Физиология езды на велосипеде (и физических нагрузок в общем) | Велотавр | Велотавр
Как всем известно, ноги наши крутят педали — ибо мы велотавры. Что же такого напихала природа в наши ноги, что они работают (а иногда и не работают)?
Сразу отмечу, что мышцы можно делить на виды по разным признакам. Для простоты условимся делить их на «быстрые» и «медленные». Скелетная мышца состоит из двух видов волокон. Процентное соотношение их обуславливается генетикой и наследственностью. Обычно медленных волокон в пучке мышц около 40%. У стайеров (бегунов на длинные дистанции) же процент этих волокон доходит до 75%!
Биохимия этих волокон (упрощаю схему для того, чтобы запоминалось).
Быстрые мышцы включаются тогда, когда необходимо совершить взрывную работу в максимально короткий срок. Финальный спурт, подъем в гору, — все это в большинстве своем делают быстрые мышцы. Энергию для них дает гликоген — «животный крахмал», переработанные углеводы. Гликоген накапливается про запас в мышцах и в печени.
Медленные мышцы работают на жирах. Жира у нас много (а у некоторых очень много ), поэтому медленные мышцы работают долго и не «устают». Мы же можем ходить целый день? Все потому, что работают медленные мышцы. Но и здесь есть подводные камни. Разложение жиров требует большого количества воды и кислорода. И если кровеносная система не будет успевать подавать все необходимые ферменты в необходимом количестве — стартуют быстрые мышцы.
Теперь введем два понятия — анаэробный и аэробный режимы.
Анаэробный (бескислородный) режим — режим, в котором работают преимущественно быстрые мышцы. Для их работы кислорода надо намного меньше (углеводы для разложения не требуют такого количества кислорода, как жиры).
Аэробный (кислородный) режим — режим, в котором работают преимущественно медленные мышцы. Возможен только когда сердце и легкие успевают снабдить клетки кислородом и водой.
Бытует мнение, что накопленная молочная кислота вызывает болевые ощущения. Споров по этому поводу много. Доводов тоже. Не буду вдаваться в дискуссии, отмечу лишь что болевые ощущения появляются только после анаэробного режима.
Итак, будем подводить итоги. Если вы хотите ездить быстро — повышайте свой анаэробный порог методом интервальных тренировок (чередуйте в одной тренировке работу на высоком пульсе и восстановление на низком). Если вы хотите ездить далеко — закрепляйте выносливость организма — продолжительные тренировки на небольшом пульсе. Очень полезен для тренировок пульсометр — позволяет объективно оценивать нагрузку.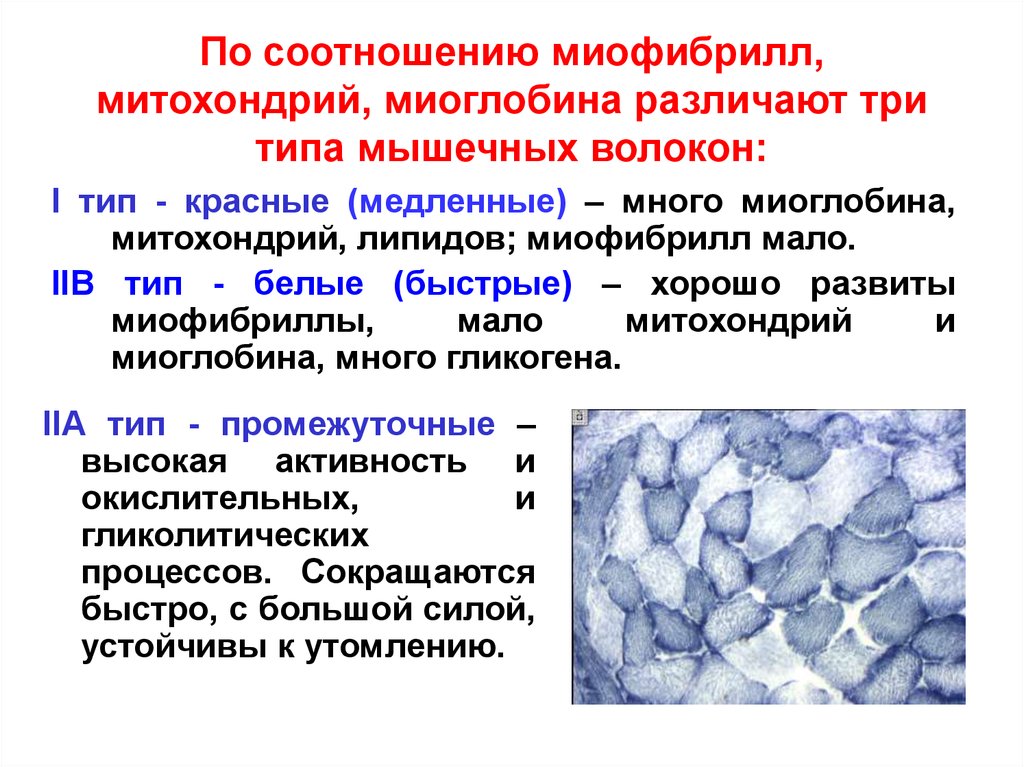
И еще небольшой совет. Если после похода или тренировки вы чувствуете слабость и боль в мышцах — не лежите. Проверено, что самым быстрым способом восстановления является аэробная нагрузка в течение 20-30 минут. Не поленитесь проехаться неспешно на велосипеде полчаса — поверьте, ваше самочувствие намного улучшится!
В чём отличия медлнных (ST) и быстрых (FT-A и FT-B) мышечных волокон
Типы мышц, их строение являются той причиной, по которой один атлет может бежать быстрее и наращивать мышечную массу легче других, а другой — способен бежать длительное время без признаков утомления.
Такую разницу создают процессы, которые происходят в мышечной ткани. Важно понимать их, например, для того чтобы выстроить правильную тренировочную программу, которая подойдет для конкретно выбранного спортсмена.
почему сокращаются мышцы?
- Пресинаптическое окончание.
- Сарколемма.
- Синаптический пузырек.
- Никотиновый ацетилхолиновый рецептор.
- Митохондрия
К волокнам скелетных мышц подходят толстые нервные волокна, которые отходят от передних отделов спинного мозга. После попадания в мышцу, каждое нервное волокно делится и снабжает своими разветвлениями до нескольких сотен мышечных волокон.
Соединение нерва и мышечного волокна образует так называемый синапс, или нервно-мышечное соединение (причем на каждом мышечном волокне формируется только один такой синапс). Под влиянием нервного сигнала возникает так называемый потенциал действия, который распространяется от спинного мозга по нервам к мышце и синапсу.
То, как скелетная мускулатура будет адаптироваться к повторяющимся стимулам, в большей степени зависит от внутренних характеристик самой мышцы. Именно типы мышечных волокон вносят наибольший вклад в возможность выполнения спортсменом той или иной тренировочной программы.
Типы мышечных волокон
У людей выделяют три типа мышечных волокон:
- Медленносокращающиеся (slow-twitch, ST или I тип) волокна характеризуются медленным временем сокращения, а также большой сопротивляемостью усталости. В своей структуре эти волокна имеют маленький мотонейрон и диаметр нервного волокна, высокую плотность митохондрий и капилляров, большое содержание миоглобина.
Этот тип волокон имеет небольшое количество креатин фосфата — высокоэнергетического субстрата, необходимого для быстрого, взрывного движения, — а значит, эти волокна не способны сокращаться быстро.
Функционально, ST-волокна используются при аэробной активности, не требующей большого приложения силы, например — ходьба и поддержание позы.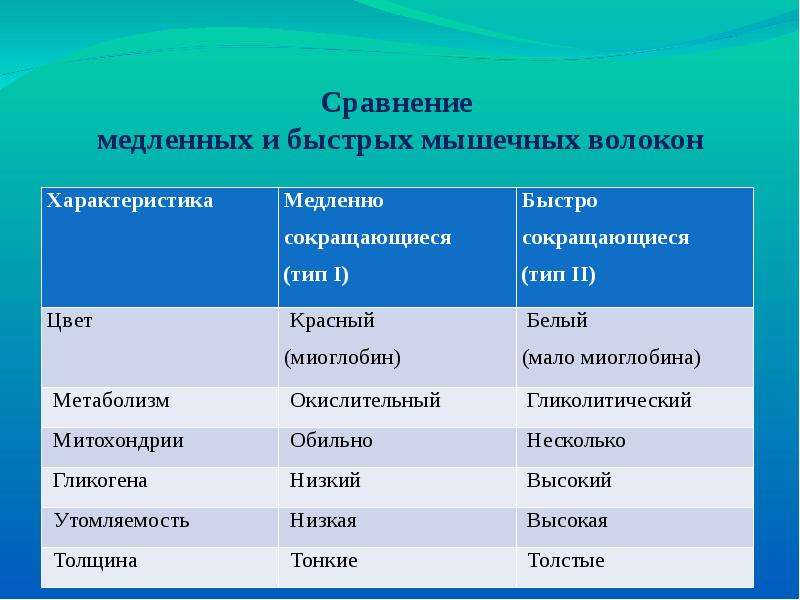 Большая часть повседневной активности задействует именно ST-волокна.
Большая часть повседневной активности задействует именно ST-волокна.
- Быстросокращающиеся (fast-twitch, FT или II тип) волокна характеризуются быстрым сокращением и низкой сопротивляемостью усталости. Разница в скорости сокращения, от которой произошло разделение волокон, может быть отчасти объяснена скоростью выделения кальция из саркоплазматического ретикулума (место в клетке, где хранится кальций), а также активностью фермента, который расщепляет АТФ внутри головки миозина (один из сократительных белков). Обе эти характеристики быстрее и в большей степени присутствуют в FT-волокнах.
Сами FT-волокна делятся на два типа: тип А (FT-A или IIA тип) и тип В (FT-B или IIB тип).
- Волокна FT-A имеют умеренную сопротивляемость усталости и представляют собой переходный тип между медленносокращающимися волокнами и волокнами типа FT -B. Функционально они используются при длительной анаэробной активности с относительно большой продукцией силы, например, беге на 400 метров.

- С другой стороны, быстросокращающиеся волокна типа B, очень чувствительны к усталости и используются для коротких анаэробных нагрузок с большой продукцией силы, такие как: спринтерские забеги, бег с барьерами, прыжки. Эти волокна также способны продуцировать больше энергии, чем ST волокна.

Краткие характеристики волокон приведены в таблице ниже.
| Тип волокна | Медленносокр. (ST) | Быстросокр. тип А (FT—A) | Быстросокр. тип B (FT—b) |
| Время сокращения | Медленное | Быстрое | Очень быстрое |
| Размер мотонейрона | Маленький | Большой | Очень большой |
| Сопротивляемость усталости | Высокая | Средняя | Низкая |
| Активность | Аэробная | Длительная анаэробная | Короткая анаэробная |
| Продукция силы | Низкая | Высокая | Очень высокая |
| Плотность митохондрий | Высокая | Высокая | Низкая |
| Плотность капилляров | Высокая | Средняя | Низкая |
| Окислительная способность | Высокая | Высокая | Низкая |
| Гликолитическая емкость | Низкая | Высокая | Высокая |
| Преимущественный запас топлива | Триглицериды (жиры) | Креатинфосфат, гликоген | Креатинфосфат, гликоген |
При конкретной скорости движения, количество продуцируемой силы определяется типом вовлеченного в движение мышечного волокна. Во время динамического сокращения, когда волокно укорачивается или удлиняется, быстросокращающиеся волокна продуцируют больше силы, чем медленносокращающиеся волокна.
Во время динамического сокращения, когда волокно укорачивается или удлиняется, быстросокращающиеся волокна продуцируют больше силы, чем медленносокращающиеся волокна.
В условиях, когда длина мышечного волокна не изменяется при сокращении, ST-волокна продуцируют столько же силы, как и FT-волокна. Разница в продукции силы наблюдается только при активном сокращении, когда изменяется длина волокна.
При конкретной скорости движения, сила, продуцируемая мышцей, возрастает с повышением содержания FT-волокон, и наоборот, при конкретной продукции силы, скорость повышается при увеличении количества FT-волокон.
Существует большой разброс в процентном соотношении волокон у атлетов. Например, хорошо известно, что участвующие в соревнованиях на выносливость имеют большее содержание медленносокращающихся волокон, тогда как спринтеры и прыгуны имеют больше быстросокращающихся волокон.
Больший процент FT-волокон у спринтеров позволяет им продуцировать большую силу и мощность, чем у атлетов с повышенным содержанием ST-волокон.
Разница в составе мышц у атлетов подняла вопрос о том, наследуется ли такой состав генетически или он может быть изменен с помощью тренировок. Исследования, проведенные на близнецах, показали, что в основном состав мышц и процент содержания в них разных типов волокон определяется генетически.
Тем не менее, есть ряд данных, доказывающих, что как структура, так и метаболическая емкость мышечных волокон может изменяться в ответ на различные типы тренировок.
Вовлечение мышечных волокон в работу
Мышца продуцирует силу путем вовлечения так называемых моторных единиц — группы мышечных волокон, которую «обслуживает» одно двигательное нервное окончание. Во время произвольного изометрического и концентрического сокращения, обычный порядок вовлечения моторных единиц контролируется их размерами — это состояние известно под названием «принцип размера».
Маленькие моторные единицы, содержащие медленносокращающиеся мышечные волокна, имеют наименьший порог активизации, т. е. для их активизации достаточно самого слабого стимула, поэтому они вовлекаются первыми. Потребность в выработке большей силы удовлетворяется вовлечением более крупных моторных единиц.
е. для их активизации достаточно самого слабого стимула, поэтому они вовлекаются первыми. Потребность в выработке большей силы удовлетворяется вовлечением более крупных моторных единиц.
Самые большие моторные единицы, содержащие быстросокращающиеся волокна типа B имеют наивысший порог активизации, и поэтому вовлекаются последними. Вне зависимости от интенсивности работы, первыми в нее вовлекаются медленносокращающиеся волокна.
Если интенсивность работы низкая, то медленносокращающиеся волокна остаются единственными, вовлеченными в нее. Если интенсивность работы высокая, например, подъем тяжелого веса, или интервальная работа на стадионе, первыми вовлекаются медленносокращающиеся волокна, затем подключаются быстросокращающиеся волокна типа A, а затем, при необходимости, волокна типа B.
Существуют данные о том, что принцип размера может быть нарушен или даже полностью изменен во время некоторых типов движений, особенно тех, которые содержат эксцентрические (с удлинением мышцы) компоненты. При этом быстросокращающиеся волокна могут активизироваться раньше медленносокращающихся.
При этом быстросокращающиеся волокна могут активизироваться раньше медленносокращающихся.
Определение типа волокон
Поскольку единственным способом напрямую определить состав мышечного волокна у атлета является проведение биопсии мышцы (прямой метод), то некоторые исследования попробовали определить состав мышечного волокна непрямым методом, путем выявления взаимосвязей между различными свойствами типа волокна и состава волокон мышц.
Интересные данные, полученные в этих исследованиях, показывают значимую взаимосвязь между содержанием быстросокращающихся волокон и мышечной силой или мощностью.
Непрямой метод, который можно использовать для определения состава волокон мышц, состоит в определении максимального веса, который спортсмен может поднять всего один раз.
После этого производится максимальное количество повторов с весом в 80% от максимального. Если общее количество повторений меньше семи, скорее всего мышцы более чем на 50% состоят из FT-волокон. Если же удается сделать двенадцать и более повторов, скорее всего, мышца более чем на 50% состоит из ST-волокон. Если число повторений между 7 и 12 — скорее всего, мышцы состоят поровну из FT и ST-волокон.
Если же удается сделать двенадцать и более повторов, скорее всего, мышца более чем на 50% состоит из ST-волокон. Если число повторений между 7 и 12 — скорее всего, мышцы состоят поровну из FT и ST-волокон.
Поскольку подъем веса вовлекает большое количество групп, этот метод не работает при определении состава изолированных мышц, а только мышечных групп.
Для определения состава волокон отдельной мышцы может потребоваться игольчатая биопсия интересующей мышцы. Другим непрямым методом, который можно использовать, является участие в различных соревнованиях. Доминантные волокна можно выявить, исходя из успеха в определенных соревнованиях, что позволит в дальнейшем развивать именно эти способности мышц.
Применение на практике
Пропорция типов волокон в мышцах будет влиять не на то, какой вес вы сможете поднять, какое количество повторений вы сможете сделать в интервальной работе, а на конечный результат — повышение силы/мощности мышц или выносливости.
Например, атлет, в мышцах которого большое содержание быстросокращающихся волокон, будет неспособен выполнить такое же количество повторений с весом, как атлет, в мышцах которого содержатся преимущественно медленносокращающиеся волокна.
Таким образом, атлет с FT-волокнами никогда не достигнет той мышечной выносливости, которая будет у атлета с ST-волокнами. Аналогично, атлет с большей пропорцией ST-волокон в мышцах не сможет поднять такой же вес, или пробежать интервалы так же быстро, как и атлет с большей пропорцией FT-волокон в мышцах. Следовательно, атлет с ST-волокнами не будет таким же сильным и мощным, как атлет с FT-волокнами.
Однако необходимо помнить, что даже внутри группы спринтеров или бегунов на длинные дистанции будет большой разброс по типам волокон в мышцах. Не все спринтеры имеют одинаковый процент FT-волокон, не все бегуны на длинные дистанции имеют одинаковый процент ST-волокон. Поэтому, одни спринтеры могут сделать работу 12х200 м, тогда как другие устанут после 8 повторов.
В зависимости от типа волокна и быстроты наступления утомления (из-за большего количества FT-волокон) необходимо решить, нужно ли больше отдыхать между интервалами для того, чтобы закончить работу, или необходимо уменьшить количество интервалов и увеличить скорость в серии.
Тренировка FT-волокон мышцы для выносливости не увеличит количество ST-волокон, а тренировка ST-волокон для силы и мощности не приведет к увеличению количества FT-волокон.
При соответствующем тренинге, FT-B волокна могут принять на себя некоторую часть выносливости, характерную для FT-BA волокон, а FT-A волокна могут принять на себя некоторую часть силы и мощности, характерной для FT-B волокон.
Однако, не существует полной взаимозаменяемости волокон. FT-волокна не могут стать ST-волокнами, и наоборот. Другими словами, то, с каким процентным соотношением волокон родился человек, с таким он будет жить и тренироваться.
Несмотря на то, что тип волокон не может быть изменен с одного на другой, тренировки могут изменить ту площадь, которую занимает определенный тип волокон в мышце. Другими словами, может произойти выборочное увеличение волокон, путем воздействия на них тренировками.
Другими словами, может произойти выборочное увеличение волокон, путем воздействия на них тренировками.
Например, у атлета в мышце может быть соотношение FT/ST-волокон 50/50, но поскольку площадь поперечного сечения FT волокон обычно больше, чем у ST-волокон, 65% площади мышцы могут занимать быстросокращающиеся, а 35% — медленносокращающиеся волокна.
При тренировках с отягощениями для повышения силы мышц, соотношение FT/ST-волокон останется таким же — 50/50, однако изменится площадь поперечного сечения, занимаемая двумя типами волокон. Это произойдет, потому что ST-волокна атрофируются, а FT-волокна гипертрофируются.
В зависимости от интенсивности тренировки, площадь мышцы может состоять на 75% из FT-волокон, и на 25% из ST-волокон. Эти изменения повлекут за собой повышение силы, но уменьшение выносливости.
Кроме этого, поскольку масса FT-волокон больше, чем ST-волокон, атлет будет набирать массу, если измерить окружности мышц.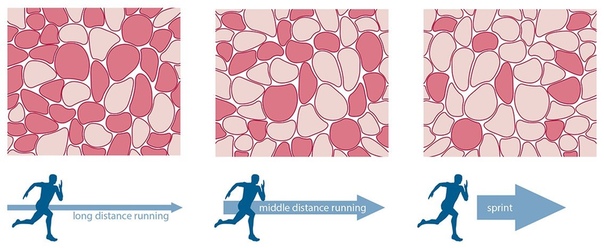 Напротив, если атлет тренируется для повышения выносливости, FT-волокна атрофируются, а ST-волокна гипертрофируются, вызывая увеличение площади поперечного сечения ST-волокон.
Напротив, если атлет тренируется для повышения выносливости, FT-волокна атрофируются, а ST-волокна гипертрофируются, вызывая увеличение площади поперечного сечения ST-волокон.
Площадь мышцы, изначально состоявшая на 65% из FT и на 35% из ST-волокон, может измениться под влиянием тренировок, и соотношение будет 50% на 50%. Кроме того, из-за того, что масса ST-волокон меньше, чем FT, наряду с повышением выносливости, произойдет снижение силы, а также потеря некоторой части мышечной массы.
Известный факт: если необходим прирост силы мышц, нужно тренироваться с тяжелыми весами и небольшим количеством повторов.
Этот режим тренировок приводит к рекрутированию FT-B волокон, которые могут развивать большее усилие, чем ST или FT-A волокна. Гипертрофия развивается только в перегруженной мышце, поэтому во время тренировки будет происходить рекрутмент FT-B волокон и их дальнейшая гипертрофия.
Тренировки с низкой или умеренной интенсивностью не всегда приводят к рекрутменту FT-B волокон, следовательно, для вовлечения этих волокон интенсивность должна быть высокой.
Выводы
Необходимо помнить, что для максимального результата необходимо тренироваться в соответствии со своей генетической предрасположенностью или на основании того, на каких соревнованиях вы показываете лучшие результаты.
Например, для атлета с преобладанием медленносокращающихся волокон, большую пользу принесет увеличение километража и тренировки с небольшими весами и большим количеством повторов.
С другой стороны, те атлеты, у которых преобладают быстросокращающиеся волокна, извлекут больше пользы из «спринтерских» методов тренировок и тренировок с тяжелыми весами и небольшим количеством повторов.
Было доказано, что длинные пробежки способствуют развитию медленносокращающихся волокон, улучшая их аэробные качества, тренируя их устойчивость к утомлению. Темповые тренировки влияют на ST и FT-A волокна, среди прочих эффектов улучшая работу этих типов волокон в связке.
Интервальная работа вовлекает FT-A и FT-B волокна, тренируя их взаимодействие и улучшая нейромышечную координацию. Скоростная работа: набегания, спринт в горку, короткие спринты на дорожке стадиона позволяют по максимуму нагрузить FT-B волокна.
Скоростная работа: набегания, спринт в горку, короткие спринты на дорожке стадиона позволяют по максимуму нагрузить FT-B волокна.
Возможно, это не принесет большой пользы тем, кто бегает марафон, но увеличение силы позволит вам бежать более плавно и эффективно.
Автор статьи: доцент кафедры анестезиологии и реаниматологии СЗГМУ им. Мечникова Евгений Суборов
Типы мышечного волокна и гипертрофия мышц
Соотношение мышечных волокон играет важнейшую роль в вопросе о том, насколько вы способны стать сильнее и массивнее. Часто люди, тренируясь одинаково, достигают разных мышечных объемов. Почему так? Общеизвестный тезис гласит: если у вас много мышечных волокон медленносокращающегося типа, то гипертрофировать их будет сложно. И, наоборот: при сильном доминировании быстросокращающихся мышечных волокон атлет легко набирает мышечную массу.
Разница между медленными и быстрыми мышечными волокнамиПочему же медленные мышечные волокна плохо гипертрофируются? Традиционно объяснение сводится к биохимическим и физиологическим свойствам этого типа волокон, отличающихся от быстрых волокон.
Однако в научных работax (напр. в этой: [2]) показана причина, связанная именно с влиянием анаболических андрогенных гормонов – тестостерона и дигидротестостерона: силовые упражнения стимулируют образование рецепторов андрогенов в цитоплазме преимущественно БЫСТРЫХ мышечных волокон, с помощью которых тестостерон из кровотока попадает в них (в частности, в ядро) и стимулирует там рост (в частности, влияет на транскрипцию генов, запуская программу синтеза сократительных белков). У медленных волокон реакция рецепторов на изменение уровня тестостерона отсутствует (кстати, об этом можно прочитать в любом хорошем учебнике по физиологии, опубликованном за последние 6-8 лет). Таким образом, повышение восприимчивости мышц к тестостерону как результат выполнения упражнений характерно для быстрых мышечных волокон.
Уровень андрогенных гормонов и рост мышечных волоконПолучается, что уровень собственных андрогенных гормонов в крови заметно будет сказываться на росте мышечных объемов лишь в том случае, если мышечные группы состоят в основном из быстросокращающихся волокон. Именно в этом случае будет работать комплекс реакций гормон-рецептор. Соответственно мышечный рост в этом случае будет сильно зависеть от уровня гормонального фона (чем больше гормонов, тем лучше рост, и наоборот).
