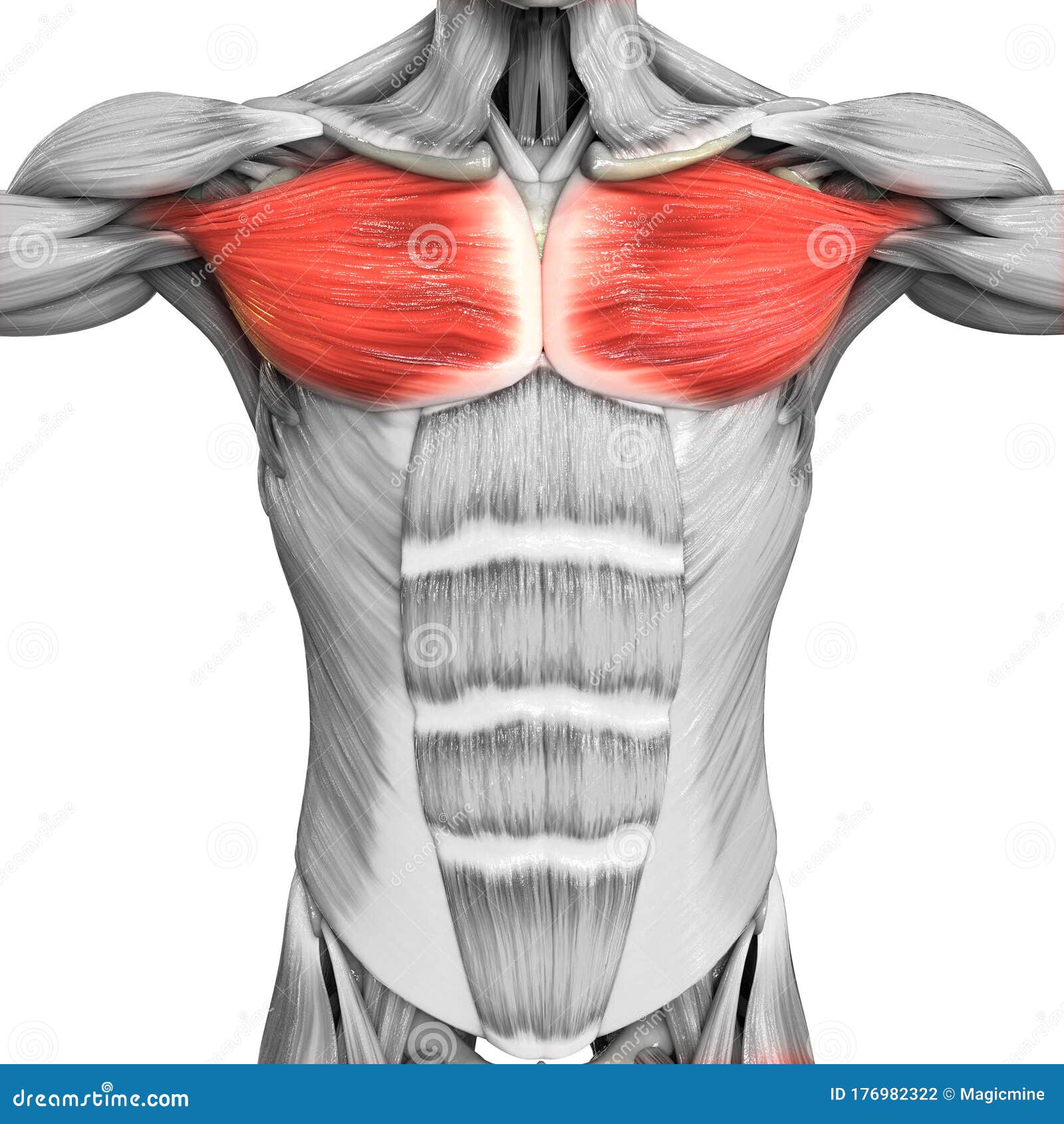 h the le& look mcre(,lb|y lon£ Ibis is what the proportions ot a regular
human skeleton is like*, even so the legs (excluding the feel) are just as long as the radius and humerus of the w ings.
Mote that height and wing size will cause variance.
Tail feathers may be necessary in some cases, if not. the majority.
A larger chest than humans with a sternal keel.
The keel is necessary as the pectoral and w ing muscles attach
r* .
to it. providing extra support for the downward strokes of the wings. There is little hope of having enough pectoral muscle to support the wings if there were no keel.
i luman rib cage from the side
Slightly smaller skull with larger eye sockets and a sagittal keel, a small ridge which serves as armour against any impact to the roof of the skull.
Useful especially if the landing part of flying is not up to scratch.
h the le& look mcre(,lb|y lon£ Ibis is what the proportions ot a regular
human skeleton is like*, even so the legs (excluding the feel) are just as long as the radius and humerus of the w ings.
Mote that height and wing size will cause variance.
Tail feathers may be necessary in some cases, if not. the majority.
A larger chest than humans with a sternal keel.
The keel is necessary as the pectoral and w ing muscles attach
r* .
to it. providing extra support for the downward strokes of the wings. There is little hope of having enough pectoral muscle to support the wings if there were no keel.
i luman rib cage from the side
Slightly smaller skull with larger eye sockets and a sagittal keel, a small ridge which serves as armour against any impact to the roof of the skull.
Useful especially if the landing part of flying is not up to scratch.ангелы,Анатомия,крылья,скелет,крылатые люди.
Еще на тему
Развернуть
Похожие темы
Похожие посты
Программа тренировки «Раскачиваем крылья»Автор: Алан Палмери Источник: CriticalBench.
Большинство бодибилдеров думает, что правильная техника приходит с опытом. К сожалению, ко многим она приходит слишком поздно, когда неправильная техника уже наработана и избавлять от ошибок и переучивать такого атлета бывает очень сложно. Чаще всего бодибилдеру удается добиться утолщения мышечных пластов, а вот ширина… Ширина «крыльев» оставляет желать лучшего. Эта проблема является следствием применения неправильной техники, которая не дается «от рождения», и не приходит сама по себе с годами, а является результатом тщательного обучения и практики. При выполнении тяг бицепсы принимают на себя значительную часть веса. Секрет правильной техники заключается в том, чтобы по возможности исключить влияние бицепсов на выполнение упражнения. Начинающие выполняют упражнение практически за счет силы одних только бицепсов, тем самым препятствуя развитию широчайших и ограничивая результаты, которых они могли бы добиться, с первых же тренировок применяя правильную технику. Считайте свои руки переходным, связующим звеном, необходимым лишь для удержания снаряда и, выполняя движение, концентрируйтесь не на руках, а на мышцах спины. Поначалу такая техника может вызвать определенные трудности, поскольку очень нелегко удерживать руки в неподвижном состоянии при выполнении упражнения. Чтобы получить максимальную пользу от упражнения, мысленно наблюдайте и контролируйте работу ваших мускулов. На примере тяги штанги к поясу в наклоне я опишу пправильную технику выполнения этого движения. Исходное положение: ноги на расстоянии чуть большем ширины плеч, корпус наклоните вперед до параллели с поверхностью пола. Спина прямая. Вгляд направлен вперед. Слегка согните ноги в коленях и захватите гриф примерно на расстоянии ширины плеч. Сконцентрируйтесь на мышцах спины и начинайте движение широчайшими мышцами (не руками). Постарайтесь свести лопатки вместе, до касания. Для большинства бодибилдеров есть только один путь обучения этой технике — постепенно нарабатывать ее, применяя очень легкие веса. Как только овладеете техникой, начинайте увеличивать веса снарядов. Часто самые нетерпеливые сразу же берутся за большие веса. Это неправильно. Несовершенная техника только увеличит риск получения травмы, и направит вас по ложному пути, препятствуя максимальному развитию ваших мышц. В подтягиваниях на перекладине и в тяге штанги в наклоне старайтесь варьировать ширину хвата. Это позволит вам разносторонне проработать мышцы спины. Не думайте ни о чем другом, кроме техники. Пока не приобретете достаточного навыка, тренируйтесь только с легкими весами. Попробуйте следующую программу тренировок, расчитанную на шесть недель. Начните тренировку с тщательной разминки. Можете использовать подтягивания на перекладине в двух подходах из десяти повторов. Если не можете выполнить сразу десять подтягиваний, делайте меньше, но старайтесь через каждые две тренировки добавлять по одному повторению. После того как разогрелись, передохните две-две с половиной минуты и приступайте к целевой части тренировки. Поскольку легкие веса бесполезны для качественной проработке мышц, я предлагаю гигантские сеты, в которых упражнения выполняются одно за другим без перерыва. Как только завершите все упражнения в указанном порядке, что составит один гигантский подход, отдохните около двух минут и приступайте к следующему. Не забывайте, что концентрироваться следует на технике, а не на том, какой вес вы сможете одолеть. Программа предназначена, прежде всего, для того, чтобы помочь вам овладеть «чувством мышцы». Также не следует проводить работать по этой программе чаще одного раза в неделю. Программа Один гигантский сет
После первого сета отдохните около двух минут, и приступайте к следующему подходу. Движение начинайте широчайшими мышцами. Старайтесь свести лопатки вместе, до касания. Не забывайте и о важности такого фактора, как умственная концентрация. Концентрируйтесь на широчайших мышцах спины на протяжении всего сета. И еще я советую, после того как закончите работу на мышцы спины, выполнить несколько сетов к любому упражнению на задние пучки дельтоидов. Это может быть любое упражнения в любом положении — сидя, стоя или в наклоне… В этот же день можно проработать голени и мышцы брюшного пресса. Резюме Старайтесь каждую неделю понемногу увеличивать веса снарядов, но если такое увеличение сказывается на технике, тогда вообще забудьте об увеличении весов. Повторяю, главное — это техника. Опытным бодибилдерам также было бы полезно время от времени возвращаться к этой программе. Часто они грешат небрежным выполнением упражнений, а периодическое возвращение к подобным программам поможет вспомнить правильную технику. И последнее. Я много лет тренирую бодибилдеров и могу со 100% вероятностью предсказать, что девять из десяти человек, прочитавших эту статью, сразу же возьмутся за больше веса, чем сведут на «нет» самое ценное преимущество этой программы. Не торопитесь. Прежде «обучите» свои широчайшие правильной работе, без помощи других мышечных групп. Умерьте свои амбиции, в конечном итоге это пойдет вам только на пользу. |
 com
com
 Сила бицепсов, предплечий и хвата должны стать второстепенным фактором при выполнении упражнений, направленных на развитие мышц спины.
Сила бицепсов, предплечий и хвата должны стать второстепенным фактором при выполнении упражнений, направленных на развитие мышц спины. Не сгибайте руки, держите их полностью выпрямленными. Помните, что руки — всего лишь связующее звено между вами и снарядом. Медленно вернитесь в исходное положение. Повторите упражнение. Эту технику можно применять во всех упражнениях для мышцы спины.
Не сгибайте руки, держите их полностью выпрямленными. Помните, что руки — всего лишь связующее звено между вами и снарядом. Медленно вернитесь в исходное положение. Повторите упражнение. Эту технику можно применять во всех упражнениях для мышцы спины.
 Вы должны выполнить три гигантских подхода. Концентрируйтесь на широчайших, ослабляйте хват на грифе, и используйте руки только как связующее звено.
Вы должны выполнить три гигантских подхода. Концентрируйтесь на широчайших, ослабляйте хват на грифе, и используйте руки только как связующее звено.
Лицевые нервные тики
39072 11 СентябряВАЖНО!
Информацию из данного раздела нельзя использовать для самодиагностики и самолечения. В случае боли или иного обострения заболевания диагностические исследования должен назначать только лечащий врач. Для постановки диагноза и правильного назначения лечения следует обращаться к Вашему лечащему врачу.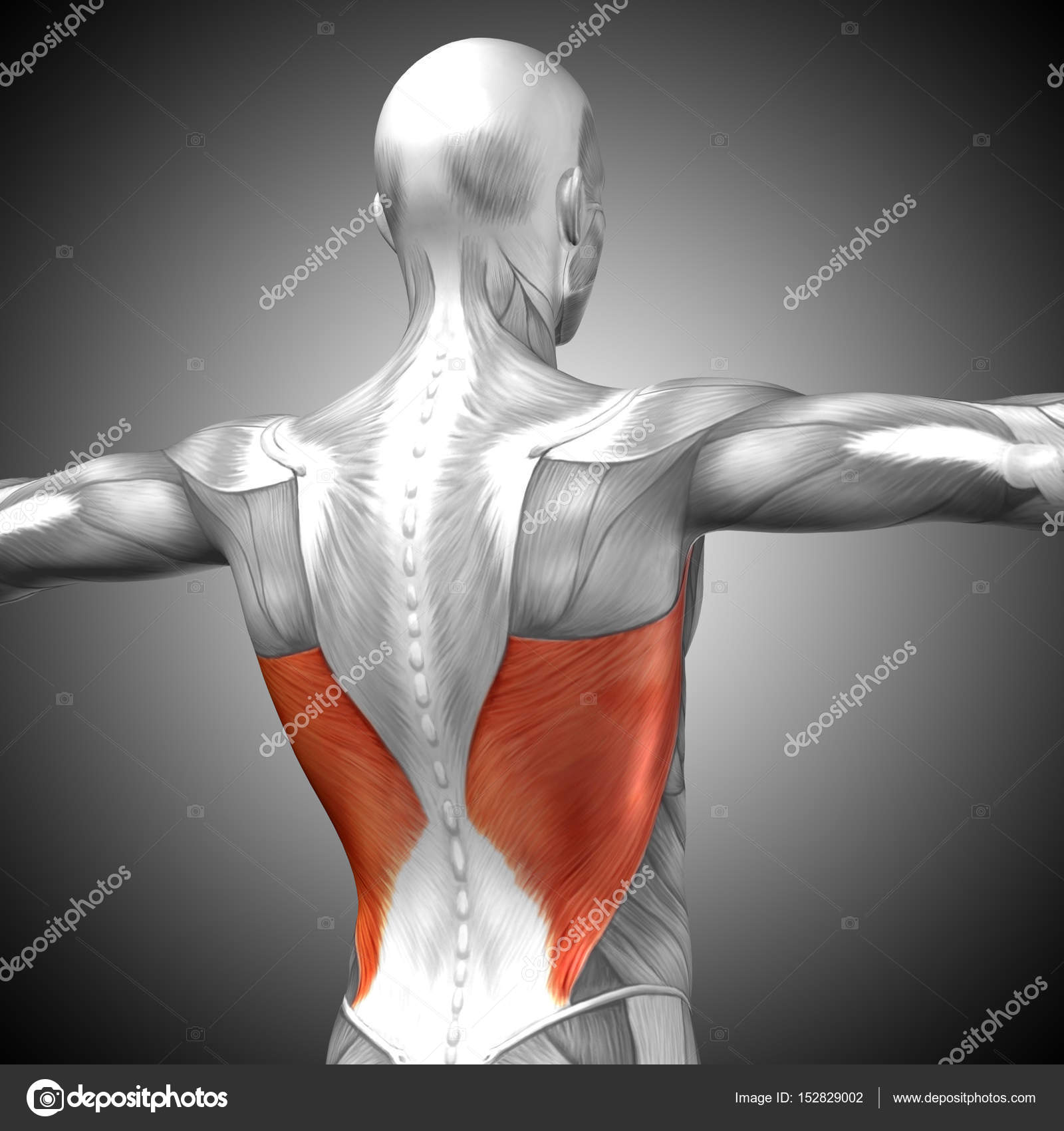
Лицевые нервные тики — причины появления, при каких заболеваниях возникает, диагностика и способы лечения.
Тики, или гиперкинезы – это повторяющиеся, непроизвольные, короткие стереотипные движения, внешне схожие с контролируемыми действиями. Чаще всего тики поражают мышцы лица, что сопровождается морганием, наморщиванием лба, раздуванием крыльев носа, облизыванием губ и различными гримасами.
Разновидности тиков
При различных заболевания гиперкинезы могут быть как единственным неврологическим проявлением (первичные формы), так и служить симптомом других расстройств нервной системы.
В числе органических (т. е. вызванных повреждением структуры мозга) выделяют гиперкинетические синдромы с преимущественным вовлечением мимических мышц, оральные (ротовые) гиперкинезы, лицевой гемиспазм (одностороннее непроизвольное сокращение лицевых мышц), постпаралитическую контрактуру лицевых мышц (результат невропатии лицевого нерва с неполным восстановлением мышечных функций). К этой же группе относят лицевые гиперкинезы, которые сочетаются или возникают на фоне других гиперкинезов и неврологических синдромов. При органических гиперкинезах в процесс могут вовлекаться не мимические, а другие мышцы: глазодвигательные, жевательные, шейные.
К этой же группе относят лицевые гиперкинезы, которые сочетаются или возникают на фоне других гиперкинезов и неврологических синдромов. При органических гиперкинезах в процесс могут вовлекаться не мимические, а другие мышцы: глазодвигательные, жевательные, шейные.
В группе неорганических тиков различают психогенные, или невропатические гиперкинезы и стереотипии (устойчивое бесцельное повторение движений, слов или фраз).
Возможные причины и заболевания, при которых встречаются тики
Чаще всего тики начинаются в детском и подростковом возрасте. Мальчики страдают в 2-4 раза чаще, чем девочки.
Запускающим фактором для развития гиперкинезов могут быть перинатальные травмы, инфекции (вирусный энцефалит, летаргический энцефалит, ВИЧ, туберкулез, сифилис и т. д.), опухоли, сосудистые поражения, психологические нарушения и лекарственные препараты.
Тики с участием мимических мышц могут затрагивать различные области лица. Например, блефароспазм (непроизвольное сокращение круговой мышцы глаза) характеризуется усиленным морганием и зажмуриванием глаз. Развитие болезни обычно происходит постепенно, первоначальное ощущение раздражения или сухости глаз сменяется морганием, эпизодами длительного зажмуривания и закрывания глаз, особенно при ярком свете. В необычной обстановке непроизвольные движения могут исчезать. Такого рода тик возникает при органических заболеваниях головного мозга (болезнь Паркинсона, рассеянный склероз и др.), а также при сосудистых, воспалительных, метаболических и токсических поражениях нервной системы. Провоцирующим фактором могут стать длительные или кратковременные воздействия на область лица при стоматологических процедурах, травмах, операциях, воспалительных заболеваниях глаз, синуситах.
Например, блефароспазм (непроизвольное сокращение круговой мышцы глаза) характеризуется усиленным морганием и зажмуриванием глаз. Развитие болезни обычно происходит постепенно, первоначальное ощущение раздражения или сухости глаз сменяется морганием, эпизодами длительного зажмуривания и закрывания глаз, особенно при ярком свете. В необычной обстановке непроизвольные движения могут исчезать. Такого рода тик возникает при органических заболеваниях головного мозга (болезнь Паркинсона, рассеянный склероз и др.), а также при сосудистых, воспалительных, метаболических и токсических поражениях нервной системы. Провоцирующим фактором могут стать длительные или кратковременные воздействия на область лица при стоматологических процедурах, травмах, операциях, воспалительных заболеваниях глаз, синуситах.
Кроме того, отмечена связь появления тиков с профессиональными вредностями (длительное напряжение зрения и мимических мышц).
Иногда за тик можно принять доброкачественную миокимию век – преходящие подергивания круговой мышцы глаз при переутомлении, волнении, повышенном употреблении кофе или курении, – которая не требует лечения.
Если при тике задействованы мышцы рта, языка и челюстей, такие расстройства называют оральными гиперкинезами. Как правило, их возникновение обусловлено применением нейролептиков, гормональных препаратов, блокаторов дофаминовых рецепторов. Однако тики в области лица могут возникать с возрастом (у пожилых людей после 60-70 лет) без приема нейролептиков. Насильственные движения при этом обычно начинаются с мышц языка, могут вовлекаться щеки и нижняя челюсть. Едва уловимые движения языком со временем переходят в нерегулярные, но частые движения языка, губ и нижней челюсти – облизывание, сосание, жевание. Во время еды, разговора дискинезия прекращается.
Разновидностью органических гиперкинезов является лицевой гемиспазм (как самостоятельное заболевание и вследствие сдавления лицевого нерва). Он проявляется серией коротких быстрых подергиваний, в основном вокруг глаза.
При этом характерно прищуривание или зажмуривание глаза, поднятие щеки и угла рта вверх. В течение суток наблюдаются сотни приступов. Произвольное зажмуривание иногда провоцирует гиперкинез, точно так же, как и эмоциональное напряжение.
Лицевые тики чаще всего бывают проявлением более распространенных по охвату мышц гиперкинезов и неврологических синдромов. К их числу относится известный синдром Туретта, который характеризуется двигательными и голосовыми тиками, дефицитом внимания и навязчивым состоянием. У мальчиков тики чаще сочетаются с синдромом дефицита внимания и гиперактивностью, а у девочек – с навязчивыми состояниями. Первые симптомы появляются в 3-7 лет и заключаются в ограниченных лицевых тиках и подергиваниях плеч. Затем охватываются мышцы верхних и нижних конечностей. Обычно максимальная выраженность заболевания отмечается в подростковом периоде. С возрастом тики уменьшаются или исчезают, а если сохраняются, то редко приводят к инвалидизации.
К генерализованным тикам относится лекарственная дискинезия, вызванная приемом антидепрессантов, леводопы (при лечении паркинсонизма). Признаки дискинезии возникают, как правило, через 2-12 недель после начала лечения нейролептиками и охватывают не только мышцы лица, но и тела.
В числе достаточно частых генерализованных тиков отмечаются хореические гиперкинезы (при хорее Гентингтона, доброкачественной наследственной хорее и хорее Сиденгама). Первые два заболевания имеют наследственную природу. Хорея Сиденгама возникает как осложнение после стрептококковой инфекции и представляет собой изолированное неврологическое проявление ревматизма. Обычно наблюдается симметричность тиков, которые проявляются быстрыми подергиваниями в области лица, туловища и конечностей. Тики сначала охватывают одну мышечную группу, затем перемещаются к другой; на фоне стресса они усиливаются, а во время сна исчезают.
Отдельно следует упомянуть лицевые гиперкинезы эпилептической природы, во время которых отмечаются повторяющиеся, ритмичные и быстрые подергивания век. Они могут сочетаться с подергиваниями рук. Каждый приступ сопровождается, по меньшей мере, тремя следующими друг за другом сокращениями век.
Достаточно большая группа лицевых гиперкинезов представлена тиками на почве неврозов, которые вызывают психологические факторы – страх, эмоциональное потрясение, психотравма.
Человек осознает невротические тики, но невозможность контролировать насильственные движения доставляет дискомфорт. Интересное занятие может отвлечь, но при волнении и переутомлении тики возобновляются. Сознательная задержка тиков часто приводит к нарастанию внутреннего напряжения и ответной реакции в виде головной боли, раздражительности и агрессивности. О психогенной природе гиперкинеза могут свидетельствовать: острое начало, непостоянство гиперкинеза с ремиссиями и обострениями, отсутствие реакции на стандартную терапию и наличие возможной выгоды, которую пациент стремиться извлечь из заболевания.
К каким врачам обращаться?
При появлении тиков у взрослых необходим осмотр невролога.
