Метаболизм: что это, как ускорить, комментарии врача :: Здоровье :: РБК Стиль
Разбираемся вместе с экспертами, как без особых усилий расходовать больше калорий.
Материал прокомментировали:
- Наталья Антонова, врач-эндокринолог, диетолог «СМ-Клиника»;
- Мария Волченкова, нутрициолог компании Best Doctor, клинический психолог, член Российского союза нутрициологов, диетологов и специалистов пищевой индустрии, эксперт по работе с ДНК-тестами
Что такое метаболизм
Метаболизм — это процесс, основным показателем которого является скорость обмена веществ [1]. Он поддерживает работу всех внутренних органов. Понятие включает в себя все жизненно важные химические функции тела: дыхание, восстановление клеток и переваривание пищи. Эти процессы требуют энергии, минимальное количество которой называется базовой скоростью обмена веществ, или уровнем метаболизма (BMR).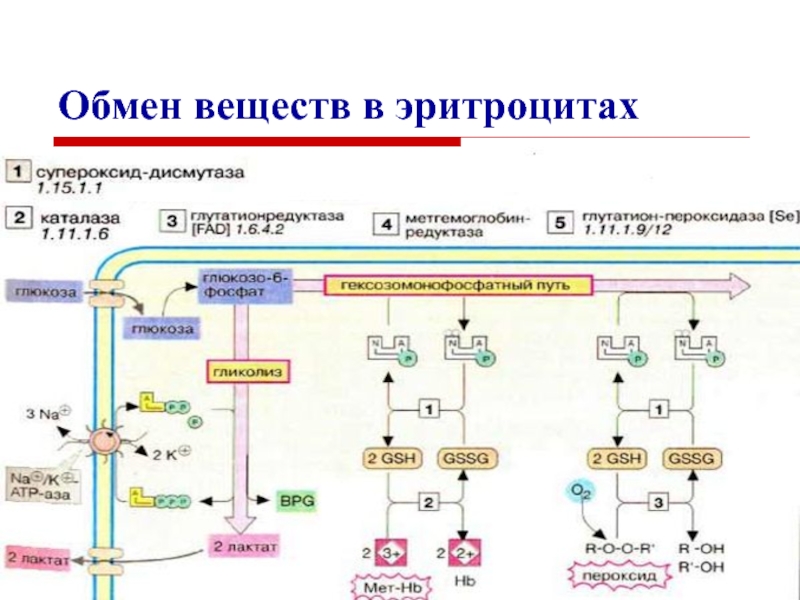
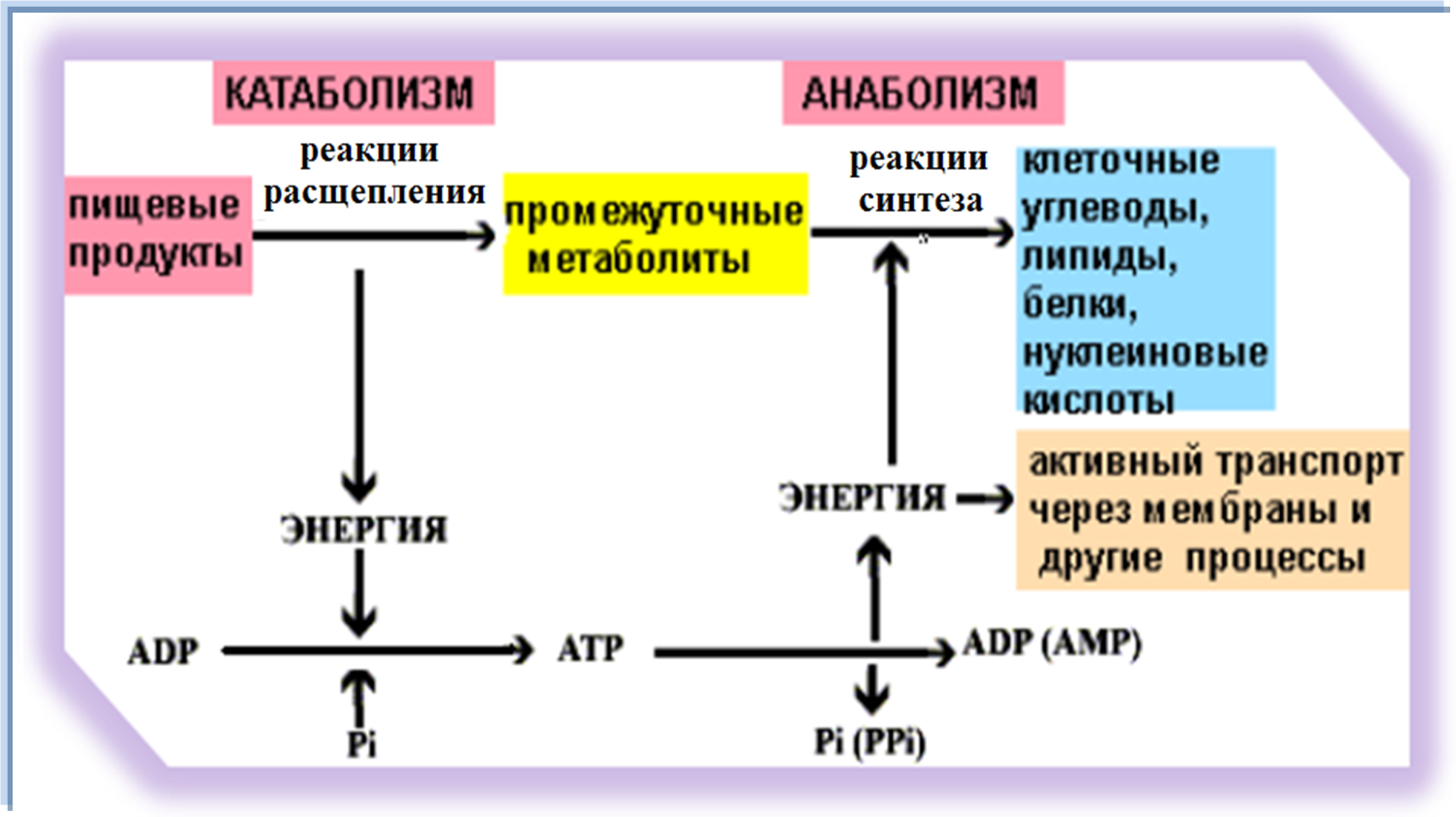
- Анаболизм — накопление запасов энергии в организме.
- Катаболизм — их расщепление.
Существует множество онлайн-калькуляторов, которые помогут рассчитать ежедневную потребность в энергии. Например, обратите внимание на уравнение Харриса — Бенедикта:
Базовый (базальный) метаболизм у женщин:
- BMR = 655,0955 + (9,5634 * вес в кг) + (1,8496 * рост в см) – (4,6756 * возраст в годах).
Базальный метаболизм у мужчин:
- BMR = 66,4730 + (13,7516 * вес в кг) + (5,0033 * рост в см) – (6,7550 * возраст в годах).
Теперь полученное значение нужно умножить на коэффициент, зависящий от физической активности:
- сидячий образ жизни — 1,2;
- умеренная активность (легкие физические нагрузки либо занятия 1–3 раз в неделю) — 1,375;
- средняя активность (занятия 3–5 раз в неделю) — 1,55;
- активные люди (интенсивные нагрузки, занятия 6–7 раз в неделю) — 1,725;
- спортсмены и люди, выполняющие сходные нагрузки (6–7 раз в неделю), — 1,9.

Например, если вы женщина 30 лет, ростом 170 см и весом 65 кг, которая большую часть времени проводит сидя за компьютером, расчет будет таким:
- BMR = 655,0955 + (9,5634 * 65) + (1,8496 * 170) – (4,6756 * 30) = 1450,9
- 1450,9 * 1,2 = 1741 ккал / сутки.
Итоговая цифра показывает дневную калорийность, которой следует придерживаться.
Мария Волченкова, нутрициолог:
«К этой формуле также можно добавить термический эффект пищи (ТЭФ), обычно он составляет 10%, которые плюсуют к обмену веществ. BMR + ТЭФ (10% от основного обмена) * фактор физической активности».