Если же мышечные группы состоят преимущественно из медленных мышечных волокон, то даже повышенный уровень собственных андрогенных гормонов в крови НЕ будет способствовать заметному увеличению мышц. Ведь тренировки НЕ вызовут экспрессии рецепторов андрогенов в медленных волокнах, а только в быстрых, которых мало.
Показательным видится одно исследование – [4], где четко показано, что корреляция между повышением тестостерона (вследствие тренировок) и улучшением спортивного результата (рост силы) наблюдается у атлетически сложенных людей (вероятнее всего, это люди с доминированием быстрых волокон). И, наоборот, у мужчин с атлетически слабой композицией тела (скорее всего, это люди с доминированием медленных волокон) данной корреляции не выявлено (то есть всплески секреции гормонов не приводят к результату).
Как же накачать мышцы?Итак, андрогенные гормоны нужны в первую очередь для запуска сократительных белков в быстрых волокнах, то есть их миофибриллярной гипертрофии. Если быстрые мышечные волокна в меньшинстве, то достигнуть их роста за счет миофибриллярной гипертрофии эффективнее всего при тренировках в меньшем объеме и интенсивности по сравнению с традиционными (классическими) занятиями. Однако чем меньший процент быстрых волокон в мышце, тем меньший потенциал гипертрофии данной мышцы (иначе говоря, генетический предел можно быстро достичь).
Можно ли достичь миофибриллярной гипертрофии в медленных мышечных волокнах? Да! Но не за счет андрогенов, а за счет других анаболических гормонов, таких как гормон роста, инсулин и инсулиноподобный фактор роста 1 (ИФР-1).
Как это сделать в тренажерном зале? В одном исследовании [3] изучено влияние разных зон интенсивности отягощений на рост тех или иных мышечных волокон. Было установлено, что традиционные для силовых видов спорта зоны интенсивности отягощений, в частности зона 75-80% от 1 ПМ, а также 90-95% от 1 ПМ, приводят к миофибриллярной гипертрофии как БЫСТРЫХ, так и МЕДЛЕННЫХ волокон. Аналогичные выводы можно найти и в других научных работах.
Таким образом, миофибриллярная гипертрофия медленных волокон достигается примерно теми же методами, что и быстрых волокон. Однако, исходя из нашей практики, целесообразно чередовать периоды на стимуляцию (тренировками и питанием) тех или иных гормонов – подробнее см. Программа XXXL.
Увеличить медленные мышечные волокна также возможно за счет так называемой саркоплазматической гипертрофии и задержке воды в мышцах (отечность), что часто наблюдается в начинающих тренирующихся (подробнее об этом: [1]). По заключению многих ученых, для новичков оптимальный режим тренировок предусматривает работу с небольшими весами (40-50% от 1 ПМ) до отказа. Именно эта схема может быть таковой, при которой достигается именно саркоплазматическая гипертрофия (причем не только в медленных, но и быстрых волокнах).
ССЫЛКИ
[1] Damas F., et al. Early resistance training-induced increases in muscle cross-sectional area are concomitant with edema-induced muscle swelling // European Journal of Applied Physiology. 2015. Vol. 116. Iss. 1.
[2] Deschenes M.R., et al. Endurance and resistance exercise induce muscle fiber type specific responses in androgen binding capacity // Journal of Steroid Biochemistry and Molecular Biology. 1994. Vol. 50. No 3-4.
[3] Campos G.E. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones // European Journal of Applied Physiology. 2002. Vol. 88. Iss. 1-2.
[4] Crewther B.T., et al. The effects of two equal-volume training protocols upon strength, body composition and salivary hormones in male rugby union players // Biology of Sport. 2016. Vol. 33. No 2.
Понятие о тонических и фазических мышечных волокнах, их физиология.
Татаренко Дарья Андреевна
Нижегородский государственный педагогический университет имени Козьмы Минина
студентка 4 курса
Библиографическая ссылка на статью:
Татаренко Д.А. Понятие о тонических и фазических мышечных волокнах, их физиология. // Современные научные исследования и инновации. 2019. № 6 [Электронный ресурс]. URL: https://web.snauka.ru/issues/2019/06/89463 (дата обращения: 21.11.2021).
Важная информация о болях и травмах, искривлениях и мышечной гипотонии, также спазмах, защемлениях. Это – причина, почему у вас нет прогресса.
Сегодня мы будем говорить о мышцах, но не, о чем обычно пишется и говорится. Давайте поднимем тему, которую я нигде не прочитал кроме книг по анатомии, хотя это очень странно. Эта информация очень важна не только для получения высоких результатов, но и понимание этого поможет вам спасти свое здоровье.
Мы будем говорить о двух типах мышц, с точки зрения людей, которые занимаются спортом, будет разумно разделить мышцы на фазические и тонические.[1]
Короче говоря, мышцы фазические используются для движения, а тонические используются для удержания определенного положения.
Мышцы, которые используются для поддержки определенного положения, для стойкости. С точки зрения эволюции они старше, имеют больше способности к длительной работе, и кровь лучше поставляется им, менее быстро утомляющиеся, но главный недостаток – то, что они имеют тенденцию сокращаться.
Принимая во внимание, что мышцы фазические более легко быстро утомляются, кровь поставляется им в меньшей степени, и они не имеют тенденцию сокращаться, но эти мышцы склонны к атрофии без постоянной нагрузки.
Только теперь действительно имеет смысл говорить о различных типах мышечных волокон:
— быстро сокращающиеся волокна ( тип II),
— медленно сокращающиеся волокна (тип I)
Как корреляция этих мышечных волокон в мышце определяют ее принадлежность или фазической или тонической группе мышц.
Тонические мышцы. [3]
У тонических мышц есть способность сокращаться в течение длительного периода времени, когда только некоторые мышечные волокна напрягаются, а другие расслаблены, как следствие, мышечное напряжение может продлиться дольше. Тонические мышцы ответственны за осанку, другими словами, действуют преимущественно, чтобы выдержать вашу положение против силы тяжести.
У них много кровеносных капилляров и митохондрий, источник Аденозинового трифосфата (ATФ) является аэробным дыханием. Это позволяет тоническим мышцам работать в течение долгого времени без истощения. Одна из их особенностей – низкая скорость хранения молочной кислоты. Они лежат глубоко в мышцах человеческих конечностей и верхней части тела. В большей степени это короткие мышцы. В результате сокращения тонической мышцы, происходит нарушение статичного положения: сближение мест крепления мускула и увеличение его в объеме.
Мышцы фазические. [1]
У мышц фазических меньшее количество митохондрий и относительно меньше кровеносных капилляров. ATФ – источник энергии, один из конечных продуктов анаэробного дыхания (без кислорода). В отклик на раздражение фазичесие мышцы сокращается намного быстрее, чем тонические волокна. Истощение происходит довольно быстро, а также кислородная задолженность. Фазические мышцы обеспечивают быстрые сокращения, и они очень важны, чтобы выполнить быстрые амплитудное движение. Эти мышцы длинные, они лежат близко к поверхности тела. У мышц, которые выполняют движения короткой продолжительности в большей степени это фазические мышечные волокна (например, мышцы конечностей). [2]
При мышечных дисфункциях Фазические мышцы, как правило, ослабевают, находятся в состоянии ослабления (утомления, растяжения мышцы). Это проявляется пере растяжением отдельных мышечных, сухожильных и фасциальных волокон, сопровождаемое повышением порога возбудимости мышцы при ее активации (Васильева Л. Ф.,1997,2002). [3]
Нарушения статики (удаление крепления мышцы и уменьшение ее размера) происходят в результате расслабления фазических мышечных волокон наряду с дисфункцией мышцы. Динамические нарушения также замечены когда мышечное сокращение наступает чуть позже в отличии от нормы. Фазические мыщцы имеют связь с мозговой корой, которая позволяет выполнять сознательные движения под контролем.
Тонические мышцы находятся в контакте с более глубокими структурами как основного ядра, мозжечка и средним мозгом. [2]
Если тонические мышцы поврежденные или усталые, они становятся слабыми, у фазических мышц нет состояния спазма. Реакцией тонических мышц является сокращение, спазмы и боль.
Согласно морфофункциональным особенностям мышц есть такие мышечные волокна как промежуточные из тонических и фазических мышц. Они быстры и имеют способность к длительной работе.
Мы не знаем мышцы, которые состоят только из одного типа мышечных волокон кроме сердечной мышцы. Например, у мышц позвоночного столба есть до 95% волокон медленного сокращения; корреляция медленных и быстрых волокон в четырехглавой мышце бедра является почти такой же.
Считалось, что преобразовать один тип мышечных волокон в другой определенной тренировочной системой было невозможно, но практика опровергла эту теорию.
Эффект от специальной подготовки действительно в состоянии преобразовать промежуточные мышечные волокна в медленные или быстро сокращающиеся волокна.
Поскольку исследование узнало еще один решающий и интересный факт: если нерв будет отделен от волокна быстрого сокращения, и будет присоединен к волокну медленного сокращения, то тип волокна изменится. Оказывается, что корреляция мышечных волокон и то как мышцы возбуждаются, тесно взаимосвязано. [1]
Корреляция различных мышечных волокон закодирована генетически в теле каждого человека и различается в каждой мышце. Мы уже рождаемся или спринтером или марафонцем. Ясно, что у кого есть больше (белые) волокна быстрого сокращения, будет больше преимущества перед человеком, у которого есть больше волокон медленного сокращения (красных) в таком спорте, которому нужны быстрые шаги в ограниченное время, но человек с белыми волокнами проиграет, где ему нужна большая стойкость. Я уверена, что большинство уже поняло важность информации, что я сказала в этой статье и как это важно для нашего здоровья и не только его улучшения, но и прежде всего сохранения здоровья на тренировках.
Очень важно укрепить фазические мышцы и растягивать тонические. Это – единственный способ, которым вы можете избежать мускульного дисбаланса, которые вызывают у вас основную проблему. Возбуждение фазических мышц (антагонисты) нарушается из-за сокращения тонических мышц, и это еще больше увеличивает состояние дисбаланса.
Библиографический список
- Понятие о фазических и тоничеких мышцах [электронный ресурс] https://medbe.ru/materials/skeletnye-myshchtsy/ponyatie-o-fazicheskikh-i-tonicheskikh-myshtsakh/, дата обращения 11.05.2019 г.
- Фазические и тонические мышечные волокна [электронный ресурс] https://studopedia.ru/17_156650_fazicheskie-i-tonicheskie-mishechnie-volokna.htm, дата обращения 11.05.2019 г.
- Саркомер. Фазические и тонические мышечные волокна [электронный ресурс] https://studfiles.net/preview/3004161/page:14/, дата обращения 11.05.2019 г.
Количество просмотров публикации: Please wait
Все статьи автора «Татаренко Дарья Андреевна»
Типы мышечных волокон
Одним из главных параметров, отличающих прирожденных марафонцев от спринтеров является соотношение быстрых и медленных типов мышечных волокон. Помимо прочего, именно это соотношение во многом определяет, легко ли конкретный человек будет сжигать жир и набирать мышечную массу.
С другой стороны, незнание основ физиологии работы мышц ведет к выбору ошибочной стратегии тренировок. Для того, чтобы успешно наращивать мышцы или обладать рельефным и подтянутым телом с минимальным количеством усилий, необходимо лишь понимать, как устроена работа организма.
Быстрые и медленные мышечные волокна
Мускулатура человека состоят из соединительной ткани, капилляров, саркоплазмы и, непосредственно, мышечных волокон. Эти волокна, в свою очередь, делятся на быстрые и медленные, в зависимости от скорости их вовлечения в работу. Также отличаются цвет и источник энергии, который они используют.
Медленные (красные) волокна, ответственные за статические или продолжительные монотонные нагрузки, используют в качестве основного источника энергии жир. Быстрые (белые) волокна, необходимые для короткой и высокоинтенсивные нагрузки — запасы гликогена (углеводов) и креатина.
Различия мышечных волокон
Наглядным примером различия физиологии типов мышечных волокон является мясо курицы. Грудка и крылья обладают характерным белым цветом и минимальным количеством жира, тогда как окорочка и бедрышки отличаются темно-красным цветом мяса и более высоким содержанием жировой ткани.
Поскольку большую часть времени курица проводит стоя, мускулатура ее ног испытывает постоянную статическую нагрузку — основную работу выполняют медленные волокна. В противоположность этому, мышцы крыльев используются для энергичных взмахов — нагрузка идет на быстрые мышечные волокна.
Медленные / Красные волокна
Несмотря на то, что сами по себе медленные волокна тонкие и слабые, они могут поддерживать нагрузку продолжительное время. Их красный цвет обусловлен наличием молекул кислорода, необходимого для окисления жиров (триглицеридов), служащих для медленных волокон главным источником энергии.
Именно поэтому аэробный тренинг и продолжительное кардио идеальны для похудения — они вовлекает в работу медленные мышечные волокна и буквально плавят жировые запасы. Однако важно тренироваться в жиросжигающей зоне пульса для оптимального питания тканей организма кислородом.
Быстрые / Белые волокна
Для высокоинтенсивных и кратковременных нагрузок мышцы человека требуют быстродоступной энергии. Жир не подойдет, поскольку его транспортировка и окисление занимает как минимум несколько минут. Энергия должна находиться в легкодоступной форме как можно ближе к мышечным волокнам.
Для взрывных усилий используются быстрые мышечные волокна, работающие преимущественно на гликогене (запасах углеводов в мышцах), АТФ и креатин фосфате. Напомним, что рост мускулатуры в результате силовых тренировок во многом обусловлен именно увеличением энергетических запасов.
Мускулатура человека состоит из сплетения мышечных волокон различных типов. В стабилизирующих мышцах корпуса и позвоночника, внутренних мышцах живота и в мышцах ног обычно преобладают волокна медленного типа, а в прочей скелетной мускулатуре — волокна быстрого типа.
Однако тело способно адаптироваться под воздействием регулярных физических нагрузок и менять это соотношение. У профессиональных бегунов на марафонские дистанции более 80% всех мышечных волокон являются медленными, в отличии от спринтеров, у которых превалируют быстрые волокна (порядка 65-70%).
Как тренировать быстрые волокна?
Для тренировок быстрых мышечных волокон лучше всего подходят силовые упражнения. Чем выше рабочий вес и чем меньше количество повторений (и, соответственно, меньше время нахождения под нагрузкой), тем активнее в работе задействованы именно быстрые мышечные волокна.
Поскольку главным источником энергии для этого типа волокон являются запасы гликогена, чрезвычайно важно поддерживать достаточное количество углеводов в питании — именно поэтому для роста мышц прежде всего необходимы углеводы, а не просто спортивный протеин, как многие ошибочно полагают.
Типы мышечных волокон — Школа фитнеса «Smart Fitness PRO»
Типы мышечных волокон
Постоянно выполняемые нами движения по перемещению тела в пространстве или, наоборот, фиксация определенного положения и сохранение позы, работа внутренних органов — все эти действия возможны благодаря функционированию мышечных тканей человеческого организма.
Мышечные волокна человека делятся на типы, отличающиеся между собою структурой клеточной организацией и структурой.
- Тонические мышечные волокна. Характерная особенность этих волокон — низкая интенсивность и скорость сокращения.Тонические волокна обладают малым коэффициентом обмена миозиновой АТФ-фазы, поэтому не способны к интенсивным физических сокращениям. Расслабление мышц, структурными единицами которых являются тонические волокна, занимает достаточно много времени;
- Медленные фазические волокна окислительного типа (МС). Функция мышц, состоящих из волокон этого типа — фиксация положения тела в определенной позе — реализовавшееся благодаря тому, что медленные физических волокна окислительного типа медленно утомляются и быстро восстанавливаются. Именно эти волокна преобладают в человеческом теле, составляя от 50 до 55% всей мышечной массы. При этом такие характеристики волокон как выносливость или сила потенциально не зависят от гендерной характеристики. Мышечные волокна имеют темно-красный цвет, так как состоя преимущественно из белка миоглобина, обладающего способностью хорошо связывать кислород;
- Быстрые фазические волокна окислительного типа (БСб). мышцы, состоящие из таких волокон, выполняют быстрые сокращения. Благодаря высокому содержанию митохондрий медленно утомляются;
- Быстрые фазические волокна с гликолитическим типом окисления (БСа).В таких мышечных волокнах АТФ синтезируется во время процесса гликолиза. Соответственно, митохондрий в них содержится меньше, поэтому мышечные волокна утомляются быстро. Способны на интенсивные, быстрые физических сокращения. Мышечные волокна имеют характерный белый цвет ввиду отсутствия миоглобина;
Типы мышечных волокон
Мышцы, предназначенные для выполнения быстрых, интенсивных сокращений, состоят из достаточно малого количества волокон, в то время как мышцы, использующиеся, к примеру, при фиксации и сохранении позы, имеют в своем составе до нескольких тысяч структурных единиц.
Число мышечных волокон конкретного вида изменчиво и зависит от тех физических нагрузок, которые испытывает организм. К примеру, вов время занятия легкой атлетикой, бегом или плаванием на дистанцию от 500 м. в работу вовлекаются быстрые фазические волокна окислительного типа. Сокращение дистанции до 100-200 м. задействует быстрые фазические волокна с гликолитическим типом окисления.
Воздействие типов тренировок на развитие мышечных волокон
При обилии в тренировочном графике анаэробных тренировок, тренировок, быстрые физических волокна окислительного типа частично изменяют свои физиологические свойства, становясь похожими на волокна с гликолитический типом окисления. В свою очередь, быстрые физических волокна с гликолитический типом окисления постепенно приобретают свойства медленных физических волокон.
При тренировках на выносливость, направленных на развитие скоростных показателей, процесс «идет» в обратную сторону: медленные волокна приобретают свойства, характерные для быстрых волокон с гликолитический типом окисления, а те — свойства, характерные для БСб волокон.
Планируя тренировочный процесс с целью повлиять на соотношение мышечных волокон в организме, следует учитывать, что эта характеристика изначально индивидуальна и определена генетически. Поэтому необходимо учесть первоначальную структуру мышечных тканей, для чего вам придется обратиться к специалисту или окончить курсы инструкторов по фитнесу.
В. Н. Селуянов. Медленные и быстрые мышечные волокна | ДомСпорт.RU
— В прошлый раз мы с вами говорили о статодинамическом комплексе как единственном способе развития силы медленных мышечных волокон и о беге в гору как средстве не только повышения силового потенциала, но и средства перевода гликолитических мышечных волокон в окислительные. А можно ли выполнять упражнения для развития силы медленных волокон в изометрическом режиме?
— Практически никакой разницы нет. Биохимические, физиологические процессы похожи, но чисто статические упражнения перенапрягают нервную систему. Их сложнее выполнять психически, так как трудно терпеть в течение 40 секунд — психика в этом случае переносит упражнение значительно труднее.
— Мы говорили, что развитие медленных волокон не мешает быстрым. А силовые тренировки для быстрых волокон не вредят медленным?
— Как правило, нет, если не допускать сверхсильное закисление. Все знают, что если бегать 400 или 800 м регулярно в полную силу, то можно перетренироваться, потеряешь и силу, и выносливость, исчерпаешь и эндокринную систему.
— Вы много проверяли на практике свой метод. Расскажите о каком-нибудь конкретном примере роста силы медленных волокон после тренировки по вашему комплексу.
— Приведу пример классического эксперимента, когда одна группа делала приседания два раза в неделю в статодинамическом режиме. Один раз — 12 подходов, а второй — 3—4, выполняя таким образом тонизирующую тренировку. За 2 месяца у спортсменов сила выросла на 20% и на столько же вырос анаэробный порог. У второй группы, которая не занималась развитием силы вообще, а два раза в неделю проводила аэробную работу по 1 часу, ничего не изменилось. Второй пример — увеличение силы медленных волокон у спринтеров (8 человек). Их результаты в беге на 100 м были улучшены на 0,2—0,3 секунды: имея средний результат 10,9, они стали бежать за 10,7.
— Кроме бега в гору существуют и другие упражнения с сопротивлением. Скажем, бег по песку, бег с покрышкой и другие…
— Их воздействие аналогично бегу в гору, но поскольку меняется направление силы тяги, то и степень воздействия на отдельные мышцы также меняется. При беге в гору основную нагрузку несут мышцы задней поверхности бедра и ягодичная, потом четырехглавая, а если завершать отталкивание, то и икроножная. В беге по песку интенсивно работают икроножные мышцы, при беге с покрышкой — задняя поверхность бедра и ягодичная. Все эти упражнения очень эффективны.
В зависимости от степени и длительности усилия идет воздействие на силовой компонент гликолитических волокон и возможен перевод этих волокон в окислительные.
— Ну, а бег с поясом?
— Это упражнение я бы точно не рекомендовал. Оно увеличивает вертикальную нагрузку на опорный аппарат, и ничего не дает для мышц, продвигающих спортсмена. А вот травму при этом получить легко.
— Для перевода гликолитических волокон в окислительные кроме упражнений с сопротивлением можно применять и бег на отрезках?
— Действительно, бег с соревновательной скоростью может этому способствовать, но важно следить за длиной отрезка, не допуская большого закисления мышц. Я вновь могу сослаться на тренировку Коэ, на его пятиярусную схему, при которой используется бег на отрезках с соревновательной скоростью дистанций 5000 м, 3000, 1500, 800, 400 м. Хотя скорость этих пробежек превышала анаэробный порог, но из-за небольшой продолжительности закисления не происходило. Высокая скорость давала возможность включать гликолитические волокна, и (так же, как при беге в гору), наращивать внутри них митохондрии.
— В своих работах вы используете термин «максимальная алактатная мощность», или МАМ.
— Это максимальная мощность, которую мышцы развивают в короткий отрезок времени, буквально в секунды, например, прыжок в длину с места, бег на 20 м.
— Почему вы считаете, что для бегуна на средние дистанции это одна из важнейших характеристик?
— Если ее определять при беге с ходу, то она характеризует силу задней поверхности бедра, которая у средневика должна быть очень сильной и быстрой. Это фактически является показателем одаренности человека к бегу на 800 м. Если квалифицированный бегун не может развить скорость близкую к 10 м/с, то мировые достижения вряд ли ему покорятся. Понятно, что я имею в виду бегунов высокой квалификации. Спортсмен может плохо стартовать, плохо бежать 100 м, скажем, хуже 11 секунд, но если он 20 м с ходу бежит как нормальный спринтер, то у него может быть будущее.
— Понятно, что задняя поверхность должна быть быстрой. Но в то же время вы отмечали, что необходимо развивать силу медленных волокон. Как это сочетать?
— Одно другому не мешает, а только помогает. Однако быстрые люди имеют, при прочих равных условиях, более высокий потенциал. При равном поперечнике мышц они будут бежать быстрее. Теоретически бегуна на длинные дистанции можно превратить в средневика. Если в два раза больше гипертрофировать мышцы задней поверхности и он будет бежать быстрее.
— Мы уже говорили, что выполнение силовых упражнений поднимает общий гормональный фон организма. Известно, что многие спортсмены в своей подготовке используют анаболические стероиды. Им не хватает своих гормонов или они просто не используют собственные?
— Когда спортсмен не проводит силовую тренировку, то он отказывается от использования собственных гормонов и ему приходится вводить чужие, чтобы достигнуть результатов. И железы внутренней секреции постепенно перестают их производить, уменьшаясь в размерах.
— Многие не верят, что без фармакологии можно достичь высоких результатов. Скажите, возможно ли без анаболиков достичь таких же результатов, как с анаболиками?
— Элементарно. Дело в том, что все хотят получить результаты быстро. Если когда-нибудь введут жесткий допингконтроль, все волей-неволей начнут тренироваться нормально. Нынешних результатов вполне можно достичь и без помощи извне. Необходимо только правильно строить тренировочный процесс.
— На сколько дольше он будет длиться?
— Что касается беговых видов на выносливость, та разницы практически нет, так как наши бегуны фактически вообще не используют силовые упражнения. Достаточно начать их применять, делая это разумно, и сила быстро вырастет. Но большинство просто не знает этого пути. Для видов, требующих большой мышечной массы, например толкания ядра, может потребоваться на 2—3 года больше.
— Чтобы поддерживать свой ежедневный гормональный фон, вы рекомендовали выполнять силовые упражнения ежедневно по вечерам. Бегун должен делать упражнения для ног или, может быть, для верхнего плечевого пояса?
— Не имеет значения для каких групп мышц проводятся тонизирующие упражнения. Главное, чтобы не росли «ненужные» мышцы, а гормоны выделялись. Гормоны пойдут для строительства нужных мышц, в данном случае — ног, и здоровье при этом будет укрепляться.