Если тики возникли у ребенка, необходима срочная консультация
педиатра и затем детского невролога. При подозрении на наследственную природу заболевания необходима консультация генетика.
Диагностика и обследования
При появлении тиков в любом возрасте невролог оценивает психосоматический статус и клиническую картину заболевания, выявляя связь с другими симптомами, наличием предшествующих травм и заболеваний. Биохимический анализ крови необходим для диагностики гиперкинезов дисметаболической и токсической этиологии.
Велосипед в небесах – Наука – Коммерсантъ
Самолет американского авиаконструктора Пола Мак-Криди Gossamer Condor («Паутинка Кондор») с педальным приводом к пропеллеру совершил полет по маршруту в виде восьмерки, облетев две вешки, стоявшие на расстоянии километра друг от друга. Такими были условия энтузиаста мускульной авиации, британского предпринимателя Кремера, учредившего премию в 50 тыс. фунтов стерлингов за полет длиной не меньше мили по замкнутому маршруту с разворотом.
фунтов стерлингов за полет длиной не меньше мили по замкнутому маршруту с разворотом.
Летать, как птицы или хотя бы птеродактили, размахивая крыльями, человеку не позволяет его анатомия: грудные мышцы слишком слабые, а кости скелета слишком тяжелые. Ноги сильнее, поэтому мускулолеты исходно строились по принципу летающего велосипеда. Различались они только движителем: у одних это был пропеллер, как у самолета, у других машущие перепончатые крылья, как у летучих мышей. Пропеллер оказался выгоднее, и после войны к машущим крыльям конструкторы уже не возвращались.
За все сто лет мускульного воздухоплавания в воздух поднялись около сотни летающих велосипедов разных конструкций. Сначала они были скорее подпрыгивающими, чем летающими. Запуски мускулолетов с катапульты давали лучшие результаты. Только это был не полет, а поддерживаемое педалями планирование, удержаться в воздухе долго сил у пилота не хватало. После войны все рекорды катапультных мускулолетов были аннулированы.
Появление углепластиков во второй половине XX века дало новые возможности. Уже второй мускулолет Мак-Криди Gossamer Albatross в 1979 году пересек Ла-Манш (33 км). А в 1988 годуDaedalus-88, построенный студентами Массачусетского технологического института, пролетел 115 км с одного греческого острова на другой, и пока это рекорд.
Уже второй мускулолет Мак-Криди Gossamer Albatross в 1979 году пересек Ла-Манш (33 км). А в 1988 годуDaedalus-88, построенный студентами Массачусетского технологического института, пролетел 115 км с одного греческого острова на другой, и пока это рекорд.
Рекордно малым был и вес «Альбатроса» и «Дедала» — 30 кг при размахе крыльев 30 и 35 м. Но даже если мускулолет будет в буквальном смысле легче паутинки, сам пилот весит около 70 кг. Чтобы создать достаточную для его веса подъемную силу крыла, надо развить и поддерживать усилие мощностью в 1/3 лошадиной силы. Профессиональные велогонщики развивают при спурте усилие больше 0,5 л. с., но для обычного велосипедиста 0,3 л. с.— это предел. Крутили педали «Альбатроса» и «Дедала» как раз велогонщики, пилот «Дедала» даже участвовал Олимпийских играх.
Таким образом, воздушный велосипед, построенный по принципу самолета или птицы, — тупиковая ветвь мускулолетов, он никогда не станет массовым. Вероятно, придется поискать другие принципы полета. Например, среди насекомых.
Например, среди насекомых.
Сергей Петухов
Маркеры обмена веществ позволяют определить, не слишком ли резко вы взяли старт
Олимпиада, приближение весны или скидки на клубные карты — есть много факторов, побуждающих нас вступить на путь оздоровления. Тут только надо убедиться, что эта дорога, выложенная, без сомнения, добрыми намерениями, ведет именно туда – к здоровью.
Всем нам свойственно бросаться в новое дело очертя голову. Вы чувствуете себя спортсменом и полны решимости перепробовать все тренажеры в спортзале – отлично, а организм об этом знает? И даже если на первые занятия взяли опытного тренера (что разумно), он не всегда может сделать из ваших слов правильные выводы.
Скажем, после тренировки все болит — это хорошо или плохо? Это смотря как болит, может, это показатель хорошей тренировки, а может, человек на пути к инфаркту. Существуют ли объективные показатели того, как организм реагирует на нагрузки? Да.
Если у вас появились новые нагрузки, значит, клеткам нужно больше питания, больше кислорода, быстрее надо удалять отходы, и организм честно старается все это обеспечить, а значит, меняется что? Состав крови.
Самое интересное, что эти показатели работают для всех, кто начал регулярно заниматься спортом, не важно, включили вас в олимпийскую сборную или вы неделю назад не могли без одышки донести тарелку до раковины. Казалось бы, как же так, ведь показатели крови должны быть такие разные?! Признаемся честно: они у всех людей разные. За редким исключением значение имеет изменение данных конкретного человека во времени.
Пять способов оценить адекватность нагрузки
Молочная кислота (лактат) — «отход производства», знакомый всем без исключения. Стоит позаниматься чуть больше обычной нормы, и мышцы сводит ноющая боль. Откуда она берется? Когда мышцы работают больше обычного, им нужно больше энергии. Организм получает энергию, окисляя глюкозу. Для этого нужны глюкоза и кислород. Запасов глюкозы в организме достаточно, а вот запасов кислорода в организме нет, его надо поставлять извне.
Организм получает энергию, окисляя глюкозу. Для этого нужны глюкоза и кислород. Запасов глюкозы в организме достаточно, а вот запасов кислорода в организме нет, его надо поставлять извне.
При увеличении нагрузки кислорода не хватает и глюкоза окисляется не до углекислого газа, а только до молочной кислоты. А потом откладывается в мышцах и причиняет дискомфорт, пока организм ее тоже потихоньку не усвоит. При регулярных тренировках поступление кислорода в организм растет и уровень лактата снижается. Снова увеличили нагрузку – содержание лактата снова растет, пока организм не адаптируется.
Слабо тренированным людям бывает трудно отличить эту боль от дискомфорта в растянутых мышцах. А вот спортсмены часто воспринимают эти ощущения как приятные, особенно после перерыва в тренировках – им организм так говорит: «Молодец, отлично потренировался».
Мочевина – очень простое и крайне интересное вещество.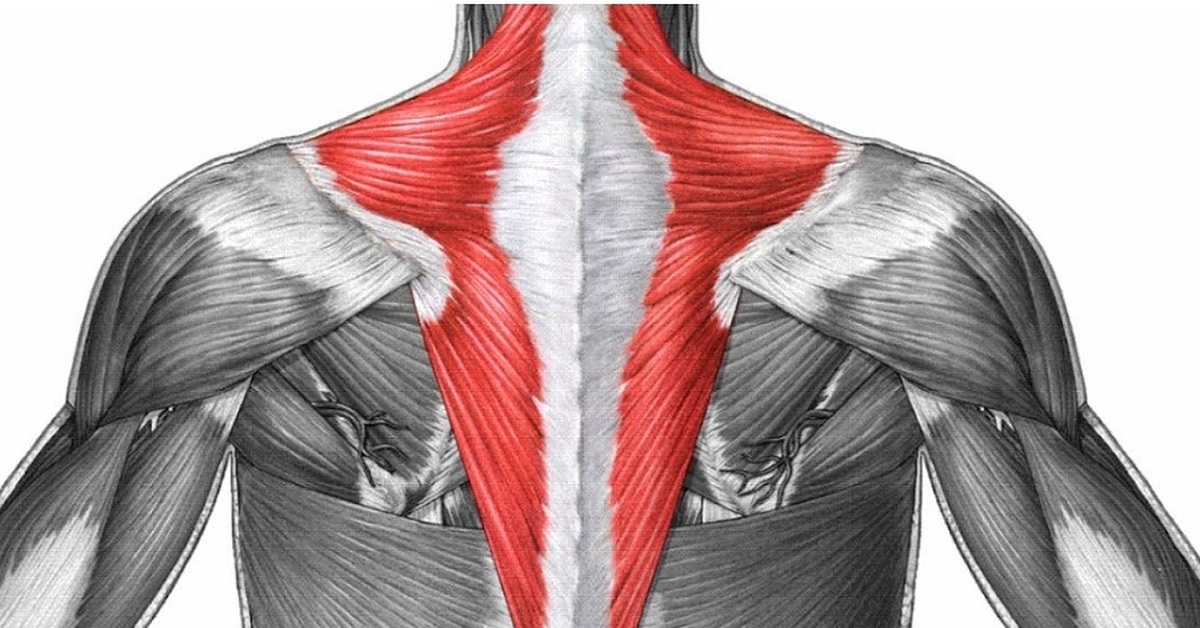 Повышение уровня мочевины в крови говорит о том, что организм под действием стресса утилизирует собственные белки, не в силах ждать, когда ему поставят те самые запасы глюкозы. Обмен веществ после увеличения нагрузки похож на неопытного спортсмена, который, вернувшись из спортзала, набрасывается на холодильник, сметая с полок все без разбора. При равномерной нагрузке организм разбирает, что к чему, налаживает поставки дополнительной глюкозы, и уровень мочевины снижается.
Повышение уровня мочевины в крови говорит о том, что организм под действием стресса утилизирует собственные белки, не в силах ждать, когда ему поставят те самые запасы глюкозы. Обмен веществ после увеличения нагрузки похож на неопытного спортсмена, который, вернувшись из спортзала, набрасывается на холодильник, сметая с полок все без разбора. При равномерной нагрузке организм разбирает, что к чему, налаживает поставки дополнительной глюкозы, и уровень мочевины снижается.
Креатинкиназа. Простые сигналы закончились. Итак, увеличение нагрузки – стресс для организма, ему не хватает энергии, и он не знает, где ее взять. Кто приходит на помощь в этот момент? Креатинкиназа – фермент — «антикризисный управляющий», он работает в скелетных мышцах, сердце, легких, мозге, то есть во всех ключевых точках, и отвечает за энергоснабжение клеток, пока организм перестраивается под новые условия. Неудивительно, что высокий уровень этого белка сигнализирует о чрезмерных нагрузках, инфаркте и травмах. При равномерных нагрузках организм стабилизирует обмен веществ, и уровень фермента падает.
При равномерных нагрузках организм стабилизирует обмен веществ, и уровень фермента падает.
Миоглобин – родной брат гемоглобина, только он переносит кислород не в крови, а в мышцах. Забирает кислород у гемоглобина и отдает его клеткам для получения энергии, в которой они так нуждаются. Миоглобин – то самое редкое исключение, чей уровень в крови не уникален для каждого человека. Потому что в норме количество его в крови минимальное, его вообще не должно быть. Более того, в крови он токсичен для организма.
Повышенное содержание миоглобина в крови кричит о повреждении мышц, а значит, стоит внимательно отнестись к своим нагрузкам.
Тропонин I – если миоглобин сигналит о повреждении любых мышц, то тропонин I, специфический маркер повреждения сердечной мышцы, предупреждает, что дело серьезное. Высокий уровень тропонина — всегда негативный признак и веский повод обратиться к врачу. Человек при этом может не ощущать никаких тревожных симптомов, и даже опытный тренер не найдет повода для беспокойства, но если уровень тропонина I высок, знайте: вы очень круто взяли. Он как предупреждающий знак на дороге – требует снизить скорость, потому что впереди опасность.
Он как предупреждающий знак на дороге – требует снизить скорость, потому что впереди опасность.
Есть, как нетрудно догадаться, две основные группы риска: спортсмены с серьезными амбициями, которые испытывают высокую кардионагрузку, и любители спорта на диване, внезапно решившие взять с места в карьер, не посоветовавшись с организмом.
Татьяна Мельникова, терапевт, врач-консультант Независимой лаборатории ИНВИТРО, отмечает: «Главное в спортивных тренировках – равномерные нагрузки, адекватные состоянию вашего организма. Человек может выдержать очень высокий темп тренировок, главное — повышать его постепенно. Увеличили нагрузку – задержитесь на ней, дайте организму адаптировать метаболизм, проконтролируйте свое состояние и только после этого двигайтесь дальше. Организм как океан, он не может сразу подстроиться к новой ситуации, а вот за постепенные и регулярные тренировки он воздаст отличной физической формой».
Организм как океан, он не может сразу подстроиться к новой ситуации, а вот за постепенные и регулярные тренировки он воздаст отличной физической формой».
Ознакомиться с другими новостями, материалами и статистикой вы можете на странице летних видов спорта.
Хороший вопрос: могли бы люди летать, если бы у нас были крылья?
Большинство из нас немного завидуют птицам. Мы давно хотели быть похожими на наших птичьих друзей, о чем свидетельствует история с эскизами птиц Икара и Леонардо да Винчи и его идеи о летающих машинах.
Да, сегодня мы можем летать, но не махая руками. Но при всех наших знаниях и технологиях, почему мы не можем купить пару крыльев в местном магазине Walmart или даже вырастить их сами? Я бы с радостью избавился от своей машины и промчался по воздуху! (А на обед я ходил в ресторан с проезжей на крыше.)
К сожалению, наука против этой мечты.
Согласно статье в Yale Scientific, «людям математически невозможно летать, как птицы». Во-первых, крылья — и размах, и сила — находятся в равновесии с размером тела птицы. Кроме того, у птиц в основном полые кости (в основном полые, потому что они усилены перекрещивающимися стойками), которые уменьшают их массу и создают воздушные карманы, чтобы дать им больше кислорода во время полета.
Во-первых, крылья — и размах, и сила — находятся в равновесии с размером тела птицы. Кроме того, у птиц в основном полые кости (в основном полые, потому что они усилены перекрещивающимися стойками), которые уменьшают их массу и создают воздушные карманы, чтобы дать им больше кислорода во время полета.
Это не потому, что мы слишком большие. Мы слишком слабы.Животные, намного крупнее людей, поднялись в небо, такие как кетцалькоатль, огромный птерозавр.
Сравнение человека с двумя разновидностями quetzalcoatlus. | С любезного разрешения Мэтт Мартынюк, Марк Уиттон и Даррен Нейш, Wikimedia «Расчеты соотношения между размером и силой человека показывают, что наш вид никогда не сможет взлететь без посторонней помощи», — говорится в статье Yale Scientific. «По мере роста организма его вес увеличивается быстрее, чем его сила. Таким образом, среднему взрослому человеку мужского пола потребуется размах крыльев не менее 6.7 метров до полета. Этот расчет даже не принимает во внимание, что сами эти крылья были бы слишком тяжелыми, чтобы функционировать ».
Другими словами, нам понадобятся крылья побольше. Но чем больше крылья, тем больше масса, а это значит, что нам понадобятся крылья еще большего размера, и… вы понимаете. Мы будем гоняться за своими хвостами, чтобы сделать крылья.
Предоставление людям рабочих крыльев не означало бы просто дополнительных или трансформированных придатков — нам пришлось бы заново спроектировать человеческое тело с нуля.
Но это не мешает людям надеяться.
Взгляните на этого парня, который в 2012 году штурмом захватил Интернет.
Какое захватывающее видео! Мечты миллионов осуществились за несколько секунд!
Это был розыгрыш. Ознакомьтесь с объяснением Сноупса и нажмите здесь, чтобы узнать, почему так сложно построить крылатый самолет с приводом от человека.
А что, если бы у нас могли быть крылья для украшения?
Я не говорю о картонных крыльях, которые маленькие дети носят в рождественских спектаклях. Я имею в виду крылья из плоти и крови. Было бы так… ангельски!
Доктор Джо Розен, пластический хирург из Дартмутской медицинской школы, говорит, что человеческие крылья возможны.
«Если бы я дал вам крылья, у вас буквально развился бы крылатый мозг. Наши тела изменяют наш мозг, и наш мозг бесконечно пластичен », — цитирует Розена в« Гардиан »в 2002 году.
« Уже существующие хирургические методы могут быть использованы для растягивания жира туловища и изменения ребер костей для создания крыла.Хотя ни один человек не сможет летать, они будут напоминать ангелов и полностью ощутить свои новые висящие, костлявые лоскуты плоти », — говорится в статье.
Однако большинству из нас придется довольствоваться этими крыльями:
Stock photo
У вас есть хороший вопрос о чем-нибудь? Отправь его Роберту! Напишите ему по адресу [email protected] или посетите его в Twitter @notthegeneral или Facebook.
Удивительные мышцы и кости, благодаря которым птицы летают
Мышцы, которые заставляют крылья двигаться вниз и вверх, выделены красным выше. Внизу показаны кости и сухожилия, составляющие оригинальный шкив птицы. Иллюстрация Дениз Такахаши.
Внизу показаны кости и сухожилия, составляющие оригинальный шкив птицы. Иллюстрация Дениз Такахаши.Полет необходим для птиц и увлекателен для орнитологов. Рассмотрим скорость ныряющего сокола, всплеск покрасневшего тетерева, беспорядочный образ ухаживания колибри или выносливость дальних мигрантов.
Каждый из них демонстрирует силу полета, для которой требуются мышцы настолько массивные, что они могут составлять треть или более веса тела птицы. Однако, когда птицы впервые появились на сцене, типичный план скелета позвоночных не мог вместить такие большие мышцы.
Птицам пришлось использовать свои инженерные навыки и внести некоторые изменения. (На самом деле естественный отбор благоприятствовал чертам, которые способствовали полету.) Чтобы понять проблему, рассмотрите нептичье позвоночное, такое как мы, люди, и подумайте о руке как о крыле.
У нас есть большая выступающая грудная мышца, называемая pectoralis major , которая берет начало вдоль грудины или грудины и прикрепляется к головке верхней кости руки (плечевой кости). Когда грудная мышца сокращается, она приближает руку к телу.(Это движение подобно удару птицы вниз.) Бодибилдеры могут наращивать грудные мышцы, но плоская грудина позвоночных не имеет достаточной площади для прикрепления увеличенных мышц, необходимых птицам.
Когда грудная мышца сокращается, она приближает руку к телу.(Это движение подобно удару птицы вниз.) Бодибилдеры могут наращивать грудные мышцы, но плоская грудина позвоночных не имеет достаточной площади для прикрепления увеличенных мышц, необходимых птицам.
Чтобы поднять руку, мы используем меньшую мышцу, известную как дельтовидная, в верхней части плеча. Положите руку на край плеча и поднимите руку. Вы почувствуете легкую выпуклость дельтовидной мышцы. Анатомия плеча позвоночного не предусматривает места для прикрепления более крупной мышцы, поднимающей руку, и это место, к сожалению, не соответствует требованиям птицы.Кстати, поскольку большая грудная мышца большая, а дельтовидная мышца маленькая, вы можете опустить руку вниз с гораздо большей силой, чем поднять.
Первым птичьим решением было добавить к грудины вертикальный киль. Киль резко увеличивает площадь поверхности для прикрепления мышц. Как вы можете видеть на диаграмме выше, горизонтальная грудина образует Т с вертикальным килем. На верхнем рисунке большая грудная мышца, нижняя мышца киля, показана красным цветом. Он вставляется в плечевую кость и сокращается, тянет крыло вниз.
На верхнем рисунке большая грудная мышца, нижняя мышца киля, показана красным цветом. Он вставляется в плечевую кость и сокращается, тянет крыло вниз.
Поскольку мышцы функционируют только за счет укорачивания, общепринятое мнение гласит, что мышца должна располагаться над крылом, чтобы поднимать его. Но птицы поднимают крылья с помощью большой мышцы, расположенной под крылом. Прикрепленная к килю грудины, мышца, известная как supracoracoideus , соединяется с верхней частью плечевой кости с помощью шкива — изобретательного механизма, который больше нигде не встречается у позвоночных.
Прочтите другие колонки Элдона Грейя.
Supracoracoideus, показанная белым на верхнем рисунке, находится чуть выше грудной мышцы.Его сухожилие перекидывается через плечо, вставляясь в верхнюю часть плечевой кости. Супракоракоидус показан красным на среднем рисунке. Он сжимается и из-за шкива поднимает крыло.
Как вы можете видеть внизу на схеме, шкив расположен там, где три кости — клювовидная, лопатка и ключица (не показаны) — соединяются, образуя плечевой сустав. Сухожилие скользит по рифленой головке коракоида.
Сухожилие скользит по рифленой головке коракоида.
Коракоиды — большие крепкие кости, соединяющие грудину с плечом.Плечевой сустав очень похож на чашевидную ладонь. Чтобы визуализировать это, представьте, что кулак вашей правой руки является головкой плечевой кости, и поместите его в сложенную чашевидную форму левой ладони. Теперь представьте, что сухожилие супракоракоидуса проходит над вашей левой рукой и входит в верхнюю часть правого запястья, а сухожилие большой грудной мышцы входит под запястье. Держа запястье неподвижно, двигайте локтем вверх и вниз, чтобы имитировать чередующиеся сокращения супракоракоидной мышцы (подъем вверх) и большой грудной мышцы (движение вниз).
Птицы претерпели множество адаптивных изменений к полету. К числу наиболее значительных можно отнести резкое увеличение мышц груди и изменения скелета, в которые они входят, а также разработка уникальной системы шкивов, которая позволяет мышце, расположенной под крылом, поднимать ее. Все это еще раз указывает на удивительное строение и функции птиц. — Элдон Грейдж, редактор-основатель
— Элдон Грейдж, редактор-основатель
Учиться во время еды
Чтобы оценить приспособление к полету, нарежьте курицу-гриль.
Осторожно удалите всю грудку, обнажив вертикальный киль, прикрепленный к плоской грудине. Обратите внимание, насколько велика грудная мышца по сравнению с размером тела и насколько киль увеличивает площадь поверхности для прикрепления грудных мышц.
Когда вы сокращаете мышцу перед килем, вы обнаружите V-образные сросшиеся ключицы, также известные как поперечные рычаги. Сразу за ним две крепкие клювовидные кости. Обратите внимание на то, как они прикрепляются к передней части грудины и к основанию крыла, и визуализируйте, как кости служат одновременно опорами для взмахов крыльев и столпами, которые удерживают сокращения летательных мышц от сжатия грудной клетки.
Эта статья из колонки Элдона Грейджа «Удивительные птицы» появилась в январском / февральском выпуске BirdWatching за 2014 год.
Иллюстрация Дениз Такахаши. Первоначально опубликовано
Первоначально опубликовано
Читайте нашу рассылку!
Подпишитесь на нашу бесплатную электронную рассылку, чтобы получать новости, фотографии птиц, советы по привлечению и идентификации и многое другое, доставляемое на ваш почтовый ящик.
Подпишитесь бесплатнограниц | Эволюция сократимости и энергетической эффективности летных мышц
Введение
Полет — занятие, требующее больших затрат энергии для животных. Мышечные полеты эволюционировали в классы Insecta (насекомые) и Aves (птицы) и в отряд Chiroptera млекопитающих (летучие мыши). Интересно исследовать, как летные мускулы этих филогенетически далеких видов эволюционировали, чтобы соответствовать требованиям высокоэффективных и устойчивых сокращений, чтобы приводить в движение их крылья во время полета.Мышцы полета насекомых и позвоночных — это поперечно-полосатые мышцы с похожей саркомерной структурой, а также моторные и регуляторные белки миофиламентов. Сравнение их сходства, а также уникальных характеристик вместе с нелетными мышцами для конвергентного эволюционного отбора может помочь понять молекулярную основу сократительной способности и энергетической эффективности, необходимых для летных мышц.