Почему подсчет калорий — опасная практика и как похудеть без диет
На что влияет метаболизм
Часто люди, которым никак не удается нормализовать вес, обвиняют в этом замедленный метаболизм. На самом деле есть мало доказательств того, что его можно считать основным барьером на пути к стройной фигуре.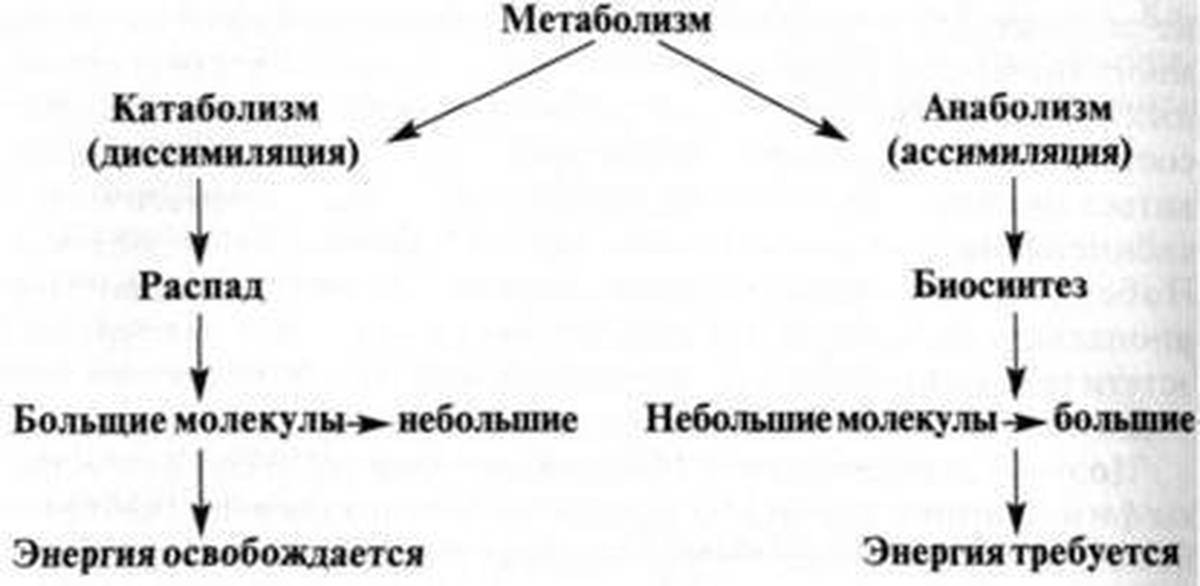
Только в редких случаях происходит прибавка в весе из-за медицинских проблем, замедляющих метаболизм, таких как синдром Кушинга или недостаточная активность щитовидной железы — гипотиреоз. При этом скорость обмена веществ напрямую связана с нормализацией веса. Чем больше калорий потрачено, тем меньше жира откладывается в теле. В условиях их дефицита происходит процесс похудения, ускоряется жировой (липидный) обмен. Метаболизм также учитывается при наборе массы. В каждом отдельном случае важно разобрать индивидуальные особенности организма и составить подходящий план питания и тренировок.
Что такое нарушение метаболизма
Норма в каждом конкретном случае будет различаться. Врач может диагностировать нарушения белкового, жирового, углеводного, минерального и водного обмена веществ [4]. Чаще всего причинами отклонений от оптимального метаболизма считают:
Врач может диагностировать нарушения белкового, жирового, углеводного, минерального и водного обмена веществ [4]. Чаще всего причинами отклонений от оптимального метаболизма считают:
Нарушение обмена веществ выражается в следующих признаках:
- ухудшение состояния кожи, волос и ногтей;
- повышенная потливость;
- проблемы с избыточным весом;
- сильные отеки;
- нарушение сна;
- болезни зубов;
- сниженная работоспособность.
При постановке диагноза врач учитывает результаты анализов и осмотра. Распространенные признаки нарушения обмена веществ: высокий уровень триглицеридов и сахара в крови, пониженный уровень «хорошего» холестерина и высокое давление [5].
Ускоренный метаболизм
Люди с ускоренным обменом веществ сжигают калории быстрее, чем те, у кого метаболизм замедлен.
Некоторые из наиболее распространенных признаков быстрого метаболизма:
- проблемы с набором веса;
- анемия;
- гиперактивность;
- высокая частота сердечных сокращений;
- повышенная температура тела даже в покое;
- низкий процент жира в организме.
Эктоморфы — люди с высоким уровнем метаболизма, от природы стройные, с трудом набирают вес и наращивают мышечную массу. На скорость обмена веществ влияют различные факторы, но в основном это генетика, физическая активность, пол и возраст. Большое значение имеют мышцы тела: по сравнению с жировыми клетками, им требуется больше энергии для поддержания жизнедеятельности.