— Бегуны на выносливость, особенно в подготовительном периоде, выполняют такие большие объемы бега при двухразовой, а то и трехразовой тренировке в день, что у них просто не хватает сил для того, чтобы проводить тренировку с отягощениями…
— На самом деле, длительный бег не стимулирует выработку гормонов. Это делают стрессовые нагрузки. Силовая либо спринтерская работа. Точнее, любая околопредельная работа, когда возникают мощные эмоции, где нужно терпеть. Если это быстрый бег, то гормоны появятся от беговых нагрузок.
— Действительно, большие объемы бега убивают силу…
— Культуристы это знают. Для строительства мышц нужна энергия, если ее направлять в другое место, то мышцы остаются на голодном пайке и строительства не происходит. Сочетать силовую и аэробную работу в одном занятии и в зависимости от величины аэробной работы в один день пользы не принесет.
— Я знаю, что вы противник больших объемов бега, но в истории бега на выносливость была эпоха Лидьярда с его марафонской тренировкой, давшей скачок результатов во всем мире, да и сейчас бегуны не мыслят достижения высоких результатов без длительного бега. Противники силовой тренировки часто ссылаются на высказывание знаменитого новозеландца: «Бегуну нужны мышцы оленя, но не льва».
— По моему мнению, длительный бег дает рост результатов, когда бегают по холмам. В это время происходит и развитие силового компонента рабочих мышц (увеличивается масса митохондрий в гликолитических мышечных волокнах). У сильнейших средневиков мира мы всегда отмечали полное отсутствие медленного бега трусцой. Я считаю его вредным, поскольку он загружает опорный аппарат ненужной нагрузкой, которая ничего не дает, кроме травм.
— Когда спортсмен бежит медленно, что работает?
— Только медленные волокна и только их малая, наиболее тренированная, часть. Поэтому в мышцах ничего не происходит. Нулевой эффект.
— Ну, а рост числа митохондрий?
— Они уже на пределе. Каждая миофибрилла оплетена митохондриями, новым просто морфологически некуда пристроиться. Если делать силовые упражнения, то добавляются новые миофибриллы и открываются новые возможности.
— И все-таки беговая практика показывает, что большой объем даже по равнине дает рост результатов…
— В этом случае прогресс идет за счет развития и совершенствования сердечно-сосудистой системы. Бегая по холмам, спортсмен тренирует и сердце, и мышцы. Если холмов нет, то мышцы останавливаются в развитии и не изменяются. Однако чем дольше бегать, если позволяет опорно-двигательный аппарат, с умеренной частотой сердечных сокращений — 120—150 уд/мин, тем больше вы воздействуете на сердечную мышцу и тем больше она растягивается. Мощное сердце может обеспечить приход кислорода порядка 6 литров к мышцам во время соревнований, но мало кто знает, что им больше 4 и не нужно. Количество рабочих мышц невелико, не сравнить с лыжниками. Но нужно отметить, что лишний кислород не помешает, бежать будет легче. Даже если анаэробный порог низкий, а сердце дает много кислорода, то он как бы повышается.
— Понятно о чем вы говорите, это то, что на потребительском рынке называется соотношением цены качеству. Огромная работа будет давать эффект совсем небольшой — несоразмерный с затраченными усилиями.
— Часто даже опытные бегуны с большим стажем допускают ошибку: по привычке, по накатанной схеме начинают подготовительный период вновь с больших объемов спокойного бега, не обращая внимания, что у них ЧСС в покое может доходить до 30 ударов в минуту. Сердце у них и так уже большое и мощное. Зачем его еще тренировать, зачем тратить понапрасну время? Это же не новичок, который бежит еле-еле и пульс у него доходит до 170.
— Мы все время говорили о тех спортсменах, у которых сердечная мышца уже максимально развита, и им необходимо заняться мышцами ног. Но у конкретного бегуна соотношение в развитии двух главных систем — транспортной и двигательной (упрощенно — сердце и ноги) может быть разным. Как это определить?
—Если спортсмен вышел на ЧСС 180 ударов в минуту и не чувствует никаких проблем с мышцами, то есть «по ногам» он может бежать и быстрее, значит сердце слабое. Значит, нужно посвятить 2—5 месяцев аэробной тренировке с умеренной скоростью. Потом опять проверить. Если в тесте на той же скорости ЧСС снизилась на 20—30 ударов, то можно заняться мышцами — снизить объем, повысить интенсивность, бегать по холмам.
Автор: Сергей Тихонов
Fast Muscle Fiber — обзор
2.2 Типы мышечных волокон
Медленно и быстро сокращающиеся мышечные волокна уже были кратко рассмотрены; здесь мы их подробно обсуждаем. Мышца состоит из разных мышечных волокон, которые различаются по внешнему виду и другим характеристикам. Например, сравнивая мышцы, выделенные у дикого и домашнего кролика, дикий кролик имеет более красноватый цвет. Также, если сравнивать куриную грудку с бедром, последнее более красноватое, чем грудка.
Таким образом, мышца, подвергающаяся постоянной нагрузке (например, мышца дикого кролика или бедра курицы), красноватая и состоит из медленно сокращающихся мышечных волокон, тогда как мышцы, которые не подвергаются постоянной нагрузке ( мышцы домашнего кролика и куриная грудка) имеют более светлый цвет и состоят из быстро сокращающихся мышечных волокон. Возникновение мышечных волокон зависит от напряжения, иннервации и типа иннервации. Внутри мышцы могут появляться мышечные волокна другого типа — например, ближе к костям мышцы более красноватые, чем у поверхности.Вообще говоря, разгибатели содержат больше быстро сокращающихся мышечных волокон, чем сгибатели. В человеческом теле есть мышцы, которые состоят в основном из медленно сокращающихся или быстро сокращающихся мышечных волокон. Мышечные волокна иннервируются альфа-мотонейронами. Моторный нейрон и все мышечные волокна, с которыми он соединяется, представляют собой двигательную единицу. Количество мышечных волокон, иннервируемых одним мотонейроном, может быть разным; например, в экстраокулярных мышцах 10 мышечных волокон иннервируются одним двигательным нейроном, в то время как мышцы бедра могут иметь 1000 волокон в каждой единице.Аксоны мотонейронов спинного мозга иннервируют периферические мышцы, и они могут иметь длину более 1 м (рис. 2.9).
Рис. 2.9. Моторные агрегаты. В человеческом теле есть три различных двигательных единицы. Моторная единица типа I обладает высокой устойчивостью к утомлению, имеет более низкий порог активации, содержит меньше мышечных волокон и имеет низкую силу, генерируемую во время сокращения. Моторный блок типа II также устойчив к усталости, имеет более высокий порог активации и создаваемое усилие выше по сравнению с типом I.Двигательная единица типа IIb утомительна, имеет высокий порог активации, иннервирует большинство мышечных волокон и создает наибольшую силу во время сокращения.
Электродвигатели различаются по размеру и порогу срабатывания. Большие двигательные единицы имеют более высокие пороги активации и содержат более бледные быстро сокращающиеся мышечные волокна, в то время как мелкие двигательные единицы имеют более низкие пороги активации и содержат красноватые медленно сокращающиеся мышечные волокна. Различия в физиологических, биохимических, гистохимических и генетических характеристиках двигательных единиц также служат полезной основой для их различения (Таблица 2.2).
Таблица 2.2. Характеристики мышечных волокон
| Характеристики | Быстросокращающиеся волокна | Медленно сокращающиеся волокна |
|---|---|---|
| Время до максимального сокращения (мс) | 50–80 | 100–200 |
| Частота для достижения тетанического сокращения (Гц / с) | 60 | 16 |
| Плотность миоглобина и митохондрий | Низкий | Высокий |
| Доминирующий путь синтеза АТФ | Анаэробный | Аэробный |
| Гликогеносодержание 900 | Высокая | Низкая |
| Миозин-АТФазная активность | Высокая | Низкая |
| Капилляризация | Низкая | Высокая |
| Устойчивость к усталости | Плохая | Высокая |
| Размер двигательного нейрона | Большой | Маленький |
| Высокий | Низкий | |
| Генерирующая сила | Высокий | Низкий |
Исходя из физиологических характеристик мышечные волокна человека быстро утомляются, устойчивы к быстрому утомлению, быстрые промежуточные или медленные волокна; по биохимическим свойствам они представляют собой быстрые гликолитические волокна типа IIb, быстрые окислительно-гликолитические волокна типа IIa или медленные окислительные волокна типа I.Другая классификация дает другой тип волокон, волокна IIi, с характеристиками между типами IIa и IIb. Красные медленно сокращающиеся волокна содержат большое количество железа, которое связано с большим количеством митохондрий и содержанием миоглобина. Красные волокна обладают более высокой окислительной способностью; они способны потреблять большое количество кислорода и уменьшать его содержание в митохондриях. Кислород всегда связан с железосодержащими молекулами; это высокое содержание железа также способствует его красному цвету. В таблице 2.2 показаны различия между типами мышечных волокон.Различия в пороге активации определяют порядок активации сокращающихся волокон разных типов. Медленно сокращающиеся мышечные волокна с отличным уровнем потребления кислорода, высоким содержанием митохондрий и активностью окислительных ферментов являются наиболее эффективными волокнами. Они способны создавать силу в точке сжатия из-за низкого порога активации. Большинство волокон антигравитационных мышц — это медленно сокращающиеся волокна, и эти волокна задействованы во время ходьбы и движений низкой интенсивности.Один из главных законов природы — это прибыльность, которая в данном случае означает задействование в первую очередь наиболее прибыльных мышечных волокон. Быстро сокращающиеся мышечные волокна с их более высокими порогами активации и генерированием огромной силы можно использовать во время полета и выживания; однако эти волокна потребляют много энергии и производят много молочной кислоты (обсуждается позже). Они могут быть активированы только стимулами высокой интенсивности из-за более высокого порога. Если использовать аналогию, медленно сокращающиеся волокна подобны экономичным городским машинам, а быстро сокращающиеся мышечные волокна — мощным гоночным автомобилям.
Типология мышечных волокон существенно влияет на время восстановления после упражнений высокой интенсивности
У людей волокна скелетных мышц подразделяются на две основные категории: медленно сокращающиеся волокна, также называемые волокнами типа I, и волокна с быстрым сокращением, или волокна типа II. . Последние делятся на подтипы IIA и IIX. Известно, что некоторые люди демонстрируют доминирующую быструю типологию (FT), тогда как другие имеют смешанную / промежуточную типологию (INT) или доминирующую медленную типологию (ST) (10). Средний тип мышечных волокон человека соответствует распределению по Гауссу, и у всех людей есть как быстро, так и медленно сокращающиеся волокна.Однако это распределение может составлять от 15% до 85% быстросокращающихся волокон (35). Несмотря на то, что типология мышечных волокон в основном определяется генетически (37), несколько исследований показывают, что между быстро сокращающимися подтипами существует смещение мышечных волокон, вызванное физической нагрузкой (1, 2). Напротив, двунаправленный сдвиг между медленными и быстро сокращающимися волокнами менее очевиден (16, 44). Таким образом, типовой состав мышечных волокон лежит в основе «типологии» спортсмена, поскольку у элитных спринтеров преобладают быстросокращающиеся волокна, тогда как у элитных спортсменов на выносливость относительно больше медленно сокращающихся волокон (10, 45).Поскольку быстро сокращающиеся волокна могут генерировать больше энергии, особенно при высокой скорости сокращения (43), а поскольку медленно сокращающиеся волокна более устойчивы к утомлению (36), можно сделать вывод, что типология мышц спортсмена является важным фактором, определяющим результативность. фактор во многих видах спорта. Однако эта информация редко применяется в повседневной спортивной научной практике.
Использование типологии мышц в спортивной науке и коучинге в основном затрудняется инвазивным характером текущей оценки состава мышечных волокон с помощью биопсии.Поэтому была разработана новая неинвазивная оценка типологии мышц, основанная на количественном определении карнозина в икроножных мышцах с помощью протонной магнитно-резонансной спектроскопии ( 1 H-MRS) (4). Поскольку карнозин, буфер рН для мышц, в большом количестве содержится в быстро сокращающихся волокнах, спортсмены с высокими концентрациями карнозина, по оценкам, обладают типологией быстрых мышц. Этот метод позволяет неинвазивно определять типологию мышц (тип I по сравнению с волокнами типа II), что может быть использовано для выявления и ориентации талантов (4, 6).Можно сомневаться, можно ли использовать эту технику для индивидуализации тренировочных циклов и циклов восстановления, поскольку известно, что одиночные быстро сокращающиеся двигательные единицы обладают более высокой утомляемостью, чем медленно сокращающиеся двигательные единицы (11).
Усталость всей мускулатуры проявляется в потере пиковой силы, скорости сокращения и / или мощности, и имеется некоторая доступная литература, указывающая на то, что мышечная усталость у людей зависит от типа состава мышечных волокон. Корреляции показали, что спортсмены с преимущественно быстросокращающимися волокнами более утомляемы по сравнению со спортсменами с преимущественно медленными волокнами во время устойчивых изометрических максимальных произвольных сокращений (MVC) (17), 50–100 изокинетических максимальных разгибаний колена (24, 41) и 60 с непрерывных прыжков (7).Наряду с утомлением типология мышечных волокон также влияет на профиль восстановления сразу после утомляющего упражнения. Colliander et al. (9) сообщили о более низком восстановлении начального пикового крутящего момента в течение 1-минутного периода отдыха между 3 × 30 максимальными изокинетическими разгибаниями колена у субъектов с преимущественно быстро сокращающимися волокнами по сравнению с медленными волокнами. Это соответствует данным Hamada et al. (18), которые показали, что восстановление электрически индуцированного пикового момента подергивания через 5 мин после шестнадцати 5-секундных максимальных произвольных изометрических сокращений разгибателей колена было значительно ниже у субъектов с преимущественно быстрым сокращением по сравнению смедленно сокращающиеся волокна. На сегодняшний день время восстановления у субъектов с дивергентным составом мышечных волокон после первых 5-минутных временных рамок восстановления еще предстоит исследовать. Тем не менее, эта информация является ключевой для индивидуализации тренировочных циклов и циклов восстановления, чтобы каждый спортсмен мог начать следующую тренировку в полностью восстановленном состоянии.
В этом исследовании мы неинвазивно оценили типологию мышц 32 субъектов и охарактеризовали их как ST, INT и FT.Впоследствии мы исследовали профиль мышечной усталости и длительного восстановления только в предварительно определенных группах ST и FT. Утомляемость вызывалась тремя повторными тестами Вингейта, а степень утомляемости и ход восстановления оценивали в течение 5 часов после Вингейтса с помощью комбинированного подхода к произвольному и электрически стимулированному сокращению разгибания колена. Мы предположили, что субъекты с FT характеризуются более выраженной утомляемостью и задержкой восстановления в течение 5 часов после высокоинтенсивных упражнений по сравнению с субъектами с ST.
Настоящее исследование было направлено на выяснение того, было ли априорное разделение спортсменов на группу быстрой или медленной мышечной типологии, основанное на неинвазивном измерении концентрации карнозина с помощью 1 H-MRS в икроножной мышце, было связано с утомляемостью. во время высокоинтенсивного упражнения и, что более важно, профиля восстановления после этого утомляющего упражнения. Несмотря на умеренные пределы включения по типологии мышц ( z балл ниже -0,5 или выше +0.5), наблюдались заметные различия в профилях утомляемости и восстановления между группами.
Во время повторных испытаний Wingate падение мощности через каждый Wingate было намного выше в группе FT по сравнению с группой ST. Это наблюдение согласуется с результатами инвазивной биопсии Bar-Or et al. (5) и Inbar et al. (21), которые обнаружили положительную взаимосвязь между падением мощности во время одного Wingate и средней площадью быстро сокращающихся / средней площадью медленных сокращений ( r = 0.75) и между падением мощности во время одного Wingate и процентом быстро сокращающихся мышечных волокон в широкой широкой мышце бедра ( r = 0,52), соответственно. Соответственно, в этом исследовании общее падение мощности по трем повторным Wingates показало самую сильную связь с типологией мышц ( r = 0,677). Более того, в группе ST практически не наблюдалось накопления утомляемости по всему Wingates, поскольку пиковая мощность была одинаковой между первым и вторым Wingate и снизилась только на 3.2% от второго к третьему Wingate. Напротив, пиковая мощность группы FT уменьшалась с каждым последующим Wingate (-4,0% с Wingate 1 до Wingate 2 и -12% с Wingate 2 до Wingate 3 ). Более высокий уровень накопленной усталости у субъектов с FT, о чем свидетельствует более высокое падение общей мощности и снижение пиковой мощности, соответствует литературным данным, показывающим более высокую накопленную усталость у субъектов с преимущественно быстро сокращающимися волокнами после трех приступов максимального одностороннего коленного сустава. расширений (−18%) и после шестнадцати 5-секундных MVC (−26%) (9, 18).
Несмотря на разные профили утомления, общая проделанная работа была одинаковой в обеих типологиях, что позволило нам исследовать ход восстановления мышечного момента после идентичного объема выполненной работы. Через десять минут после повторных тестов Вингейта восстановление крутящего момента MVC было на 18% ниже в группе FT по сравнению с группой ST. Эта разница в восстановлении соответствует данным Hamada et al. (18), которые обнаружили, что состояние восстановления в их быстро сокращающейся группе на 16% ниже, чем в их медленно сокращающейся группе, через 5 минут после шестнадцати 5-секундных максимальных произвольных изометрических сокращений.До сих пор было неизвестно, присутствует ли эта разница между типологическими группами только в первые минуты фазы восстановления или будет длиться несколько часов. Это исследование впервые демонстрирует, что через 2 часа после тренировки высокой интенсивности разница в максимальном восстановлении крутящего момента между группами по типологии все еще составляла 11%. Группа ST полностью восстановилась до исходного уровня через 20 минут после Wingates, тогда как группа FT еще не восстановилась через 5 часов, и их время на восстановление, таким образом, превысило временные рамки этого исследования.Эти результаты имеют значение для дальнейших исследований, поскольку ранее считалось, что MVC быстро восстанавливается после высокоинтенсивных велосипедных упражнений (14, 39). Однако это могло бы ввести в заблуждение, если бы тип мышечных волокон не принимался во внимание, поскольку в этом исследовании быстрое восстановление наблюдалось при ST, но не при FT. Более того, если бы это исследование было сосредоточено на влиянии трех повторных Wingates, например, на ЗГТ после стимуляции 10 Гц без какого-либо разделения на типологии, не было бы никакого эффекта для этого параметра ( P = 0.177; Рис.10). Тем не менее, высокое стандартное отклонение через 10 минут после повторных тестов Вингейта можно объяснить противоположными эффектами в обеих группах, с увеличением ЗГТ в группе FT и небольшим снижением (значимым при 20, но не 10-минутном восстановлении) в группе ST. Таким образом, дифференциация между группами мышечной типологии может привести к новому пониманию мышечной усталости человека и механизмов восстановления.
Рис. 10. Противодействующие эффекты утомления на время половинного расслабления (HRT) дублета 10 Гц в обеих группах типологии: увеличение HRT в быстрой типологии (FT) через 10 мин после повторения Wingates и уменьшение медленная типология (ST) через 20 мин после повторного Wingates. A : результаты по всем предметам (ST и FT). B : результаты для отдельных субъектов. * Значительно отличается между группами; $ значительно отличается от базового уровня.
Восстановление произвольных параметров сочеталось с электрически индуцированными реакциями, чтобы понять причину утомления (центральная или периферическая). Путем администрирования SID во время MVC была проведена VAL. Это измерение в сочетании с отношением RMS-волны к M показало, что центральная усталость не присутствовала в обеих группах от 10 минут до 5 часов после выполнения задачи, вызывающей утомляемость.Это согласуется с современной литературой, поскольку центральное утомление чаще всего возникает после длительных тренировок на выносливость, но не после коротких интенсивных упражнений (8). Возможно, что центральная усталость присутствовала во время Wingates, поскольку литература по усталости во время тотальной повторяющейся езды на велосипеде обычно показывает, что центральная двигательная команда ограничивает производительность, чтобы предотвратить чрезмерное утомление опорно-двигательных мышц. Поскольку в этом исследовании центральная утомляемость измерялась только через 10 минут после Уингейтса, она могла восстановиться в этот период времени (20, 34).Когда iEMG / крутящий момент был проанализирован через 10 минут после Wingate, стало ясно, что, несмотря на неизменную центральную активацию, группа FT не смогла создать такое же количество крутящего момента после повторных тестов Wingate. Более высокое отношение iEMG к крутящему моменту в FT также наблюдалось Nilsson et al. (31), которые обнаружили положительную корреляцию между увеличением отношения ЭМГ к крутящему моменту и процентным содержанием быстро сокращающихся волокон, что указывает на то, что местные факторы в быстро сокращающихся волокнах вызывают развитие утомляемости (31).Прежде чем можно было интерпретировать периферическую усталость, были исследованы изменения в нервно-мышечном соединении. Область M-волны показала усталость во время передачи потенциала действия по нервно-мышечному соединению в обеих группах через 20 мин после повторных тестов Вингейта. На основании литературных данных не ожидалось никаких различий в свойствах M-волн после трех повторных тестов Вингейта (32). В группе ST это снижение присутствовало до 50 минут, тогда как в группе FT оно наблюдалось до 2 часов.Этот паттерн уже наблюдали Макфадден и МакКомас (28), которые предположили, что это снижение возникло не из-за нарушения нервно-мышечной передачи, а из-за проблемы с мембранным потенциалом, инициированной, например, повреждением сарколеммы. Хотя после повторных тестов Вингейта повреждения не ожидается, поскольку оно в основном характеризуется концентрическими сокращениями (32), его нельзя полностью исключить в некоторых быстро сокращающихся волокнах.
Поскольку максимальные вызванные усталостью дефициты в дублете 100 Гц (-23%), дублете 10 Гц (-43%) и синглете (-37%) больше, чем изменения в M-волне (-18 %), и эти дефициты электрически вызванного крутящего момента уже присутствуют в начале восстановления, в отличие от дефицита М-волны, важные угнетающие крутящий момент изменения будут иметь место и за пределами мышечной мембраны.Ранняя периферическая усталость может быть связана с увеличением побочных продуктов P и , H + , а также АДФ и местной депривацией энергии (3). Накопление этих продуктов может быть выше в быстро сокращающихся волокнах из-за большей зависимости от анаэробных процессов для обеспечения энергией (29). Более того, накопление может по-разному влиять на производство крутящего момента в FT по сравнению с группой ST. Например, увеличение P и оказывает большее влияние на эффективность быстро сокращающегося волокна по сравнению с медленным волокном (25) и отрицательное влияние увеличения H + на переход к высокоскоростному волокну. Силовое состояние присутствует в быстрых, но не в медленных волокнах (30).Кроме того, местное истощение АТФ могло присутствовать вокруг насосов саркоплазматического ретикулума Ca 2+ в быстро сокращающихся волокнах, поскольку они имеют более высокую плотность насосов саркоплазматического ретикулума Ca 2+ и, следовательно, потребляют большее количество АТФ ( 13, 22). Теоретически все побочные продукты должны быть извлечены через 2 часа и, таким образом, не могут объяснить вывод о том, что восстановление занимает более 5 часов на всех частотах стимуляции.
Во время утомления относительное уменьшение производства крутящего момента часто больше на высоких частотах по сравнению с низкими частотами активации, потому что во время утомления присутствует удлинение переходного процесса Ca 2+ , что приводит к усиленному слиянию стимуляций на более низких частотах и следовательно, приводит к более высокому крутящему моменту (23).Напротив, в этом исследовании отношение 10 Гц к 100 Гц уменьшается в утомленном состоянии; таким образом, 10 Гц понижены в большей степени, чем 100 Гц. Это явление можно объяснить наличием длительной депрессии низкочастотной силы, которая может быть причиной длительного утомления (12). Во время длительного низкочастотного подавления силы изменения в обработке Ca 2+ будут влиять на производство крутящего момента больше на более низких частотах, поскольку более низкие частоты стимуляции находятся на более крутой части внутриклеточной концентрации Ca 2+ ([Ca 2+ ] i ), чем более высокая стимуляция.Механизмом длительной низкочастотной депрессии силы являются либо снижение высвобождения Са 2+ саркоплазматического ретикулума, либо снижение чувствительности миофибрилл к Са 2+ . Точнее, причиной различной обработки Ca 2+ после трех повторных Wingates у рекреационно тренированных субъектов может быть фрагментация саркоплазматического ретикулума канала высвобождения Ca 2+ (32). Поскольку отношение 10 Гц к 100 Гц было значительно ниже в группе FT, эта фрагментация могла быть выше в этой группе.