Сравнение их сходства, а также уникальных характеристик вместе с нелетными мышцами для конвергентного эволюционного отбора может помочь понять молекулярную основу сократительной способности и энергетической эффективности, необходимых для летных мышц.
Эволюция механического полета у насекомых, птиц и летучих мышей
Летная способность насекомых, птиц и летучих мышей является типичным примером конвергентной эволюции.Эти животные независимо эволюционировали от разных предков в разное время с аналогичной функцией силового полета в результате сходного естественного селективного давления (Gleiss et al., 2011).
Истоки полета
Определенные общие факторы окружающей среды могли иметь решающее значение для возникновения и конвергентной эволюции полета животных. Геофизические данные показывают, что резкое повышение уровня кислорода в атмосфере происходило дважды в истории Земли (Lyons et al., 2014). Первое повышение уровня кислорода до 35% произошло ∼360 миллионов лет назад (Mya) в результате земледелия растений и глобального отложения углерода в период от позднего девона до позднего каменноугольного периода, когда появились летающие насекомые (Dudley, 2000), а второе повышение уровня кислорода до 25–30% приходилось на позднюю юрскую и меловую эры, когда появились летающие позвоночные, т. Е. Птицы (∼150 млн лет назад) и летучие мыши (∼52 млн лет назад) (Рисунок 1) (Dudley, 2000).
Е. Птицы (∼150 млн лет назад) и летучие мыши (∼52 млн лет назад) (Рисунок 1) (Dudley, 2000).
Рисунок 1. Эволюционное появление насекомых, птиц и летучих мышей.Филогенетическое дерево, обобщенное на основе литературных данных (Nelson and Tidwell, 1987; Tokita et al., 2012; Brusatte et al., 2015), иллюстрирует геологические времена, когда классы Insecta и Aves и отряд Chiroptera летающих животных появился. Насекомые начали летать около 400 миллионов лет назад (млн лет назад), древние летающие птицы появились 150 млн лет назад, а летучие мыши появились около 50 млн лет назад. Историческая кривая содержания кислорода в атмосфере (Dudley, 1998) иллюстрирует аналогичные условия гипероксии, когда у этих летающих животных возникло аналогичное воздушное передвижение.
Бескрылые насекомые появились 395–390 млн лет назад, и, хотя происхождение летающих насекомых неясно, предком, вероятно, был гексапод крылатого животного (крылатое насекомое), на что указывают самые ранние окаменелости, датированные примерно 325 млн лет назад (рис. 1) (Nelson and Tidwell, 1987). Морфологическое происхождение крыльев остается спорным, поскольку нет прямых доказательств переходных форм между бескрылыми и крылатыми насекомыми. Убедительная теория, основанная на примитивной летописи окаменелостей нимф крылатых насекомых, предполагает, что крылья насекомых произошли от комбинации дорсальной части грудной клетки и стенки тела (Prokop et al., 2017). Расширенные грудные доли развивали сочленение, позволяющее скользить, и мускулатура, сформированная позже, чтобы приводить в движение взмахи крыльями.
1) (Nelson and Tidwell, 1987). Морфологическое происхождение крыльев остается спорным, поскольку нет прямых доказательств переходных форм между бескрылыми и крылатыми насекомыми. Убедительная теория, основанная на примитивной летописи окаменелостей нимф крылатых насекомых, предполагает, что крылья насекомых произошли от комбинации дорсальной части грудной клетки и стенки тела (Prokop et al., 2017). Расширенные грудные доли развивали сочленение, позволяющее скользить, и мускулатура, сформированная позже, чтобы приводить в движение взмахи крыльями.
Из-за повышенной метаболической потребности в взмахе крыльев во время полета грудные мышцы насекомых имеют самый высокий удельный массовый расход кислорода по сравнению с другими типами тканей (Harrison and Lighton, 1998). Более высокое парциальное давление кислорода (PO 2 ) компенсировало диффузионные пределы в трахеальных системах насекомых во время дыхания, уменьшая ограничения на максимальный размер тела, а атмосферная гипероксия, возможно, сыграла роль в эволюции гигантизма (Dudley, 1998). Все гигантские членистоногие из позднепалеозойской эры вымерли к концу перми, возможно, из-за снижения концентрации кислорода в атмосфере (Dudley, 1998).
Все гигантские членистоногие из позднепалеозойской эры вымерли к концу перми, возможно, из-за снижения концентрации кислорода в атмосфере (Dudley, 1998).
Летающие птицы появились позже летающих насекомых (рис. 1). Ископаемые остатки указывают на то, что птицы произошли от мелких плотоядных динозавров в конце юрской эры ∼150 млн лет назад (Brusatte et al., 2015). Археоптерикс считается переходной формой между нептичьими пернатыми динозаврами и современными птицами (Xu et al., 2011). Примечательно, что второе повышение уровня кислорода в атмосфере произошло во время начала полета птиц (Dudley, 2000). Существуют две гипотезы эволюционного происхождения полета птиц: гипотеза «прямого воздушного спуска» и гипотеза «фундаментального взлета крыла» (Askew and Ellerby, 2007). Основная дискуссия между этими двумя гипотезами заключается в том, произошел ли полет птиц от предков-планеров или взмахов крыльев. Тем не менее, более высокое значение PO 2 в атмосфере предположительно способствовало развитию производства аэродинамической силы и усилению перфузии кислорода в грудных мышцах птиц для обеспечения взмахов крыльев в полете.
Летучие мыши — единственная сохранившаяся линия млекопитающих, обладающая способностью к полету. Возникновение летучих мышей можно проследить до эпохи раннего палеоцена — позднего мелового периода ∼52 млн лет назад (рис. 1), которые находились в гипероксической среде, подобной той, когда возникла птичья стая. Молекулярно-филогенетический анализ продемонстрировал монофилию летучих мышей, предполагая, что у летучих мышей механический полет произошел от единственного предкового источника (Tokita et al., 2012). Широко признано, что летающие млекопитающие произошли от мелких древесных млекопитающих, а не от наземных бегунов, и окаменелость микрокрылых животных Onychonycteris из 52 млн лет назад в эоценовый период подтверждает эту точку зрения (Bishop, 2008). Onychonycteris имел более длинные задние лапы и более короткие предплечья по сравнению с современными летучими мышами, что предполагало примитивные короткие перелеты с дерева на дерево. Эта теория эволюции полета «с дерева вниз» предполагает, что летучие мыши приобрели способность летать, упав с дерева, а не от быстрого бега для взлета с земли (Bishop, 2008). Никакие промежуточные ископаемые свидетельства напрямую не связывают летучих мышей с нелетающими предками, предполагая довольно быстрое появление летающих летучих мышей. Эволюция уникальной мускулатуры крыльев летучих мышей (Cheney et al., 2017) еще предстоит изучить.
Предлагаемый в качестве важного фактора окружающей среды, способствующего развитию полета животных, высокий уровень PO 2 в атмосфере мог бы повысить системный метаболический потенциал и физическую активность животных для достижения потребности в полете. Помимо эффекта атмосферной гипероксии на начало полета, более высокая плотность воздуха в те геологические эпохи могла также способствовать аэродинамическим подъемным эффектам взмахов и аэродинамической поверхности крыльев (Combes and Daniel, 2003).Помимо увеличения мышечной массы для увеличения выходной мощности во время полета, кости птиц были сращены для устранения суставов и уменьшения массы (Foth et al., 2014). Эта адаптация летающих видов сосредоточена на увеличении мышечной силы и запаса энергии, а также на повышении эффективности работы для поддержания высокой стоимости полета.
Независимая эволюция сходящихся черт в линиях трех летающих животных
Способность летать позволяет животным достигать дополнительных источников пищи, преодолевать физические препятствия для жизни в более благоприятных средах обитания и убегать от наземных хищников.Хотя многие виды животных развили способность летать, единственные сохранившиеся группы, которые приобрели механический полет, — это насекомые, птицы и летучие мыши. Эволюция полета животных — один из увлекательных примеров фенотипической конвергенции, в которой отдаленные клоны развили одну и ту же физиологическую функцию, чтобы адаптироваться к аналогичному выбору окружающей среды.
Насекомые были первой группой животных, у которых развились крылья, и их способность летать с ископаемыми останками свидетельствует о том, что они эволюционировали только один раз (Ross, 2017).Общепринятая гипотеза предполагает, что полет насекомых возник на суше, а не на поверхности моря. Окаменелости наиболее примитивных насекомоподобных коллембол Rhyniella praecursor из 407 млн лет назад подтверждают эту гипотезу (Dunlop and Garwood, 2018). Коллембол, как родственная группа насекомых, представляет собой бескрылое шестиногое гексапод. Уникальная структура коллембол, фуркула под их телом, способна поднимать животное в воздух, избегая опасных сред. Способность древних коллембол в воздухе указывать на возможность развития крылатых насекомых на суше (Engel and Grimaldi, 2004).Строение крыльев насекомых произошло от расширенных грудных долей, которые использовались для терморегуляции у предковых крылатых животных. Впоследствии расширенные грудные доли были сочленены для функции скольжения, а мышцы появились, чтобы махать силой при полете (Ross, 2017).
Позвоночные животные позже эволюционировали в отдельные линии. Птицы — один из самых разных классов современных позвоночных из-за их способности летать. Свидетельства ископаемых и исследования молекулярной эволюции последних десятилетий показывают, что птицы произошли от тероподных динозавров юрского периода (Brusatte et al., 2015). Маленькое, легкое, оперенное и крылатое тело в форме тела впервые появилось в мезозойском периоде. Высказывается гипотеза, что истоки полета птиц с двигателем возникли у выживших видов предков теропод после массового вымирания (Brusatte et al., 2015). Наряду с увеличением массы грудных мышц у птиц возникли пневматизация посткраниального скелета (полые, заполненные воздухом кости) и сращения костей, чтобы облегчить вес тела в процессе эволюции полета (Britt et al., 1998).
Летучие мыши — единственное летающее млекопитающее, которое произошло от предка-планера, который лазил по деревьям и поднимался в воздух, чтобы увеличить дальность полета (Bishop, 2008). Летописи окаменелостей свидетельствуют о появлении летучих мышей, полностью способных летать ∼52 млн лет назад. Летучие мыши разделились на два подотряда: Microchiroptera и Megachiroptera . Эти две группы имеют значительные различия в размере, питании и среде обитания. Megachiroptera (мегабаты) имеют гораздо более крупные размеры тела по сравнению с Microchiroptera (микробаты).Микробаты развили способность эхолокации, используя высокочастотные звуки, чтобы обнаруживать мелких жертв, таких как насекомые, в то время как мегабаты являются нектароядными (Dechmann et al., 2006).
Конвергентная эволюция полета у насекомых, птиц и летучих мышей разделяет общую адаптацию движений крыльев с помощью мускулов для создания устойчивой аэродинамической подъемной силы, которая требует энергии. Следовательно, эволюция летных мышц у этих филогенетически дивергентных видов должна отражать адаптации из-за выбора высокоэффективных генераторов энергии.
Анатомические и физиологические особенности летных мышц насекомых, птиц и летучих мышей
Мышцы насекомых и поперечно-полосатые мышцы позвоночных являются аналогичными органами, тогда как летные мышцы птиц и летучих мышей являются гомологичными органами, специально выбранными для полета. Следовательно, сравнение мускулов трех независимо развившихся групп летающих животных может дать представление о том, как естественный отбор управляет эволюцией аналогичных структур и их функций, а также радикальными расхождениями.
Полетные мышцы насекомых
Насекомые — единственные летающие беспозвоночные на Земле, и они развили способность летать с небольшими размерами тела, чтобы максимизировать частоту взмахов крыльев (Conley and Lindstedt, 2002). Мышцы полета насекомых расположены в грудной клетке и функционируют прямым или косвенным образом (Bullard and Pastore, 2011; Baumler et al., 2018) (Рисунок 2) с синхронными или асинхронными сокращениями (Ellington, 1985).
Рисунок 2. Прямые и непрямые мышцы полета насекомых. (слева) Движение крыльев приводится в движение синхронными мышцами прямого полета. Более крупные насекомые, такие как стрекозы и саранча, используют мышцы прямого полета, чтобы взмахивать крыльями во время полета. Сокращение мышц-подъемников тянет крыло вверх, а мышцы-депрессоры тянут крылья вниз. (Справа) Движение крыльев плодовой мушки приводится в движение асинхронными мышцами непрямого полета. Дорсальные вертикальные мышцы (DVM) тянут верхнюю часть грудной клетки, чтобы произвести движение крыльев вверх при растяжении продольных спинных мышц (DLM).Последующее сокращение DLM вызывает укорочение переднего и заднего концов грудной клетки, что приводит к движению вниз крыльев и растяжению DVM, чтобы вызвать следующий цикл, активируемый растяжением. Иллюстрации резюмированы из литературных источников (Iwamoto, 2011).
Синхронные мышцы прямого полета обнаруживаются у низших видов насекомых, таких как саранча ( Orthoptera ) и стрекоза ( Odonata ), у которых взмах крыльев происходит на более низких частотах (<100 Гц). Подобно действиям птичьих грудных мышц, сокращение подъемных мышц в грудной клетке поднимает крылья, а сокращения депрессорных мышц приводят в действие крылья (Baumler et al., 2018) (рисунок 2). Режим сокращения и расслабления мышц синхронного полета насекомых аналогичен режиму скелетных мышц позвоночных, в котором сокращение вызывается деполяризацией мембраны миоцитов в ответ на импульсы, поступающие от двигательных нервов, а цикл сокращения-релаксации зависит от подъема и распад цитозоля [Ca 2+ ] (Ellington, 1985; Iwamoto, 2011).
Большинство современных видов насекомых имеют асинхронные мышцы непрямого полета (IFMs) для обеспечения высокочастотного биения крыльев посредством деформации грудного экзоскелета, а не прямого управления движениями крыльев (Conley and Lindstedt, 2002).При сохранении структуры поперечно-полосатых мышц асинхронные IFM эволюционировали с механизмом активации растяжения. В грудной клетке есть два набора IFM: спинная продольная мышца (DLM) и дорсальная вентральная мышца (DVM). Два набора мышц расположены поперечно и работают вместе, чтобы управлять колебательными деформациями грудной клетки и взмахами крыльев (Conley and Lindstedt, 2002). При взлете нервный импульс заставляет DVM сокращаться, что деформирует грудную клетку, заставляя растяжение DLM вызывать замедленную активацию растяжения.Отсроченное сокращение DLM, в свою очередь, заставляет DVM расширяться и активируется растяжением (Deora et al., 2017) (Рисунок 2). Этот цикл растяжения и сокращения повторяется с высокой частотой с помощью низкочастотных стимулирующих импульсов от двигательных нервов, благодаря которым промежуточное повышение цитозоля [Ca 2+ ] будет поддерживать непрерывные колебания сокращения и расслабления асинхронного IFM насекомых (Iwamoto, 2011 ).
Грудные мышцы птиц
Полет птицы в основном обеспечивается грудными мышцами, которые перемещают плечевую кость крыла вокруг плеча.Грудные мышцы большинства взрослых птиц составляют примерно 8–11% от общей массы тела (George, Berger, 1966; Biewener, 2011). Основная роль этих больших скелетных мышц — создавать механическую силу для движений крыла вниз во время полета (Poore et al., 1997). Еще одна важная летающая мышца птиц — это супракоракоидус, который составляет около одной пятой массы грудной мышцы. Меньшая мышца supracoracoideus в первую очередь отвечает за подъем крыльев при взмахе вверх, что особенно важно для движений с медленной и умеренной скоростью, а также за парение при быстрых движениях крыльев (Crandell and Tobalske, 2015).
Чтобы обеспечить аэродинамическую мощность, достаточную для поддержания полета, летательные мышцы птиц должны постоянно сокращаться на высоких частотах, требуя значительных усилий и больших затрат энергии. Состав волокон летательных мышц птиц различается у разных видов. Грудная мышца содержит в основном быстроокисляющие волокна (~ 85% у голубей) и небольшую часть быстро гликолитических волокон, в то время как у supracoracoideus пропорции быстро гликолитических и быстроокисляющих волокон могут сильно различаться у разных видов (Biewener, 2011).
Мышцы полета летучей мыши
Как и у большинства летающих птиц, у летучих мышей на грудине имеется киль, к которому прикрепляются большие грудные мышцы (Gaudioso et al., 2017). Макроскопические исследования анатомии и электромиографии также выявили многочисленные мышцы в руке и предплечье летучих мышей, которые участвуют в движении крыльев и контролируют форму и ориентацию крыльев во время полета (Hermanson and Altenbach, 1981). Подобно птицам, грудная мышца летучей мыши является основной мышцей, приводящей в движение крылья вниз во время полета, а двуглавая мышца плеча действует, складывая крылья в локтевом суставе во время движения крыльев вниз (Von Busse et al., 2012). Летучие мыши развили крылья для полета от передних конечностей с суставами внутри пальцев. Эти сочленения влияют на трехмерную форму крыла, чтобы контролировать создание аэродинамических сил (Hedenstrom and Johansson, 2015). Поскольку большое количество мышц участвует в управлении аэродинамикой во время полета летучей мыши, затраты энергии, связанные с грудной мышцей, составляют меньшую часть от общей суммы летных мышц, чем у птиц (Cheney et al., 2014).
В отличие от хлопающих крыльев птиц и насекомых, у летучих мышей развились уникальные структуры крыльев, которые больше похожи на патагии летающих животных.Крылья летучей мыши образованы перепончатой кожей, называемой плагиопатагиумом, покрывающей кости передней конечности и соединяющей области между конечностью, пальцами и даже хвостом. Большая часть поверхности крыльев летучей мыши представляет собой кожу, толщина которой варьируется от 27 до 270 мкм в зависимости от вида. Кожа крыла летучей мыши сложна и уникальна и состоит из пучков эластина и встроенных крошечных скелетных мышц, называемых плагиопатагиальными (Skulborstad et al., 2015). Пучки эластина ориентированы преимущественно в направлении размаха, что способствует облегчению складывания и раскладывания крыльев во время полета (Cheney et al., 2017). Исследования 130 видов летучих мышей обнаружили пять различных мышечных массивов в плагиопатагиуме крыльев, которые потенциально контролируют жесткость мембран (Cheney et al., 2017). Благодаря тонкой и эластичной коже мышечные массивы и эластиновые пучки контролируют форму и точно регулируют натяжение перепончатых крыльев, чтобы регулировать аэродинамику во время полета.
Сократимость и механизмы регуляции различных летных мышц
Летные мускулы насекомых, птиц и летучих мышей развивались независимо друг от друга с функциональной конвергенцией.Многие из их анатомических и физиологических сходств отражают адаптацию к обычным селективным давлениям, таким как высокая выходная мощность с энергетической эффективностью для устойчивых полетов. С другой стороны, уникальные свойства летающих мышц насекомых, птиц и летучих мышей, такие как длина саркомера в состоянии покоя, пассивная жесткость и режим сокращений, активируемых Ca 2+ или активируемых растяжением, указывают на адаптацию к определенным селективным давлениям для конкретный тип летной активности (Cutts, 1986; Cao et al., 2019).
Общие черты поперечно-полосатых мышц беспозвоночных и позвоночных
В то время как насекомые, птицы и летучие мыши эволюционировали независимо от разных предков, их летательные мышцы аналогичны поперечно-полосатым мышцам (Iwamoto et al., 2002; Homberger and de Silva, 2015; Cheney et al., 2017). Скелетные мышцы позвоночных и IFM насекомых были широко изучены как представители поперечно-полосатых мышц, которые содержат пучки миофибрилл, состоящих из тандемных повторов саркомеров в качестве сократительной единицы (Iwamoto, 2011; Sweeney and Hammers, 2018).Отражая аналогичные механизмы функции, структурная организация саркомера похожа у летательных мышц насекомых, птиц и летучих мышей с перекрывающимися толстыми филаментами миозина и тонкими филаментами актина и связанными с ними каркасными и регуляторными белками (Рисунок 3) (Cutts, 1986; Cao и др., 2019). В саркомере толстые филаменты миозина закрепляются на М-линии, тонкие филаменты актина закрепляются на Z-диске, а тайтин или тайтин-подобные эластичные белки соединяют толстые и тонкие филаменты, образуя структурную целостность (Рис. 3).Исходя из природы поперечно-полосатых мышц как высокодифференцированного типа ткани, сократительный аппарат и регуляторные механизмы сокращения и расслабления летных мышц насекомых, птиц и летучих мышей в значительной степени сохранены от их общего предка, который жил 500-700 млн лет назад (Cao et al. др., 2019).
Рисунок 3. Сравнение структур саркомеров (A) позвоночных и (B) летательных мышц беспозвоночных. Летные мышцы беспозвоночных и позвоночных содержат сходные саркомерные структуры, состоящие из перекрывающихся миозиновых толстых филаментов и актиновых тонких филаментов с гомологичными, но расходящимися каркасными белками.Мышцы непрямого полета (IFM) беспозвоночных имеют более длинные саркомеры по сравнению с летательными мышцами позвоночных и более узкие I-образные перемычки по сравнению с саркомерами позвоночных. Иллюстрации были обобщены из литературной информации (Hooper et al., 2008; Lange et al., 2020).
Содержание белка миофиламентов как в летных мышцах беспозвоночных, так и в позвоночных сохраняется, как и в других поперечнополосатых мышцах (Рис. 3). Все они содержат активируемые F-актином миозиновые двигатели и связанные с тонкими филаментами регуляторные белки тропомиозин и тропонин (Tn).Комплекс Tn состоит из трех белковых субъединиц: Ca 2+ -связывающей субъединицы тропонина C (TnC), ингибирующей субъединицы тропонина I (TnI) и заякоренной субъединицы тонкого филамента тропонина T (TnT) (Gordon et al. , 2000; Jin et al., 2008). Когда мышца сокращается, деполяризация плазматической мембраны миоцитов вызывает приток небольшого количества Ca 2+ и запускает высвобождение более значительного количества Ca 2+ из саркоплазматической сети (SR). Увеличение цитозольного [Ca 2+ ] способствует связыванию Ca 2+ с TnC и вызывает серию конформационных изменений Tn и тропомиозина, позволяя миозиновым головкам образовывать прочные поперечные мостики с тонким актиновым филаментом для активации миозин-АТФаза и генерируют силовые удары.Расслабление мышц происходит, когда свободный Ca 2+ уменьшается под действием SR и насосов Ca 2+ плазматической мембраны, которые удаляют Ca 2+ из цитозоля, что приводит к диссоциации Ca 2+ из TnC (Gordon et al. , 2000; Ивамото, 2011).
Основываясь на консервативных субклеточных структурах, биохимической основе и функциях, уникальные особенности сократительных и регуляторных механизмов летающих мышц насекомых, птиц и летучих мышей могут дать представление об эволюции их сократительной способности и энергетической эффективности.
Мышца синхронного полета насекомых
Тот факт, что летательные мышцы низших насекомых, таких как саранча и стрекозы, с частотой взмахов крыльев ниже 100 Гц, имеют синхронные летательные мышцы, которые функционируют как нелетные мышцы насекомых и поперечно-полосатые мышцы позвоночных (Snelling et al., 2012). эта синхронная мышца прямого полета изначально присутствовала, и асинхронные IFM, возможно, произошли от них, чтобы поддерживать более частые движения крыльев и потенциально с более высокой энергетической эффективностью.