Лучший способ определить, почему у вас ускоренный метаболизм и нужно ли предпринимать какие-то действия по его нормализации, — консультация с врачом. Специалист поможет определить, насколько это нормально в вашем случае, не является ли побочным эффектом заболевания и стоит ли что-то менять.
Замедленный метаболизм
На замедление обмена веществ влияют многие факторы:
- хронический стресс;
- недостаток микроэлементов, особенно железа;
- несбалансированное, нерегулярное питание;
- некачественные продукты;
- отсутствие режима дня, поздние подъемы и отходы ко сну.
Метаболизм замедляется при недостаточном количестве калорий в рационе, поэтому с ним часто сталкиваются те, кто придерживается строгих ограничительных диет. Это чревато снижением работоспособности, уровня энергии и производства гормонов щитовидной железы. При замедлении обмена веществ развивается атрофия мышц и рост жировых отложений. Диагностировать это состояние должен врач. Например, для эндоморфов сниженный метаболизм и склонность к набору веса — разновидность нормы.
Это чревато снижением работоспособности, уровня энергии и производства гормонов щитовидной железы. При замедлении обмена веществ развивается атрофия мышц и рост жировых отложений. Диагностировать это состояние должен врач. Например, для эндоморфов сниженный метаболизм и склонность к набору веса — разновидность нормы.
Что такое монодиеты и почему они опасны
Что может ускорить метаболизм
Некоторые исследования подтверждают, что скорость обмена веществ не всегда снижается с возрастом, а в основном зависит от образа жизни [6]. Замедление химических реакций приводит к недостатку питания тканей организма, появляются различные дисфункции. Проконсультируйтесь с врачом, чтобы подобрать оптимальную терапию и систему питания, если необходимо нормализовать обмен веществ. Есть несколько простых и доступных способов, которые можно использовать дополнительно.
1. Лежите на диване
Обычно лежание на диване ассоциируют только с неподвижностью и лишним весом, но в то же время это моменты комфорта и расслабления.
2. Откажитесь от вечернего бокала вина
Все тот же стресс может привести к привычке расслабляться вечером за счет бокала вина. Но известно, что даже одна порция алкоголя может отрицательно влиять на метаболизм, вызывая такие болезни обмена веществ, как, например, диабет [8]. Если вам нужен ритуал для вечернего отдыха, попробуйте строить его не вокруг алкоголя, а вокруг теплых полезных напитков — травяных и ягодных чаев.
3. Выбирайте здоровые десерты
Простые углеводы — сладкое и продукты из белой муки — быстро поднимают уровень сахара в крови, запускают процесс отложения жира и делают организм нечувствительным к инсулину [9]. В то же время десерты — это важная часть социального ритуала, который часто совсем не хочется пропускать. Компромисс — десерты из фруктов, творожные угощения с орехами без добавления сахара, печенье из цельнозерновой муки с бананом и изюмом. Да, во фруктах тоже содержится сахар, но вместе с клетчаткой он усваивается и попадает в кровь медленнее.
В то же время десерты — это важная часть социального ритуала, который часто совсем не хочется пропускать. Компромисс — десерты из фруктов, творожные угощения с орехами без добавления сахара, печенье из цельнозерновой муки с бананом и изюмом. Да, во фруктах тоже содержится сахар, но вместе с клетчаткой он усваивается и попадает в кровь медленнее.
4. Пританцовывайте
Даже небольшая активность в течение дня — пританцовывание в очереди, отбивание ритма ступнями во время работы за столом, прогулка вокруг остановки в ожидании автобуса — сжигает калории и заставляет избавляться от лишних запасов. Час такой активности в день без изменения других факторов может позволить сбросить до одного килограмма в месяц.
5. Пейте кофе
Кофеин стимулирует нервную систему, а вместе с ней и обмен веществ, который может ускориться на 5–8% (100–150 килокалорий в день). Кроме того, чашка кофе перед тренировкой поможет взбодриться и сделать упражнения эффективнее. Важно обсудить этот вопрос с вашим гастроэнтерологом — при некоторых проблемах ЖКТ употребление этого напитка нежелательно.
Важно обсудить этот вопрос с вашим гастроэнтерологом — при некоторых проблемах ЖКТ употребление этого напитка нежелательно.