Настоящее исследование предоставляет доказательства длительного дефицита максимальной произвольной мышечной силы у некоторых, но не у всех людей, что, по-видимому, в основном связано с определенной типологией мышц. Поскольку максимальное произвольное (изометрическое) сокращение требует одновременного задействования почти всех мышечных волокон, можно предположить, что медленно сокращающиеся волокна полностью восстанавливаются после тотальных тестов Вингейта в течение нескольких минут, тогда как быстро сокращающимся волокнам, вероятно, потребуются часы, чтобы восстановить полный исходный сократительный потенциал.В этом случае люди с преобладающим распределением медленно сокращающихся волокон будут демонстрировать небольшой дефицит силы и быстрое восстановление, тогда как люди с преобладающим распределением быстро сокращающихся волокон будут иметь большую долю своих мышечных волокон в состоянии длительного дефицита, что приведет к скомпрометированный крутящий момент всей мышцы в течение нескольких часов. Чтобы проверить эту гипотезу, мы сравнили абсолютные сократительные кинетические свойства в трех электрически стимулированных условиях как в состоянии покоя, так и в состоянии утомления.Исходно скорость развития крутящего момента, пиковый крутящий момент и скорость релаксации крутящего момента были выше в группе FT по сравнению с группой ST. Несмотря на эти глубокие базовые различия, средний след группы FT больше не отличался от следа группы ST через 10 минут после повторного теста Вингейта (рис. 8). Более того, большинство этих сократительных кинетических свойств оставались одинаковыми между группами до 5 часов восстановления, за исключением пикового крутящего момента на частоте 100 Гц и скорости релаксации крутящего момента в синглете, которые значительно различались через 2 часа, и скорости развития крутящего момента. при 100 Гц, что значительно изменилось через 50 мин.Поскольку эти кривые отражают совокупность всех мышечных волокон, которые участвуют в производстве крутящего момента, все быстро сокращающиеся мышечные волокна, возможно, были утомлены, и поэтому только менее утомленные медленно сокращающиеся мышечные волокна вносили свой вклад в электрически индуцированный мышечный момент.
Мы должны признать, что различия в исходных характеристиках в весе, безжировой массе тела и массе четырехглавой мышцы (оцениваемой по скорректированной окружности) между группами типологии мышц были ограничением исследования, которое могло привести к различиям в крутящем моменте при MVC. и электрическая стимуляция в покое, хотя мы не ожидаем, что эти различия вызовут все базовые различия в скорости развития вращающего момента и расслабления.Последнее можно объяснить другой типологией мышечных волокон и, следовательно, подтвердить, что измерение карнозина связано с типологией мышц. Более того, это также доказывает наличие поперечного мышечного фенотипа, поскольку карнозин измерялся в икроножной мышце, но дифференциальные исходные ответы на электрическую стимуляцию были обнаружены в четырехглавой мышце. Этот поперечно-мышечный фенотип предполагает, что соотношение типов волокон в одной мышце является показателем доли всего тела (42).Мы решили измерить типологию мышц икроножной мышцы, так как это позволило нам сравнить концентрацию карнозина с большой эталонной базой данных (98 рекреационно активных контрольных мужчин-мужчин), чтобы разделить группу на репрезентативные группы медленной и быстрой типологии.
Наряду с функциональными измерениями с помощью электрической стимуляции были предприняты другие попытки неинвазивной оценки типологии мышечных волокон, такие как прыжок в противоположном направлении (7) и тензиомиография (38). Однако все эти измерения имеют одни и те же недостатки: на них могут влиять разминка, тренировка, техника, усталость, мотивация и острый прием пищи.Наша неинвазивная оценка типологии мышц, основанная на измерении карнозина с помощью 1 H-MRS, проводится в состоянии покоя и, следовательно, не подвержена влиянию техники, мотивации или утомления. Более того, карнозин является очень стабильным метаболитом, на который не влияют тренировки или острый ежедневный прием пищи (4).
Хотя упражнения на утомление в этом исследовании (3 повторения Wingates) были в основном концентрическими, мы предполагаем, что более высокая утомляемость и время на восстановление в группе FT также будут присутствовать после упражнений с эксцентрическим компонентом, таких как бег.Более того, эксцентрический компонент может даже преувеличивать различия между группами, поскольку было высказано предположение, что быстро сокращающиеся волокна легче повреждаются после эксцентрических упражнений, чем медленно сокращающиеся волокна (15). Следовательно, эта информация важна для спортсменов, особенно если они характеризуются высокой разнородностью типологии мышц внутри одной команды. До сих пор тренировки не корректировались в соответствии с типологией мышечных волокон, и, таким образом, все спортсмены выполняют одни и те же тренировочные циклы и циклы восстановления, хотя это вызывает различные реакции у спортсменов с разной типологией мышц.Таким образом, некоторые спортсмены могут подвергаться риску накопленной усталости, перетренированности и потенциально травм. Предлагаемый подход к неинвазивной оценке типологии мышц может быть значимым инструментом для индивидуализации тренировочных циклов и циклов восстановления.
Таким образом, настоящее исследование показывает, что разные группы мышечной типологии утомляются по-разному во время трех повторных тестов Вингейта, с более высокой степенью утомляемости во время Вингейтса у субъектов с FT по сравнению с субъектами с ST.Более того, всего лишь 90 секунд упражнений высокой интенсивности вызвали длительную усталость и нарушения сократительной функции после этих упражнений Wingates. Эта послетренировочная усталость была более выражена в группе FT и привела к неполному восстановлению произвольных сокращений в течение 5 часов после Вингейта. С другой стороны, произвольный крутящий момент группы ST уже восстановился через 20 минут после повторения Уингейтса. Эти результаты могут иметь отношение к циклам тренировок и восстановления и могут открыть возможности для более индивидуальной подготовки спортсменов на основе их типологии мышц.
10.5 Типы мышечных волокон — анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Опишите типы волокон скелетных мышц
- Объясните быстрые и медленные мышечные волокна
Два критерия, которые следует учитывать при классификации типов мышечных волокон, — это скорость сокращения одних волокон по сравнению с другими и то, как волокна производят АТФ. Используя эти критерии, можно выделить три основных типа волокон скелетных мышц.Медленные окислительные (SO) волокна сокращаются относительно медленно и используют аэробное дыхание (кислород и глюкоза) для производства АТФ. Волокна быстрого окисления (FO) быстро сокращаются и в основном используют аэробное дыхание, но поскольку они могут переключаться на анаэробное дыхание (гликолиз), они утомляются быстрее, чем волокна SO. Наконец, быстрые гликолитические (ФГ) волокна быстро сокращаются и в основном используют анаэробный гликолиз. Волокна FG устают быстрее других. Большинство скелетных мышц человека содержат все три типа, хотя и в разных пропорциях.
Скорость сокращения зависит от того, насколько быстро АТФаза миозина гидролизует АТФ, чтобы вызвать действие поперечного мостика. Быстрые волокна гидролизуют АТФ примерно в два раза быстрее, чем медленные волокна, что приводит к гораздо более быстрой смене поперечного мостика (которая тянет тонкие волокна к центру саркомеров с большей скоростью). Первичный метаболический путь, используемый мышечным волокном, определяет, классифицируется ли волокно как окислительное или гликолитическое. Если волокно в основном производит АТФ посредством аэробных путей, оно является окислительным.Во время каждого метаболического цикла может производиться больше АТФ, что делает клетчатку более устойчивой к утомлению. Гликолитические волокна в основном создают АТФ посредством анаэробного гликолиза, который производит меньше АТФ за цикл. В результате гликолитические волокна утомляются быстрее.
Окислительные волокна содержат намного больше митохондрий, чем гликолитические волокна, потому что аэробный метаболизм, который использует кислород (O 2 ) в метаболическом пути, происходит в митохондриях. Волокна SO содержат большое количество митохондрий и способны сокращаться на более длительные периоды из-за большого количества АТФ, которое они могут производить, но они имеют относительно небольшой диаметр и не создают большого напряжения.Волокна SO широко снабжаются кровеносными капиллярами, чтобы поставлять O 2 из красных кровяных телец в кровотоке. Волокна SO также содержат миоглобин, молекулу, несущую O 2 , аналогичную O 2 , несущую гемоглобин в красных кровяных тельцах. Миоглобин хранит часть необходимого O 2 внутри самих волокон (и придает волокнам SO их красный цвет). Все эти особенности позволяют волокнам SO производить большое количество АТФ, который может поддерживать мышечную активность без утомления в течение длительных периодов времени.
Тот факт, что волокна SO могут функционировать в течение длительного времени без утомления, делает их полезными для поддержания осанки, создания изометрических сокращений, стабилизации костей и суставов и выполнения небольших движений, которые происходят часто, но не требуют большого количества энергии. Они не создают высокого напряжения, и поэтому не используются для мощных, быстрых движений, требующих большого количества энергии и быстрой езды на велосипеде по мосту.
ВолокнаFO иногда называют промежуточными волокнами, потому что они обладают промежуточными характеристиками между быстрыми и медленными волокнами.Они производят АТФ относительно быстро, быстрее, чем волокна SO, и, таким образом, могут создавать относительно высокое напряжение. Они окислительны, потому что производят АТФ аэробно, обладают большим количеством митохондрий и не утомляются быстро. Однако волокна FO не содержат значительного количества миоглобина, что придает им более светлый цвет, чем волокна SO красного цвета. Волокна FO используются в основном для движений, таких как ходьба, которые требуют больше энергии, чем контроль позы, но меньше энергии, чем взрывные движения, такие как спринт.Волокна FO полезны для этого типа движения, потому что они создают большее натяжение, чем волокна SO, но они более устойчивы к усталости, чем волокна FG.
ВолокнаFG в основном используют анаэробный гликолиз в качестве источника АТФ. Они имеют большой диаметр и содержат большое количество гликогена, который используется в гликолизе для быстрого образования АТФ и создания высокого уровня напряжения. Поскольку они в первую очередь не используют аэробный метаболизм, они не обладают значительным количеством митохондрий или значительным количеством миоглобина и поэтому имеют белый цвет.Волокна FG используются для создания быстрых, сильных сокращений для быстрых и мощных движений. Эти волокна быстро устают, что позволяет использовать их только в течение коротких периодов времени. Большинство мышц содержат смесь волокон каждого типа. Преобладающий тип волокна в мышце определяется основной функцией мышцы.
Волокна дыхательных мышц: специализация и пластичность
Основная функция легких — вентиляция крови. Переменный поток воздуха к альвеолярной поверхности и от нее управляется градиентами давления, создаваемыми дыхательными мышцами.Несмотря на их конкретную задачу, которая не позволяет им отдыхать в течение всей жизни, дыхательные мышцы имеют ту же структуру и функции, что и все другие мышцы конечностей и туловища. Специализация дыхательных мышц проистекает непосредственно из характеристик волокон, из которых они состоят. Хорошо известно, что волокна скелетных мышц очень разнородны: цель этого обзора — выявить особенности волокон дыхательных мышц, которые позволяют им работать, не утомляясь.Однако волокна дыхательных мышц не только узкоспециализированы для выполнения своих функциональных задач, но также могут изменять свои свойства, чтобы адаптироваться к новым требованиям, которые могут возникать в результате физиологических условий, таких как физические упражнения, заболевания легких или дыхательных путей. Точно так же изменения в дыхательных мышечных волокнах в результате мышечного заболевания могут серьезно нарушить дыхательную функцию.
ГЕТЕРОГЕННОСТЬ МЫШЕЧНЫХ ВОЛОКН
Мышечные волокна не равны: способность генерировать силу и механическую мощность, временной ход сократительной реакции и сопротивление утомлению заметно различаются от волокна к волокну.На основе их функциональных свойств волокна можно разделить на большие группы, обычно известные как «типы волокон». 1, 2 Существует несколько классификаций типов волокон, наиболее распространенные из которых показаны в таблице 1.
Таблица 1Классификация волокон скелетных мышц человека и отличительные функциональные или молекулярные свойства каждого типа волокон
Современная молекулярная биология показывает, что каждый тип волокон является результатом определенного профиля экспрессии генов, основанного на двух основных механизмах: 1 (1) количественная регуляция экспрессии (повышающая или понижающая регуляция) для многих генов; и (2) избирательная экспрессия других генов, которые существуют во множестве копий с небольшими различиями («изогены») и дают начало изоформам, то есть белкам, достаточно похожим, чтобы заменять друг друга, но достаточно разнообразным, чтобы придать волокну отличные функциональные свойства.Недавнее сравнение, основанное на методах гибридизации микрочипов на наборе из 6519 генов, показало, что 177 генов (около 3%) по-разному экспрессируются в медленных и быстрых мышцах. 3
Способность мышечного волокна генерировать напряжение, укорачиваться и производить механическую энергию в основном определяется составом миофибриллярного белка и, в первую очередь, составом изоформ миозина. Миозин — это двигатель сокращения мышц, то есть он способен преобразовывать химическую энергию АТФ в механическую энергию (работу).Молекула миозина, гексамерный белок, состоит из двух тяжелых цепей (MyHC) и четырех легких цепей (MyLC): как MyHC, так и MyLC существуют в нескольких изоформах. Четыре изоформы MyHC экспрессируются в скелетных мышцах туловища и конечностей взрослых млекопитающих: медленные, 2A, 2X и 2B. MyHC-2B, однако, не экспрессируется в мышцах человека. 2 Кроме того, другие изоформы экспрессируются в специализированных мышцах — например, экстраокулярный MyHC и жевательный MyHC экспрессируются во внешних мышцах глаза и мышцах челюсти, соответственно.Во время развития волокна дыхательных мышц, как и все другие волокна скелетных мышц, экспрессируют изоформы развития: эмбриональный MyHC и неонатальный MyHC. 1
ИзоформыMyHC обычно используются для классификации типов волокон, потому что:
они являются основными детерминантами сократительных свойств;
MyHC — самый распространенный белок; и
несколько методов, используемых для идентификации типов волокон, включая гистохимическую реакцию mATPase на моноклональные антитела и электрофорез в полиакриламидном геле, основаны на изоформах MyHC.
С учетом изоформного состава MyHC мышечные волокна классифицируются как медленные (содержащие MyHC медленный) или быстрые 2A, быстрые 2X и быстрые 2B (содержащие MyHC 2A, 2X или 2B соответственно; таблица 1). В мышечных волокнах человека максимальная скорость укорочения (рис. 1A) и скорость гидролиза АТФ (рис. 1B) постепенно увеличиваются от медленных до волокон 2A и 2X. 1 Натяжение, развиваемое во время изометрических сокращений, обычно меньше в медленных, чем в быстрых волокнах, хотя эта разница намного меньше, чем для скорости укорачивания. 4
Рисунок 1Сократительные и энергетические свойства отдельных мышечных волокон мускулов человека (латеральная широкая мышца бедра). (A) Соотношение силы и скорости трех репрезентативных одиночных мышечных волокон, содержащих изоформы MyHC-1 (непрерывная линия), MyHC-2A (пунктирная линия) и MyHC-2X (пунктирная линия). Скорость и сила увеличиваются от медленного → 2A → 2X. (B) Средние значения скорости потребления АТФ во время изометрических сокращений (активность АТФазы) трех отдельных волокон, содержащих изоформы MyHC-1, MyHC-2A и MyHC-2X.Активность АТФазы волокон также увеличивается при переходе от медленного → 2A → 2X.
Быстрые и медленные волокна также различаются во времени сократительной реакции — то есть во время сокращения повышенное напряжение достигает пика и снижается быстрее в быстрых, чем в медленных волокнах (таблица 1). Временные параметры сокращения определяются (1) скоростью, с которой кальций высвобождается и поглощается саркоплазматической сетью, (2) чувствительностью миофибрилл к кальцию и (3) скоростью, с которой миозин формирует силу. образование связей (поперечных мостиков) с актином.В свою очередь, скорость высвобождения и поглощения кальция зависит от свойств саркоплазматического ретикулума: плотность высвобождающих единиц и плотность кальциевых насосов выше в быстрых волокнах, чем в медленных волокнах 5– 8 и конкретных изоформах кальциевого насоса (SERCA) выражаются в медленных и быстрых волокнах. 9
Медленные волокна обычно более устойчивы к усталости, чем быстрые волокна, то есть они могут сохранять свои сократительные свойства в течение более длительного времени (таблица 1).Устойчивость к усталости зависит от баланса между производством и использованием энергии. Потребление АТФ для выработки механической выходной мощности, а также для контроля иона (первоначально концентрации кальция) соответствует производству АТФ. Производство АТФ происходит по двум основным метаболическим маршрутам: (1) анаэробный гликолиз, который генерирует одну АТФ и две молекулы лактата или пирувата из одной молекулы глюкозы, и (2) аэробное окисление в митохондриях, где 18 молекул АТФ производятся из одного ацетатного остатка (который образует либо от β-окисления жирных кислот или декарбоксилирования пирувата) только с диоксидом углерода и водой в качестве побочных продуктов.Для аэробного окисления требуется кислород, поэтому вокруг мышечных волокон и миоглобина имеется хорошо развитая капиллярная сеть для хранения кислорода в волокнах. Медленные волокна устойчивы к утомлению, потому что они потребляют мало АТФ, и это потребление может быть полностью компенсировано аэробным окислительным производством АТФ; таким образом, сократительная активность мало меняет состав цитозоля. Напротив, у быстрых волокон очень высока анаэробная метаболическая мощность; однако подкисление цитозоля из-за побочных продуктов анаэробного гликолиза (молекул лактата) подавляет сократительную реакцию и вызывает утомляемость.Потребление АТФ для сокращения быстрых волокон настолько велико, что не может сравниться с производством митохондриального АТФ, и значительное увеличение концентрации неорганического фосфата следует за сократительной активностью. Накопление лактата и неорганического фосфата вызывает усталость, поэтому потеря сократительной способности у быстрых волокон происходит быстрее, чем у медленных.
ТИПЫ ВОЛОКНА ДЫХАТЕЛЬНЫХ МЫШЦ
Наличие в дыхательных мышцах медленных и быстрых волокон отражает их функциональные задачи.В то время как в спокойном дыхании используются в основном медленные волокна, быстрые мышечные волокна задействуются именно при увеличении частоты дыхания — сдвиг в привлечении медленных мышечных волокон к быстрым во время тахипноэ был продемонстрирован в диафрагме и парастернальных мышцах кролика. 10, 11 Доля быстрых волокон в дыхательных мышцах выше у мелких лабораторных животных (мышей и крыс), чем у людей или крупных млекопитающих: это может отражать скорость вентиляции, которая снижается от мелких к крупным животным. 12, 13 В зависимости от характера активности дыхательные мышцы можно разделить на две основные группы. Во время вдоха можно регистрировать электромиографическую (ЭМГ) активность диафрагмы, лестничной мышцы, грудино-сосцевидных, парастернальных, внутренних межреберных и внешних межреберных мышц, в то время как во время выдоха внутренние межреберные мышцы и мышцы живота активны. Во время спокойного дыхания большая часть дыхательной работы выполняется диафрагмой, и другие дыхательные мышцы (вспомогательные дыхательные мышцы) становятся важными только тогда, когда этого требует увеличенная глубина вентиляции.Таким образом, диафрагма находится в постоянной ритмической активности, которая не допускает паузы для отдыха, поэтому волокна диафрагмы должны быть очень устойчивы к утомлению. Помимо вентиляции, дыхательные мышцы участвуют в других двигательных действиях, включая кашель, разговор и пение; эти действия носят поэтапный и эпизодический характер. Некоторые дыхательные мышцы также выполняют постуральную роль, предполагающую более длительную активность.
Наилучшие оценки распределения типов волокон в диафрагме взрослого человека показывают около 55% медленных волокон, 21% быстрых окислительных и 24% быстрых гликолитических. 14– 16 Распределение изоформ миозина показывает, что медленный миозин составляет менее 50% от общего количества, в то время как миозин 2A составляет около 40% (рис. 2A). Несоответствие в распределении типов волокон можно объяснить разным размером волокон каждого типа.
Рисунок 2Изоформный состав MyHC респираторных мышц человека в (A) диафрагме и (B) в наружной межреберной мышце. В обеих мышцах медленные (MyHC-1) и быстрые (MyHC-2A + MyHC-2X) MyHC экспрессируются почти в равных пропорциях.Данные взяты из Levine и др. 168 для диафрагмы и D’Antona (неопубликованное наблюдение) для наружной межреберной мышцы.
Сведения о межреберных мышцах более отрывочны. У человека доля медленных волокон превышает 60% (то есть немного выше, чем в диафрагме 16– 20 ) как во внутренних, так и во внешних межреберных мышцах. Распределение изоформ миозина в наружных межреберных мышцах (рис. 2В) аналогично распределению в диафрагме, несоответствие связано с большим размером волокон, содержащих быстрый миозин.
При сравнении у данного вида животных быстрые волокна 2Х и 2В имеют более низкую устойчивость к утомлению и более низкую аэробную окислительную способность, чем медленные и быстрые волокна 2А (таблица 1), на что указывает более низкая активность сукцинатдегидрогеназы или цитратсинтазы, обоих из которых являются ключевыми ферментами митохондриального метаболизма. 21– 23 Это различие также присутствует в дыхательных мышцах (диафрагма крысы, 24 диафрагма кошки 25 ), хотя существует большая вариабельность в каждом типе волокон, особенно в волокнах 2Х.Значительные различия наблюдаются в активности аэробных окислительных ферментов у разных видов. Окислительный метаболизм, измеряемый по активности сукцинатдегидрогеназы, выше у мелких животных прямо пропорционально частоте дыхания и доле быстрых волокон 12, 26 и обратно пропорционален экспрессии медленного миозина. 13 Таким образом, дыхательные мышцы мелких животных не только быстрее, но и обладают большей аэробной окислительной активностью, чем у крупных животных.Повышенный уровень аэробного окислительного метаболизма в респираторных мышцах приводит к повышенной сопротивляемости усталости, которая требуется при постоянной ритмической активности. Утомляемость дыхательных мышц ограничивает физическую работоспособность, как показано у спортсменов и пациентов с хронической обструктивной болезнью легких (ХОБЛ). 27, 28
Волокна диафрагмы обычно имеют меньшую площадь поперечного сечения, чем мышцы конечностей. Поскольку количество капиллярных сосудов, окружающих каждое волокно, одинаково, расстояние диффузии уменьшается, что делает подачу кислорода в диафрагму более эффективно, чем в другие мышцы. 16 Сообщалось об обратной зависимости между активностью аэробных окислительных ферментов и площадью поперечного сечения. 29 Это может улучшить диффузию кислорода и способствовать повышению устойчивости диафрагмы к усталости.
Недавние исследования также выявили особые особенности сцепления возбуждения-сокращения в волокнах диафрагмы по сравнению с другими дыхательными мышцами и мышцами задних конечностей. Реакция на такие агенты, как кофеин, которые вызывают высвобождение кальция без деполяризации мембраны, определенно больше в диафрагме, чем в других быстрых мышцах, и эта реакция находит свою молекулярную основу в экспрессии двух изоформ каналов высвобождения кальция саркоплазматического ретикулума — RyR1 и RyR3. 30, 31 В то время как RyR1 (который присутствует во всех скелетных мышцах) опосредует высвобождение кальция, вызванное деполяризацией мембраны, RyR3 (который экспрессируется только в диафрагме) опосредует индуцированное кальцием высвобождение кальция и более реагирует, чем RyR1, на стимуляцию кофеином. высвобождения кальция. 30– 32 Преимущество, которое дает волокнам диафрагмы такое специализированное расположение, все еще не ясно, хотя большая чувствительность диафрагмы к соединениям, таким как метилксантин, которые стимулируют сократимость мышц, можно объяснить присутствием RyR3.