Сокращение и расслабление мышц синхронного полета насекомых происходит посредством того же механизма, что и у скелетных мышц позвоночных, включая синхронное соединение возбуждения-сокращения (Peron et al., 2009), как описано ранее. Сокращение инициируется стимулированными двигательными нейронами возбуждениями в нервно-мышечных соединениях, что приводит к деполяризации плазматической мембраны миоцитов и притоку Ca 2 + , который вызывает высвобождение большего количества Ca 2+ из SR.Поднимающийся свободный Ca 2+ связывается с TnC, что вызывает конформационные изменения комплекса Tn и тропомиозина в тонких филаментах актина, что приводит к активации миозиновой АТФазы. Гидролиз АТФ вызывает энергетический удар миозиновой головки, укорачивая саркомер и сокращая мышцу. Во время релаксации цитозольный Ca 2+ перекачивается обратно в SR и внеклеточное пространство. С помощью этого механизма скорость циклирования Ca 2+ ограничивается скоростью диффузии Ca 2+ и насосов SR Ca 2+ , ограничивая частоту ударов крыльями насекомых с синхронными летательными мышцами до <100 Гц ( Ивамото, 2011).В соответствии с законсервированным механизмом сокращения и расслабления, длина саркомера покоя синхронных летных мышц саранчи составляет ~ 3.5 мкм, как и у скелетных мышц позвоночных (Nave and Weber, 1990).
Асинхронный IFM для насекомых и активация растяжения
Представляя более совершенную адаптацию к полету, асинхронный IFM мелких насекомых эволюционировал с уникальной активацией растяжения для высокочастотных сокращений. Для генерации высокочастотных импульсов крыльев низкочастотные нейронные импульсы активируют асинхронный IFM, чтобы вызвать повышение цитозольного Ca 2+ до промежуточного уровня, который не колеблется во время последующего сокращения и расслабления.Вместо зависящих от волны циклов активации-деактивации миофиламентов Ca 2+ , сокращение и расслабление асинхронных IFM насекомых используют механизм активации взаимного растяжения двух пар анатомически перекрестно ориентированных грудных мышц, из которых сокращение одной группы растягивает другой набор, чтобы активировать последующее сокращение, и цикл повторяется, приводя к автономным колебаниям (Squire, 1992) (Рисунок 2). Этот механизм позволяет избежать ограничений скорости, связанных с равновесной динамикой переходного процесса Ca 2+ и циклической смены Ca 2+ , управляемой насосом, обеспечивая более быстрое биение крыльев без дорогостоящих работ при обращении с Ca 2+ (Dickinson et al., 2005).
IFM насекомых также представляют собой поперечно-полосатые мышцы, подобные скелетным мышцам позвоночных, с саркомерной структурой и консервативными белками миофиламентов, миозином, актином, тропонином и белками эластичного каркаса (Iwamoto et al., 2002) (Figure 3). Длина покоящегося саркомера IFM насекомых колеблется от 3,7 мкм у Drosophila (Bullard et al., 2002) до 2,7–2,9 мкм у пчелы (Nave and Weber, 1990). В отличие от поперечно-полосатых мышц позвоночных, высокая пассивная жесткость является характерной чертой асинхронных IFM насекомых вместе с узкими I-полосами, длина которых не изменяется во время сокращения и расслабления (Hao et al., 2006; Хендерсон и др., 2017). Очевидно, жесткая саркомерная структура, вероятно, является адаптацией миофиламентов для восприятия сигнала растяжения во время активации растяжения.
Моторные функции саркомерных миофиламентов в мышцах позвоночных и насекомых требуют, чтобы миозин в толстом филаменте взаимодействовал с актином в тонком филаменте в соответствии с Ca 2+ -зависимой регуляцией Tn (Gordon et al., 2000; Sweeney and Hammers, 2018 ; Cao et al., 2019). Все больше данных свидетельствует о том, что конформационные искажения как миозина в толстом филаменте, так и регуляторных белков в тонком филаменте играют роль в активации растяжения асинхронных IFM (Perz-Edwards et al., 2011; Ивамото и Яги, 2013). Исследования на Drosophila IFM продемонстрировали, что домен конвертера миозина является критическим для генерации достаточной аэродинамической мощности для полета (Swank et al., 2002). Нарушение домена конвертера миозина снижает напряжение, активируемое растяжением, в первую очередь за счет уменьшения пассивной жесткости мышечных волокон (Wang et al., 2014). N -концевой домен регуляторной легкой цепи миозина играет структурную роль в определении количества прочно связанных поперечных мостиков миозина и генерации энергии (Miller et al., 2011). Исследование на Drosophila показало, что альтернативно сплайсированный N -конечный регион тяжелой цепи миозина является регуляторным сайтом для оптимизации выработки энергии в IFM (Swank et al., 2004). Последовательные исследования рентгеновской дифракции в реальном времени выявили Tn-мостики, которые механически оттягивают тропомиозин в сторону, когда растягивается волокно от Lethocerus IMF (Perz-Edwards et al., 2011). Было обнаружено, что различные альтернативные формы сплайсинга TnT изменяют чувствительность Ca 2+ и выходную мощность IFM насекомых (Marden et al., 1999). Хотя эти молекулярные механизмы могут совместно способствовать активации растяжения IFM насекомых, стоит проверить гипотезу, что сигнал напряжения механического растяжения саркомерных миофиламентов может изменять конформацию тонких волокон и повышать чувствительность тропонина к постоянному промежуточному уровню цитозольного Ca 2+. , который активирует актомиозин-АТФазу и перекрестный мостиковый цикл.
Птичьи летные мышцы
По сравнению с другими летающими видами, летающие птицы имеют массивные грудные и надкожные мышцы, обеспечивающие движение крыльев вниз и вверх во время полета.Грудные мышцы птиц содержат преимущественно быстро сокращающиеся волокна и были изучены как классические скелетные мышцы позвоночных в отношении их типичных миофибрилл и саркомерных структур, а также содержания белка миофиламентов (Cutts, 1986). Детерминирующий механизм (механизмы) эволюционировал уникальным образом в грудной мышце птиц в процессе адаптации к длительному полету, например, во время непрерывной миграции на большие расстояния, еще предстоит установить. Парение — это уникальный способ полета, при котором птицы сохраняют в воздухе почти неподвижное положение (Vejdani et al., 2018). Колибри, самые маленькие виды птиц, — единственные птицы, которые могут выдерживать парение. Их небольшой размер тела и пропорционально большие грудные мышцы позволяют им выдерживать высоту и парение (Achache et al., 2017). По сравнению с другими птицами, у колибри значительно более высокая частота взмахов крыльев (~ 34 Гц) при гораздо меньшей силе и напряжении, создаваемых грудными мышцами (Tobalske, 2016). Продолжительность нервного импульса во время активации грудных мышц колибри короче, чем у других птиц, что соответствует более короткому времени взаимодействия возбуждения и сокращения во время высокочастотных взмахов крыльев и предположительно меньшему притоку Ca 2+ в миоциты с более низкой активацией миозина. моторы (Тобальское, 2016).
Птичьи летящие мышцы имеют типичные саркомерные структуры (рис. 3В). Длина покоящегося саркомера составляет 2,08–2,14 мкм у грудной мышцы курицы (McGinnis et al., 1989; Burkholder, Lieber, 2001), ~ 1,89 мкм в грудной мышце колибри и ~ 2,5 мкм в мышце ноги колибри. Напротив, саркомеры скелетных мышц млекопитающих длиннее, ∼2,3 мкм в медленных и быстрых скелетных мышцах крыс, 2,27 мкм у кролика и 2,64 мкм у человека (Burkholder and Lieber, 2001; Reiser et al., 2013). Более длинный саркомер в состоянии покоя будет ближе к оптимальной длине для создания максимальной силы (Reiser et al., 2013). Следовательно, функциональное значение компромисса более короткой длины саркомера в грудных мышцах птиц еще предстоит исследовать. Несмотря на различную сократительную потребность летных мышц у разных видов, изоформа тяжелой цепи миозина (MHC) удивительно однородна в грудных мышцах большинства видов птиц с очень ограниченными вариациями (Velten and WelchJr., 2014; Tobalske, 2016). Поскольку один и тот же миозиновый двигатель способен удовлетворить общие требования к различным типам силового полета, различные сократительные и функциональные характеристики летных мышц разных видов птиц могут зависеть от адаптации и вариаций в регуляторных белках тонких филаментов.
N -концевая область TnT имеет вариабельную структуру из-за генетических вариаций, альтернативного сплайсинга и протеолитической модификации, которая производит конформационные модуляции дальнего действия для изменения функции Tn и мышечной сократимости (Wang and Jin, 1998; Wei and Джин, 2016). Ген быстрого TnT птиц эволюционировал с помощью до семи уникальных экзонов P, кодирующих повторяющиеся мотивы последовательностей, богатых His-Glu, в N -концевой вариабельной области (Jin and Samanez, 2001).В дополнение к альтернативному сплайсингу для исключения эмбриональных экзонов для уменьшения размера и отрицательного заряда N -концевой области, наблюдаемого в регуляции развития как генов изоформ TnT в скелетных мышцах, так и сердечных генов у позвоночных (Wang and Jin, 1997; Jin) et al., 2008), быстрый TnT грудной мышцы птицы подвергается параллельному пост-штриховому альтернативному сплайсингу — экзонам P для увеличения размера и отрицательного заряда вариабельной области N -конца (Ogut et al., 1999). Этот фенотип не наблюдается в мышцах ног и сопровождается быстрым ростом массы грудных мышц (Jin and Samanez, 2001). Возрастное и функциональное значение этой интригующей адаптации, специфичной для летных мышц, еще предстоит установить.
Грудные и крылатые мышцы летучих мышей
Лётные мышцы летучей мыши имеют консервативную структуру, сократительные и регуляторные механизмы, аналогичные таковым у других поперечнополосатых мышц млекопитающих. Длина саркомера покоя грудной мышцы летучей мыши составляет ∼2.2 мкм и аналогичен скелетным мышцам крысы (Mathieu-Costello et al., 1992). Особенностью полета летучих мышей является то, что набор мышечных волокон грудной мышцы регулируется скоростью полета, аналогичной скорости полета птиц (Konow et al., 2017). Производство энергии связано с изменениями скорости полета (Hermanson et al., 1998). Различная скорость полета разных видов летучих мышей может быть связана с различным составом волокон. Грудная мышца летучих мышей, питающихся насекомыми, содержит только неутомляемые быстрые окислительно-гликолитические волокна, тогда как грудная мышца летучих мышей, питающихся фруктами, содержит несколько типов волокон (Hermanson et al., 1998; Konow et al., 2017).
Крылья летучей мыши могут изменять свою морфологию в различных аэродинамических условиях с помощью плагиопатагиальных мышц. Эти мышцы берут начало в скелете и вставлены в мембрану плагиопатагиума. Параллельно хорде расположены внутримембранные мышцы, не связанные с костью. Сокращения этих мышц создают напряжение в мембране плагиопатагиума и задних краях синхронно, чтобы сформировать крылья летучей мыши для противодействия аэродинамической нагрузке во время циклов взмахов крыльев для устойчивого полета (Cheney et al., 2014).
Очевидное отсутствие специфических для полета адаптаций грудных мышц летучих мышей может быть причиной того, что летучим мышам необходимо исчерпывающе использовать мышечную силу и метаболизм всего тела для поддержания полета (O’Shea et al., 2014). Этот механизм отличается от эволюции полета насекомых и птиц и подвергает летучих мышей постоянному метаболическому стрессу, который создает уникальное иммунологическое состояние, которое служит резервуаром для различных вирусов, которые являются патогенами человека и других млекопитающих (O’Shea et al., 2014). Это понятие подразумевает, что эволюция и адаптация летных мышц может иметь системное влияние на жизнь животных, при этом интенсивные упражнения и использование скелетных мышц обеспечивает потенциальный механизм для выработки иммунотолерантности (Calder and Yaqoob, 1999) для лечения воспалительного заболевания. в людях.
Энергетическая эффективность летных мышц насекомых, птиц и летучих мышей
Передвижение, приводимое в движение мышцами, является основным расходом энергии для жизни животных, и для снижения затрат на сокращение мышц были разработаны различные стратегии.Помимо увеличения массы летных мышц и улучшения структуры крыльев и снижения веса тела для уменьшения мышечной работы, для летающих видов важна энергетическая эффективность сокращения и расслабления летных мышц для поддержания функции высокой стоимости полета с двигателем (Ellington, 1985; Ward et al. др., 2001; Hedenstrom, Johansson, 2015). Энергоэффективность мышц можно описать как отношение механической мощности к расходу энергии, которое показывает долю химической энергии, которая преобразуется в механическую работу (Morris et al., 2010). Независимо развившиеся летные мускулы насекомых, птиц и летучих мышей приобрели различные особенности сократительной кинетики и снабжения АТФ для сбалансированного расхода энергии.
Насекомые
У летающих насекомых самый высокий удельный аэробный метаболизм среди всех животных (Snelling et al., 2012). Терморегуляция используется медоносными пчелами для оптимизации затрат на электроэнергию при различных температурах окружающей среды. При поисках пищи в тени они демонстрируют высокий уровень оборота энергии для ускорения процесса кормодобывания, который снижается на 18–76% при солнечном нагреве (Kovac et al., 2010; Stabentheiner and Kovac, 2014).
Исследования измеряли скорость метаболизма во время максимального полета по общему количеству выброшенного CO 2 (Mattila, 2015) или потребления O 2 во время полета на привязи (Snelling et al., 2012). Эффективность мышц увеличивается с увеличением силы, тогда как аэродинамическая эффективность подъемной силы снижается с увеличением силы. Уменьшение размера крыльев насекомых связано с уменьшением летной мышечной массы, выработки механической энергии и общей эффективности полета.Следовательно, насекомые менее эффективны в преобразовании метаболической энергии в механическую, а общая энергетическая эффективность полета Drosophila составляет всего 2–3% из-за высокой скорости метаболизма всего тела (Lehmann, 2001). Основным источником энергии в мышцах насекомых является жир, а жировые клетки тела контролируют использование энергии из жира и гликогена (Arrese and Soulages, 2010).
Ca 2+ , циркулирующий через плазматические и SR мембраны во время сокращения и расслабления поперечно-полосатых мышц позвоночных, и синхронный IFM насекомых потребляет большое количество энергии, используя ~ 30% общего АТФ, продуцируемого в мышечных клетках (Szentesi et al., 2001). Напротив, асинхронный IFM насекомых работает без биения, чтобы побить Ca 2+ на велосипеде, демонстрируя уникальный режим энергосбережения для создания работы большой мощности для полета (Conley and Lindstedt, 2002).
Цикл Ca 2+ во время сокращения и расслабления мышц с помощью SR и насосов Ca 2+ плазматической мембраны требует значительных затрат энергии и потребляет большое количество АТФ. Исследования показали, что кардиомиоциты млекопитающих используют ~ 30% общего АТФ, вырабатываемого для работы насосов Ca 2+ (Starling et al., 1998). Исследования скелетных мышц человека показали, что активность SR-АТФазы потребляет такое же количество общего АТФ, что и миозин-АТФаза в медленных мышцах типа I и примерно половину этого количества в быстро сокращающихся волокнах типа IIA (Szentesi et al., 2001). Устраняя необходимость в высокочастотном циклировании больших количеств Ca 2+ во время каждого эпизода сокращения и релаксации, количество SR и мембранных насосов Ca 2+ сводится к минимуму в асинхронном IFM насекомых, что приводит к меньшему потреблению АТФ по сравнению с который используется миозиновыми двигателями во время силового полета (Conley and Lindstedt, 2002).Следовательно, снижение затрат на обработку Ca 2+ в миоцитах является потенциальным направлением повышения энергетической эффективности поперечно-полосатых мышц позвоночных.
Регуляторные белки миофиламентов, особенно Tn, играют центральную роль в регулировании кинетики производства силы и, следовательно, вносят вклад в энергетическую эффективность поперечно-полосатых мышц. TnC представляет собой субъединицу рецептора Ca 2+ Tn. Во время активации мышц количество внутриклеточного Ca 2+ напрямую определяет динамическую активацию TnC и производство силы.Мышцы насекомых имеют две изоформы TnC. Форма F1 повсеместно экспрессируется во всех мышцах и имеет только один сайт связывания Ca 2+ , как в сердечном / медленном TnC позвоночных, тогда как форма F2 является IFM-специфичной и имеет два сайта связывания Ca 2+ , как в быстрые TnC позвоночных (Herranz et al., 2005). F2 TnC важен для активации растяжения IFM, и чем выше доля F2 TnC, тем выше частота, которую может достигать IFM (Herranz et al., 2005).
TnT в мышцах насекомых имеет длинное богатое глутаминовой кислотой С-концевое удлинение (Рис. 4), которое отсутствует в TnT позвоночных.Хотя он не является частью консервативной основной структуры TnT, делеция Glu-богатого С-концевого удлинения Drosophila TnT значительно снижает мышечные и сердечные функции (Cao et al., 2020). В качестве потенциально аналогичной структуры N -концевая вариабельная область TnT позвоночных, особенно быстрый TnT в грудных мышцах взрослых птиц с множественными встроенными экзонами P (Ogut et al., 1999; Jin and Samanez, 2001), является также богат Glu и связывает Ca 2+ с физиологически значимым сродством (Zhang et al., 2004), предполагая функцию связанного с миофиламентом Ca 2+ буфера / резервуара. Подтверждая аналогичную функцию, точечный анализ аминокислотных последовательностей выявил значительное сходство между богатым Glu C-концевым удлинением TnT пчелы и богатым гистидином кальций-связывающим белком (HRC) SR (рис. 5), который представляет собой известный Ca . 2+ буфер / резервуар в мышечных клетках позвоночных (Arvanitis et al., 2011). Гипотеза, подлежащая проверке, заключается в том, что этот буфер / резервуар для Ca 2+ обеспечивает механизм для локального циклирования Ca 2+ с TnC во время энергосберегающих циклов активации-деактивации растяжения, когда TnC претерпевает конформационно-модулированные изменения в Ca . 2+ -аффинность связывания.
Рис. 4. Glu-богатое С-концевое удлинение TnT насекомых. Аминокислотные последовательности, кодируемые экзоном 11 или экзоном 12 репрезентативных генов TnT насекомых, выравнивают, чтобы идентифицировать сходство остатков и проиллюстрировать специфичное для насекомых С-концевое удлинение, обогащенное остатками Glu. Содержание Glu положительно коррелирует с частотой взмахов крыльев этих видов (Snelling et al., 2012). Регистрационные номера используемых последовательностей в GenBank: Dragonfly TnT, AAD33604.1; саранча TnT, AVCP010016941; Drosophila TnT, NP_525088; Пчела ТнТ, NP_001035348.1.
Рис. 5. Парные точечные диаграммы сходства аминокислотных последовательностей между Glu-богатым сегментом TnT насекомых и богатым гистидином кальций-связывающим белком саркоплазматического ретикулума (HRC). При анализе с использованием метода попарного выравнивания программного обеспечения DNAStar MegAlign Dotplot C-концевое удлинение TnT пчелы с высокой степенью сходства совпадает с сегментами в человеческом (A) и мышином (B) HRC (обозначено пунктирными кружками), что указывает на аналогичный функционируют как резервуар миофиламента Ca 2+ .Соответствующие области и степень сходства последовательностей обозначены разными цветами диагональных линий. Красный цвет, показанный на цветной полосе под панелями, представляет наибольшее сходство, а фиолетовый — наименьшее. Номера доступа GenBank для используемых последовательностей: HRC-связывающий белок человека, AAH94691.1; белок сердца HRC мыши, AAD42061.1; Bee TnT, PBC29029.1.
Птицы
Измерение кинематики взмахов крыльев как механической мощности и метаболической мощности (потребления кислорода) показало, что эффективность летных мышц птиц у мелких птиц составляет от 13% до 23% (Ward et al., 2001; Аскью и Эллерби, 2007). Грудные мышцы птиц приводят в действие взмахи крыльев со значительной долей укорочения (33–42%) во время полета (Biewener, 2011). На способность птиц летать или удерживать положение в воздухе влияет размер тела, при этом боевая эффективность снижается по мере увеличения размера тела. Объяснение заключается в том, что скорость выработки энергии не успевает за увеличением размера тела, таким образом, удельная выработка энергии снижается, хотя это частично компенсируется увеличением количества гликолитических волокон, производящих более высокую мощность (Askew et al., 2001). Гипотеза «сердце для полета» предполагает, что эффективная сердечно-сосудистая система с большим сердцем необходима для доставки кислорода к грудным мышцам для полета с двигателем (Altimiras et al., 2017). В соответствии с этой гипотезой, парящий полет развился только у колибри с маленьким размером тела, который производит наиболее энергетически затратную мышечную работу с самой высокой удельной массой метаболизма у позвоночных (Evangelista et al., 2010).
В отличие от насекомых и млекопитающих, летающие птицы демонстрируют высокий дневной обмен энергии и более продолжительную продолжительность жизни по сравнению с млекопитающими с такой же массой тела.Из-за хронически повышенного уровня глюкозы в крови и инсулинорезистентности птичьи летные мускулы преимущественно используют окисление липидов для обеспечения движения, которое дает больше энергии, чем метаболизм углеводов. Чтобы предотвратить образование свободных радикалов при высокой скорости метаболизма, митохондрии грудной мышцы воробья обладают гораздо большей способностью окислять жирные кислоты, чем митохондрии мышц задних конечностей крысы. Кроме того, летучая мышца птиц имеет высокое соотношение цепи переноса электронов / оксидазы, что указывает на низкий окислительно-восстановительный потенциал для производства активных форм кислорода (Kuzmiak et al., 2012).
Птицы-мигранты могут совершать беспосадочные перелеты на дальние расстояния без еды и воды. Скорость обмена веществ у перелетных птиц регулируется в режиме реального времени в зависимости от их миграционных потребностей. В основном они используют липиды и минимальное количество белка в качестве топлива во время полета (Jenni-Eiermann, 2017). Во время остановок первой реакцией на снижение скорости метаболизма является увеличение содержания свободных жирных кислот в плазме, чтобы остановить дальнейший липолиз. Показывая конвергентную эволюцию мышечного метаболизма в процессе адаптации к мигрирующим полетам, перелетные летучие мыши используют аналогичную стратегию для адаптации типов топлива к топливу перелетных птиц (таблица 1) (McNab and O’Donnell, 2018), хотя летучие мыши также полагаются на оцепенение между полетами. для экономии энергии (Knight, 2019).
Таблица 1. Сократительные характеристики, метаболическое обеспечение, другие энергетические затраты и энергетическая эффективность летных мышц насекомых, птиц и летучих мышей.