РАЗВИТИЕ
Перинатальное развитие человека характеризуется значительным увеличением сократительной способности диафрагмы, о чем свидетельствует положительная корреляция между трансдиафрагмальным давлением (Pdi) и максимальным давлением на вдохе (Pimax) и возрастом после зачатия в диапазоне от 32 до 44 недель. . 33, 34 Подобное возрастающее увеличение сократительной силы было измерено in vitro на полосках диафрагмы крысы 35 и хомяка. 36 Из-за более позднего развития мелких грызунов по сравнению с людьми увеличение прочности диафрагмы можно легко наблюдать в раннем постнатальном периоде. Недавнее исследование 37 предоставило исчерпывающую картину механизма, лежащего в основе увеличения сократительной силы в диафрагме крысы. Взаимодействуют три различных фактора, а именно: (1) повышенная плотность миозина на саркомере, (2) замена медленного и неонатального миозина быстрым миозином и (3) более высокая сила, развиваемая отдельными молекулами миозина в волокнах, экспрессирующих быстрый миозин.Вероятно, аналогичные механизмы актуальны и для диафрагмы человека, где замена миозина неонатального миозина на взрослый медленный и быстрый миозин происходит быстро в перинатальном периоде — например, доля медленных волокон увеличивается с 9% к 27 неделе (срок беременности). ) до 25% в срок и достигает взрослого уровня во втором послеродовом году. 38 Увеличение медленной экспрессии миозина сопровождается увеличением окислительной способности, оцениваемой по активности НАДН-дегидрогеназы, что приводит к повышению устойчивости диафрагмы к утомлению. 38
ОБУЧЕНИЕ И ИСПОЛЬЗОВАНИЕ
Сократительная активность — один из наиболее эффективных стимулов, изменяющих размер и тип мышечных волокон. 39 У людей неиспользование вызывает уменьшение мышечной массы 40 , что становится очевидным всего через пару недель. 41 Хотя работа, проведенная на мышцах крыс и мышей, показала, что неиспользование вызывает сдвиг в сторону более быстрого мышечного фенотипа, 42 явное преобразование от медленного к быстрому типу волокон наблюдалось только у людей после строгой иммобилизации 43 в то время как в других моделях неиспользования (таких как космический полет и постельный режим) сдвиг вряд ли будет значительным. 44– 46 Интересно, что мышцы людей с параличом нижних конечностей развивают однородный состав быстрых волокон в течение 6–12 месяцев после травмы. 47
Считается, что два основных типа мышечных упражнений — тренировка на выносливость (длительные сеансы субмаксимальных сокращений) и тренировки с отягощениями (короткие сеансы максимальных сокращений 48 ) — приводят к различным адаптациям скелетных мышц. Тренировка на выносливость не увеличивает мышечную массу и силу 49 и усиливает активность аэробных ферментов, 50 , тогда как тренировки с отягощениями вызывают значительное увеличение мышечной массы и силы 51 с неопределенным влиянием на метаболические ферменты митохондрий. 52, 53 В мышцах конечностей человека тренировки на выносливость и сопротивление приводят к преобразованию волокон из типа 2X в 2A, 54– 58 , тогда как обратное преобразование (из 2A в 2X) не описано ни в одном парадигма упражнений 58, 59 и будет иметь место только при неиспользовании и иммобилизации. Трансформация в направлении от типа 1 к типу 2А была продемонстрирована после интенсивных тренировок на короткие дистанции 54, 56, 57 , а трансформация от 2А к 1 была зарегистрирована после тренировки на выносливость. 60
Дыхательные мышечные волокна не претерпевают изменений из-за тренировок и бездействия так же, как мышечные волокна конечностей. Был поставлен вопрос о том, можно ли тренировать дыхательные мышцы, и хроническая активность дыхательных мышц, кажется, предполагает, что реального неиспользования не может произойти. Наблюдение за тем, что вентиляция во время напряженных упражнений никогда не достигает уровня максимальной произвольной вентиляции и что вентиляция может быть увеличена, даже когда испытуемые тренируются выше анаэробного порога, в течение длительного времени привело к выводу, что дыхательная система «создана для упражнений». 61 и что дыхательные мышцы нельзя тренировать.Однако интерес к реакции дыхательных мышц на упражнения значительно вырос, поскольку было показано, что утомление дыхательных мышц может возникать во время тяжелых упражнений на выносливость 62 и что нарушение работы дыхательных мышц и мышц конечностей у пациентов с ХОБЛ является важным фактором. в определении снижения переносимости физических упражнений и качества жизни. 63
У крыс аэробная способность дыхательных мышц может увеличиваться после тренировки выносливости всего животного, 64– 68 , несмотря на более ранние данные, показывающие противоположный эффект. 69– 71 Увеличение аэробных возможностей наблюдалось не только для реберной диафрагмы, но и для голеностопной, 66 парастернальных, 66 и наружных межреберных мышц 70 . Интересно, что это увеличение происходит во всех типах волокон (1, 2A и 2X 64, 65, 72 и не вызвано сдвигом в составе типа волокна. Вряд ли какие-либо изменения в составе типа волокна после износостойкости тренировка фактически наблюдалась. 73, 74 Это еще одно свидетельство того, что в дыхательных мышцах метаболические ферменты и миофибриллярные белки не связаны строго, как показано в мышцах конечностей (см. Выше). Однако было указано, что реакция аэробного метаболизма дыхательных мышц на тренировку на выносливость, 72 хотя и ясна, но менее выражена, чем в мышцах конечностей. 75 Площадь поперечного сечения волокон уменьшается после тренировки на выносливость, и это может поддерживать наблюдаемое увеличение активности аэробных ферментов 64, 65 , определяющих увеличение концентрации митохондрий.
Тренировка инспираторных мышц у людей стала важной частью программы легочной реабилитации пациентов с ХОБЛ со слабостью инспираторных мышц. 76 Влияние тренировки инспираторных мышц на выполнение упражнений еще не установлено, 76 , и до сих пор ведется много споров о том, улучшаются ли показатели упражнений на выносливость за счет дыхательной тренировки у спортсменов 77, 78 и у субъектов. с ХОБЛ. 76, 79
Увеличение скорости укорочения диафрагмы может быть вызвано умеренной программой тренировок, но маловероятно, что это изменение отражает структурную или биохимическую адаптацию мышечных волокон. 80 Имеется мало прямой информации о структурных изменениях волокон дыхательных мышц человека в связи с дыхательной тренировкой. Недавнее исследование показало, что функциональное улучшение, вызванное 5-недельной программой инспираторных тренировок в группе пациентов с ХОБЛ, было связано с увеличением доли медленных волокон (примерно на 38%) и размера быстрых волокон (на примерно 21%) в наружных межреберных мышцах. 81
Несмотря на то, что дыхательные мышцы хронически активны, адаптация к уменьшенной механической нагрузке действительно может происходить и может быть причиной проблем, с которыми сталкиваются пациенты, отлученные от ИВЛ.Исследования на животных показали, что снижение силы диафрагмы наблюдается уже через 12 часов ИВЛ, а повреждение диафрагмальных миофибрилл — через 3 дня. В волокнах диафрагмы снижается регуляция инсулиноподобного фактора роста 1 (IGF-1) и мессенджеров MyoD / миогенина, а также повышается активность окислительного стресса и протеазы; атрофия волокон появляется через 1-2 дня контролируемой ИВЛ. 82
СТАРЕНИЕ
Старение влияет на силу, массу и структуру скелетных мышц.У человека сила скелетных мышц in vivo значительно снижается (на 20-40%) с возрастом, 83- 89 наиболее вероятным объяснением такого снижения является потеря мышечной массы. Мышцы могут потерять 20–30% своей массы к 70 годам. 87, 90 Считается, что уменьшение мышечной массы зависит от уменьшения как размера, так и количества мышечных волокон. Размер волокон скелетных мышц уменьшается у субъектов старше 70 лет с избирательной атрофией быстрых волокон. 85, 91 Частичная денервация мышечных волокон происходит с возрастом 92, 93 и, несмотря на частичную реиннервацию, обе двигательные единицы 92 и мышечные волокна 90, 93, 94 теряются при старении. Медленные двигательные единицы реиннервируют мышечные волокна более эффективно, чем быстрые двигательные единицы, и поэтому значительно увеличиваются в размерах. 92, 93 Это может указывать на общий сдвиг в сторону фенотипа медленных волокон.Однако есть несколько исследований, показывающих либо отсутствие изменений в распределении типов волокон 95, 96 , либо даже увеличение количества быстрых волокон. 97– 99 Возможное объяснение этому представлено сопутствующим неиспользованием мышц, связанным с общим снижением двигательной активности, типичным для старения. Повышенная доля медленных волокон не делает метаболизм мышц более окислительным. Максимальная окислительная способность снижается с возрастом в связи с ухудшением функции митохондрий. 100, 101
Имеется ограниченная информация о влиянии старения на дыхательные мышцы, особенно у людей. Более того, тот факт, что дыхательные мышцы должны быть хронически активными, предполагает, что старение может по-разному влиять на дыхательные мышцы и мышцы конечностей, и поэтому трудно применить информацию о мышцах конечностей к дыхательным мышцам. То, что старение действительно влияет на дыхательные мышцы, подтверждается значительным снижением прочности диафрагмы, наблюдаемым как у мелких млекопитающих 102, 103 , так и у людей. 104, 105 В соответствии с неопределенным влиянием старения на мышцы конечностей, van Lunteren и др. 106 обнаружили небольшой, но значительный сдвиг в сторону волокон типа 2B в диафрагме старых крыс, в то время как другие рабочие 103, 107 не сообщили об изменениях. Отсутствие атрофии отдельных мышечных волокон, наблюдаемое в мышцах конечностей, находит свое отражение в диафрагме без изменения площади поперечного сечения отдельных волокон у старых крыс. 106, 107 Наконец, неопределенное влияние старения на метаболические ферменты согласуется с отсутствием значительных изменений окислительной способности диафрагмы, наблюдаемых у старых крыс. 107– 109 Таким образом, до настоящего времени не было обнаружено специфического воздействия старения на дыхательные мышцы, несмотря на их особую функциональную роль.
МЫШЕЧНЫЕ ДИСТРОФИИ
Болезни, поражающие в первую очередь дыхательные мышцы, представляют собой мышечные дистрофии. Мутации генов, кодирующих цитоскелет, сарколемму, ядерную оболочку и внеклеточный матрикс, вызывают раннюю гибель мышечных волокон, которая, вероятно, опосредована потерей целостности мембраны.По неизвестным причинам группы мышц, которые отмечают начало заболевания, различаются в зависимости от мышечных дистрофий, но в основном представлены мышцами конечностей. Дыхательная недостаточность возникает поздно у пациентов с дистрофией Дюшена, Беккера и пояса конечностей, являясь одной из наиболее частых причин смерти, 110 и очень редко у пациентов с лицевой-лопаточно-плечевой мышечной дистрофией. 111 Врожденные миопатии, такие как врожденная миопатия Ульриха 112 , которые характеризуются ранним тяжелым респираторным нарушением, являются исключением.Специализация и особый паттерн активности дыхательных мышечных волокон могут объяснять их вариабельность и либо позднее, либо отсутствие вовлечения в мышечные дистрофии.
Нет прямой информации об изменениях волокон скелетных мышц дыхательных мышц у людей с мышечной дистрофией. Косвенные доказательства псевдогиптрофии диафрагмы у пациентов с мышечной дистрофией Дюшенна были получены в недавнем исследовании, в котором сочеталось ультразвуковое измерение толщины диафрагмы с регистрацией трансдиафрагмального давления. 113 В отличие от человеческого аналога, мышечная модель мышечной дистрофии Дюшенна (мыши MDX) характеризуется первичным поражением диафрагмы 114 , в то время как мышцы конечностей не имеют серьезных функциональных нарушений, сравнимых с клиническими симптомами, наблюдаемыми у мальчиков с условием. 115 Диафрагма мышей MDX содержит регенерирующие волокна, которые выражают MyHC в процессе развития, и имеет общий сдвиг в сторону более медленных типов волокон (1 и 2A) за счет самых быстрых типов волокон (2X и 2B) и очень значительной потери удельной силы. . 114 Нарушение диафрагмальной функции также было замечено на двух моделях мышечной дистрофии пояса конечностей: у хомяков Bio 14.6, лишенных дельта-саркогликана, 116 и у мышей с нулевым альфа-саркогликаном. 117 Несмотря на различное поражение дыхательных мышц и мышц конечностей, мышиные модели мышечных дистрофий человека предоставляют полезную информацию об их патогенезе и широко используются для тестирования возможных методов лечения.
Попытки спасти мышцы, пораженные мышечной дистрофией, в настоящее время предпринимаются по трем основным направлениям: 118
Генная терапия, направленная на доставку мутированного гена к дистрофическим мышечным волокнам аденовирусом и аденоассоциированными вирусами.Внутримышечная инъекция таких векторов определила экспрессию мини-дистрофина (полноразмерный дистрофин слишком большой, чтобы его можно было успешно переносить) у мышей mdx 119, 120 и восстановила экспрессию α-саркогликана 121 в α-саркогликане. нулевых мышей и дельта-саркогликана 122 у хомяков Bio 1.6. Однако длительная доставка трансгена по всему телу без иммунной реакции остается серьезной проблемой. 123
Фармакологическое лечение, основанное на наблюдении, что большинство мутаций дистрофина происходит в области молекулы, которая не является строго необходимой для функционирования, но просто останавливает транскрипцию до того, как может быть синтезирована жизнеспособная молекула.Гентамицин , 124, и олигонуклеотиды 125 были успешно использованы для определения пропуска экзонов и обеспечения транскрипции молекулы дистрофина почти нормальной длины и последовательности. Эффективность лечения гентамицином недавно подверглась сомнению 126 , и клинические испытания пока дали отрицательные результаты. 118
Клеточная терапия, которая направлена на доставку к дистрофическим мышцам сателлитных клеток или плюрипотентных стволовых клеток, которые могут дифференцироваться в скелетные мышцы, либо для восстановления дегенерирующих клеток скелетных мышц, либо для создания новых клеток скелетных мышц.Ограниченная способность к самообновлению и миграции сателлитных клеток и миобластов до сих пор не позволяла добиться значительного успеха. Внутриартериальная доставка нового класса ассоциированных с сосудами фетальных стволовых клеток (мезоангиобластов) у мышей с отсутствием α-саркогликана недавно показала резкое улучшение морфологии и функции нижележащих мышц, 127 открывая новые перспективы для клеток терапия.
Дыхательные мышцы до сих пор не подвергались лечению на животных моделях мышечных дистрофий.У мелких животных, таких как мыши, трудно достичь дыхательных мышц при внутримышечной инъекции или внутриартериальной доставке. Однако у людей доставка не является проблемой, и респираторные мышцы будут лечиться в дополнение к мышцам конечностей, когда будут установлены методы лечения людей.
ЗДОРОВЬЕ
Длительное недоедание / недоедание вызывает потерю массы тела параллельно с истощением скелетных мышц, при этом респираторные мышцы не щадят, причем диафрагма часто оказывается наиболее пораженной из респираторных мышц.Потеря веса диафрагмы — которая, по крайней мере, у грызунов, кажется, связана с полом, более заметным у мужчин 128 — пропорциональна потере веса других скелетных мышц (диафрагма атрофируется так же, как мышцы конечностей 129 ) или даже превышает потерю массы тела, как видно из некроскопических исследований недоедающих субъектов. 130 Неясно, происходит ли потеря мышечной массы из-за уменьшения площади поперечного сечения волокон или из-за уменьшения количества волокон. 131, 132 Некоторые исследования показали, что количество волокон снижается из-за голодания, 133 другие не сообщили об изменениях 131, 132 и другие, не исключая при этом возможное влияние лишения пищи на общую клеточность , показали, что недоедание в первую очередь влияет на размер клетчатки. 129
Несколько исследований показали избирательное влияние недостаточного питания на быстрые волокна как в дыхательных 134– 136 , так и в мышцах задних конечностей 131, 137, 138 , которые, по всей видимости, атрофируются исключительно 134, 135 или в большей степени 129, 131, 136, 139 , чем медленные волокна.Изменение состава диафрагмальных волокон в сторону более медленного фенотипа, вызванное недостаточным питанием 134 может быть частично связано с переходом от быстрых волокон к медленным, вызванным увеличением циркулирующих глюкокортикоидов в результате голодания 140 и частично с тем, что медленное волокна более активны, чем быстрые волокна. 141
Потеря мышечной массы и более сильное влияние недостаточного питания на быстрые волокна оказывают значительное влияние на сократительные и утомляющие свойства диафрагмы.Значительное снижение как пиковой силы сокращения (Pt), так и пикового тетанического напряжения (Po) полосок диафрагмы без изменений удельной силы (Pt и Po с поправкой на вес мышцы диафрагмы или площадь поперечного сечения), а также увеличение времени полураспада подергивания (RT 0,5 , время, когда Pt упадет до 0,5 от максимума) и о повышении Pt / Po. 129, 142– 145 Отсутствие вызванных голоданием изменений удельной силы предполагает, что падение генерации силы, наблюдаемое в диафрагме, должно быть связано исключительно с уменьшением мышечной массы, а не с сопутствующими изменениями миофибриллярного аппарата. .Кроме того, наблюдаемые изменения RT 0,5 и Pt / Po могут быть связаны с увеличением доли медленных волокон. 129, 144, 145 В то время как сопротивление низкочастотной усталости увеличивается, 129, 144– 146 сопротивление высокочастотной усталости снижается 144 , и эти изменения могут быть обнаружены путем оценки сократимости диафрагмы посредством непрямой диафрагмальной стимуляции. 142, 143
ГЛЮКОКОРТИКОИДЫ И БЕТА-АГОНИСТЫ
Некоторые фармакологические соединения могут изменять размер и / или тип волокон скелетных мышц, из которых глюкокортикоиды и β-агонисты имеют отношение к лечению респираторных заболеваний.
Постоянное употребление больших доз глюкокортикоидов вызывает атрофию и слабость мышц, состояние, известное как стероидная миопатия, которое хорошо известно у людей и животных. Нарушение сократимости может быть достаточно серьезным, чтобы снизить двигательную активность и дыхательную функцию. 147, 148 Стероидная миопатия характеризуется как в мышцах животных, так и человека атрофией волокон, которая преимущественно поражает быстрые волокна, сдвигом от быстрой к медленной экспрессии изоформы миозина и увеличением вариации площади поперечного сечения волокон с несколькими атрофическими углами. волокна и некротические волокна. 148, 149 Поскольку возможным объяснением относительного сопротивления медленных волокон является их более устойчивая активность, можно предположить, что мышечные волокна в дыхательных мышцах поражаются меньше, чем в мышцах конечностей. Однако типичные признаки стероидной миопатии также хорошо документированы в диафрагме крыс 149, 150 и хомяков 151 и в межреберных мышцах крыс. 149
Хроническое введение β-агонистов для расслабления гладких мышц дыхательных путей вызывает гипертрофию скелетных мышц, которая более выражена в быстрых, чем в медленных волокнах, и переход от медленных к быстрым изоформ миозина.Эффект проявляется в мышцах конечностей и дыхательных мышцах различных видов животных. 152– 154 Таким образом, бета-агонисты улучшают сократительную способность дыхательных мышц, как показано на примере хомяков 155 и крыс. 156 Следует помнить, что усиление сократительной способности диафрагмы также следует за острым введением некоторых β-агонистов, таких как сальбутамол. 156
Интересно, что острый эффект β-адренергической активации имеет противоположные эффекты в быстрых и медленных мышцах, 157 амплитуда и продолжительность сокращений увеличивается в быстрых мышцах и уменьшается в медленных.Это означает, что как хроническое, так и острое введение β-агонистов оказывает различное воздействие на медленные и быстрые волокна. Распределение β 2 адренорецепторов неоднородно. В мышцах задних конечностей имеется 85% рецепторов β 2 и 15% рецепторов β 1 , тогда как в диафрагме присутствуют только рецепторы β 2 . Следовательно, можно ожидать, что диафрагма будет более чувствительной к агонистам β 2 , чем мышцы задних конечностей. Кроме того, плотность рецепторов β 2 больше в медленных волокнах, чем в быстрых волокнах. 158
ХРОНИЧЕСКАЯ ОБСТРУКТИВНАЯ БОЛЕЗНЬ ЛЕГКИХ
Одна из первых жалоб пациентов с ХОБЛ — снижение толерантности к физической нагрузке, частично вызванное структурными и функциональными нарушениями скелетных мышц. Анализ биоптатов четырехглавой мышцы бедра у пациентов с ХОБЛ показал значительную атрофию волокон с уменьшением доли медленных волокон, сопровождаемым увеличением быстрых волокон 2Х. 159, 160 Напротив, никаких изменений в двуглавой мышце плеча у пациентов с ХОБЛ по сравнению с контрольной группой того же возраста, 161 и сила захвата менее снижена, чем сила четырехглавой мышцы. 162 Значительно повышенная экспрессия MyHC-2X и быстрых изоформ MyLC наблюдалась в латеральной широкой мышце бедра у пациентов с ХОБЛ по сравнению с контрольной группой. 163 Интересно, что плотность капилляров резко снижается, а концентрация миоглобина снижается в мышцах пациентов с ХОБЛ, 164 и активность окислительных аэробных ферментов также снижается. 165, 166 Липофусцин накапливается в волокнах, что позволяет предположить, что окислительное повреждение мышечной ткани может быть связано с дисфункцией скелетных мышц и истощением при ХОБЛ. 167
Пациенты с ХОБЛ имеют более высокий процент медленных MyHC (рис. 3) и более низкий процент быстрых MyHC-2A и MyHC-2X в диафрагме, чем в контрольной группе. 168 Кроме того, диафрагма пациентов с ХОБЛ содержит более высокий процент медленных изоформ MyLC, тропонинов и тропомиозина, чем диафрагма контрольных субъектов. 168 Окислительная способность митохондрий относительно потребности в АТФ (SDH / mATPase) выше в каждом из типов волокон диафрагмы у пациентов с ХОБЛ, чем в контрольной группе, что указывает на повышенную устойчивость к усталости. 169 По-видимому, повышенная функциональная потребность вызывает адаптацию волокон диафрагмы у пациентов с ХОБЛ. Удивительно, но развивающиеся (эмбриональные и неонатальные) изоформы MyHC были обнаружены в диафрагмах пациентов с тяжелой ХОБЛ, но значение этого еще предстоит выяснить. 170 Ультраструктурные исследования также выявили субклеточные изменения в диафрагме пациентов с ХОБЛ, включая уменьшение средней длины саркомера в состоянии покоя и увеличение концентрации митохондрий. 171 Изменения в мышцах выдоха могут отличаться от изменений в диафрагме: в недавнем исследовании было показано снижение силы и выносливости. 172
Рисунок 3Относительная доля изоформы MyHC-1 в образцах биопсии диафрагмы и латеральной широкой мышцы бедра здоровых субъектов (CTR) и пациентов с хронической обструктивной болезнью легких (ХОБЛ). Данные Levine и др. 168 для мышцы диафрагмы и Satta и др. 163 для латеральной широкой мышцы бедра; * р <0.05.
ГИПОКСИЯ
Нормобарическая гипоксия — частое осложнение ряда респираторных заболеваний. В то время как острая тяжелая гипоксия вызывает дыхательную недостаточность, связанную с серьезным повреждением дыхательных мышечных волокон в течение нескольких часов, 173 хроническая гипоксия вызывает умеренную адаптацию мышечных волокон.
В мышцах конечностей крысы адаптация к хронической гипоксии вызывает уменьшение медленных волокон и увеличение быстрых волокон и сдвиг в сторону более гликолитического метаболизма с течением времени в течение многих недель. 174– 176 Эти изменения могут быть связаны с малоподвижностью и анорексией, которые также вызваны недостатком кислорода, но они также наблюдались у крыс, подвергшихся тренировкам на выносливость в условиях гипоксии. 174 Диафрагма, похоже, не отличается от мышц конечностей в своей адаптации к гипоксии, хотя недавнее исследование показало, что периодическая гипоксия вызывает адаптивные реакции в диафрагме, которые могут повысить ее устойчивость к острой аноксии. 177
У пациентов с хронической гипоксией из-за ХОБЛ доля быстрых волокон увеличена, а доля медленных волокон снижена, 160 , но трудно с уверенностью приписать этот эффект только гипоксии.У альпинистов, подвергавшихся в течение нескольких недель высотной гипоксии, наблюдалась миопатия, характеризующаяся значительным уменьшением размера мышечных волокон (-20%) и потерей окислительной способности мышц (-25%); объем митохондрий уменьшается, и липофусцин накапливается в цитозоле волокон. 178, 179 Эти данные были получены из образцов биопсии латеральной широкой мышцы бедра, и неизвестно, происходят ли аналогичные изменения и в дыхательных мышцах.
ВЫВОДЫ
Картина структурных и функциональных особенностей дыхательных мышечных волокон, которая появляется из доступной в настоящее время информации, далека от полной (таблица 2), хотя были описаны признаки их специализированной работы и их адаптации к нескольким условиям, которые изменяют функциональные требования. в литературе.Некоторая описательная информация по-прежнему отсутствует, например, касающаяся возрастных изменений или влияния тренировок на дыхательные мышцы человека. Новые инструменты протеомики и транскриптомики еще не применялись для изучения дыхательных мышечных волокон, а анализ сигналов, регулирующих транскрипцию генов и посттрансляционную модификацию белков, еще не проводился.
Таблица 2Сводка изменений, происходящих в волокнах дыхательных мышц в связи с изменениями физиологических условий, заболеваний и фармакологического лечения
ССЫЛКИ
- ↵
Скьяффино S , Реджиани К.Молекулярное разнообразие миофибриллярных белков: регуляция генов и функциональное значение. Physiol Rev 1996; 76: 371–423.
- ↵
Bottinelli R , Reggiani C. Волокна скелетных мышц человека: молекулярное и функциональное разнообразие. Программа Biophys Mol Biol2000; 73: 195–262.
- ↵
Campbell WG , Gordon SE, Carlson CJ, et al. Дифференциальная глобальная экспрессия генов в красных и белых скелетных мышцах.Am J Physiol Cell Physiol 2001; 280: C763-8.
- ↵
He Z — H , Боттинелли Р., Пеллегрино М.А., и др. Потребление АТФ и эффективность отдельных мышечных волокон человека с различным изоформным составом миозина. Biophys J2000; 79: 945–61.
- ↵
Everts ME , Andersen J, Clausen T, et al. Количественное определение Са 2+ -зависимой Mg 2+ -АТФазы из саркоплазматического ретикулума в биопсиях мышц.Biochem J1989; 260: 443–8.
Leberer E , Pette D. Иммунохимическая количественная оценка Са-АТФазы саркоплазматического ретикулума, кальсеквестрина и парвальбумина в скелетных мышцах кролика с определенным составом волокон. Eur J Biochem 1986; 156: 489–96.
Wu KD , Lytton J. Молекулярное клонирование и количественная оценка изоформ Са (2 +) — АТФазы саркоплазматического ретикулума в мышцах крыс.Am J Physiol1993; 264: C331–41.
- ↵
Францини-Армстронг C , Протаси Ф., Рамеш В. Формы, размеры и распределение высвобождающих единиц и куплонов Ca 2+ в различных скелетных и сердечных мышцах. Biophys J1999; 77: 1528–39.
- ↵
Lytton J , Westlin M, Burk SE, и др. Функциональные сравнения изоформ семейства кальциевых насосов саркоплазматического или эндоплазматического ретикулума.J Biol Chem1992; 267: 14483–9.
- ↵
Citterio G , Agostoni E, Piccoli S, et al. Выборочная активация парастернальных мышечных волокон в зависимости от частоты дыхания. Respir Physiol 1982; 48: 281–95.
- ↵
Citterio G , Sironi S, Piccoli S, и др. Сдвиг от медленного к быстрому в мышечных волокнах вдоха во время теплового тахипноэ. Respir Physiol 1983; 51: 259–74.
- ↵
Blank S , Chen V, Iannuzzo C. Биохимические характеристики диафрагм млекопитающих. Respir Physiol 1988; 74: 115–26.
- ↵
Hodge K , Powers SK, Coombes J, et al. Биоэнергетические характеристики реберной и бедренной диафрагмы млекопитающих. Respir Physiol1997; 109: 149–54.
- ↵
Либерман Д.А. , Фолкнер Дж. А., Крейг А.Б., и др. Характеристики и гистохимический состав диафрагмы морской свинки и человека. J Appl Physiol 1973; 34: 233–7.
Sanchez J , Medrano G, Debesse B, et al. Типы мышечных волокон реберной и голевой диафрагмы у нормальных мужчин и пациентов с умеренными хроническими респираторными заболеваниями. Bull Eur Physiopathol Respir 1985; 21: 351–6.
- ↵
Mizuno M .Дыхательные мышцы человека: морфология волокон и капиллярное снабжение. Eur Respir J1991; 4: 587–601.
Ramirez-Sarmiento A , Orozco-Levi M, Guell R, et al. Тренировка дыхательных мышц у пациентов с хронической обструктивной болезнью легких: структурная адаптация и физиологические исходы. Am J Respir Crit Care Med, 2002; 166: 1491–7.
- ↵
Ruff RL , Whittlesey D.Плотность тока Na + и зависимость от напряжения в межреберных мышечных волокнах человека. J. Physiol1992; 458: 85–97.
- ↵
Keens TG , Bryan AC, Levison H, et al. Образец развития типов мышечных волокон в дыхательных мышцах человека. J Appl Physiol 1978; 44: 909–13.
- ↵
Keens TG , Chen V, Patel P, et al. Клеточные адаптации дыхательных мышц к хронической повышенной респираторной нагрузке.J Appl Physiol 1978; 44: 905–8.
- ↵
Burke RE , Levine DN, Tsairis P, et al. Физиологические типы и гистохимические профили моторных единиц икроножной мышцы кошки. J. Physiol1973; 234: 723–48.
Burke RE , Levine DN, Zajac FE. Двигательные единицы млекопитающих: физиолого-гистохимическая корреляция трех типов в икроножной мышце кошки. Science 1971; 174: 709–12.
- ↵
Кугельберг E . Гистохимический состав, скорость сокращения и утомляемость двигательных единиц камбаловидной мышцы крыс. J Neurol Sci 1973; 20: 177–98.
- ↵
Sieck GC , Blanco CE. Постнатальные изменения в распределении активности сукцинатдегидрогеназы среди мышечных волокон диафрагмы. Педиатр Res1991; 29: 586–93.
- ↵
Sieck GC , Fournier M, Prakash YS, et al. Фенотип миозина и вариабельность фермента SDH среди волокон двигательных единиц. J Appl Physiol 1996; 80: 2179–89.
- ↵
Green HJ , Reichmann H, Pette D. Межвидовые и внутривидовые сравнения распределения типов волокон и активности сукцинатдегидрогеназы в волокнах типов I, IIA и IIB диафрагм млекопитающих. Гистохимия 1984; 81: 67–73.
- ↵
Фитинг JW . Усталость дыхательных мышц, ограничивающая физические упражнения? Eur Respir J1991; 4: 103–8.
- ↵
Johnson BD , Aaron EA, Babcock MA, et al. Усталость дыхательных мышц во время упражнений: влияние на производительность. Med Sci Sports Exerc1996; 28: 1129–37.
- ↵
Green HJ , Plyley MJ, Smith DM, et al. Тренировка на экстремальную выносливость и адаптация типа волокна в диафрагме крысы. J Appl Physiol1989; 66: 1914–20.
- ↵
Bertocchini F , Ovitt CE, Conti A, et al. Потребность в рецепторе рианодина типа 3 для эффективного сокращения скелетных мышц новорожденных. Embo J1997; 16: 6956–63.
- ↵
Росси Р. , Боттинелли Р., Соррентино В., и др. Ответ на изоформы рецепторов кофеина и рианодина в скелетных мышцах мышей. Am J Physiol Cell Physiol 2001; 281: C585–94.
- ↵
Conklin MW , Ahern CA, Vallejo P, et al. Сравнение искр Ca (2+), независимо продуцируемых двумя изоформами рианодинового рецептора (тип 1 или тип 3). Biophys J2000; 78: 1777–85.
- ↵
Dimitriou G , Greenhough A, Rafferty GF, et al. Влияние зрелости на максимальное трансдиафрагмальное давление у младенцев во время плача. Am J Respir Crit Care Med, 2001; 164: 433–6.
- ↵
Dimitriou G , Greenhough A, Moxham J, et al. Влияние созревания на функцию диафрагмы младенца, оцениваемую с помощью магнитной стимуляции диафрагмальных нервов. Педиатр Пульмонол, 2003; 35: 17–22.
- ↵
Johnson BD , Wilson LE, Zhan WZ, et al. Сократительные свойства развивающейся диафрагмы коррелируют с фенотипом тяжелой цепи миозина. J Appl Physiol1994; 77: 481–7.
- ↵
Coirault C , Lambert F, Joseph T, et al. Изменения в развитии свойств поперечного мостика и изоформы миозина в диафрагме хомяка. Am J Respir Crit Care Med, 1997; 156: 959–67.
- ↵
Geiger PC , Коди MJ, Macken RL, et al. Механизмы, лежащие в основе повышенной генерации силы волокнами диафрагмы крысы во время развития. J Appl Physiol2001; 90: 380–8.
- ↵
Keens TG , Bryan AC, Levison H, et al. Образец развития типов мышечных волокон в дыхательных мышцах человека. J Appl Physiol 1978; 44: 909–13.
- ↵
Болдуин К.М. , Хаддад Ф. Пластичность скелетных мышц: клеточные и молекулярные ответы на измененные парадигмы физической активности. Am J Phys Med Rehabil, 2002; 81: S40–51.
- ↵
LeBlanc A , Rowe R, Evans H, et al. Атрофия мышц при длительном постельном режиме.Int J Sports Med, 1997; 18 (Приложение 4): S283–5.
- ↵
Наричи М.В. , Кайзер Б., Бараттини П., и др. Изменения электрически вызванных сокращений скелетных мышц во время 17-дневного космического полета и постельного режима. Int J Sports Med, 1997; 18 (Приложение 4): S290–2.
- ↵
Stevens L , Mounier Y. Доказательства медленных и быстрых изменений сократительных белков камбаловидной мышцы крысы после подвешивания задних конечностей: исследования волокон с кожурой.Физиолог 1990; 33: S90–1.
- ↵
Hortobagyi T , Dempsey L, Fraser D, et al. Изменения силы мышц, размера мышечных волокон и экспрессии миофибриллярных генов у людей после иммобилизации и переподготовки. J. Physiol2000; 524: 293–304.
- ↵
Edgerton VR , Zhou MY, Ohira Y, et al. Размер человеческих волокон и ферментативные свойства после 5 и 11 дней космического полета.J Appl Physiol1995; 78: 1733–9.
Фитинги RH , Riley DR, Widrick JJ. Функциональные и структурные адаптации скелетных мышц к микрогравитации. J Exp Biol2001; 204: 3201–8.
- ↵
Berg HE , Larsson L, Tesch PA. Функция скелетных мышц нижних конечностей после 6 недель постельного режима. J Appl Physiol 1997; 82: 182–8.
- ↵
Round JM , Barr FM, Moffat B, et al. Участки волокон и типы гистохимических волокон в четырехглавой мышце у пациентов с параличом нижних конечностей. J Neurol Sci, 1993; 116: 207–11.
- ↵
Стенд FW , Thomason DB. Молекулярная и клеточная адаптация мышц в ответ на упражнения: перспективы различных моделей. Physiol Rev1991; 71: 541–85.
- ↵
Holloszy JO , будка FW. Биохимическая адаптация к упражнениям на выносливость в мышцах.Анну Рев Physiol 1976; 38: 273–91.
- ↵
Baldwin KM , Klinkerfuss GH, Terjung RL, et al. Дыхательная способность белой, красной и промежуточной мышцы: адаптивный ответ на упражнение. Am J Physiol1972; 222: 373–8.
- ↵
Хаккинен К , Ален М, Коми ПВ. Изменения изометрической силы и времени релаксации, электромиографических характеристик и характеристик мышечных волокон скелетных мышц человека во время силовых тренировок и разгрузки.Acta Physiol Scand 1985; 125: 573–85.
- ↵
MacDougall JD , Sale DG, Moroz JR, et al. Объемная плотность митохондрий в скелетных мышцах человека после тяжелых тренировок с отягощениями. Med Sci Sports, 1979; 11: 164–6.
- ↵
Saltin B , Голлник, PD. Адаптивность скелетных мышц: значение для метаболизма и работоспособности. В кн .: Справочник по физиологии скелетных мышц.Бетесда, Мэриленд: Американское физиологическое общество, 1983: 555–631.
- ↵
Allemeier CA , Fry AC, Johnson P, et al. Влияние тренировки цикла спринта на скелетные мышцы человека. J Appl Physiol1994; 77: 2385–90.
Andersen JL , Klitgaard H, Saltin B. Изоформы тяжелой цепи миозина в отдельных волокнах из m. Wastus lateralis спринтеров: влияние тренировки, Acta Physiol Scand, 1994; 151: 135–42.
- ↵
Cadefau J , Casademont J, Grau JM, et al. Биохимическая и гистохимическая адаптация к спринтерской тренировке юных спортсменов. Acta Physiol Scand1990; 140: 341–51.
- ↵
Esbjornsson M , Hellsten-Westing Y, Balsom PD, et al. Тип мышечных волокон изменяется во время спринтерской тренировки: влияние схемы тренировки. Acta Physiol Scand1993; 149: 245–6.
- ↵
Pette D , Staron RS. Переходы типов волокон в скелетных мышцах млекопитающих. Int Rev Cytol1997; 170: 143–223.
- ↵
Харридж SD , Боттинелли Р., Канепари М., и др. Спринтерская тренировка, функция мышц in vitro и in vivo и экспрессия тяжелых цепей миозина. J Appl Physiol 1998; 84: 442–9.
- ↵
Baumann H , Jaggi M, Soland F, et al. Тренировка с физической нагрузкой вызывает переход субъединиц изоформы миозина в мышечные волокна человека с гистохимическим типом. Пфлюгерс Арк 2987; 409: 349–60.
- ↵
Демпси JA . Мемориальная лекция Дж. Б. Вольфа. Легкие созданы для упражнений? Медико-научные спортивные упражнения 1986; 18: 143–55.
- ↵
Johnson BD , Babcock MA, Suman OE, et al. Утомление диафрагмы у здоровых людей, вызванное физической нагрузкой.J. Physiol1993; 460: 385–405.
- ↵
Рочестер DF . Диафрагма при ХОБЛ. Лучше, чем ожидалось, но недостаточно. N Engl J Med1991; 325: 961–2.
- ↵
Пауэрс SK , Criswell D, Lieu FK, et al. Специальная адаптация диафрагмальных волокон к упражнениям на выносливость. Респир Физиол 1992; 89: 195–207.
- ↵
Пауэрс SK , Criswell D, Lieu FK, et al. Клеточные изменения диафрагмы, вызванные физической нагрузкой. Am J Physiol1992; 263: R1093–8.
- ↵
Пауэрс СК , Крисвелл Д., Лоулер Дж., и др. Региональные изменения диафрагмальных окислительных и антиоксидантных ферментов, вызванные тренировками. Respir Physiol1994; 95: 227–37.
Пауэрс СК , Крисвелл Д., Лоулер Дж., и др. Влияние физических упражнений и типа волокон на активность антиоксидантных ферментов в скелетных мышцах крыс.Am J Physiol1994; 266: R375–80.
- ↵
Дадли GA , Abraham WM, Terjung RL. Влияние интенсивности и продолжительности упражнений на биохимические адаптации скелетных мышц. J Appl Physiol 1982; 53: 844–50.
- ↵
Fregosi RF , Sanjak M, Paulson DJ. Тренировка на выносливость не влияет на митохондриальное дыхание диафрагмы. Respir Physiol1987; 67: 225–37.
- ↵
Зеленый HJ , Райхманн Х.Дифференциальный ответ активности ферментов в диафрагме и межреберных мышцах крыс на тренировку. J Neurol Sci 1988; 84: 157–65.
- ↵
Metzger JM , Фитинги RH. Сократительные и биохимические свойства диафрагмы: эффекты тренировок и утомления. J Appl Physiol 1986; 60: 1752–8.
- ↵
Gosselin LE , Betlach M, Vailas AC, et al. Изменения в мышцах диафрагмы молодых и стареющих крыс, вызванные тренировками.J Appl Physiol 1992; 72: 1506–11.
- ↵
Сугиура Т , Моримото А., Мураками Н. Влияние тренировки на выносливость на изоформы тяжелых цепей миозина и активность ферментов в диафрагме крысы. Pflugers Arch2992; 421: 77–81.
- ↵
Sugiura T , Morimoto A, Sakata Y, et al. Изменения изоформы тяжелой цепи миозина в диафрагме крысы индуцируются тренировкой на выносливость.Jpn J Physiol1990; 40: 759–63.
- ↵
Halseth AE , Fogt DL, Fregosi RF, et al. Метаболические реакции дыхательных мышц крыс на произвольную тренировку. J Appl Physiol1995; 79: 902–7.
- ↵
Lotters F , van Tol B, Kwakkel G, et al. Эффекты контролируемой тренировки инспираторных мышц у пациентов с ХОБЛ: метаанализ. Eur Respir J2002; 20: 570–6.
- ↵
Romer LM , McConnell AK, Jones DA. Влияние тренировки инспираторных мышц на результаты тренировок на время у тренированных велосипедистов. J Sports Sci, 2002; 20: 547–62.
- ↵
Williams JS , Wongsathikun J, Boon SM, et al. Тренировка дыхательных мышц не способствует повышению выносливости у спортсменов. Med Sci Sports Exerc2002; 34: 1194–8.
- ↵
Sturdy G , Hillman D, Green D, et al. Возможность высокоинтенсивной интервальной тренировки дыхательных мышц при ХОБЛ. Chest2003; 123: 142–50.
- ↵
Darnley GM , Gray AC, McClure SJ, et al. Влияние резистивного дыхания на способность к физической нагрузке и функцию диафрагмы у пациентов с ишемической болезнью сердца. Eur J Heart Fail, 1999; 1: 297–300.
- ↵
Ramirez-Sarmiento A , Orozco-Levi M, Guell R, et al. Тренировка дыхательных мышц у пациентов с хронической обструктивной болезнью легких: структурная адаптация и физиологические исходы. Am J Respir Crit Care Med, 2002; 166: 1491–7.
- ↵
Gayan-Ramirez G , Decramer M. Влияние механической вентиляции на функцию диафрагмы и биологию. Eur Respir J2002; 20: 1579–86.
- ↵
Брюс С.А. , Ньютон Д., Уолледж RC.Влияние возраста на произвольную силу и площадь поперечного сечения приводящей мышцы большого пальца. QJ Exp Physiol1989; 74: 359–62.
Davies CT , White MJ, Young K. Электрически вызванное и произвольное максимальное изометрическое напряжение по отношению к динамической работе мышц у пожилых мужчин в возрасте 69 лет. Eur J Appl Physiol Occup Physiol 1983; 51: 37–43.
- ↵
Ларссон L , Гримби Г., Карлссон Дж.Сила мышц и скорость движения в зависимости от возраста и морфологии мышц. J Appl Physiol1979; 46: 451–6.
Ларссон Л . Морфофункциональная характеристика стареющих скелетных мышц человека. Поперечное исследование. Acta Physiol Scand Suppl1978; 457: 1–36.
- ↵
Young A , Stokes M, Crowe M. Размер и сила четырехглавых мышц пожилых и молодых мужчин.Clin Physiol 1985; 5: 145–54.
Young A , Stokes M, Crowe M. Размер и сила четырехглавой мышцы пожилых и молодых женщин. Eur J Clin Invest 1984; 14: 282–7.
- ↵
Наричи М.В. , Бордини М., Черретелли П. Влияние старения на функцию приводящей мышцы большого пальца человека. J Appl Physiol1991; 71: 1277–81.
- ↵
Гримби G , Солтин Б.Старение мышц. Clin Physiol 1983; 3: 209–18.
- ↵
Grimby G , Danneskiold-Samsoe B, Hvid K, et al. Морфология и ферментативная способность мышц рук и ног у мужчин и женщин в возрасте 78–81 лет. Acta Physiol Scand 1982; 115: 125–34.
- ↵
Кэмпбелл М.Дж. , МакКомас А.Дж., Петито Ф. Физиологические изменения стареющих мышц. J Neurol Neurosurg Psychiatry 1973; 36: 174–82.
- ↵
Лекселл Дж . Старение человека, мышечная масса и тип волокон. Журнал «Джеронтол», биол. И мед. Наука, 1995; 50: 11–6.
- ↵
Caccia MR , Харрис Дж. Б., Джонсон Массачусетс. Морфология и физиология скелетных мышц у стареющих грызунов. Мышечный нерв 1979; 2: 202–12.
- ↵
Klitgaard H , Mantoni M, Schiaffino S, et al. Функция, морфология и экспрессия белка стареющих скелетных мышц: перекрестное исследование пожилых мужчин с разным уровнем подготовки. Acta Physiol Scand1990; 140: 41–54.
- ↵
Aoyagi Y , Shephard RJ. Старение и функция мышц. Sports Med, 1992; 14: 376–96.
- ↵
Frontera WR , Hughes VA, Fielding RA, et al. Старение скелетных мышц: 12-летнее продольное исследование.J Appl Physiol 2000; 88: 1321–6.
Klitgaard H , Zhou M, Schiaffino S, et al. Старение изменяет состав тяжелых цепей миозина отдельных волокон скелетных мышц человека. Acta Physiol Scand1990; 140: 55–62.
- ↵
D’Antona G , Pellegrino MA, Adami R, et al. Влияние старения и иммобилизации на структуру и функцию волокон скелетных мышц человека.J. Physiol, 2003; 552: 499–511.
- ↵
Conley KE , Jubrian SA, Esselman PC. Окислительная способность и старение мышц человека. J. Physiol2000; 526: 203–10.
- ↵
Tonkonogi M , Fernstrom M, Walsh B, et al. У пожилых людей снижена окислительная способность, но не изменилась способность скелетных мышц. Пфлюгерс Arch3002; 446: 261–9.
- ↵
Criswell DS , Powers SK, Herb RA, et al. Механизм дефицита удельной силы в диафрагме стареющих крыс. Respir Physiol1997; 107: 149–55.
- ↵
Gosselin LE , Johnson BD, Sieck GC. Возрастные изменения сократительных свойств мышц диафрагмы и изоформ тяжелых цепей миозина. Am J Respir Crit Care Med, 1994; 150: 174–8.
- ↵
Polkey MI , Harris ML, Hughes PD, et al. Сократительные свойства диафрагмы человека пожилого возраста.Am J Respir Crit Care Med, 1997; 155: 1560–4.
- ↵
Tolep K , Higgins N, Muza S, et al. Сравнение прочности диафрагмы у здоровых взрослых пожилых людей и молодых людей. Am J Respir Crit Care Med, 1995; 152: 677–82.
- ↵
van Lunteren E , Vafaie H, Salomone RJ. Сравнительные эффекты старения на мышцы глотки и диафрагмы. Respir Physiol1995; 99: 113–25.
- ↵
Oliven A , Carmi N, Coleman R, et al. Возрастные изменения морфологических и окислительных свойств мышц верхних дыхательных путей. Exp Gerontol2001; 36: 1673–86.
Пауэрс СК , Лоулер Дж., Крисвелл Д., и др. Изменения диафрагмальных окислительных и антиоксидантных ферментов у стареющих крыс Fischer 344. J Appl Physiol 1992; 72: 2317–21.
- ↵
Пауэрс СК , Лоулер Дж., Крисвелл Д., и др. Старение и метаболическая пластичность дыхательных мышц: эффекты тренировки на выносливость. J Appl Physiol 1992; 72: 1068–73.
- ↵
Гозал Д . Легочные проявления нервно-мышечной болезни с особым упором на мышечную дистрофию Дюшенна и спинальную мышечную атрофию. Педиатр Пульмонол 2000; 29: 141–50.
- ↵
Padberg GW , Lunt PW, Koch M, et al. Диагностические критерии фасциально-плечевой мышечной дистрофии. Нейромышечное расстройство, 1991; 1: 231–4.
- ↵
Mercuri E , Yuva Y, Brown SC, et al. Участие коллагена VI в синдроме Ульриха: клиническое, генетическое и иммуногистохимическое исследование. Неврология, 2002; 58: 1354–9.
- ↵
De Bruin PF , Ueki J, Bush A, et al. Толщина диафрагмы и сила вдоха у пациентов с мышечной дистрофией Дюшенна. Thorax1997; 52: 472–5.
- ↵
Petrof BJ , Stedman HH, Shrager JB, et al. Адаптации в экспрессии тяжелой цепи миозина и сократительной функции в дистрофической диафрагме мыши. Am J Physiol1993; 265: C834–41.
- ↵
Stedman HH , Sweeney HL, Shrager JB, et al. Диафрагма мыши MDX воспроизводит дегенеративные изменения мышечной дистрофии Дюшенна. Nature1991; 352: 536–9.
- ↵
Burbach JA , Schlenker EH, Johnson JL. Морфометрия, гистохимия и сократимость дистрофической диафрагмы хомяка. Am J Physiol1987; 253: R275–84.
- ↵
Patel ND , Jannapureddy SR, Hwang W, et al. Изменение мышечной силы и жесткости скелетных мышц у мышей с дефицитом альфа-саркогликана.Am J Physiol Cell Physiol, 2003; 284: C962-8.
- ↵
Skuk D , Vilquin JT, Tremblay JP. Экспериментальные и терапевтические подходы к мышечным дистрофиям. Курр Опин Neurol2002; 15: 563–9.