Во время эмбрионального и постнатального развития позвоночных животных белки миофиламентов в сердечных и скелетных мышцах подвергаются переключениям изоформ и / или сплайс-форм в ответ на изменения функциональной потребности. Альтернативный сплайсинг вызывает постнатальное переключение форм сплайсинга TnT быстрых скелетных мышц на сплайсинг экзонов плода в N -концевой вариабельной области (Wang and Jin, 1997; Ogut and Jin, 1998).В дополнение к тому, что у других видов позвоночных и в мышцах ног птиц, быстрый TnT грудных мышц птиц имеет уникальный параллельный переключатель развития после вылупления, чтобы встраивать несколько специфичных для птиц экзонов P, кодирующих длинный богатый Glu сегмент для значительного увеличения длины. концевой вариабельной области N (Ogut, Jin, 1998). Эта быстрая TnT-специфическая структура грудных мышц взрослых птиц не экспрессируется в мышцах ног птиц (Ogut and Jin, 1998) и отсутствует в быстрых TnT нелетающих эму (Cao et al., 2020). Однако он демонстрирует поразительное сходство последовательностей с богатым Glu C-концевым удлинением TnT насекомых, предполагая аналогичную функцию из конвергентного эволюционного отбора для полета (Cao et al., 2020). Гипотеза о том, что Ca 2+ -связывающая способность N -концевого сегмента быстрого TnT грудной мышцы птицы может служить в качестве буфера / резервуара Ca 2+ для локального циклирования Ca 2+ с TnC и Снижение работы и затрат энергии циклических насосов Ca 2+ подтверждается открытием, что N -конечный Glu-богатый сегмент TnT грудной мышцы птицы связывает Ca 2+ с физиологическим сродством (∼14 мкМ) ( Zhang et al., 2004).
Летучие мыши
Подобно птицам, летучие мыши обладают высокой скоростью обмена веществ и относительно небольшими размерами тела, но при этом обладают исключительной продолжительностью жизни. В отличие от птиц, летучие мыши не используют триацилглицерин в качестве окислительного топлива из-за отсутствия у млекопитающих эффективных транспортных ферментов (McGuire et al., 2013). Следовательно, летучие мыши напрямую и быстро усваивают полученные питательные вещества, что является возможной причиной их неспособности выдерживать дальние перелеты, как птицы. Метаболизм диетического сахара является прямым источником энергии для летучих мышей-нектароядных.Напротив, летучие мыши, питающиеся насекомыми, используют богатую белком диету в качестве основного источника энергии с быстрым окислением, чтобы обеспечить высокую потребность в энергии во время полета (таблица 1) (Voigt et al., 2017).
Высокая скорость метаболизма летучих мышей важна для их летных способностей, но также приводит к системному накоплению вредных свободных радикалов. Геномные исследования показали, что у летучих мышей эволюционировали генетические варианты, чтобы противостоять токсичности свободных радикалов, образующихся во время полета (Jebb et al., 2018). Было замечено, что чрезвычайно высокая скорость метаболизма летучих мышей во время полетов может вызывать постоянный стресс, стимулирующий иммунную систему.Генетические варианты, которые минимизируют повреждение свободными радикалами, также могут способствовать уникальному иммунному гомеостазу летучих мышей, что позволяет им быть естественным резервуаром многих вирусов, которые являются крайне патогенными для других млекопитающих (Wang and Anderson, 2019).
Предлагаемый механизм повышения энергоэффективности летучих мышей — это компромисс с маневренностью. Эксперименты в аэродинамической трубе показали, что, хотя летучие мыши менее энергоэффективны в крейсерском полете, чем птицы, они имеют преимущество перед птицами, когда дело доходит до маневрирования (таблица 1).В то время как птицы уменьшили вес крыльев из-за слияния костей, крылья летучих мышей имеют суставы для трехмерного хлопания, чтобы увеличить производство аэродинамической силы (Hedenstrom and Johansson, 2015).
Чтобы исследовать, развились ли аналогичные богатые Glu сегменты в грудных мышцах насекомых и птиц, TnT (Cao et al., 2020) также конвергентно возникли в мышцах полета летучих мышей в качестве потенциального механизма для снижения расхода энергии, анализ точечной диаграммы быстрых мышечных мышей TnT не обнаружил значительного структурного сходства с N -концевой Glu-богатой областью быстрого TnT грудной мышцы птицы, тогда как его общая структура консервативна, аналогична структуре быстрого TnT мышцы ноги птицы и TnT быстрого скелетного мускула мыши (Рисунок 6). .
Рисунок 6. Парные точечные графики сходства аминокислотных последовательностей между TnT из быстро сокращающихся скелетных мышц летучих мышей, грудных мышц и мышц ног птиц и быстро сокращающихся скелетных мышц мыши. При анализе с использованием метода Dotplot программного обеспечения DNAStar MegAlign, TnT летучей мыши не имеет Glu-богатого N -концевого сегмента, обнаруженного в грудном TnT птиц ( A , обозначен пунктирным кругом), тогда как он имеет общую структуру, аналогичную структуре мышцы ноги птицы. TnT (B) и быстрый TnT (C) .Красный цвет, показанный на цветной полосе под панелями, представляет наибольшее сходство, а фиолетовый — наименьшее. Номера доступа GenBank для используемых последовательностей: Быстрая скелетная мышца летучей мыши TnT, XP_028371248.1; птичий грудной быстрый скелет TnT, AAG44258.1; мышца ноги птицы TnT, BAC76600.1; быстрые скелетные мышцы мыши TnT, AAB67285.1.
Биомедицинские перспективы
Путем сравнения летных мышц насекомых, птиц и летучих мышей на предмет их эволюции сократительных свойств и механизмов повышения энергетической эффективности, данные, обобщенные в этом целевом обзоре, показывают, что, хотя эти филогенетически разные животные независимо развили летные способности в разное время от разных предков. естественный отбор дал аналогичные функции полета.Хотя остается много гипотез и спорных точек зрения на происхождение полета, существующие данные дают информативную информацию о понимании мышечной сократимости и энергетической эффективности. Чтобы достичь и поддерживать полет, всем летающим видам необходимо сбалансировать летные характеристики и экономию энергии, что также важно для других мышц, особенно когда рассматривается лечение мышечной слабости и сердечной недостаточности.
Лучшее понимание молекулярной эволюции белков миофиламентов в контексте физиологических особенностей летательных мышц беспозвоночных и позвоночных может помочь изучить потенциальные механизмы и трансляционные приложения для улучшения здоровья человека.Одним из любопытных моментов является то, что лежащие в основе молекулярные механизмы, позволяющие мышцам полета птиц и летучих мышей функционировать с очень разными скоростями метаболизма, отражают разную энергетическую эффективность, хотя у них схожий сократительный механизм и при выборе одинаковых природных условий.
Еще одним привлекательным направлением для будущих исследований является то, что низкая сократительная эффективность асинхронного IFM насекомых поддерживается за счет минимизации работы и затрат энергии системы обработки Ca 2+ , которая является основным потребителем АТФ в поперечно-полосатых мышцах позвоночных (Starling et al., 1998; Szentesi et al., 2001). Прошлые и недавние исследования по лечению мышечной слабости и сердечной недостаточности в основном были сосредоточены на увеличении сократительной способности и / или увеличении выработки АТФ в миоцитах, которые имеют ограниченный потенциал и часто снижают энергетическую эффективность, ограничивая долгосрочную пользу. Таким образом, выборочное снижение расхода энергии на обработку Ca 2+ по сравнению с миозиновыми моторами миофиламентов в поперечно-полосатых мышцах млекопитающих является привлекательным направлением для повышения энергетической эффективности для лечения мышечной слабости и сердечной недостаточности.
Авторские взносы
Оба автора внесли свой вклад в обзор литературы, анализ данных, составление рисунков и таблиц, черновик и редакцию рукописи.
Финансирование
Эта работа была частично поддержана грантами Национальных институтов здравоохранения HL138007 и 127691, предоставленных J-PJ.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Ачаче, Ю., Сапир, Н., Элимелех, Ю. (2017). Аэродинамика крыла парящего колибри в течение годового цикла. I. Полное крыло. R. Soc. Open Sci. 4: 170183. DOI: 10.1098 / RSOS.170183
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Альтимирас, Дж., Линдгрен, И., Хиральдо-Дек, Л. М., Маттей, А., и Гаритано-Завала, Á (2017). Аэробная производительность у тинамусов ограничена их маленьким сердцем. Новая гипотеза эволюции полета птиц. Sci. Отчет 7: 15964.
Google Scholar
Аррезе, Э. Л., и Сулаж, Дж. Л. (2010). Жировое тело насекомых: энергия, метаболизм и регуляция. Annu. Преподобный Энтомол. 55, 207–225. DOI: 10.1007 / 978-1-4615-9221-1_8
CrossRef Полный текст | Google Scholar
Арванитис Д. А., Вафиадаки Э., Сануду Д. и Краниас Э. Г. (2011). Богатый гистидином кальций-связывающий белок: новый регулятор кальциевого цикла саркоплазматического ретикулума. Дж.Мол. Cell Cardiol. 50, 43–49. DOI: 10.1016 / j.yjmcc.2010.08.021
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Аскью, Г. Н., Марш, Р. Л., и Эллингтон, К. П. (2001). Выходная механическая мощность летных мышц голубогрудого перепела ( Coturnix chinensis ) во время взлета. J. Exp. Биол. 204: 3601.
Google Scholar
Баумлер Ф., Горб С. Н., Буссе С. (2018). Сравнительная морфология мускулатуры грудной клетки взрослых Anisoptera (Insecta: Odonata): функциональные аспекты летательного аппарата. Членистоногие. Struct. Dev. 47, 430–441. DOI: 10.1016 / j.asd.2018.04.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бритт Б. Б., Маковицки П. Дж., Готье Дж. И Бонд Н. (1998). Посткраниальная пневматизация археоптерикса. Природа 395, 374–376. DOI: 10.1038 / 26469
CrossRef Полный текст | Google Scholar
Брусатте, С. Л., О’Коннор, Дж. К., и Джарвис, Э. Д. (2015). Происхождение и разнообразие птиц. Curr. Биол. 25, R888 – R898.
Google Scholar
Буллард Б., Линке У. А. и Леонард К. (2002). Разновидности эластичного белка в мышцах беспозвоночных. J. Muscle Res. Cell Motil. 23, 435–447. DOI: 10.1007 / 978-94-010-0147-2_5
CrossRef Полный текст | Google Scholar
Буркхолдер, Т. Дж., И Либер, Р. Л. (2001). Длина саркомера, рабочий диапазон мышц позвоночных во время движения. J. Exp. Биол. 204 (Pt 9), 1529–1536.
Google Scholar
Колдер П. К. и Якуб П. (1999). Глютамин и иммунная система. Аминокислоты 17, 227–241.
Google Scholar
Цао, Т., Суйковски, А., Кобб, Т., Уэсселс, Р. Дж., И Джин, Дж. П. (2020). Обогащенное глутаминовой кислотой длинное С-концевое удлинение тропонина Т играет решающую роль в функциях мышц насекомых. J. Biol. Chem. 295, 3794–3807. DOI: 10.1074 / jbc.ra119.012014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цао, Т., Тонгам У. и Джин Дж. П. (2019). Тропонин беспозвоночных: понимание эволюции и регуляции сокращения поперечно-полосатых мышц. Arch. Biochem. Биофиз. 666, 40–45. DOI: 10.1016 / j.abb.2019.03.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чейни, Дж. А., Аллен, Дж. Дж., И Шварц, С. М. (2017). Разнообразие в организации эластиновых пучков и внутримембранозных мышц крыльев летучих мышей. J. Anat. 230, 510–523. DOI: 10.1111 / joa.12580
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чейни, Дж. А., Коноу, Н., Миддлтон, К. М., Брейер, К. С., Робертс, Т. Дж., Гиблин, Э. Л. и др. (2014). Функция мембранных мышц податливых крыльев летучих мышей. Bioinspir. Биомими. 9: 025007. DOI: 10.1088 / 1748-3182 / 9/2/025007
CrossRef Полный текст | Google Scholar
Комб, С. А., и Дэниел, Т. Л. (2003). В воздух: вклад аэродинамических и инерционно-упругих сил в изгиб крыла у ястреба Manduca sexta. J. Exp. Биол. 206 (Pt 17), 2999–3006. DOI: 10.1242 / jeb.00502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Конли, К. Э., и Линдстедт, С. Л. (2002). Энергосберегающие механизмы в мышцах: стратегия минимизации. J. Exp. Биол. 205 (Pt 15), 2175–2181.
Google Scholar
Каттс, А. (1986). Длина саркомеров в мышцах крыльев изменяется в цикле взмахов крыльев у двух видов птиц. J. Zool. 209, 183–185.DOI: 10.1111 / j.1469-7998.1986.tb03574.x
CrossRef Полный текст | Google Scholar
Дехманн, Д. К. Н., Сафи, К., и Вонхоф, М. Дж. (2006). Соответствие морфологии и рациона дискокрылых летучих мышей Thyroptera tricolor (Chiroptera). J. Mammal. 87, 1013–1019. DOI: 10.1644 / 05-mamm-a-424r2.1
CrossRef Полный текст | Google Scholar
Дикинсон, М., Фарман, Г., Фрай, М., Бекьярова, Т., Гор, Д., Моган, Д. и др. (2005). Молекулярная динамика циклически сокращающихся летных мышц насекомых in vivo. Природа 433, 330–334. DOI: 10.1038 / nature03230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дадли Р. (1998). Атмосферный кислород, гигантские палеозойские насекомые и эволюция воздушных двигательных способностей. J. Exp. Биол. 201: 1043.
Google Scholar
Данлоп, Дж. А., и Гарвуд, Р. Дж. (2018). Наземные беспозвоночные в экосистеме Rhynie Chert. Philos. Пер. R. Soc. Лондон. B Biol. Sci. 373: 20160493.DOI: 10.1098 / rstb.2016.0493
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эллингтон, К. П. (1985). Мощность и эффективность мускулов полета насекомых. J. Exp. Биол. 115: 293.
Google Scholar
Евангелиста Д., Фернандес М. Дж., Бернс М. С., Гувер А. и Дадли Р. (2010). Парящая энергия и тепловой баланс у колибри Анны ( Calypte anna ). Physiol. Biochem. Zool. 83, 406–413. DOI: 10.1086/651460
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фот, К., Тишлингер, Х., и Раухут, О. В. М. (2014). Новый образец археоптерикса дает представление об эволюции меловых перьев. Природа 511, 79–82. DOI: 10.1038 / природа13467
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гаудиозо, П. Дж., Диас, М. М., и Баркес, Р. М. (2017). Морфология осевого скелета семи родов рукокрылых (Chiroptera: Phyllostomidae). An. Акад. Бюстгальтеры. Cienc. 89, (3 доп.), 2341–2358. DOI: 10.1590 / 0001-3765201720170076
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джордж Дж. К. и Бергер А. Дж. (1966). Птичья миология. Кембридж, Массачусетс: Academic Press.
Google Scholar
Gleiss, A.C., Jorgensen, S.J., Liebsch, N., Sala, J.E., Norman, B., Hays, G.C., et al. (2011). Конвергентная эволюция моделей движения летающих и плавающих животных. Nat. Commun. 2: 352.
Google Scholar
Гордон А. М., Хомшер Э. и Ренье М. (2000). Регулирование сокращения поперечно-полосатой мышцы. Physiol. Ред. 80, 853–924.
Google Scholar
Hao, Y., Miller, M. S., Swank, D. M., Liu, H., Bernstein, S. I., Maughan, D. W., et al. (2006). Пассивная жесткость в мышцах непрямого полета Drosophila снижается за счет нарушения фосфорилирования парамиозина, но не за счет замещения шарнира S2 эмбрионального миозина. Biophys. J. 91, 4500–4506. DOI: 10.1529 / biophysj.106.088492
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Харрисон, Дж. Ф., и Лайтон, Дж. Р. (1998). Кислородно-чувствительный полетный метаболизм у стрекозы erythemis simplicicollis. J. Exp. Биол. 201: 1739.
Google Scholar
Хендерсон, К. А., Гомес, К. Г., Новак, С. М., Ми-Ми, Л., и Грегорио, К. С. (2017). Обзор мышечного цитоскелета. Компр. Physiol. 7, 891–944. DOI: 10.1002 / cphy.c160033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хермансон, Дж. У., и Альтенбах, Дж. С. (1981). Функциональная анатомия мышц, ведущих вниз, у бледной летучей мыши. Антрозоус паллидус . J. Mammal. 62, 795–800. DOI: 10.2307 / 1380600
CrossRef Полный текст | Google Scholar
Хермансон, Дж. У., Райан, Дж. М., Кобб, М. А., Бентли, Дж., И Шаттс, Дж. У. А. (1998). Гистохимический и электрофоретический анализ основной летучей мышцы нескольких летучих мышей-филлостомид. Оттава: Canadian Science Publishing, 1983–1992.
Google Scholar
Хомбергер, Д. Г., и де Сильва, К. Н. (2015). Функциональная микроанатомия покровов оперения: значение для эволюции птиц и птичьего полета 1. Am. Zool. 40, 553–574. DOI: 10.1668 / 0003-1569 (2000) 040 [0553: fmotfb] 2.0.co; 2
CrossRef Полный текст | Google Scholar
Хупер, С. Л., Хоббс, К. Х., и Тума, Дж. Б. (2008). Мышцы беспозвоночных: тонкая и толстая филаментная структура; молекулярная основа сокращения и его регуляции, ловля и асинхронность мышцы. Прог. Neurobiol. 86, 72–127. DOI: 10.1016 / j.pneurobio.2008.06.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ивамото, Х., Нисикава, Ю., Вакаяма, Дж., И Фудзисава, Т. (2002). Прямое рентгеновское наблюдение одиночной гексагональной решетки миофиламентов в нативных миофибриллах поперечно-полосатой мышцы. Biophys. J. 83, 1074–1081. DOI: 10.1016 / s0006-3495 (02) 75231-4
CrossRef Полный текст | Google Scholar
Джебб Д., Фоли Н.М., Уилан, К. В., Тузалин, Ф., Пуэчмайл, С. Дж., И Тилинг, Е. К. (2018). Митогеномика на уровне популяции долгоживущих летучих мышей выявляет динамическую гетероплазмию и ставит под сомнение теорию свободных радикалов старения. Sci. Отчет 8: 13634.
Google Scholar
Дженни-Эйерманн, С. (2017). Энергетический обмен во время продолжительного полета и послеполетной фазы восстановления. J. Comp. Physiol. Нейроэтол. Sens. Neural. Behav. Physiol. 203, 431–438. DOI: 10.1007 / s00359-017-1150-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джин, Дж.П. и Саманез Р. А. (2001). Эволюция металлсвязывающего кластера в NH (2) -концевой вариабельной области тропонина Т быстрых скелетных мышц птиц: функциональное расхождение на основе устойчивости к структурному дрейфу. J. Mol. Evol. 52, 103–116. DOI: 10.1007 / s0023
139
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзинь, Дж. П., Чжан, З., и Баутиста, Дж. А. (2008). Разнообразие изоформ, регуляция и функциональная адаптация тропонина и кальпонина. Crit. Преподобный Эукариот. Gene Expr. 18, 93–124. DOI: 10.1615 / critreveukargeneexpr.v18.i2.10
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Найт, К. (2019). Мигрирующие седовласые летучие мыши точно настраивают оцепенение для экономии энергии. J. Exp. Биол. 222: jeb202366. DOI: 10.1242 / jeb.202366
CrossRef Полный текст | Google Scholar
Коноу, Н., Чейни, Дж. А., Робертс, Т. Дж., Ириарте-Диас, Дж., Брейер, К. С., Вальдман, Дж. Р. С. и др.(2017). Зависимая от скорости модуляция интенсивности и кинематики набора крыловых мышц у двух видов летучих мышей. J. Exp. Биол. 220: 1820. DOI: 10.1242 / jeb.144550
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ковач, Х., Стабентхайнер, А., Шмаранцер, С. (2010). Терморегуляция медоносных пчел, добывающих пищу в воде — Баланс эндотермической активности с радиационным притоком тепла и функциональными требованиями. J. Insect Physiol. 56, 1834–1845. DOI: 10.1016 / j.jinsphys.2010.08.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ланге, С., Пиноцис, Н., Агаркова, И., Элер, Э. (2020). М-группа: недооцененная часть саркомера. Biochim. Биофиз. Acta Mol. Cell Res. 1867: 118440. DOI: 10.1016 / j.bbamcr.2019.02.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леманн, Ф. О. (2001). Эффективность производства аэродинамической силы у дрозофилы. Комп.Biochem. Physiol. A. Mol. Интегр. Physiol. 131, 77–88. DOI: 10.1016 / s1095-6433 (01) 00467-6
CrossRef Полный текст | Google Scholar
Марден, Дж. Х., Фитцхью, Г. Х., Вольф, М. Р., Арнольд, К. Д., и Роуэн, Б. (1999). Альтернативное сращивание, чувствительность мышц к кальцию и модуляция летных характеристик стрекозы. Proc. Natl. Акад. Sci. США 96, 15304–15309. DOI: 10.1073 / pnas.96.26.15304
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Матье-Костелло, О., Szewczak, J.M., Logemann, R.B., и Agey, P.J. (1992). Геометрия кроветворения в летучей мышце летучей мыши по сравнению с задней конечностью летучей мыши и камбаловидной мышцей крысы. Am. J. Physiol. 262 (6 баллов 2), R955 – R965.
Google Scholar
Макгиннис, Дж. П., Папа, К. М., и Флетчер, Д. Л. (1989). Влияние пентобарбитала натрия на pH, длину саркомера и болезненность мышц большой грудной мышцы бройлера. Poult. Sci. 68, 244–248. DOI: 10.3382 / пс. 0680244
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Макгуайр, Л.П., Фентон, М. Б., Гульельмо, К. Г. (2013). Сезонная регуляция катаболических ферментов и переносчиков жирных кислот в летательных мышцах мигрирующих седых летучих мышей, Lasiurus cinereus . Комп. Biochem. Physiol. B. Biochem. Мол. Биол. 165, 138–143. DOI: 10.1016 / j.cbpb.2013.03.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Макнаб, Б.К., и О’Доннелл, К. (2018). Поведенческая энергетика летучих мышей Новой Зеландии: ежедневное оцепенение и спячка, непрерывность. Комп. Biochem. Physiol. A. Mol. Интегр. Physiol. 223, 18–22. DOI: 10.1016 / j.cbpa.2018.05.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Миллер, М.С., Фарман, Г.П., Брэддок, Дж. М., Сото-Адамес, Ф. Н., Ирвинг, Т. К., Вигоро, Дж. О. и др. (2011). Регуляторное фосфорилирование легкой цепи и N-концевое удлинение увеличивают связывание поперечного мостика и выходную мощность у дрозофилы при расстоянии между решетками миофиламентов in vivo. Biophys. J. 100, 1737–1746.DOI: 10.1016 / j.bpj.2011.02.028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моррис К. Р., Нельсон Ф. Э. и Аскью Г. Н. (2010). Требования к метаболической мощности для полета и оценка эффективности летных мышц у кореллы ( Nymphicus hollandicus ). J. Exp. Биол. 213, 2788–2796. DOI: 10.1242 / jeb.035717
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нейв Р. и Вебер К. (1990). Миофибриллярный белок мышц насекомых, связанный с тайтином позвоночных, соединяет полосу Z и полосу A: очистка и молекулярная характеристика мини-тайтина беспозвоночных. J. Cell Sci. 95 (Pt 4), 535–544.
Google Scholar
Нельсон К. Р. и Тидуэлл В. Д. (1987). Brodioptera Stricklani N. Sp. (Megasecoptera: Brodiopteridae), новое ископаемое насекомое из сланцевой формации Верхнего Мэннинг-Каньона, Юта (самый нижний намурский период B). Психея 94, 309–316. DOI: 10.1155 / 1987/52032
CrossRef Полный текст | Google Scholar
Огут О., Гранзье Х. и Джин Дж. П. (1999). Кислые и основные изоформы тропонина Т в зрелых быстро сокращающихся скелетных мышцах и влияние на сократимость. Am. J. Physiol. 276, C1162 – C1170.
Google Scholar
Огут, О., и Джин, Дж. П. (1998). Онтогенетически регулируемые, генерируемые сплайсингом альтернативной РНК специфичные для грудных мышц изоформы тропонина Т и роль Nh3-концевой гипервариабельной области в толерантности к ацидозу. J. Biol. Chem. 273, 27858–27866. DOI: 10.1074 / jbc.273.43.27858
PubMed Аннотация | CrossRef Полный текст | Google Scholar
О’Ши, Т.Дж., Крайан, П.М., Каннингем А. А., Фукс А. Р., Хейман Д. Т., Луис А. Д. и др. (2014). Полет летучих мышей и зоонозные вирусы. Emerg. Заразить. Дис. 20, 741–745. DOI: 10.3201 / eid2005.130539
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перон С., Зордан М. А., Магнабоско А., Реджиани К. и Мегигиан А. (2009). От потенциала действия к сокращению: нервный контроль и связь возбуждения-сокращения в личиночных мышцах дрозофилы. Комп. Biochem. Physiol.A. Mol. Интегр. Physiol. 154, 173–183. DOI: 10.1016 / j.cbpa.2009.04.626
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перц-Эдвардс, Р. Дж., Ирвинг, Т. К., Бауман, Б. А., Гор, Д., Хатчинсон, Д. К., Кшич, У. и др. (2011). Дифракция рентгеновских лучей свидетельствует о связях миозин-тропонин и движении тропомиозина во время активации растяжения летающих мышц насекомых. Proc. Natl. Акад. Sci. США 108, 120–125. DOI: 10.1073 / pnas.1014599107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пур, С.О., Санчес-Хайман А. и Гослоу Г. Э. (1997). Взмах крыла вверх и эволюция махающего полета. Природа 387, 799–802. DOI: 10.1038 / 42930
CrossRef Полный текст | Google Scholar
Prokop, J., Pecharova, M., Nel, A., Hornschemeyer, T., Krzeminska, E., Krzeminski, W., et al. (2017). Палеозойские зачатки крыльев нимфалий подтверждают двойную модель происхождения крыльев насекомых. Curr. Биол. 27, 263–269. DOI: 10.1016 / j.cub.2016.11.021
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райзер, П.Дж., Уэлч, К. К., Суарес, Р. К., и Альтшулер, Д. Л. (2013). Очень низкая способность генерировать силу и необычно высокая температурная зависимость в мышечных волокнах полета колибри. J. Exp. Биол. 216: 2247. DOI: 10.1242 / jeb.068825
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Росс, А. (2017). Эволюция насекомых: происхождение крыльев. Curr. Биол. 27, R113 – R115.
Google Scholar
Скулборстад, А.Дж., Шварц, С.М., и Гоулборн, Н.С. (2015). Биаксиальная механическая характеристика кожи крыла летучей мыши. Bioinspir. Биомим. 10: 036004. DOI: 10.1088 / 1748-3190 / 10/3/036004
CrossRef Полный текст | Google Scholar
Снеллинг, Э. П., Сеймур, Р. С., Мэтьюз, П. Г. Д. и Уайт, К. Р. (2012). Максимальная скорость метаболизма, относительная подъемная сила, частота взмахов крыльев и амплитуда ударов во время полета на привязи взрослой саранчи Locusta migratoria. J. Exp. Биол. 215, 3317–3323. DOI: 10.1242 / jeb.069799
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Stabentheiner, А., и Ковач, Х. (2014). Энергетическая оптимизация кормления медоносных пчел: гибкое изменение стратегии в ответ на экологические проблемы. PLoS One 9: e105432. DOI: 10.1371 / journal.pone.0105432
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Старлинг Р. К., Хаммер Д. Ф. и Альтшульд Р. А. (1998). Содержание АТФ в миокарде человека и сократительная функция in vivo. Мол. Клетка. Biochem. 180, 171–177. DOI: 10.1007 / 978-1-4615-5687-9_19
CrossRef Полный текст | Google Scholar
Суонк, Д.М., Ноулз, А.Ф., Саггс, Дж. А., Сарсоза, Ф., Ли, А., Моган, Д. У. и др. (2002). Домен-преобразователь миозина модулирует работу мышц. Nat. Cell Biol. 4, 312–316. DOI: 10.1038 / ncb776
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Суонк, Д. М., Кронерт, В. А., Бернштейн, С. И., и Моган, Д. В. (2004). Альтернативные N-концевые области тяжелой цепи миозина дрозофилы настраивают мышечную кинетику для оптимальной выходной мощности. Biophys. J. 87, 1805–1814.DOI: 10.1529 / biophysj.103.032078
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Szentesi, P., Zaremba, R., van Mechelen, W., and Stienen, G.J. (2001). Использование АТФ для поглощения кальция и выработки силы в различных типах волокон скелетных мышц человека. J. Physiol. 531 (Pt 2), 393–403. DOI: 10.1111 / j.1469-7793.2001.0393i.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Токита, М., Абэ, Т., и Судзуки, К.(2012). Основа развития мышцы крыла летучей мыши. Природа 3: 1302.
Google Scholar
Вейдани, Х. Р., Бурма, Д. Б., Шварц, С. М., и Брейер, К. С. (2018). Динамика парящего полета у колибри, насекомых и летучих мышей с последствиями для воздушной робототехники. Bioinspir. Биомим. 14: 016003. DOI: 10.1088 / 1748-3190 / aaea56
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Велтен, Б. П., Уэлч, К. К. Мл. (2014).Изоформы тяжелых цепей миозина в мышцах полета и ног колибри и зебровых вьюрков. Am. J. Physiol. Regul. Интегр. Комп. Physiol. 306, R845 – R851.
Google Scholar
Войт, К. К., Фрик, В. Ф., Холдерид, М. В., Холланд, Р., Керт, Г., Мелло, М. А. Р. и др. (2017). Принципы и закономерности движений летучих мышей: от аэродинамики до экологии. Q. Rev. Biol. 92, 267–287. DOI: 10.1086 / 693847
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фон Буссе, Р., Хеденстрём А., Винтер Ю. и Йоханссон Л. К. (2012). Кинематика и форма крыла в зависимости от скорости полета у летучей мыши, & it; em & gt; Leptonycteris yerbabuenae & it; / em & gt. Biol. Открыть 1
Google Scholar
Ван Дж. И Джин Дж. П. (1997). Первичная структура и переход из кислой в основную при развитии 13 альтернативно сплайсированных изоформ Т тропонина быстрых скелетных мышц мыши. Ген 193, 105–114. DOI: 10.1016 / s0378-1119 (97) 00100-5
CrossRef Полный текст | Google Scholar
Ван, Дж., и Джин, Ж.-П. (1998). Конформационная модуляция тропонина Т за счет конфигурации вариабельной области Nh3-терминала и функциональных эффектов †. Биохимия 37, 14519–14528. DOI: 10.1021 / bi9812322
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, К., Ньюхард, С., Раманат, С., Шеппард, Д., Суонк, Д. М. (2014). Эмбриональный домен конвертера миозина влияет на активацию растяжения мышц непрямого полета, выработку энергии и полет у дрозофилы. J. Exp. Биол. 217, 290–298. DOI: 10.1242 / jeb.091769
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уорд, С., Моллер, У., Райнер, Дж. М., Джексон, Д. М., Било, Д., Нахтигол, В. и др. (2001). Метаболическая мощность, механическая мощность и эффективность во время полета в аэродинамической трубе европейского скворца Sturnus vulgaris . J. Exp. Биол. 204 (Pt 19), 3311–3322.
Google Scholar
Функция мышц в птичьем полете: достижение силы и контроля
Philos Trans R Soc Lond B Biol Sci.2011 27 мая; 366 (1570): 1496–1506.
Полевая станция Конкорд, Гарвардский университет, 100 Old Causeway Road, Бедфорд, Массачусетс 01730, США
Авторские права на этот журнал принадлежат © 2011 Королевское обществоЭта статья цитировалась другими статьями в PMC.Abstract
Хлопающий полет предъявляет жесткие требования к физиологической работоспособности животного. Мышцы полета птиц, особенно при меньших размерах тела, обычно сокращаются на высоких частотах и выполняют значительную работу для создания аэродинамической мощности, необходимой для поддержки веса животного в воздухе и преодоления сопротивления.Это контрастирует с наземным движением, которое предлагает механизмы для минимизации потерь энергии, связанных с движением тела, в сочетании с экономией упругой энергии для снижения требований к работе скелетных мышц. Мышцы также производят значительную силу во время плавания, но в основном это делается для преодоления сопротивления тела, а не для поддержки веса животного. Здесь я рассмотрю функцию и архитектуру ключевых летных мышц, связанных с тем, как эти мышцы способствуют выработке мощности, необходимой для взмахов, как мышцы задействуются для управления движением крыльев и как они используются при маневрировании.Эмерджентным свойством основных летных мышц, согласующимся с их необходимостью производить значительную работу, перемещая крылья через большие отклонения во время каждого взмаха крыла, является то, что грудные и надкожные мышцы укорачиваются на большей части длины их покоящихся волокон (33–42). %). Обе мышцы активируются при удлинении или при почти изометрическом развитии силы, улучшая работу, которую они выполняют при последующем сокращении. Две меньшие мышцы, трицепс и бицепс, работают в меньшем диапазоне сократительных напряжений (12–23%), что отражает их роль в контроле формы крыла посредством сгибания и разгибания локтя.Примечательно, что голуби регулируют плоскость хода своего крыла в основном за счет изменения шага всего тела во время взлета и посадки относительно горизонтального полета, позволяя мышцам крыла работать с небольшим изменением времени активации, величины напряжения и характера.
Ключевые слова: полет птицы, нервно-мышечная функция, сила мышц, деформация пучка
1. Введение
Птицы управляют полетом главным образом за счет больших грудных мышц, которые вдавливают крылья в плече. Таким образом, доминирующая роль и большой размер грудной мышцы позволяют критически оценить, как функция мышц приспособлена для удовлетворения требований к механической мощности при машущем полете в различных условиях полета.Меньшая супракоракоидная мышца птиц, составляющая примерно одну пятую размера грудной мышцы, является основным подъемником крыла, активным во время движения вверх, особенно на медленных и умеренных скоростях и во время зависания (на более высоких скоростях полета подъем крыла, вероятно, пассивно создается аэродинамическими силами. воздействуя на крылья, которые остаются вытянутыми во время движения вверх, чтобы поддерживать подъемную силу за счет связанной циркуляции [1,2]). Меньшие внешние и внутренние мышцы крыла помогают модулировать ориентацию крыла и контролировать форму крыла.Эти мышцы, вероятно, вносят вклад в регулировку характеристик крыла как крыла [3–7] и, таким образом, могут косвенно влиять на требования к мощности полета. Однако из-за своего небольшого размера внутренние мышцы крыла, вероятно, вносят небольшую дополнительную механическую мощность для полета.
Предыдущие анализы архитектуры мышцы-сухожилия показали, что мышцы сильно различаются по своей конструкции для изменения длины при создании силы, но из-за их консервативных свойств для создания силы и относительной деформации волокон (отношение изменения активированной длины к длине волокна в состоянии покоя) скелетные мышцы обычно выполняют примерно одинаковое количество работы пропорционально своей массе [8–11].Однако более длинные волокнистые мышцы, такие как птичья грудная мышца, хорошо подходят для выполнения больших движений, необходимых для движения крыльев, для создания эффективной аэродинамической мощности для поддержки веса и преодоления сопротивления. Помимо более длинных волокон, большие рабочие нагрузки также увеличивают диапазон движений, которые производит мышца. Таким образом, ожидается, что рабочие напряжения некоторых летательных мышц будут выше, чем напряжения мышц, которые поддерживают вес животного во время наземного передвижения [12], которые сокращаются в более ограниченных диапазонах напряжения, позволяя производить более экономичное производство силы.Мышцы, имеющие короткие волокна, которые прикрепляются к более длинным сухожилиям, таким как те, которые находятся в ногах наземных животных, производят большие силы и могут восстанавливать значительную упругую энергию из своих сухожилий и апоневроза [12–15]. Эти мышцы лучше всего использовать для движений, требующих небольшого сокращения или удлинения мышцы. Следовательно, перистые мышцы, обладающие этими архитектурными особенностями, обычно встречаются в дистальных отделах конечностей. Внутренние мышцы крыльев птиц обычно коротковолокнистые, перистые и имеют длинные сухожилия.Это позволяет этим мышцам управлять дистальными движениями крыла, в то же время будучи маленькими и легкими. Их функция на сегодняшний день мало изучена, за исключением нескольких сравнительных функционально-анатомических описаний [7,16,17] и оценки паттернов их нервно-мышечной активности [3,16,17]. Тем не менее, эти исследования важны, потому что они обеспечивают основу для будущих исследований, направленных на оценку того, как меньшие внутренние мышцы крыльев используются для достижения полета в разных условиях и у птиц с разным дизайном крыльев и стилями полета.
В контексте этой более ранней работы функции двух основных летательных мышц птиц, грудной и супракоракоидной, рассматриваются здесь в связи с механической мощностью, необходимой для удовлетворения аэродинамических требований для взмахивания крыльев. Подавляющее большинство морфологической и физиологической работы в основном сосредоточено на грудной мышце из-за ее доминирующей роли в обеспечении полета птиц. Следовательно, большая часть обзора функции птичьих мышц будет сосредоточена на грудной мышце, с особым сравнением с ее антагонистом, супракоракоидусом.Предварительный анализ in vivo трехглавой и двуглавой мышц, которые контролируют форму крыла посредством разгибания и сгибания локтя, также рассматриваются в отношении изменений летных характеристик, необходимых для взлета, посадки и маневрирования. Также определены будущие направления исследований для улучшения нашего понимания нервно-мышечного контроля и функционального дизайна полета птиц.
2. Функциональная анатомия основных летательных мышц птиц
Грудная мышца — большая мышца (прибл.8–11% массы тела; [15,16]), который прикрепляется к плечевой кости крыла у дельтовидного гребня (DPC;). Его основная часть (sternobrachialis, SB) происходит от увеличенного грудинного киля с большим количеством передних волокон, отходящих от фуркулы, или «поперечного рычага». Гораздо меньшая часть (thoracobrachialis, TB) берет начало от ребер дорсально. Волокна TB и задняя часть вставки SB на внутреннем апоневрозе, которая сливается с более передними волокнами SB перед прикреплением к DPC. В дополнение к механической работе во время движения вниз грудная мышца также пронатирует крыло.Более мелкая супракоракоидус лежит глубоко в грудной мышце, также берет начало от киля грудины и составляет около одной пятой грудной мышцы (около 2% массы тела). С помощью своего сухожилия, которое вставляется и действует на дорсальной стороне плеча как шкив, supracoracoideus поднимает и супинирует крыло во время движения вверх [18–21]. В то время как грудная мышца состоит, как правило, из длинных волокон с умеренным перистым оперением (голубь: 31–67 мм, в среднем 41 мм), супракоракоидус — это классическая двуногая мышца с короткими волокнами (голубь: 16–23 мм, в среднем 20 мм).Он обеспечивает подъем и супинацию крыла с помощью длинного сухожилия, которое проходит дорсально через плечо через трехкостное отверстие грудного пояса птиц, прежде чем прикрепиться к дорсальной поверхности проксимального отдела плечевой кости, прилегающей к DPC. Грудная мышца состоит в основном из быстроокисляющих (тип IIa) волокон (около 85% у голубей) с меньшим компонентом из быстро-гликолитических волокон (тип IIb) [20,21]. Насколько мне известно, волокнистый состав supracoracoideus не исследовался на голубях, но у европейского скворца он состоит из большей доли (68%) быстро гликолитических волокон по сравнению с быстроокисляющими волокнами [22]; тогда как у зебровых вьюрков, колибри Анны [23] и атлантических тупиков [24] супракоракоидус состоит исключительно из быстроокисляющих волокон.
( a ) Анатомическая организация мускулатуры крыла птиц (адаптировано из Dial [3]), показывающая ключевые мышцы, которые были изучены, и (b ), показывающая общие места, используемые для регистрации силы грудной мышцы через дельтопекторальный гребень (DPC). ) деформация костей, деформация грудного пучка и нервно-мышечная активация (ЭМГ).
3.
In vivo оценка функции мышц птиц во время полетаИз-за его фокального прикрепления на вентральной поверхности DPC у голубей (b ), голубей, корелл, волнистых попугаев, сорок и некоторых других видов птиц. силы, создаваемые грудной мышцей, можно оценить непосредственно с помощью деформаций, регистрируемых с помощью тензодатчика, прикрепленного к дорсальной поверхности DPC (у некоторых видов птиц грудная мышца также вставляется вдоль брюшной проксимальной части плечевой кости, предотвращая этот подход) .Детали для экспонирования и прикрепления тензодатчиков из металлической фольги для получения калиброванных по деформации in vivo записей грудной мышцы описаны в другом месте [25,26]. Хотя существует некоторая неопределенность в калибровке деформации DPC по силе грудной мышцы [27], такие записи обеспечивают надежную и детализированную во времени запись изменяющейся во времени мышечной силы. Другие методы получения мышечной силы и оценки выходной механической мощности для полета птицы также имеют свои ограничения [28,29].Подобный подход, основанный на растяжении скелета, также использовался для извлечения изменяющейся во времени силы, передаваемой супракоракоидной мышцей через прикрепление сухожилия мышцы к проксимальному дорсальному стержню плечевой кости [30].
В сочетании с регистрацией деформации-силы DPC грудной и супракоракоидной мышц, in vivo измерений деформации мышечных пучков в локализованных участках мышц получают с помощью сономикрометрии, метода, основанного на измерениях распространения звуковых импульсов внутри мышца для определения изменения длины [31].Поскольку преобразователи сономикрометрии расположены рядом с пучками мышечных пучков, они обеспечивают измерение деформации пучка, а не напряжения мышечных волокон per se . Тем не менее, эти две меры, скорее всего, будут очень похожи. В большой грудной мышце сонометрические измерения, полученные в нескольких местах (передняя и задняя SB и TB) у голубей, показали одинаковые уровни деформации пучка в большей части SB мышцы, но меньшие деформации в самых задних частях SB и TB мышцы [ 32].Усредняя данные сономикрометрии для деформации пучка по участкам записи (взвешенные по расчетной доле мышечной массы, которую представляет каждый участок) или полагаясь на один участок записи в мышце и предполагая, что этот участок является репрезентативным для мышцы в целом, Общую работу мышцы можно оценить по изменению ее длины. Таким образом, работа мышц определяется деформацией пучка, умноженной на среднюю длину пучка, по отношению к изменяющейся во времени силе, производимой мышцей.Результат изменения длины и силы мышечного пучка визуализируется как рабочая петля в течение цикла взмахов крыльев или сокращений мышц. Время активации мышц регистрируется одновременно с использованием электродов для тонкопроволочной электромиографии (ЭМГ), вставленных и закрепленных рядом с теми пучками, для которых регистрируется сонометрическая оценка напряжения [31]. ЭМГ обеспечивает измерение времени активации мышц и относительного моторного набора в зависимости от изменения силы и длины мышц.В целом сила, напряжение и нервно-мышечная активация, зарегистрированные в мышце, служат для описания временной динамики сократительной способности мышцы в различных условиях полета.
4. Функциональный анализ грудных и супракоракоидных мышц во время полета
Грудная мышца активируется, чтобы сокращаться в конце хода вверх, до разворота крыла ( a ). Развитие силы следует вскоре после начала активации (примерно 2–8 мс у голубей и корелл) и достигает пика в начале хода вниз, продолжаясь до конца хода вниз.Грудная мышца слегка растягивается или остается почти изометричной (в зависимости от изучаемого вида и условий полета), поскольку сила развивается в конце взмаха вверх и за счет разворота крыльев, чтобы начать движение вниз (рисунки и). Развивая силу, когда она почти изометрична или при кратковременном растяжении, скорость нарастания силы и величина пиковой силы заметно увеличиваются из-за эффектов сила-скорость [35,36]. В результате работа, которую выполняет грудная мышца, существенно увеличивается, в то время как мышца укорачивается в течение оставшейся части хода вниз.Деактивация грудной мышцы происходит в начале хода вниз, почти одновременно с моментом выработки пиковой силы. Это позволяет мышце расслабиться почти до нулевой силы перед пассивным растяжением при движении вверх. Важно отметить, что это уменьшает антагонистическую («негативную») работу, необходимую супракоракоидусу для поднятия крыла. Время деактивации грудной мышцы относительно ее продолжающегося производства силы указывает на проблематичный характер вывода о производстве мышечной силы на основании только записей ЭМГ.
Типичный образец in vivo записей деформации грудного пучка, нервно-мышечной активации (ЭМГ) и силы для трех взмахов крыльев у кореллы, летящей со скоростью 7 м с -1 в аэродинамической трубе. Сплошная линия, запястье; пунктирная линия, кончик крыла. По материалам Hedrick et al . [33].
Типичный образец in vivo рабочих петель, создаваемых грудной клеткой ( a ) кореллы ( Nymphicus hollandicus ) на трех разных скоростях полета (адаптировано из Hedrick et al .[33]) и ( b ) грудные клетки трех других видов: голуби с круглой шеей ( Streptopelia risoria ), голуби ( Columba livia ) и утки кряквы ( Anas platyrynchos ) (адаптировано из Tobalske ). и др. [27], Бивенер и др. [31] и Уильямсон и др. [34]). Сила, создаваемая мышцей, отображается в зависимости от деформации ее пучка ( L / L o , где L o — длина мышцы в состоянии покоя: напряжение = 1.0). На первой панели ( a ) пунктирный прямоугольник обозначает максимальную работу, которую мышца может произвести при максимальной силе и напряжении; Реализованная работа мышцы составляет 68% от теоретического максимума. Диапазон напряжения для всех мышц одинаков (0,9–1,3, или 40% диапазона изменения длины мышцы), но диапазоны сил различаются ( b ) из-за разного размера мышц. Жирная серая часть каждой рабочей петли представляет период нервно-мышечной активации, измеренный с помощью ЭМГ.Стрелки обозначают направление изменения силы и длины пучка.