- ↵
Harper SQ , Hauser MA, DelloRusso C, и др. Модульная гибкость дистрофина: значение для генной терапии мышечной дистрофии Дюшенна. Nat Med, 2002; 8: 253–61.
- ↵
Watchko J , O’Day T, Wang B, et al. Аденоассоциированный вирусный вектор-опосредованная генная терапия минидистрофином улучшает сократительную функцию дистрофических мышц у мышей MDX. Hum Gene Ther2002; 13: 1451–60.
- ↵
Allamand V , Donahue KM, Straub V, et al. Ранний перенос гена, опосредованный аденовирусом, эффективно предотвращает мышечную дистрофию у мышей с дефицитом альфа-саркогликана.Джин Ther2000; 7: 1385–91.
- ↵
Сяо X , Ли Дж., Цао Ю.П., и др. Полное функциональное восстановление полной мышцы (TA) у дистрофических хомяков с помощью генной терапии, направленной на вектор аденоассоциированного вируса. Дж. Вирол, 2000; 74: 1436–42.
- ↵
Грегоревич П , Чемберлен Дж. С.. Генная терапия мышечной дистрофии — обзор многообещающих успехов. Мнение эксперта Biol Ther2003; 3: 803–14.
- ↵
Barton-Davis ER , Cordier L, Shoturma DI, et al. Аминогликозидные антибиотики восстанавливают функцию дистрофина в скелетных мышцах мышей MDX. Дж. Клин Инвест, 1999; 104: 375–81.
- ↵
Lu QL , Mann CJ, Lou F, et al. Функциональные количества дистрофина, продуцируемого пропуском мутировавшего экзона у mdx-дистрофической мыши. Нат Мед2003; 9: 1009–14.
- ↵
Dunant P , Walter MC, Karpati G, et al. Гентамицин не увеличивает экспрессию дистрофина в мышцах с дефицитом дистрофина. Мышечный нерв 2003; 27: 624-7.
- ↵
Sampaolesi M , Torrente Y, Innocenzi A, et al. Клеточная терапия мышей с дистрофией альфа-саркогликан-нуль посредством внутриартериальной доставки мезоангиобластов. Science2003; 301: 487–92.
- ↵
Prezant DJ , Richner B, Aldrich TK, et al. Влияние длительного недоедания на сократительную способность диафрагмы самцов и самок крыс, утомляемость и типы волокон. J Appl Physiol1994; 76: 1540–7.
- ↵
Lewis MI , Sieck GC, Fournier M, et al. Влияние пищевой депривации на сократительную способность диафрагмы и размер мышечных волокон. J Appl Physiol 1986; 60: 596–603.
- ↵
Арора NS , Рочестер, округ Колумбия. Сила дыхательных мышц и максимальная произвольная вентиляция у пациентов с недостаточным питанием. Am Rev Respir Dis 1982; 126: 5–8.
- ↵
Goldspink DF . Влияние лишения пищи на белковый обмен и концентрацию нуклеиновых кислот в активных и иммобилизованных мышцах длинного разгибателя пальцев крысы. Biochem J1978; 176: 603-6.
- ↵
Парсонс Д. , Риди М., Мур Р.Л., и др. Острое голодание и количество волокон в камбаловидной мышце крысы. J Appl Physiol 1982; 53: 1234–8.
- ↵
Hegarty PV , Kim KO. Изменения клеточности скелетных мышц у голодных и голодных молодых крыс. Br J Nutr1980; 44: 123–7.
- ↵
Kelsen SG , Ference M, Kapoor S.Влияние длительного недоедания на структуру и функцию диафрагмы. J Appl Physiol 1985; 58: 1354–9.
- ↵
Lanz JK , Donahoe M, Rogers RM, et al. Влияние гормона роста на восстановление диафрагмы после истощения. J Appl Physiol 1992; 73: 801–5.
- ↵
Dekhuijzen PN , Gayan-Ramirez G, Bisschop A, et al. Лечение кортикостероидами и недостаток питания вызывают другую картину атрофии диафрагмы крысы.J Appl Physiol1995; 78: 629–37.
- ↵
Gardiner PF , Montanaro G, Simpson DR, и др. Влияние лечения глюкокортикоидами и ограничения пищи на мышцы задних конечностей крыс. Am J Physiol1980; 238: E124–30.
- ↵
Goodman MN , McElaney MA, Ruderman NB. Адаптация к длительному голоданию у крыс: сокращение протеолиза скелетных мышц. Am J Physiol1981; 241: E321–7.
- ↵
Oldfors A , Mair WG, Sourander P. Изменения мышц у молодых крыс, лишенных белка. Морфометрическое, гистохимическое и ультраструктурное исследование. J. Neurol Sci. 1983; 59: 291–302.
- ↵
Mlekusch W , Paletta B, Truppe W, et al. Концентрации глюкозы, кортикостерона, глюкагона и инсулина в плазме крови и содержание метаболических субстратов и ферментов в печени во время голодания и дополнительной гипоксии у крыс.Horm Metab Res 1981; 13: 612–4.
- ↵
Hennig R , Lomo T. Модели активации двигательных единиц у нормальных крыс. Nature1985; 314: 164–6.
- ↵
Dureuil B , Viires N, Veber B, et al. Острые изменения диафрагмы, вызванные голоданием у крыс. Am J Clin Nutr 1989; 49: 738–44.
- ↵
Льюис MI , Sieck GC.Влияние острого недоедания на структуру и функцию диафрагмы. J Appl Physiol 1990; 68: 1938–44.
- ↵
Prezant DJ , Valentine DE, Kim HH, et al. Влияние голодания и возобновления питания на сократительную способность диафрагмы взрослых самцов крыс, утомляемость и типы волокон. J Appl Physiol1993; 74: 742–9.
- ↵
Льюис М.И. , Лоруссо Т.Дж., Жан В.З., и др. Взаимодействующие эффекты денервации и недостаточности питания на структуру и функцию диафрагмы. J Appl Physiol 1996; 81: 2165–72.
- ↵
Koerts-De Lang E , Schols AM, Wouters EF, et al. Сократительные свойства и гистохимические характеристики диафрагмы крысы после длительного лечения триамцинолоном и депривации питания. J. Muscle Res Cell Motil, 1998; 19: 549–55.
- ↵
Decramer M , Стас К.Дж.Миопатия, вызванная кортикостероидами, затрагивающая дыхательные мышцы, у пациентов с хронической обструктивной болезнью легких или астмой. Am Rev Respir Dis 1992; 146: 800–2.
- ↵
Decramer M , де Бок В., Дом Р. Функциональная и гистологическая картина стероид-индуцированной миопатии при хронической обструктивной болезни легких. Am J Respir Crit Care Med 1996; 153: 1958–64.
- ↵
Polla B , Bottinelli R, Sandoli D, et al. Кортизон-индуцированные изменения в распределении тяжелой цепи миозина в дыхательных мышцах и мышцах задних конечностей. Acta Physiol Scand1994; 151: 353–61.
- ↵
Lieu FK , Powers SK, Herb RA, et al. Физические упражнения и диафрагмальная миопатия, вызванная глюкокортикоидами. J Appl Physiol1993; 75: 763–71.
- ↵
Льюис М.И. , Монн С.А., Зик Г.К. Влияние кортикостероидов на утомляемость диафрагмы, активность СДГ и размер мышечных волокон.J Appl Physiol 1992; 72: 293–301.
- ↵
Zeman RJ , Ludemann R, Easton TG, и др. Медленные или быстрые изменения в волокнах скелетных мышц, вызванные кленбутеролом, агонистом бета 2 -рецепторов. Am J Physiol1988; 254: E726–32.
Polla B , Cappelli V, Morello F, et al. Эффекты бета-2 агониста кленбутерола на дыхательные мышцы и мышцы конечностей крыс-отъемышей.Am J Physiol2001; 280: R862–9.
- ↵
Pellegrino MA , D’Antona G, Bortolotto S, et al. Кленбутерол противодействует вызванной глюкокортикоидами атрофии и трансформации типов волокон у мышей. Exp Physiol2004; 89: 89–100.
- ↵
Van Der Heijden HF , Dekhuijzen PN, Folgering H, et al. Долгосрочные эффекты кленбутерола на морфологию диафрагмы и сократительные свойства у эмфизематозных хомяков.J Appl Physiol 1998; 85: 215–22.
- ↵
Van der Heijden HFM , Zhan WZ, Prakash YS, et al. Сальбутамол усиливает изотонические сократительные свойства диафрагмальной мышцы крысы. J Appl Physiol1998; 85: 525–9.
- ↵
Bowman WC . Влияние адренергических активаторов и ингибиторов на скелетные мышцы. В: Szerekes L, ed. Адренергические активаторы и ингибиторы.Гейдельберг: Springer-Verlag, 1980: 47–128.
- ↵
Williams RS , Caron MG, Daniel K. Бета-адренорецепторы скелетных мышц: различия в зависимости от типа волокна и тренировок. Am J Physiol1984; 246: E160-7.
- ↵
Jakobsson P , Jorfeldt L, Brundin A. Метаболиты скелетных мышц и типы волокон у пациентов с запущенной хронической обструктивной болезнью легких (ХОБЛ) с хронической дыхательной недостаточностью и без нее.Eur Respir J1990; 3: 192–6.
- ↵
Hildebrand IL , Sylven C, Esbjornsson M, et al. Вызывает ли хроническая гипоксемия преобразование типов волокон? Acta Physiol Scand1991; 141: 435–9.
- ↵
Sato Y , Asoh T, Honda Y, и др. Морфологическая и гистохимическая оценка мышц у пациентов с хронической эмфиземой легких, проявляющейся генерализованным истощением.Eur Neurol1997; 37: 116–21.
- ↵
Gosselink R , Troosters T, Decramer M. Слабость периферических мышц способствует ограничению физических упражнений при ХОБЛ. Am J Respir Crit Care Med 1996; 153: 976–80.
- ↵
Satta A , Migliori GB, Spanevello A, et al. Типы волокон в скелетных мышцах пациентов с хронической обструктивной болезнью легких, связанные с функцией дыхания и толерантностью к физической нагрузке.Eur Respir J1997; 10: 2853–60.
- ↵
Whittom F , Jobin J, Simard PM, et al. Гистохимические и морфологические характеристики латеральной широкой мышцы бедра у пациентов с хронической обструктивной болезнью легких. Med Sci Sports Exerc1998; 30: 1467–74.
- ↵
Maltais F , LeBlanc P, Simard C, et al. Адаптация скелетных мышц к тренировкам на выносливость у пациентов с хронической обструктивной болезнью легких.Am J Respir Crit Care Med 1996; 154: 442–7.
- ↵
Jakobsson P , Jorfeldt L, Henriksson J. Активность метаболических ферментов в четырехглавой мышце бедра у пациентов с тяжелой хронической обструктивной болезнью легких. Am J Respir Crit Care Med, 1995; 151: 374–7.
- ↵
Allaire J , Maltais F, LeBlanc P, et al. Накопление липофусцина в латеральной широкой мышце у пациентов с хронической обструктивной болезнью легких.Мышечный нерв, 2002; 25: 383–9.
- ↵
Levine S , Kaiser L, Leferovich J, et al. Клеточные адаптации диафрагмы при хронической обструктивной болезни легких. N Engl J Med1997; 337: 1799–806.
- ↵
Левин С. , Грегори С., Нгуен Т., и др. Биоэнергетическая адаптация отдельных диафрагмальных миофибрилл человека к тяжелой форме ХОБЛ. J Appl Physiol, 2002; 92: 1205–13.
- ↵
Nguyen T , Shrager J, Kaiser L, et al. Тяжелые цепи миозина развития в диафрагме взрослого человека: паттерны коэкспрессии и эффект ХОБЛ. J Appl Physiol 2000; 88: 1446–56.
- ↵
Orozco-Levi M , Gea J, Lloreta JL, et al. Субклеточная адаптация диафрагмы человека при хронической обструктивной болезни легких. Eur Respir J1999; 13: 371–8.
- ↵
Ramirez-Sarmiento A , Orozco-Levi M, Barreiro E, et al. Выносливость выдыхательной мышцы при хронической обструктивной болезни легких. Thorax2002; 57: 132–6.
- ↵
Simpson JA , van Eyk JE, Iscoe S. Модификация тропонинов I и T, индуцированная гипоксемией, в диафрагме собаки. J Appl Physiol 2000; 88: 753–60.
- ↵
Бигард AX , Брюне А, Серрюрье Б, и др. Влияние тренировок на выносливость на большой высоте на свойства диафрагмальных мышц. Pflugers Arch2992; 422: 239–44.
Ито К. , Моритани Т., Исида К., и др. Преобразование типа волокон в мышцах задних конечностей крыс, вызванное гипоксией. Гистохимические и электромеханические изменения. Eur J Appl Physiol Occup Physiol 1990; 60: 331–6.
- ↵
Мортола JP , Насо Л.Электрофоретический анализ сократительных белков диафрагмы крыс с хронической гипоксией. Am J Physiol1995; 269: L371–6.
- ↵
Clanton TL , Klawitter PF. Приглашенный обзор: Адаптивные реакции скелетных мышц на перемежающуюся гипоксию: известное и неизвестное. J Appl Physiol2001; 90: 2476–87.
- ↵
Hoppeler H , Howald H, Cerretelli P. Структура мышц человека после воздействия экстремальной высоты.Experientia1990; 46: 1185–7.
- ↵
Howald H , Pette D, Simoneau JA, et al. Влияние хронической гипоксии на активность мышечных ферментов. Int J Sports Med, 1990; 11 (Приложение 1): S10–4.
РАСТЯГИВАНИЕ И ГИБКОСТЬ — Физиология растяжения
Перейти к предыдущей, следующей главе.
Цель этой главы — познакомить вас с некоторыми из основных физиологические концепции, которые вступают в действие при растяжении мышцы.Сначала будут представлены концепции с общим обзором, а затем (для желающих узнать кровавые подробности) будет обсуждаться в дальнейшие детали. Если вас не очень интересует этот аспект растяжка, вы можете пропустить эту главу. Другие разделы будут ссылаться на важные концепции из этой главы, и вы можете легко найти их основа «необходимости знать».
Вместе мышцы и кости составляют то, что называется опорно-двигательный аппарат тела. Кости обеспечивают осанку и структурная поддержка тела и мускулов обеспечивают телу способность двигаться (сокращаясь и тем самым создавая напряжение).В опорно-двигательный аппарат также обеспечивает защиту внутренних органов тела. органы. Чтобы кости выполняли свою функцию, они должны быть соединены вместе. чем-то. Точка, где кости соединяются друг с другом, называется шарнир , причем это соединение выполнено в основном связками (вместе с помощью мышц). Мышцы прикрепляются к кости с помощью сухожилий . Кости, сухожилия и связки не обладают способностью (как мышцы), чтобы ваше тело двигалось. Мышцы очень уникальны в этом уважать.
Мышцы различаются по форме и размеру и служат разным целям. Большинство крупных мышц, таких как подколенные сухожилия и квадрицепсы, контролируют движения. Другие мышцы, такие как сердце и мышцы внутреннего уха, выполняют другие функции. Однако на микроскопическом уровне все мышцы разделяют та же основная структура.
На самом высоком уровне (целая) мышца состоит из множества нитей ткань называется пучков . Это мышечные волокна, которые мы посмотрите, когда мы режем красное мясо или птицу.Каждый пучок состоит из пучков , которые представляют собой пучки мышечных волокон . Мышца волокна, в свою очередь, состоят из десятков тысяч нитевидных миофибриллы , которые могут сокращаться, расслабляться и удлиняться (удлиняться). Миофибриллы (в свою очередь) состоят из миллионов полосок, уложенных сквозной называется саркомеров . Каждый саркомер сделан из перекрывающие друг друга толстые и тонкие нити, называемые миофиламентами , . В толстые и тонкие миофиламенты состоят из сократительных белков , , в первую очередь актин и миозин.
Как сокращаются мышцы
То, как работают все эти различные уровни мышц, таково: следующим образом: Нервы соединяют позвоночник с мышцами. Место, где соединение нерва и мышцы называется нервно-мышечным соединением . Когда электрический сигнал проходит через нервно-мышечное соединение, он передается глубоко внутрь мышечных волокон. Внутри мышечных волокон сигнал стимулирует поток кальция, который вызывает густой и тонкие миофиламенты скользят друг по другу.Когда это происходит, это заставляет саркомер укорачиваться, что создает силу. Когда миллиарды саркомеры в мышцах сразу укорачиваются, что приводит к сокращению всего мышечного волокна.
Когда мышечное волокно сокращается, оно сокращается полностью. Такого нет вещь как частично сокращенное мышечное волокно. Мышечные волокна не в состоянии варьировать интенсивность их сжатия по отношению к нагрузке против которые они действуют. Если это так, то как сила мышечные сокращения различаются по силе от сильного до слабого? Что происходит состоит в том, что задействуется больше мышечных волокон, если они необходимы для выполнения работа под рукой.Чем больше мышечных волокон задействовано центральной нервной системы, тем сильнее сила, создаваемая мышечной сокращение.
Быстрые и медленные мышечные волокна
Энергия, которая производит поток кальция в мышечных волокнах, поступает от Митохондрия , часть мышечной клетки, которая преобразует глюкозу (уровень сахара в крови) в энергию. Различные типы мышечных волокон имеют разное количество митохондрий. Чем больше митохондрий в мышце волокно, тем больше энергии оно способно произвести.Мышечные волокна подразделяются на медленных волокон и быстрых волокон . Медленно сокращающиеся волокна (также называемые мышечными волокнами типа 1 ) медленно растут. сокращаются, но они также очень медленно утомляются. Быстро сокращающиеся волокна очень быстро сжимаются и бывают двух разновидностей: Тип 2A мышечные волокна , которые утомляются со средней скоростью, и Тип 2B мышечные волокна , которые очень быстро утомляются. Основная причина медленные волокна медленно утомляются из-за того, что они содержат больше митохондрии, чем быстро сокращающиеся волокна, и, следовательно, способны производить больше энергия.Медленно сокращающиеся волокна также меньше в диаметре, чем быстро сокращающиеся. волокна и имеют увеличенный капиллярный кровоток вокруг них. Потому что они имеют меньший диаметр и повышенный кровоток, медленные волокна способны доставлять больше кислорода и удалять больше отходов из мышечных волокон (что снижает их «утомляемость»).
Эти три типа мышечных волокон (типы 1, 2A и 2B) содержатся в все мышцы в разном количестве. Мышцы, которые нужно сильно сокращать времени (как и сердце) имеют большее количество Тип 1 (медленный) волокна.Когда мышца впервые начинает сокращаться, это в первую очередь 1-й тип. сначала активируются волокна, затем волокна Типа 2А и Типа 2В активируются (при необходимости) в указанном порядке. Дело в том, что мышечные волокна набранных в этой последовательности — вот что дает возможность выполнять мозговые команды с такими точно настроенными мышечными ответами. Это также делает волокна типа 2B трудно тренировать, потому что они не активируются пока не будет задействовано большинство волокон Типа 1 и Типа 2А.
HFLTA утверждает, что лучший способ запомнить разница между мышцами с преимущественно медленными волокнами и мышцы с преимущественно быстросокращающимися волокнами следует думать о «белых мясо »и« темное мясо ».Темное мясо темное, потому что в нем больше медленно сокращающихся мышечных волокон и, следовательно, большего количества митохондрий, которые темные. Белое мясо состоит в основном из мышечных волокон, которые большую часть времени отдыхают, но их часто просят кратко приступы интенсивной активности. Эта мышечная ткань может быстро сокращаться, но быстро утомляется и медленно восстанавливается. Белое мясо светлее чем темное мясо, потому что в нем меньше митохондрий.
Располагается вокруг мышцы, а ее волокна состоят из соединительных волокон. ткани .Соединительная ткань состоит из основного вещества и двух виды клетчатки на белковой основе. Два типа волокна: коллагеновая соединительная ткань и эластичная соединительная ткань . Коллагеновая соединительная ткань состоит в основном из коллагена (отсюда и ее название) и обеспечивает прочность на разрыв. Эластичная соединительная ткань состоит в основном из эластина и (как можно догадаться по названию) обеспечивает эластичность. Основное вещество называется мукополисахаридом и действует как смазка (позволяя волокнам легко скользить по одному другой), и в качестве клея (удерживая волокна ткани вместе в связки).Более эластичная соединительная ткань вокруг сустава, тем больше диапазон движений в этом суставе. Соединительные ткани состоящие из сухожилий, связок и фасциальных влагалищ, которые охватывают, или связать мышцы в отдельные группы. Эти фасциальные оболочки или фасция , названы в соответствии с их расположением в мышцы:
- эндомизий
- Самая внутренняя фасциальная оболочка, охватывающая отдельные мышечные волокна.
- перимий
- Фасциальная оболочка, которая связывает группы мышечных волокон в отдельные fasciculi (см. раздел «Состав мышц»).
- эпимизий
- Внешняя фасциальная оболочка, которая связывает целые пучки (см. Раздел «Состав мышц»).
Эти соединительные ткани помогают обеспечить эластичность и тонус мышцы.
Когда мышцы заставляют конечность перемещаться по диапазону движения сустава, они обычно действуют в следующих сотрудничающих группах:- агонистов
- Эти мышцы вызывают движение. Они создают нормальный диапазон движения в суставе сокращением.Агонисты также называют тягачей , поскольку именно они отвечает за создание движения.
- антагонистов
- Эти мышцы действуют противоположно движению, производимому агонисты и несут ответственность за возвращение конечности в исходное позиция.
- синергистов
- Эти мышцы выполняют или помогают в выполнении одного и того же набора суставов. движение как агонисты. Синергистов иногда называют нейтрализаторов , потому что они помогают нейтрализовать или нейтрализовать лишние движение от агонистов, чтобы убедиться, что генерируемая сила работает в желаемой плоскости движения.
- фиксаторы
- Эти мышцы обеспечивают необходимую поддержку, помогая удерживать остальная часть тела на месте во время движения. Фиксаторы также иногда называют стабилизаторы .
Например, когда вы сгибаете колено, подколенное сухожилие сокращается и, в некоторой степени то же самое происходит с икроножной мышцей и нижними ягодицами. Между тем, квадрицепсы заторможены (расслаблены и растянуты. несколько), чтобы не сопротивляться сгибанию (см. раздел «Взаимное торможение»).В этом примере подколенное сухожилие служит агонистом или первичный двигатель; квадрицепс служит антагонистом; и теленок и нижние ягодицы служат синергистами. Агонисты и антагонисты обычно располагается на противоположных сторонах пораженного сустава (например, ваш подколенные сухожилия и квадрицепсы или ваши трицепсы и бицепсы), в то время как синергисты обычно располагаются на той же стороне сустава рядом с агонистами. Более крупные мышцы часто призывают своих меньших соседей действовать как синергисты.
Ниже приводится список наиболее часто используемых мышц-агонистов / антагонистов. пары:
- грудные / широчайшие мышцы спины (грудные и широчайшие)
- передние дельтоиды / задние дельты (переднее и заднее плечо)
- трапеции / дельты (трапеции и дельты)
- брюшной пресс / выпрямители позвоночника (пресс и поясница)
- левый и правый внешние косые (стороны)
- квадрицепсы / подколенные сухожилия (квадрицепсы и бедра)
- голени / икры
- бицепс / трицепс
- сгибатели / разгибатели предплечья
Сокращение мышцы не обязательно означает, что мышца укорачивается; это только означает, что возникло напряжение.Мышцы могут заключить договор следующими способами:
- изометрическое сжатие
- Это сокращение, при котором не происходит никакого движения, потому что нагрузка на мышцу превышает напряжение, создаваемое сокращающейся мышцей. Это происходит, когда мышца пытается толкнуть или потянуть неподвижный объект.
- изотоническое сокращение
- Это сокращение, в котором происходит движение и , потому что
напряжение, создаваемое сокращающейся мышцей, превышает нагрузку на
мышца.Это происходит, когда вы используете свои мышцы для успешного толчка или
тянуть объект.
Изотонические сокращения делятся на два типа:
- концентрическое сжатие
- Это сокращение, при котором мышца уменьшается в длину (укорачивается). против встречного груза, например, подняв тяжесть.
- эксцентрическое сжатие
- Это сокращение, при котором мышца увеличивается в длину. (удлиняется) при сопротивлении нагрузке, например, при нажатии на что-либо.
Во время концентрического сокращения сокращающиеся мышцы служат агонистами и, следовательно, выполняют всю работу. Во время эксцентрическое сокращение, мышцы, которые удлиняются, служат агонисты (и делают всю работу). См. Раздел «Взаимодействие групп мышц».