Для тех изученных видов [27,33,34], поведение грудной мышцы in vivo с силой-длиной в целом одинаково во всем диапазоне скоростей полета и условий (). Как отмечалось выше, активация грудной мышцы у этих видов происходит на поздних стадиях движения вверх, когда мышца удлиняется (это наиболее экстремально у кряквы, b ) или почти изометрична, что позволяет мышце быстро развивать силу в течение заданного промежутка времени. уровень активации.В отличие от классических ожиданий в отношении деформации рабочего пучка мышцы (приблизительно 10-15% длины покоя на основе изометрических свойств длины и силы [35,36]), грудная мышца этих видов подвергается деформации 32-40% во время различные условия полета (взлет, подъем и спуск, изменение скорости во время горизонтального полета), растяжение на 20–30% за пределы длины покоя мышцы (измеряется, когда крылья сложены относительно тела птицы на жердочке) и укорачивание 8 –12% меньше длины покоя.Этот большой скачок напряжения лежит в основе способности грудной мышцы выполнять значительную работу во время движения вниз каждого цикла сокращения. Было обнаружено, что силы, создаваемые грудной мышцей голубя, различаются примерно на 40% в зависимости от условий полета, начиная от взлета и восходящего полета и заканчивая приземлением и спуском [26]. Было обнаружено, что сила, создаваемая грудной корелой во время горизонтального полета на скоростях от 1 до 14 м / с –1 в аэродинамической трубе, варьируется на 65% [33].По оценкам, эти силы составляют менее 40–60% максимальной изометрической силы, которую может генерировать мышца [26], частично отражая быстрое укорачивание, которому мышца подвергается, чтобы произвести работу. У корелл, голубей и голубей грудная мышца достигает 58–73% максимальной теоретической выходной мощности, возможной для наблюдаемой силы и диапазона активной деформации [30,33] ( a ).
Неудивительно, что супракоракоидус голубей демонстрирует зеркальные характеристики силы, длины и времени активации относительно грудной мышцы [30] ().Как основная мышца, совершающая движение вверх, supracoracoideus активируется в конце хода вниз, незадолго до разворота крыльев. Мышца быстро развивает силу, будучи почти изометричной, достигая пика силы очень рано при движении вверх. Раннее начало развития силы с помощью supracoracoideus, вероятно, отражает его роль в замедлении и повторном ускорении крыла во время перехода вниз-вверх, а также его роль в супинации крыльев [19]. Оценки запаса упругой энергии в сухожилии надкоракоидной мышцы (51-62 мДж во время нивелирования и 88-85 мДж во время восходящего полета) согласуются с этой ролью, учитывая, что величина инерционной кинетической энергии превышает количество запасенной и возвращаемой упругой энергии сухожилие supracoracoideus [30].Дополнительная инерционная сила движения крыла, вероятно, преобразуется в полезную аэродинамическую мощность в основном при движении вниз, как это традиционно предполагалось [37]. Быстрая супинация крыла, производимая supracoracoideus, важна для достижения кратковременного подъема вверх, с потенциалом создания положительной подъемной силы у птиц с кинематикой полета с изменением кончика крыла [38] или для минимизации нежелательной отрицательной подъемной силы. Это также максимизирует продолжительность подъема подъемной силы вниз и, вероятно, было важной особенностью в развитии активного взмахивающего взмаха в полете [19].Быстрая супинация крыла для инициирования подъема вверх у рыжих колибри [39] является ключом к способности этого вида генерировать положительную подъемную силу подъема вверх, которая, по оценкам, составляет 25–33% от их общей подъемной силы [40]. У голубей величина силы, создаваемой антагонистически между двумя мышцами, была оценена как небольшая [30]. Во время медленного горизонтального полета отрицательная работа грудной мышцы голубя непосредственно перед концом движения вверх составляет около 18 процентов от положительной работы, которую мышцы выполняют во время движения вниз.Это вполне может отражать роль в поглощении инерционной энергии крыла, поскольку оно замедляется в конце хода вверх [30]. Для сравнения, отрицательная работа голубя supracoracoideus составляет 14 процентов положительной работы, которую выполняет мышца, и происходит в конце хода вниз, чтобы замедлить крыло в это время.
Репрезентативные записи деформации пучка голубя супракоракоид (высота крыла), ЭМГ и силы, деформации пучка грудной мышцы (депрессия крыла), ЭМГ и силы, записанные во время взлета с приподнятой платформы и горизонтального полета примерно при 4.5 м с −1 (показаны семь взмахов крыльев). Серые панели представляют собой ход вниз для начальных четырех циклов взмахов крыльев, при этом ход вверх на белом фоне (адаптировано из Tobalske & Biewener [30]).
Короткие волокна двупоясной надкожной мышцы требуют, чтобы они действовали на большие нагрузки, подобные таковым у грудной мышцы. Деформации пучка Supracoracoideus составляют от 33 до 40 процентов длины покоя мышцы во время спуска, подъема и горизонтального полета [30].Пучки supracoracoideus также претерпевают меньшую степень растяжения относительно их длины покоя (6–12% в условиях полета) по сравнению с их результирующей деформацией укорочения (−27% для всех условий полета). Этот паттерн изменения длины пучка относительно длины покоя противоположен паттерну деформации, наблюдаемому в пучках грудной мышцы, которые удлиняются на 20–30% от их длины покоя, прежде чем укорачиваться примерно на 10% меньше, чем в состоянии покоя в конце хода вниз () . Интересно, что модуляция мышечного напряжения в супракоракоиде отражает в основном различия в степени депрессии крыла (растяжение супракоракоидуса и его сухожилия), которое происходит в конце хода вниз в трех изученных условиях полета.Из-за своего относительно небольшого размера супракоракоидный голубь производит в 1,6 раза больше мышечной мощности, чем грудная мышца. Это отражает гораздо более высокие рабочие напряжения (сила, нормированная на физиологическую площадь поперечного сечения) супракоракоидной мышцы, которая составляла от 85 до 125 кПа при спуске по сравнению с восходящим полетом, по сравнению с напряжениями 50–58 кПа в грудной мышце в тех же условиях полета. [30] и 57–76 кПа в более раннем исследовании грудной мышцы голубя с поправкой на предполагаемую миофибриллярную площадь мышцы [26].
5. Сравнительные данные выходной мощности грудной мышцы птиц в зависимости от скорости
Поскольку грудная мышца является доминирующей летной мышцей птиц (у голубей грудная мышца составляет 60% от общей массы мышц крыла, А.А. Бивенер 2010, неопубликованные данные), мощность мышц Выходная мощность может использоваться для оценки того, как выходная мощность всего тела и, косвенно, выходная аэродинамическая мощность изменяются в зависимости от условий полета и скорости птицы. Измерения механической выходной мощности грудных мышц и частоты взмахов крыльев были опубликованы для черноклювых сорок ( Pica pica ), корелл ( Nymphicus hollandicus ) и голубей с кольчатой шеей ( Streptopelia risoria ) в диапазоне скоростей полета во время полета. ровный и устойчивый в аэродинамической трубе [27,41] ().За исключением сорок, два других вида показали U-образную кривую зависимости мощности от скорости полета, что в целом согласуется с аэродинамической теорией. Это отражает высокие затраты на индуцированную мощность при низких скоростях полета и зависании, которые уменьшаются с увеличением скорости, а также высокие затраты на мощность профиля и паразитов (из-за увеличения лобового сопротивления крыла и корпуса) при более высоких скоростях полета. Отсутствие наблюдаемого увеличения силы грудных мышц при более высоких скоростях полета у сорок может отражать либо неспособность этого вида с его более низким удлинением и менее заостренными крыльями достигать достаточной тяги, чтобы преодолеть профиль, и сопротивление паразитов обходится ему. влечет за собой более высокие скорости полета, ограничивающие максимальную скорость, которую он может достичь [27], или то, что птицы не желали летать на более высоких скоростях в аэродинамической трубе.Хотя аэродинамическая труба, используемая для изучения сорок, была меньше (на 50% меньше поперечных размеров рабочего участка), чем та, которая использовалась для изучения корелл и голубей, такие артефакты, как возможное влияние земли или стены [42], не оценивались авторы должны стать основой более низкой стоимости мощности сорок при более высоких скоростях полета. У двух других видов (корелл и голубей) мощность грудных мышц на самых высоких скоростях полета превышала мощность, производимую, когда птицы почти зависали ().Таким образом, хотя выходная мощность грудной мышцы была высокой, как и ожидалось, в течение 1 м с -1 полета у сорок, остается неясным, почему выходная мощность мышцы не достигает или не превышает этого уровня при более высоких скоростях полета.
Сравнительные кривые мощности полета для трех видов птиц, показывающие изменения в мышечной мощности грудной мышцы (определяемой на основе калиброванных записей DPC-деформации-силы и деформации пучка) от скорости полета в аэродинамической трубе (адаптировано из Tobalske et al .[27]).
Учитывая, что другие мышцы участвуют в махающем полете и выполняют механическую работу, это, безусловно, тот случай, когда общая потребность в механической мощности мышц для полета больше, чем оценки, основанные только на грудных мышцах. В исследовании голубей, для которых определялась мощность грудных и надкожных мышц [30], включение мощности надкоракоидных мышц увеличивает общую мощность полета почти на 25%. Мощность грудной клетки на всех режимах полета составила 3.В 2 раза больше, чем у supracoracoideus, но меньше, чем почти пятикратная разница в мышечной массе. Вместе эти две мышцы составляют 71 процент (A. A. Biewener 2010, неопубликованные данные) от общей массы боевых мышц голубя. Если оставшиеся меньшие внешние и внутренние мышцы крыльев выполняют ту же относительную удельную работу по массе, это предполагает общую потребность в мощности, которая может быть почти на 40 процентов больше, чем определенная для одной только грудной мышцы.
Аэродинамические модели для оценки требований к мощности полета птиц на разных скоростях [43–45] обычно используются для вывода экологических стратегий увеличения дальности полета птиц или минимизации потребности в метаболической энергии для полета в зависимости от времени [46 ].Хотя измерения выходной механической мощности грудной мышцы согласуются с общим изменением мощности в зависимости от скорости полета (наибольшее значение на малых и высоких скоростях и минимум на промежуточной скорости полета), абсолютная величина затрат энергии на колебательный полет между видами и скорости остаются неопределенными. Аргументы в пользу того, что один подход и / или метод превосходит другой, остаются неубедительными. Это происходит из-за допущений и упрощений, которые квазистационарная аэродинамическая теория делает для оценки требований к мощности полета, а также из-за неопределенностей в калибровке силы грудной мышцы и оценке региональных профилей деформации пучка на основе записей локализованных пучков на экспериментальной стороне.Более поздние попытки оценить выходную мощность мышц на основе отдельных измерений мышц рабочей петли относительно записей ЭМГ, сделанных во время полета [28,29], также имеют свои ограничения. Сюда входит оценка набора мышц по относительной величине ЭМГ на разных скоростях полета для корректировки измерений максимальной стимулированной мышечной силы, полученных из рабочих экспериментов in vitro и . Такой подход обязательно определяет изменение требований к мощности полета на основе изменений зарегистрированной интенсивности ЭМГ.Это также приводит к более низким оценкам требований к мощности летных мышц корелл (минимальная стоимость мощности = приблизительно 40 Вт кг −1 при 7 мс −1 ) по сравнению с таковыми (74–79 Вт кг −1 при 5 мс). –7 мс -1 ), полученные с использованием измерений силы на основе DPC [27,33]. Дополнительные исследования, которые уточняют использование этих подходов или используют другие методы [47], улучшат нашу способность количественно оценивать абсолютные затраты энергии на колебательный полет для конкретных видов, действующих в различных условиях полета.В соответствии с результатами исследования in vitro, мышечной работы и интенсивности ЭМГ, которые приписывают изменение выходной мощности мышц в зависимости от скорости полета из-за изменений в интенсивности ЭМГ [28,29], результаты основаны на in vivo, деформации пучка, ЭМГ и DPC-деформации. -калиброванные записи силы [27,33] также показали, что интенсивность ЭМГ сильно коррелирует с мышечной силой ( R 2 = 0,92). В последних исследованиях на изменения интенсивности ЭМГ приходилось 65% модуляции мышечной силы, при этом изменения амплитуды деформации пучка составляли 25%, а изменения частоты взмахов крыльев составляли лишь 10% модуляции мышечной силы [27, 33].
Используя измерения силы грудной мышцы, откалиброванной с помощью DPC, и деформации пучка для определения выходной мощности in vivo pectoralis, сравнительные кривые мощности для различных видов, изученных на сегодняшний день, позволяют предположить, что нагрузка на крыло, а также форма крыла и хвоста вероятно, важный фактор, определяющий относительную стоимость мышечной силы вида. Голуби имеют самую высокую нагрузку на крыло (36 Н · м −2 ) среди видов, изученных на сегодняшний день [2], и, соответственно, имеют самые высокие относительные затраты на мощность полета в широком диапазоне скоростей ().У сорок крылья с самым низким соотношением сторон (5,0; по сравнению с волнистыми попугайчиками: 7,3, кореллы: 7,0 и голуби: 5,7) и закругленные кончики крыльев, что, вероятно, помогает снизить требования к мощности, зависящей от их мышечной массы, но также может ограничивать максимальную скорость, которую они могут достичь.
В настоящее время было бы неосмотрительно сильно полагаться на точность результатов экспериментального или теоретического моделирования, чтобы точно указать, имеет ли вид минимальные затраты энергии при определенной скорости полета, учитывая неопределенность и ограничения в разрешающей способности и точности доступные в настоящее время подходы, используемые для оценки затрат на полетную мощность.Например, в то время как данные о потреблении кислорода кореллами [48,49] указывают на минимальную стоимость метаболической энергии на уровне 10 мс -1 , измерения мощности грудных мышц предполагают минимум в диапазоне 5-7 мс -1 [ 27,29]. Комбинирование результатов метаболической мощности корелл с результатами их механической мышечной силы [49] показывает, что эффективность мышц увеличивается с увеличением скорости полета, в пределах от 6,9 до 11,2%, основываясь на данных о мышечной силе Морриса и Аскью [29], или от 12.От 2 до 28,3% на основании данных силы DPC-pectoralis и деформации пучка Tobalske et al . [27].
Различия в эффективности мышц, вероятно, обусловлены тем, что скорость сокращения пучков грудных мышц зависит от скорости полета. У корелл [27,33] скорости укорочения пучков варьировались от 5,19 до 6,73 длины мускулов в секунду при скоростях полета от 1 до 13 м с -1 . Диапазон эффективности, полученный из измерений in vitro мышц с поправкой на интенсивность ЭМГ [29], низок по сравнению с ожидаемыми для скелетных мышц позвоночных, которые колеблются от 20 до 28 процентов при оптимальных скоростях сокращения [50].Кажется удивительным, что эволюция функции летных мышц у корелл и других птиц была ограничена существенно более низкой эффективностью. Хотя частота взмахов крыльев незначительно меняется в зависимости от скорости полета (10% у корелл), величина деформации грудного пучка изменяется в виде неглубокой U-образной формы, параллельно изменениям силы грудной мышцы [29], что приводит к соотношению общей силы мышц и скорости. что наблюдается у корелл (). Хотя скорость деформации пучка зависит от скорости полета, обычно однородные сократительные свойства грудной мышцы в диапазоне скоростей полета [27,29] () отражают поразительно однородные характеристики типа волокон грудной мышцы птиц [21–23].Это контрастирует с гораздо большим изменением скорости укорочения пучка со скоростью бега, которое происходит в мышцах ног наземных животных [51–54].
6. Мышечная функция, связанная с контролем взлета, посадки и маневрирования в полете
Принимая во внимание, что грудная и супракоракоидная мышцы в основном отвечают за выработку механической энергии, необходимой для продолжительного взмахивающего полета у птиц, неясно, действует ли активность Эти большие летные мышцы модулируются для достижения маневренного поведения в полете или для того, чтобы задействовать меньшие внешние и внутренние мышцы крыла для регулировки ориентации крыла и формы крыла.Прошлые работы, основанные на записях трехмерной кинематики, силы мышц и ЭМГ, предполагают две возможности. У голубей [6,55] левая и правая грудные мышцы, по-видимому, демонстрируют разное время развития силы и величины, при этом фаза движения наружного крыла вниз опережает фазу поворота внутреннего крыла. В галах с розовой грудью [5] небольшая разница в кинематическом времени удара вниз или ЭМГ-активности грудной клетки была отмечена во время поворотов на 90 °. Вместо этого были доказательства дифференциальной активации левой и правой мышц двуглавой мышцы, при этом внутренняя часть двуглавой мышцы показывала более сильную активность, что свидетельствовало об увеличении сгибания локтя и уменьшении внутреннего размаха крыльев.Однако в обоих исследованиях более подробная кинематика формы и движения крыла во время этих маневров не была доступна из-за ограниченного разрешения систем анализа движения, используемых в то время. Будущие исследования выиграют от улучшенного кинематического разрешения во время полета с разворотом в сочетании с дальнейшим изучением сократительной асимметрии мышц левого и правого крыла.
В исследованиях голубей, взлетающих с возвышенной платформы, уровня полета и приземления на аналогичную площадку, измерения кинематики крыльев, тела и хвоста показывают незначительные изменения в движениях крыльев или хвоста относительно тела птицы [56].Вместо этого большинство изменений в глобальных ориентациях хвостовой части, крыла и плоскости хода крыла, которые определяют аэродинамические свойства взмаха птицы, достигаются за счет изменения наклона корпуса ( a ). Во время взлета голуби наклоняются вперед (голова опущена), наклоняя плоскость гребка в более вертикальную ориентацию, чтобы обеспечить повышенную тягу для ускорения после взлета и прыжка с жердочки. Во время приземления голубь отклоняется назад (поднимает голову), меняя плоскость удара на более горизонтальную ориентацию, чтобы помочь замедлить движение при приземлении.Изменения в общем угле наклона плоскости хода во время взлета и посадки значительно больше и меньше, соответственно, чем наблюдаемые при горизонтальном полете.
( a ) Изменения в плоскости хода крыла (SPA loc ) и угла наклона корпуса (в глобальном пространстве) голубя во время последовательных взмахов крыльев при взлете, среднем полете и посадке (адаптировано из Berg & Biewener) [56]). Справа показана сильная корреляция угла наклона плоскости гребка крыла с углом корпуса. Закрашенные треугольники, углы тела; закрашенные квадраты, SPA loc .(b ) Типичный образец in vivo записей напряжения мышц и активации (ЭМГ) внешних и внутренних мышц крыльев голубя во время взлета, уровня (прибл. 4,5 мс -1 ) и полета при посадке, соответствующем полету. аналогичная последовательность показана в ( a ) выше (адаптировано из Berg [57]).
Равномерное движение крыльев голубя относительно его тела во время взлета, горизонтального полета и приземления предполагает, что управление движением крыла и тела на этих ключевых этапах полета зависит от тонких сдвигов в аэродинамических и инерционных силах, создаваемых хвостом. и крылья относительно корпуса для управления углом наклона корпуса.Момент инерции тангажа птицы, хотя и превышает момент инерции крена, все же довольно мал. В результате небольшие сдвиги в ориентации чистой аэродинамической силы вызывают наблюдаемое ускорение по тангажу. У голубей смещение в направлении чистой аэродинамической силы должно составлять всего около 8 мм относительно его центра масс, чтобы получить наблюдаемый момент тангажа [56]. В соответствии с этим, не наблюдалось значительных различий в нервно-мышечной активации (ЭМГ) или в поведении сократительной деформации исследованных крыловых мышц ( b ) [57].Этот результат предполагает, что контроль ориентации тела и движения крыльев относительно тела не требует существенных изменений в активации летных мышц и сократительной функции. Вместо этого, очень маневренные тела многих птиц (моменты инерции малого тангажа, крена и рыскания) позволяют им достигать изменений в ориентации тела и крыла, которые обеспечивают быстрый резкий поворот или переход от взлета к посадке с небольшими изменениями. в нервно-мышечной функции, которую, вероятно, будет сложно идентифицировать.
7. Обсуждение и подведение итогов
Функция мышц в птичьем полете зависит от производства значительной механической работы, выполняемой с высокой скоростью. Хотя скелетные мышцы, как правило, обладают аналогичной способностью производить работу, ориентированную на массу тела, грудная мышца птиц хорошо подходит для выполнения работы с большими длинами экскурсий. Это предварительное условие для обеспечения полета, потому что крылья должны совершать большой размах во время движения вниз, чтобы обеспечить эффективную аэродинамическую подъемную силу. Грудная мышца достигает этого за счет относительно длинных пучков, которые укорачиваются на значительной части (до 42%) своей длины.Выбор времени для активации мышц в конце хода вверх также позволяет грудной мышце быстро развивать силу в почти изометрических условиях или в условиях растяжения. Это увеличивает работу, которую выполняет мышца при сокращении ().
Из-за большого размера и принципиальной роли в создании аэродинамической подъемной силы сократительная функция грудной мышцы птиц обеспечивает ценный показатель требований к мощности полета, основанный на измерениях ее производства силы, сократительной деформации и нервно-мышечной активации.Это контрастирует с множеством групп мышц конечностей бегущих животных, которые способствуют мышечной силе для движения. Тем не менее, функциональное исследование более широкого набора крыловых мускулов необходимо, чтобы понять, как управляются движения в полете, особенно во время маневрирования. Хотя гораздо меньшие мускулы крыла могут не вносить значительного вклада в механическую мощность, лежащую в основе полета, регулируя ориентацию и форму крыла, они могут изменять аэродинамические свойства крыла и, таким образом, влиять на то, как аэродинамические силы и сила перемещаются между крыльями для маневрирования. .
Неожиданный результат заключается в том, что смещения тела, хвоста и крыльев во время взлета, горизонтального и приземляющего полета голубей достигаются в основном за счет изменения шага всего тела, а не за счет изменений движения крыльев или хвоста относительно тела. сам. Остается неясным, в какой степени разворот достигается за счет левосторонней и правой асимметрии меньших мышц крыльев, действующих для «управления» птицей при повороте, в отличие от модуляции более крупных, производящих энергию грудных и / или надкоракоидных мышц.Существуют доказательства того, что оба набора мышц могут вносить вклад в необходимую аэродинамическую асимметрию, которая приводит к маневру при повороте. Низкие моменты инерции и высокая маневренность тела птиц означают, что левосторонняя и правосторонняя асимметрии при развороте или продольная асимметрия в создании аэродинамической силы во время взлета и посадки, вероятно, будут небольшими и их сложно идентифицировать.
Будущие исследования выиграют от улучшенной визуализации, которая позволит количественно определить подробные изменения формы, ориентации и движения крыла и связать их со временем и величиной активации мышц и, где это возможно, изменениями длины, силы и работы мышц.Эти измерения становятся все более трудными для более мелких мышц, расположенных дистальнее в крыле. В частности, для большинства мышц трудно получить измерения силы, что затрудняет возможность оценки мышечной силы и производительности в отношении маневрирования в полете. В случае, когда мышцы слишком малы или сила не может быть записана напрямую, in vitro, или in situ, измерения мышечной силы [29] могут сыграть важную роль для оценки сократительных свойств и роли мышцы в полете. .Замечательная способность птиц летать с разным диапазоном скоростей, часто маневрируя в сложных условиях, делает понимание нейромышечных и аэродинамических характеристик этого поведения в полете значительным интересом для физиологов, биомехаников и авиационных инженеров.
Точно так же требования к аэродинамической и метаболической мощности для полета представляют значительный интерес для птичьих и эволюционных экологов, интересующихся стратегиями, которые птицы используют для кормления и миграции, чтобы обеспечить успешную жизненную историю.По этой причине дополнительные данные о свободном полете о метаболизме птиц, характерных скоростях полета и поведении необходимо увязать с дополнительными экспериментальными оценками энергетического метаболизма полета и функции опорно-двигательного аппарата. В то время как квазистатические аэродинамические модели могут дать приблизительную оценку затрат на полет, важность нестационарных аэродинамических эффектов для затрат на мощность полета в настоящее время хорошо осознается, и ее нельзя игнорировать. Таким образом, необходимо дополнительное моделирование и экспериментальные исследования, которые стремятся дать улучшенные измерения мышечной функции и аэродинамической выходной мощности.
Благодарности
Автор благодарит своих многочисленных бывших студентов, постдоков и сотрудников за веселую и захватывающую работу по изучению функции мышц птиц в отношении летных характеристик. Он также благодарит г-на Педро Рамиреса за его экспертную и преданную помощь в уходе за птицами. Большая часть работы автора финансируется Национальным научным фондом, последний раз — IOS-0744056.
Ссылки
1. Райнер Дж. В. М. 1988. Форма и функции в птичьем полете.В современной орнитологии, т. 5 (изд. Джонстон Р. Ф.), стр. 1–66 Нью-Йорк, Нью-Йорк: Plenum Press [Google Scholar] 4. Диал К. П., Гейтси С. М. 1993. Нервно-мышечный контроль и кинематика крыльев и хвоста при маневренном полете. Являюсь. Zool. 33, 5 [Google Scholar] 5. Хедрик Т. Л., Ашервуд Дж. Р., Бивенер А. А. 2007. Полет розогрудого какаду на низкой скорости ( Eolophus roseicapillus ). II: Инерционная и аэродинамическая переориентация. J. Exp. Биол. 210, 1912–1924 10.1242 / jeb.002063 (DOI: 10.1242 / jeb.002063) [PubMed] [CrossRef] [Google Scholar] 6. Уоррик Д. Р., Диал К. П. 1998. Кинематические, аэродинамические и анатомические механизмы в медленном маневренном полете голубей. J. Exp. Биол. 201, 655–672 [PubMed] [Google Scholar] 9. Александр Р. М. 2003. Принципы передвижения животных. Принстон, Нью-Джерси: Издательство Принстонского университета [Google Scholar] 10. Бивенер А. А. 2003. Передвижение животных. Оксфорд, Великобритания: Oxford University Press [Google Scholar] 11. Марш Р. Л. 1999. Как мышцы справляются с реальными нагрузками: влияние траектории длины на работу мышц.J. Exp. Биол. 202, 3377–3385 [PubMed] [Google Scholar] 12. Бивенер А. А. 1998. Функция мышц in vivo : конструкция мышц, используемых в качестве пружин, по сравнению с мышцами, используемыми для выработки механической энергии. Являюсь. Zool. 38, 703–717 [Google Scholar] 13. Александр Р. М. 1988. Упругие механизмы в движении животных. Кембридж, Великобритания: Издательство Кембриджского университета [Google Scholar] 14. Бивенер А. А., Робертс Т. Дж. 2000. Вклад мышц и сухожилий в экономию силы, работы и упругой энергии: сравнительная перспектива.Exer. Спорт. Sci. Ред. 28, 99–107 [PubMed] [Google Scholar] 16. Джордж Дж. К., Бергер А. Дж. 1966. Птичья миология. Нью-Йорк, Нью-Йорк: Academic Press [Google Scholar] 17. Райков Р. 1985. Опорно-двигательная система. В книге «Форма и функции птиц» (ред. Кинг А. С., Маклелланд Дж.), Стр. 57–147, Лондон, Великобритания: Academic Press [Google Scholar] 18. Пур С. О., Эшкрофт А., Санчес-Хайман А., Гослоу Г. Э., мл. 1997. Сократительные свойства m. supracoracoideus у голубя и скворца: случай вращения плечевой кости вдоль длинной оси.J. Exp. Биол. 200, 2987–3002 [PubMed] [Google Scholar] 20. Каплан С. Р., Гослоу Г. Э., младший 1989. Нервно-мышечная организация грудной мышцы (pars thoracicus) голубя ( Columba livia ): значение для моторного контроля. Анат. Рек. 224, 426–43010.1002 / ar.1092240311 (doi: 10.1002 / ar.1092240311) [PubMed] [CrossRef] [Google Scholar] 21. Россер Б. В. К., Джордж Дж. С. 1986. Грудная мышца птиц: гистохимическая характеристика и распределение типов мышечных волокон. Жестяная банка. J. Zool. 64, 1174–118510.1139 / z86-176 (doi: 10.1139 / z86-176) [CrossRef] [Google Scholar] 26. Диал К. П., Бивенер А. А. 1993. Сила и мощность грудных мышц во время различных режимов полета у голубей ( Columba livia ). J. Exp. Биол. 176, 31–54 [Google Scholar] 28. Моррис С. Р., Аскью Г. Н. 2010. Выходная механическая мощность грудной мышцы кореллы ( Nymphicus hollandicus ): траектория длины и активности мышцы in vivo и модели активности и их значение для модуляции мощности.J. Exp. Биол. 213, 2770–278010.1242 / jeb.035691 (doi: 10.1242 / jeb.035691) [PubMed] [CrossRef] [Google Scholar] 29. Моррис С. Р., Аскью Г. Н. 2010. Сравнение между потребностями в механической мощности для полета, оцененными с использованием аэродинамической модели, и мышечными характеристиками in vitro и у кореллы ( Nymphicus hollandicus, ). J. Exp. Биол. 213, 2781–278710.1242 / jeb.035709 (doi: 10.1242 / jeb.035709) [PubMed] [CrossRef] [Google Scholar] 31. Бивенер А. А., Корнинг В. Р., Тобальске Б. Т.1998. In vivo поведение силы-длины грудной мышцы во время горизонтального полета у голубей ( Columba livia ). J. Exp. Биол. 201, 3293–3307 [PubMed] [Google Scholar] 32. Соман А., Хедрик Т. Л., Бивенер А. А. 2005. Региональные особенности деформации грудного пучка у голубя Columba livia во время горизонтального полета. J. Exp. Биол. 208, 771–78610.1242 / jeb.01432 (doi: 10.1242 / jeb.01432) [PubMed] [CrossRef] [Google Scholar] 33. Хедрик Т. Л., Тобальске Б. В., Бивенер А. А.2003. Как кореллы ( Nymphicus hollandicus ) регулируют мощность грудной мышцы в зависимости от скорости полета. J. Exp. Биол. 206, 1363–137810.1242 / jeb.00272 (doi: 10.1242 / jeb.00272) [PubMed] [CrossRef] [Google Scholar] 34. Уильямсон М. Р., Диал К. П., Бивенер А. А. 2001. Производительность грудных мышц во время подъема и медленного горизонтального полета у крякв ( Anas platyrynchos ). J. Exp. Биол. 204, 495–507 [PubMed] [Google Scholar] 35. Либер Р. Л. 1992. Структура и функция скелетных мышц.Балтимор, Мэриленд: Уильямс и Уилкинс [Google Scholar] 36. МакМахон Т. А. 1984. Мышцы, рефлексы и движение. Принстон, Нью-Джерси: Издательство Принстонского университета [Google Scholar] 37. Пенникуик К. Дж., Хеденстрём А., Розен М. 2000. Горизонтальный полет ласточки ( Hirundo rustica ), наблюдаемый в аэродинамической трубе, с помощью нового метода прямого измерения механической мощности. J. Exp. Биол. 203, 1755–1765 [PubMed] [Google Scholar] 38. Тобальске Б., Диал К. П. 1996. Кинематика полета черноклювых сорок и голубей в широком диапазоне скоростей.J. Exp. Биол. 199, 263–280 [PubMed] [Google Scholar] 39. Тобальске Б. В., Уоррик Д. Р., Кларк К. Дж., Пауэрс Д. Р., Хедрик Т. Л., Хайдер Г. А., Бивенер А. А. 2007. Трехмерная кинематика полета колибри. J. Exp. Биол. 210, 2368–238210.1242 / jeb.005686 (doi: 10.1242 / jeb.005686) [PubMed] [CrossRef] [Google Scholar] 41. Диал К. П., Бивенер А. А., Тобальске Б. В., Уоррик Д. Р. 1997. Прямая оценка выходной механической мощности птицы в полете. Nature 390, 67–7010.1038 / 36330 (DOI: 10.1038 / 36330) [CrossRef] [Google Scholar] 43.Пенникуик К. Дж. 1968. Требования к мощности для горизонтального полета голубя Columba livia . J. Exp. Биол. 49, 527–555 [Google Scholar] 44. Пенникуик К. Дж. 1989. Летные характеристики птицы. Практическое руководство по расчету. Оксфорд, Великобритания: Oxford University Press [Google Scholar] 45. Райнер Дж. В. М. 1999. Оценка силовых кривых летающих позвоночных. J. Exp. Биол. 202, 3449–3461 [PubMed] [Google Scholar] 47. Usherwood J. R., Hedrick T. L., McGowan C. P., Biewener A. A. 2005. Карты динамического давления для крыльев и хвостов голубей при медленном, хлопающем полете и их энергетические последствия.J. Exp. Биол. 208, 355–36910.1242 / jeb.01359 (doi: 10.1242 / jeb.01359) [PubMed] [CrossRef] [Google Scholar] 48. Связка М. В., Хансен К. С., Диал К. П. 2007. Различается ли соотношение между скоростью метаболизма и скоростью полета у геометрически схожих птиц разной массы? J. Exp. Биол. 210, 1075–108310.1242 / jeb.02727 (doi: 10.1242 / jeb.02727) [PubMed] [CrossRef] [Google Scholar] 49. Моррис С. Р., Нельсон Ф. Э., Аскью Г. Н. 2010. Требования к метаболической мощности для полета и оценки эффективности летных мышц у кореллы ( Nymphicus hollandicus ).J. Exp. Биол. 213, 2788–279610.1242 / jeb.035717 (doi: 10.1242 / jeb.035717) [PubMed] [CrossRef] [Google Scholar] 50. Волледж Р. К., Куртин Н. А., Хомшер Э. 1985. Энергетические аспекты сокращения мышц. Лондон, Великобритания: Academic Press [PubMed] [Google Scholar] 51. Дейли М. А., Бивенер А. А. 2003. Динамика мышечной силы и длины во время горизонтального и наклонного движения: сравнение показателей in vivo и двух разгибателей голеностопного сустава цесарок. J. Exp. Биол. 206, 2941–295810.1242 / jeb.00503 (DOI: 10.1242 / jeb.00503) [PubMed] [CrossRef] [Google Scholar] 52. Габальдон А. М., Нельсон Ф. Э., Робертс Т. Дж. 2004. Механическая функция двух разгибателей голеностопного сустава у диких индеек: переход от выработки энергии к поглощению энергии во время бега на наклонной поверхности по сравнению с бегом на спуске. J. Exp. Биол. 207, 2277–228810.1242 / jeb.01006 (doi: 10.1242 / jeb.01006) [PubMed] [CrossRef] [Google Scholar] 53. Гиллис Г. Б., Бивенер А. А. 2001. Функция мышц задних конечностей в зависимости от скорости и походки: in vivo, напряжение и активация в разгибателях бедра и колена крысы ( Rattus norvegicus ).J. Exp. Биол. 204, 2717–2731 [PubMed] [Google Scholar] 54. Гиллис Г. Б., Флинн Дж. П., МакГиган П., Бивенер А. А. 2005. Паттерны напряжения и активации мышц бедра коз при походке во время ровного передвижения. J. Exp. Биол. 208, 4599–461110.1242 / jeb.01940 (doi: 10.1242 / jeb.01940) [PubMed] [CrossRef] [Google Scholar] 55. Уоррик Д. Р., Диал К. П., Бивенер А. А. 1998. Производство асимметричной силы при маневрировании голубей. Auk 115, 916–928 [Google Scholar] 57. Берг А. М. 2010.Кинематика, аэродинамика и нервно-мышечные функции полета птиц: взлет и посадка, подъем и спуск. Докторская диссертация, Гарвардский университет, Кембридж, Массачусетс [Google Scholar]Диссекция куриного крылышка
Диссекция куриного крылаРассечение куриного крылышка
Может показаться, что передние конечности позвоночных человека, птицы и летучей мыши не имеют много общего, но более тщательное изучение анатомии показывает, что одни и те же кости использовались для выполнения множества различных работ у каждого из этих видов.Эволюция не планирует наперед. Он может работать только с тем, что есть в наличии. Цветовая кодировка показывает, каким образом костные элементы были изменены у каждого вида. Вы можете подумать, что два крыла будут более похожи по конструкции из-за выполняемой ими работы, но на самом деле крыло птицы и человеческая рука больше похожи по внутренней части. Однако птичья «рука» сокращена, тогда как у летучей мыши и человеческой руки больше общего. Посмотрите, сможете ли вы найти похожее изображение дельфина или плавника кита для другого сравнения. Какие прогнозы вы можете сделать, прежде чем увидеть внутреннюю анатомию?
Анатомия передних конечностей позвоночных
Просмотрите следующие термины в Википедии:
Анатомия куриного крыла : Посмотрите следующее видео в качестве общего руководства по процедуре.
Правила техники безопасности:
- Поскольку вы будете иметь дело с сырой курицей, на поверхности которой могут быть микробы, вымойте руки водой с мылом до и после этой процедуры.
- При желании вы можете использовать резиновые перчатки или голые руки, если хотите.
- Чтобы уменьшить вероятность порезаться или порезаться, вам понадобятся только ножницы и пинцет.
- По завершении вымойте все оборудование горячей мыльной водой.
- Выбросьте куриное крылышко и его части в конце этого упражнения.
Осмотреть целое крыло
- Удерживая крыло за заплечик и кончик крыла, полностью выдвиньте крыло.
- Отметьте места соединений в местах изгиба крыла.
- Обратите внимание на направления изгиба шарниров.
- Пальцами нащупайте кости и мышечные пучки под кожей.
- Отметьте места, где перья были вставлены в кожу.
Куриное крылышко с прикрепленной кожей и мышцами
Снимаем только кожу
- Ножницами удалите как можно больше кожи, разрезая по линии костей.
- Будьте осторожны, чтобы не порезать мышцы, связки и сухожилия.
- Промокните весь жир бумажным полотенцем и удалите его, чтобы обнажить анатомические структуры.
- «Манус» происходит от того же корня, что и «руководство». Какая эквивалентная часть на вашем теле?
- На этом рисунке обозначен локтевой сустав. Где лучезапястный сустав?
- Какая часть вашей руки соответствует «alula»?
Кости куриного крыла
Осмотреть мягкие ткани
- Найдите два мышечных пучка, которые сгибают и распрямляют нижнюю часть крыла ниже локтевого сустава, по очереди потянув за каждую мышцу.
- Обратите внимание, что эти мышцы находятся напротив друг друга. Когда один сжимается, другой удлиняется.
- Отметьте точки, в которых эти мышцы прикрепляются к кости, и осторожно удалите их одну за другой.
- Можно ли найти хрящ в суставе плечевой кости и лучевой / локтевой кости?
- Вы заметили, что мышцы, двигающие нижнее крыло, находятся в верхнем крыле? Где мышцы, контролирующие верхнее крыло курицы?
- Сможете ли вы найти связки, которые скрепляют кости?
- Согните руку в локте и крепко возьмите ее.Обратите внимание на то, что ваша двуглавая мышца укорачивается. Теперь вытяните руку и почувствуйте, как удлиняется бицепс.
- Посмотрите на свою руку. Где мышцы, которые двигают пальцы? Подсказка: надавите на запястье и предплечье, шевеля пальцами.
- Почему большой палец так важен для захвата предметов? Попробуйте взять что-нибудь, не используя большой палец.
- Поднимайте и опускайте тяжелый предмет, как учебник, сгибая руку в локте. Попросите партнера почувствовать, какие мышцы действуют во время подъема и опускания.
Контрольные вопросы :
- В чем разница между связкой и сухожилием?
- В чем разница между сгибанием и разгибанием?
- В чем разница между приведением и отведением?
- Что означает, когда мы говорим, что мышцы находятся в оппозиции?
- Выполняет ли мышца какое-либо действие, когда она сокращается или расслабляется?
- Для чего нужен хрящ?
- Изучите различные типы суставов.Опишите три или более типов, включая допустимый диапазон движений.
- Что означает противопоставление большого пальца человека? Чему это противопоставляется?
- Исследуйте переднюю крестообразную связку или ПКС. Почему он обычно повреждается при травмах в футболе и на лыжах?
- Исследуйте ахиллово сухожилие. Для чего нужны сухожилия? Что делают сухожилия в руке?
Дополнительный кредит :
- Используя резиновые ленты или эластичные шнуры в качестве мышц и палочек для мороженого в качестве костей, с клейкой лентой или петлями в качестве суставов или с другими подходящими материалами создайте рабочую модель руки или кисти человека.
BBC Science & Nature — Human Body and Mind
Два волокна: в мышцах есть два разных волокнаМедленно сокращающиеся мышечные волокна: сокращаются медленно, но продолжаются в течение длительного времени
Быстро сокращающиеся мышечные волокна: сокращаются быстро, но быстро усталость
Выносливость или скорость
Медленно сокращающиеся мышечные волокна хороши для тренировок на выносливость, таких как бег на длинные дистанции или езда на велосипеде. Они могут работать долго, не уставая. Быстро сокращающиеся мышцы хороши для быстрых движений, таких как прыжки, чтобы поймать мяч, или бег на автобусе.Они быстро сокращаются, но быстро устают, так как потребляют много энергии.
Большинство ваших мышц состоит из смеси медленных и быстро сокращающихся мышечных волокон. Но камбаловидная мышца голени и мышцы спины, участвующие в поддержании осанки, содержат в основном медленно сокращающиеся мышечные волокна. А мышцы, которые двигают ваши глаза, состоят из быстро сокращающихся мышечных волокон.
Темно-белое мясо
У цыплят также есть быстро и медленно сокращающиеся мышцы. Темное мясо, как и куриные окорочка, в основном состоит из медленно сокращающихся волокон.Белое мясо, как и куриные крылышки и грудка, в основном состоит из быстро сокращающихся мышечных волокон. Курицы используют свои ноги для ходьбы и стояния, что они и делают большую часть времени. Это не требует много энергии. Они используют свои крылья для коротких взлетов. Это требует много энергии, и задействованные мышцы очень быстро устают.
Медленно сокращающиеся и быстро сокращающиеся мышечные волокна вырабатывают энергию по-разному.
Мышцы, содержащие много медленных волокон, имеют красный цвет, потому что они содержат много кровеносных сосудов.Медленно сокращающиеся мышечные волокна зависят от обильного поступления насыщенной кислородом крови, поскольку они используют кислород для выработки энергии для сокращения мышц.
Быстро сокращающиеся мышечные волокна не используют кислород для производства энергии, поэтому им не нужно такое обильное кровоснабжение. Вот почему быстро сокращающиеся мышцы светлее, чем мышцы, содержащие много медленных мышечных волокон.
Быстро сокращающиеся мышечные волокна могут очень быстро производить небольшое количество энергии, тогда как медленно сокращающиеся мышцы могут медленно производить большое количество энергии.
Вернуться к началу
Крылья ангела — Performance Medicine
Танцовщица постоянно работает над идеальной осанкой и элегантными линиями.
Взмах лопаток — обычная проблема танцоров, их чаще называют крылышками ангела или куриными крылышками (рис. А).
Это изменение осанки часто вызвано слабостью лопатных мышц и мышц, стабилизирующих плечевой пояс.Хотя ангельские крылья часто не вызывают боли, важно решить проблему раньше, чем позже, поскольку это может увеличить вероятность развития чрезмерного использования плечевого сустава, такого как удар плеча или нестабильность. Для танцоров, которые несут тяжесть руками (например, танцоры-мужчины, поднимающие своих партнеров, танцоры-танцоры, выполняющие акробатические трюки и трюки), очень важно, чтобы мышцы плеча были сильными, чтобы предотвратить травмы.
Основная биомеханика плеча
Плечевой пояс состоит из акромиально-ключичного сустава, лопатко-грудного сустава и плечевого сустава.Плечевой сустав, шаровидный сустав, образован неглубокой суставной поверхностью («впадиной») лопатки, в которой находится головка плечевой кости («шар»). Это позволяет плечу двигаться в больших дугах движений, но именно с этой повышенной подвижностью суставов приносится в жертву стабильность. Поэтому важно укрепить мышцы, которые стабилизируют лопатки на грудной клетке, чтобы создать стабильную основу для движения плечевого сустава, а также исправить куриные крылышки или крылья ангела.
Плечевой пояс сложен и поэтому требует как стабилизирующих, так и создающих силу мышц, чтобы гарантировать, что он функционирует на высоком уровне, необходимом для танцора. К крупным мышцам, которые производят сильные и мощные движения, относятся грудные, широчайшие мышцы спины и трапеции (рис. B). Меньшие, стабилизирующие и контролирующие мышцы включают вращающую манжету (надостной, подостной, подлопаточной, большой круглой) и переднюю зубчатую мышцу (рис. C). Крылья ангела часто появляются при дисбалансе набора мышц и активации этих больших и малых групп мышц.
Какие упражнения мне делать?
Есть много способов укрепить стабилизирующие мышцы лопатки.
Ниже приведены некоторые базовые упражнения, которые помогут исправить ваши ангельские крылья. Щелкните здесь, чтобы увидеть наши фотографии упражнений.
I Если вы испытываете боль или беспокоитесь, что эти упражнения не для вас, важно проконсультироваться с вашим физиотерапевтом, который может предложить вам конкретную программу.
Ретракция лопатки лежа на животе
- Лягте на живот лицом к полу.Вы можете свернуть полотенце и подложить его себе под лоб, чтобы вам было удобнее.
- Держите руки по бокам и медленно поднимайте лопатки вверх, обхватывая ими заднюю часть грудной клетки.
- Медленно опустите их обратно.
- Повторите два или три подхода по 10. (Рисунок d)
Прочность поворотной манжеты
Внутреннее вращение
- Привяжите низкопрочную (обычно желтую, красную или зеленую) ленту для упражнений к чему-нибудь прочному.
- Встаньте или сядьте перпендикулярно ленте для упражнений и возьмитесь за руку, находящуюся ближе всего к ней (лента должна находиться с внешней стороны вашего тела).
- Установите лопатку в нужное положение, повернув ее вверх и назад (аналогично приведенному выше упражнению). Это создаст хорошую лопаточную основу для движения гельно-плечевого сустава.
- Медленно протяните ленту через тело, сохраняя положение лопатки.
- Медленно вернитесь в исходное положение.
- Повторите два или три подхода по 10. (Рисунок e)
Внешнее вращение
- После того, как вы закончите, продолжайте удерживать эластичную ленту той же рукой и поверните ее на 180 °, чтобы смотреть в другую сторону (лента теперь будет поперек вашего тела).
- Повторите описанное выше упражнение, на этот раз оттягивая ремешок от тела. ( Рисунок f)
Ряд упражнений с лентами
- Привяжите ленту для упражнений к чему-нибудь прочному так, чтобы оба конца были свободны.