Растяжение мышечного волокна начинается с саркомера. (см. раздел «Состав мышц»), основная единица сокращения в мышечное волокно. По мере сужения саркомера область перекрытия между толстые и тонкие миофиламенты увеличиваются.По мере того, как он растягивается, эта область перекрытия уменьшается, позволяя мышечным волокнам удлиняться. Однажды мышечное волокно находится на максимальной длине покоя (все саркомеры полностью растянуты), дополнительное растяжение накладывает силу на окружающие соединительная ткань (см. раздел «Соединительная ткань»). По мере увеличения напряжения волокна коллагена в соединительной ткани выравниваются вдоль та же силовая линия, что и напряжение. Следовательно, когда вы растягиваете, мышца волокно вытягивается саркомером на всю длину саркомера, а затем соединительная ткань принимает на себя оставшуюся слабину.Когда это происходит, это помогает перестроить любые неорганизованные волокна в направлении напряжение. Эта перестройка помогает восстановить поврежденную ткань. вернуться к здоровью.
Когда мышца растягивается, некоторые ее волокна удлиняются, а другие волокна могут оставаться в покое. Текущая длина всей мышцы зависит от количества растянутых волокон (аналогично тому, как общая сила сокращающейся мышцы зависит от количества набираемые волокна сокращаются). Согласно SynerStretch вы следует думать о «маленьких карманах волокон, распределенных по всей растяжение мышечного тела, а другие волокна просто поездка».Чем больше растянуты волокна, тем больше длина развита растянутой мышцей.
Проприорецепторы
Нервные окончания, передающие всю информацию о опорно-двигательном аппарате. Система центральной нервной системы называются проприорецепторами , . Проприорецепторы (также называемые механорецепторами ) являются источником всех проприоцепция : восприятие собственного положения тела и движение. Проприорецепторы обнаруживают любые изменения физического смещения (движение или положение) и любые изменения напряжения или силы в пределах тело.Они находятся во всех нервных окончаниях суставов, мышц и сухожилия. Проприорецепторы, связанные с растяжением, расположены в в сухожилиях и в мышечных волокнах.
Есть два типа мышечных волокон: интрафузальных мышечных волокон, и экстрафузальные мышечные волокна . Волокна Extrafusil — это те волокна, которые содержат миофибриллы (см. раздел «Состав мышц») и обычно имеется в виду, когда мы говорим о мышечных волокнах. Интрафузальные волокна также называется мышечными веретенами и лежат параллельно экстрафузальным волокнам.Мышечные веретена, или рецепторов растяжения, , являются первичными проприорецепторы в мышце. Еще один проприорецептор, который вступает в игру при растяжении располагается в сухожилии возле конца мышцы волокно и называется органом сухожилия Гольджи . Третий тип проприоцептор, называемый пачинским тельцем , расположен недалеко от орган сухожилия Гольджи и отвечает за обнаружение изменений в движение и давление внутри тела.
Когда экстрафузальные волокна мышцы удлиняются, растут и интрафузальные волокна. волокна (мышечные веретена).Мышечное веретено содержит два разных типы волокон (или рецепторов растяжения), которые чувствительны к изменению длины мышцы и скорости изменения длины мышцы. Когда мышцы сокращается, это создает напряжение в сухожилиях там, где орган сухожилия Гольджи расположен. Орган сухожилия Гольджи чувствителен к изменению напряжения и скорость изменения напряжения.
Рефлекс растяжения
Когда мышца растягивается, растягивается и мышечное веретено. (см. раздел Проприорецепторы).Мышечное веретено фиксирует изменение длины (и как быстро) и посылает сигналы к позвоночнику, которые передают это Информация. Это вызывает рефлекс растяжения (также называемый миотатический рефлекс ), который пытается противостоять изменению мышцы длины, заставляя растянутую мышцу сокращаться. Чем внезапнее изменение длины мышцы, тем сильнее будут сокращения мышц (плиометрическая, или «прыжковая» тренировка основана на этом факте). Этот базовый функция мышечного веретена помогает поддерживать мышечный тонус и защитить тело от травм.
Одна из причин проведения растяжки в течение длительного периода времени заключается в том, что когда вы удерживаете мышцу в растянутом положении, мышца веретено приживается (привыкает к новой длине) и сокращает его сигнализация. Постепенно вы можете тренировать рецепторы растяжения, чтобы большее удлинение мышц.
Некоторые источники предполагают, что при длительных тренировках растяжка рефлекс определенных мышц можно контролировать, так что или нет рефлекторного сокращения в ответ на внезапное растяжение.В то время как этот тип контроля дает возможность для максимального увеличивает гибкость, а также обеспечивает наибольший риск травм при неправильном использовании. Только непревзойденные профессиональные спортсмены и считается, что танцоры, достигшие вершины своего вида спорта (или искусства), на самом деле обладают таким уровнем мышечного контроля.
Компоненты рефлекса растяжения
Рефлекс растяжения имеет как динамический, так и статический компоненты. Статический компонент рефлекса растяжения сохраняется до тех пор, пока мышца растягивается.Динамический компонент рефлекса растяжения (который может быть очень мощным) длится всего мгновение и отвечает к первоначальному внезапному увеличению длины мышцы. Причина, по которой Рефлекс растяжения состоит из двух компонентов, потому что на самом деле их два виды интрафузионных мышечных волокон: волокон ядерной цепи , которые являются отвечает за статическую составляющую; и ядерный мешок волокна , которые отвечают за динамическую составляющую.
Волокна ядерной цепи длинные и тонкие, и они постепенно удлиняются при растянулся.Когда эти волокна растягиваются, нервы рефлекса растяжения увеличивать их скорострельность (сигнализацию) по мере того, как их длина постоянно увеличивается. Это статический компонент рефлекса растяжения.
Волокна ядерного мешка выступают посередине, где они больше всего эластичный. Нервные окончания этих волокон, чувствительные к растяжению, обернуты вокруг этой средней области, которая быстро удлиняется, когда волокно растянулся. Внешне-средние области, напротив, действуют так, как будто они наполнен вязкой жидкостью; они сопротивляются быстрому растяжению, затем постепенно расширяются при длительном напряжении.Итак, когда требуется быстрое растяжение у этих волокон поначалу больше всего растягивается середина; тогда, как внешне-средние части расширяются, средняя может несколько укорачиваться. Так что нерв, который ощущает растяжение этих волокон, быстро срабатывает вместе с начало быстрого растяжения, затем замедляется по мере того, как средний участок волокна разрешено снова сокращаться. Это динамический компонент рефлекс растяжения: сильный сигнал к сокращению в начале быстрого увеличение длины мышц, за которым следует немного «выше нормы» сигнализация, которая постепенно уменьшается по мере того, как скорость изменения мышцы длина уменьшается.
Реакция удлинения
Когда мышцы сокращаются (возможно, из-за рефлекса растяжения), они производят напряжение в точке, где мышца соединяется с сухожилием, где расположен орган сухожилия Гольджи. Орган сухожилия Гольджи регистрирует изменение напряжения, и скорость изменения напряжения, и отправляет сигналы к позвоночнику, чтобы передать эту информацию (см. раздел Проприорецепторы). Когда это напряжение превышает определенный порог, оно запускает реакция удлинения , которая препятствует сокращению мышц и заставляет их расслабиться.Другие названия этого рефлекса — обратный миотатический рефлекс , аутогенное торможение и складной нож рефлекс . Эта основная функция сухожилия Гольджи орган помогает защитить мышцы, сухожилия и связки от травм. Реакция удлинения возможна только потому, что сигнал Гольджи орган сухожилия спинного мозга достаточно мощный, чтобы преодолеть сигнализация мышечных веретен, приказывающих мышце сокращаться.
Еще одна причина для растяжки в течение длительного периода времени: чтобы позволить этой реакции удлинения произойти, тем самым помогая растянутому мышцы, чтобы расслабиться.Мышцу легче растянуть или удлинить, когда она не пытается заключить контракт.
Взаимное ингибирование
Когда агонист сокращается, чтобы вызвать желаемое движение, он обычно заставляет антагонистов расслабиться (см. раздел «Взаимодействие групп мышц»). Это явление называется реципрокным торможением , потому что антагонистам запрещено сокращаться. Иногда это называют реципрокная иннервация , но этот термин на самом деле неверный, поскольку он агонисты, которые подавляют (расслабляют) антагонисты.Антагонисты действительно ли , а не , действительно иннервируют (вызывают сокращение) агонистов.
Такое подавление мышц-антагонистов необязательно. На самом деле может произойти совместное сокращение. Когда вы выполняете приседания, можно обычно предполагают, что мышцы живота препятствуют сокращению мышцы поясничного или нижнего отдела спины. В этом конкретном Однако, например, мышцы спины (выпрямители позвоночника) также сокращаются. Этот это одна из причин, почему приседания хороши как для укрепления спины, так и для желудок.
При растяжении расслабленную мышцу растянуть легче, чем чтобы растянуть сокращающуюся мышцу. Воспользовавшись ситуации, когда происходит реципрокное торможение , можно получить более эффективная растяжка за счет расслабления антагонистов во время растягиваются из-за сокращения агонистов. Вы также хотите расслабиться любые мышцы, которые используются как синергисты мышцы, которую вы пытаетесь растянуть. Например, когда вы растягиваете икры, вы хотите сжать голень. мышцы (антагонисты голени), сгибая стопу.Однако подколенные сухожилия используют икроножные мышцы как синергист, поэтому вы также хотите расслабить подколенные сухожилия, сокращая четырехглавую мышцу (т. е. удерживая ногу прямой).
Перейти к предыдущей, следующей главе.
В чем разница между медленными и быстро сокращающимися мышечными волокнами?
Мышечные волокна тела просты, но сложны в том, как мы их понимаем и тренируем. То, как мы тренируемся, будь то силовая или аэробная, будет влиять на состав наших мышечных волокон. Это происходит из-за того, что мышечные волокна приспосабливаются к нагрузке, которую мы испытываем во время тренировок.
Стресс, вызванный различными режимами тренировки, будет влиять на состав миофибрилл в мышцах (миофибриллы состоят из актиновых и миозиновых нитей) , что помогает волокнам получить титулы Типа I, IIA и IIX. Мышцы нашего тела состоят из различного количества волокон каждого типа.
- Тип I (медленные сокращения) — Активная деятельность, основанная на продолжительности и выносливости.
- Тип IIA (быстрое сокращение) — Относительная сила и взрывные движения.
- Тип IIX (быстрое подергивание (эр) ) — Взрывные движения.
Между волокнами существует множество физиологических различий, благодаря которым они получили названия быстрых и медленных сокращений. В конце каждого сокращения выходное усилие у разных типов очень похоже, но скорость сокращения различается, отсюда их названия быстрых и медленных сокращений.
Некоторые из этих различий касаются того, как типы мышц используют энергию, порядок, в котором они задействованы, и насколько быстро они производят силу.
Определения мышечных волокон
Существует три метода описания или определения того, к какому типу волокна относится мышечное волокно (также называемые методами, которые исследователи используют при классификации мышцы как медленно или быстро сокращающейся) . К ним относятся: гистиологическое окрашивание миозина, идентификация изоформ MHC и идентификация метаболических ферментов.
Тип I (медленные сокращения) : Это мышечное волокно эффективно использует кислород для производства АТФ, который является формой энергии в наших мышцах.Они сокращаются медленно и более устойчивы к усталости из-за их способности использовать кислород с большей скоростью.
Тип IIA (быстрое сокращение) : Это мышечное волокно вырабатывает энергию анаэробно (иначе говоря, без использования свободного кислорода), и сокращается намного быстрее, чем волокна типа I. Из-за отсутствия аэробного энергетического обмена они быстро утомляются. Волокно IIA часто считается промежуточным волокном.
Тип IIX (быстрый (эр) подергивания) : Подобно мышечному волокну типа IIA, это волокно также создает энергию анаэробно и будет быстро сокращаться с той же (но несколько более быстрой) скоростью утомления.Это мышечное волокно сокращается быстрее всего из трех, что делает его наиболее подверженным преждевременному утомлению.
Мышечные волокна и энергетические системы
Энергетические системы организма будут играть роль в том, как каждое мышечное волокно сокращается и используется. В организме есть три типа энергетических систем, в том числе: АТФ-ПК, гликолитическая и окислительная. Каждая энергетическая система будет играть определенную роль в различных видах деятельности, и их приблизительные оценки времени, основанные на максимальных потребностях в энергии, приведены ниже.
- Энергетическая система ATP-PC : (+/-) 12 секунд
- Гликолитическая система : 30 секунд — 2 минуты
- Окислительная система : 2+ минуты
Если у вас есть представление об энергетических системах, вы можете увидеть, как они влияют на способность мышечных волокон сокращаться.Ознакомьтесь с таблицей ниже, которая дает краткое представление о различных энергетических системах и мышечных волокнах, на которые они оказывают наибольшее влияние.
| Тип I — медленное сокращение | Выносливость и высокая репутация |
| Тип IIA — Fast Twitch | Тяжелые бега и относительная силовая тренировка |
| Тип IIX — быстрый (er) Twitch | Спринты и взрывные движения |
Имейте в виду, что приведенная выше таблица не является полностью идеальным примером того, какие волокна будут присутствовать в определенных видах деятельности.В каждом действии, которое вы выполняете, будут сочетаться почти все двигательные нейроны и типы мышечных волокон, но их количество будет отличаться.
Приведенная ниже таблица представляет собой примерный пример того, какие спортсмены высокого уровня будут иметь самый высокий состав каждого мышечного волокна. Эта таблица, очевидно, не предназначена для использования в качестве сценария «на каждый случай», а для того, чтобы дать наглядное представление о том, как каждый элитный спортсмен будет чаще всего тренироваться, чтобы влиять на определенные волокна. Для силовых атлетов всегда будет кроссовер.
| Тип I — медленное сокращение | Триатлонисты и марафонцы |
| Тип IIA — Fast Twitch | Спортсмены и бодибилдеры функционального фитнеса |
| Тип IIX — быстрый (er) Twitch | Тяжелоатлеты и пауэрлифтеры |
Эти таблицы представляют собой иллюстрации, помогающие представить контекст волокон, которые вы будете чаще всего видеть при определенных занятиях и у спортсменов, что может быть дополнительно объяснено с помощью принципа размера.
Принцип размера
Этот принцип представляет собой научную концепцию, согласно которой наша нервная система задействует двигательные нейроны с мышечными волокнами в предсказанном порядке. Порядок обычно включает в себя задействование более мелких нейронов и более медленных мышц, а затем — последних.
Например, представьте себе подтягивание на 10 повторений. Если это движение дается вам легко, то вы можете обнаружить, что на ваши первые 4-5 повторений сильно влияют меньшие двигательные нейроны / медленно сокращающиеся мышцы, а затем крупные нейроны / быстро сокращающиеся волокна включаются примерно на 6+ повторений.Напротив, если это движение затруднено, тогда ваши более крупные нейроны / быстро сокращающиеся волокна сработают гораздо раньше.
Во многих исследованиях этот принцип верен, но есть одна проблема. Исследование, проведенное в 2014 году, показывает, что гораздо сложнее оценить модели набора персонала с учетом различных изменений реальных жизненных сил. Например, спортсмен, выполняющий несколько движений в одной последовательности, может испытывать колебания в различных моделях набора. Эта концепция затрудняет определение принципа размера с четким определением того, как наше тело задействует различные типы мышечных волокон.
Можно ли увеличить количество мышечных волокон?
Короче да и нет. В какой-то степени мы можем.
Принято считать, что мы рождаемся с одинаковым количеством медленных и быстрых волокон 50/50. Во время тренировки мы влияем на эффективность каждого целевого типа мышц. Это приведет к увеличению размера и скорости, с которой мы можем создавать силу с помощью этой мышцы (подумайте, прогрессивная перегрузка в нашей тренировке) . Мы знаем, что можем увеличить размер волокна, но можем ли мы изменить что-то вроде волокна типа I на волокно типа II?
И снова наука противоречит этому ответу.Многие исследования показывают, что мы можем влиять примерно на 10% нашего мышечного типа, но мы не можем полностью преобразовать одни волокна в другие. Например, мы не можем превратить свое тело в худощавую среднюю на 100% быстросокращающуюся машину Типа IIX (… если только) .
Прежде чем терять надежду, исследование 2012 г. показало, что у нас есть способность влиять на промежуточные волокна. Например, мы можем заменить некоторые волокна типа IIA на тип IIX (так мы со временем становимся более мощными) .Тело способно адаптироваться и изменять промежуточное быстро сокращающееся волокно (Тип IIA), чтобы производить еще более быстрое сокращение, что сделало его Типом IIX. Кроме того, исследователи отмечают, что при более длительных тренировках организм может преобразовывать волокна типа IIA в волокна типа I.
Промежуточные волокна демонстрируют наибольшую способность к изменению от тренировочного (предполагается, что это связано с их более высокой кислородной емкостью). Зависание включает в себя переключение между истинными волокнами типа I и типа IIX.Эта область исследований все еще отсутствует.
Мы состоят из нескольких типов мышечных волокон, и когда мы рождаемся, они распределены довольно равномерно. По мере того, как мы продвигаемся по карьерной лестнице, мы влияем на то, как наше тело адаптируется и реагирует на стресс, тем, как быстро мы можем создавать силу. У одних спортсменов естественно больше определенных волокон, чем у других (или способность лучше адаптироваться) .
В конце концов, очень важно тренироваться таким образом, чтобы вы могли оптимально развиваться и преуспевать в своем виде спорта.
Примечание редактора: Мария Дальзот, диетолог, бегун по горам / тропам и читатель BarBend, после прочтения этой статьи сказала следующее:
Как спортсмен, работающий на выносливость, кажется, что единственный раз, когда мои быстро сокращающиеся мышцы активируются, это когда я тянусь за едой после 4-часовой пробежки в горах. Говоря о еде, энергетические системы организма (АТФ-ПК, гликолитическая и окислительная) не только будут играть роль в том, как каждое мышечное волокно сокращается и используется, но также будут определять, какое топливо используется для окисления.Углеводы являются эффективным источником топлива во время анаэробных упражнений, когда организм не может обрабатывать достаточно кислорода для удовлетворения своих потребностей. Чем больше кислорода становится доступным во время менее интенсивных упражнений, жир становится основным источником топлива, экономя запасы гликогена в мышцах. Но так же, как в каждом действии, которое вы выполняете, будет задействовано сочетание
моторных нейронов и типов мышечных волокон, ваше тело будет использовать комбинацию углеводов и жиров в качестве топлива. Нет переключателя, который полностью включает или выключает окисление углеводов или жиров.Ничто в теле не происходит изолированно.
В чем разница между быстро и медленно сокращающимися мышечными волокнами?
Четверг, 24 августа 2017 г. Автор Джеральдин Кэмпбелл
Что за «зазор» с быстрым и медленным подергиванием?
Фразы «быстро сокращающийся» и «медленно сокращающийся» часто возникают в фитнес-индустрии (и во всей зоне нашего Участника) в отношении наших мышечных волокон.Но что это и чем они отличаются?
В организме человека есть три основных типа мышечных волокон: Тип I, Тип IIa и Тип IIb. Их можно различить по уровню активности фермента миозин-аденозинтрифосфаза (АТФаза), а также по другим характеристикам. Волокна типа I медленно сокращаются, в то время как оба волокна типа II классифицируются как быстро сокращающиеся. Процент каждого из них в вашем теле определяется как вашей генетикой (40% для медленных и 60% для быстро сокращающихся), так и факторами окружающей среды, такими как опыт тренировок.
Медленно сокращающиеся волокна
Эти мышечные волокна — лучший друг спортсмена на выносливость. Их много в митохондриях, миоглобине (связывающем кислород белке) и капиллярах, которые позволяют использовать кислород для производства энергии посредством окислительного фосфорилирования. Капилляры транспортируют кислород, связанный с миоглобином, в мышцы, где он может использоваться митохондриями для синтеза АТФ; позволяя вам тренироваться дольше. Следовательно, медленно сокращающиеся мышечные волокна обладают высокой устойчивостью к утомлению, поэтому такие мышцы, как икроножная мышца (икроножная мышца), содержат больше медленно сокращающихся волокон; позволяя вам продолжать ходить, стоять и выполнять все повседневные дела, не утомляясь быстро.Основным топливом для медленно сокращающихся мышечных волокон являются триглицериды (жиры), поскольку этот источник энергии может обеспечить большое количество АТФ. Узнайте больше о том, как ваше тело использует жир в качестве топлива здесь.
В то время как медленно сокращающиеся мышечные волокна помогают вам во время длительных пробежек или заездов, они не подходят для взрывных подъемов или спринтов, поскольку они медленнее сокращаются.
Быстро сокращающиеся волокна
Два типа быстро сокращающихся мышечных волокон отличаются от медленных, потому что они обладают способностью производить энергию в отсутствие кислорода (гликолитическое окисление).Это позволяет им быстрее вырабатывать энергию, используя фосфокреатин и гликоген (подробнее об этих энергетических системах читайте здесь), чтобы подпитывать эти быстрые взрывные движения, такие как прыжки и спринт. Это означает, что у них более высокая скорость сокращения, чем у медленных волокон. Эти два типа быстро сокращающихся волокон также отличаются друг от друга, при этом волокна типа IIa имеют некоторое сходство с медленными волокнами.
Волокнатипа IIa могут вырабатывать энергию как гликолитическим, так и окислительным путями, что делает их немного более устойчивыми к усталости, чем волокна типа IIb.Тип IIa также имеет больше капилляров, чем IIb; помогая их способности производить энергию с помощью кислорода.
Таблица ниже суммирует различия между различными мышечными волокнами:
Итак, для каких видов спорта лучше всего подходят?
Волокна типа I идеально подходят для продолжительных упражнений, выполняемых с низкой или средней интенсивностью. Если вам нравятся марафоны, полумарафоны, дистанции 10 км, длинные прогулки или велосипеды; это мышечные волокна, через которые вы проходите.
Волокна типа IIa предназначены для упражнений средней продолжительности с более высокой интенсивностью.Такие упражнения, как бег на 400 м и использование умеренно тяжелых весов в диапазоне 8-12 повторений во время тренировки с отягощениями, будут преимущественно использовать эти волокна.
Волокнатипа IIb отлично подходят для коротких тренировок с высокой интенсивностью, требующих взрывной выработки силы мышцами. Спринт на 100 м или пауэрлифтинг являются примерами видов спорта, в которых эти волокна процветают.
Что FitnessGenes может рассказать мне о типах моих мышечных волокон?
Существует множество вариантов генов, которые могут влиять на ваш состав этих различных типов волокон.ACTN3 является одним из таких примеров, белок, кодируемый этим геном, наиболее часто встречается в быстро сокращающихся мышечных волокнах. Было показано, что наличие генотипа, который приводит к нокауту этого белка, больше связано с большей выносливостью, чем с мощностью или скоростью.
Но ваша генетика лишь частично контролирует состав ваших мышечных волокон. Если вы хотите тренироваться, чтобы стать спринтером, но имеете генетическое преимущество в отношении выносливости, тренировки, которые вы проводите, чтобы стать спринтером, помогут вашим мышечным волокнам адаптироваться к стимулам более высокой интенсивности и улучшат ваши результаты!
Если вам понравился этот блог, прочтите другие мои сообщения:
Использование жира в качестве топлива
Тяга к еде
Ваш обеденный перерыв предназначен не только для еды.Обновите его до ранчо!
Тренировка с эспандером
Познай свое сердце
Как алкоголь может ограничивать ваш прогресс
Бег и генетика
Внимательность
Генетическое преобладание бегунов из Восточной Африки и Ямайки
Спринт и мощность
Скандинавская диета
Морские водоросли
Окислительный стресс
Ссылки
Эдгертон В.Р., Смит Дж. Л. и Симпсон Д.Р., 1975. Типы мышечных волокон мышц ног человека. Гистохимический журнал , 7 (3), стр. 259-266.
Thorstensson, A. и Karlsson, J., 1976. Утомляемость и состав волокон скелетных мышц человека. Acta Physiologica, 98 (3), стр. 318-322.
Бушар, К., Симоно, Дж. А., Лорти, Г., Буле, М. Р., Маркотт, М. и Тибо, М. К., 1986. Генетические эффекты в распределении типов волокон скелетных мышц человека и активности ферментов. Канадский журнал физиологии и фармакологии , 64 (9), стр.1245-1251.
Эссен, Б., Янссон, Э., Хенрикссон, Дж., Тейлор, А.В. и Saltin, B., 1975. Метаболические характеристики типов волокон в скелетных мышцах человека. Acta Physiologica , 95 (2), стр 153-165.
.