6. Компенсируйте недостатки сидячего образа жизни
Сидячий образ жизни вызывает целый спектр проблем со здоровьем и замедляет обмен веществ [10]. Чтобы лучше себя чувствовать и повысить дневную активность, введите в привычку каждый час делать 10-минутную разминку. Кроме того, можно установить стол, который позволяет работать стоя.
7. Ешьте больше овощей
Клетчатка в овощах требует от организма дополнительной энергии (калорий) для переваривания и усваивания. Кроме того, овощи в большинстве случаев содержат низкое количество калорий и много витаминов, что также полезно для нормализации веса.
8. Не переживайте о перекусах
Скорость обмена веществ снижается при длительных, а не при коротких промежутках времени без еды.
Мария Волченкова:
«Интервальное голодание увеличивает выработку нескольких гормонов, участвующих в жиросжигании и влияющих на набор веса. Во-первых, это инсулин. Слишком высокий инсулин дает телу сигнал запасать жир и ассоциирован с повышенным риском онкологических и сердечно-сосудистых заболеваний, диабета 2-го типа и ожирения. Гиперинсулинемия — ранний индикатор метаболических дисфункций [11]. Короткие циклы голода могут снижать индекс инсулинорезистентности [12]. В одном интересном исследовании Университета Алабамы небольшой группы мужчин с ожирением и преддиабетом было установлено, что после пяти недель интервального голодания с приемами пищи в восьмичасовой промежуток времени наблюдалось значительное снижение инсулина и улучшение чувствительности к нему [13]. Кроме того, у участников снизилось артериальное давление и аппетит. А вот что действительно замедляет обмен веществ, так это диеты, основанные на экстремальном ограничении калорий вкупе с неадекватными физическими нагрузками.
Достаточно обратить внимания на участников знаменитого американского шоу The biggest loser, в котором участники всеми способами старались сбросить вес. Врачи установили, что через шесть лет после шоу большинство героев вернули себе весь потерянный с таким трудом вес, а вот уровень их обмена веществ при этом не вырос и оставался в среднем на 500 калорий ниже, чем должен быть при их массе тела [14]».
9. Занимайтесь спортом хотя бы пять минут в день
Интенсивная тренировка с кардионагрузкой сжигает калории в первую очередь за счет высокого потребления кислорода. К тому же она повышает активность обмена веществ на 24 часа, даже если речь идет о пяти минутах в день. Выделите для себя это время — и уже через пару недель почувствуете себя более подтянутыми.
10. Старайтесь больше гулять
Прогулка — та нагрузка, которую несложно выполнять каждый день большинству из нас. Она помогает размяться, снять стресс, разогнать обмен веществ и стимулировать организм сжигать больше калорий. Полчаса в день — серьезная инвестиция в ваше здоровье.
Полчаса в день — серьезная инвестиция в ваше здоровье.
Комментарии экспертов
Наталья Антонова, врач-эндокринолог, диетолог «СМ-Клиника»
Чаще всего признаками замедленного метаболизма являются набор веса, сухость кожных покровов, ломкость, слоистость ногтей, редкий пульс, ухудшение памяти, зябкость, сонливость, снижение либидо. При ускоренном метаболизме, как правило, наблюдаются тахикардия, потливость, снижение веса, повышенная возбудимость, раздражительность, бессонница. В любом случае при появлении этих признаков требуется консультация специалиста.
К врачу необходимо обратиться в том случае, если на фоне обычного образа жизни пациент начинает замечать быстро прогрессирующие изменения в своем теле: потерю или набор веса, сопровождающиеся изменением пульса, артериального давления, появлением и прогрессированием отеков, выраженной сухостью, гиперпигментацией кожи, перепадами настроения, отсутствием жизненных сил и энергии. Предпочтительнее начинать с консультации терапевта. Он оценит состояние пациента, проведет основные необходимые обследования и диагностирует изменения со стороны органов ЖКТ (жировой гепатоз, нарушение желчеоттока, дисбиоз кишечника), ухудшение состояния кожных покровов (сухость кожи, высыпания, образование и рост папиллом), половых органов (снижение половой функции у мужчин, нарушение менструального цикла у женщин) или гормонального фона (набор веса, стрии), после чего своевременно направит пациента для обследования к профильным специалистам.
Предпочтительнее начинать с консультации терапевта. Он оценит состояние пациента, проведет основные необходимые обследования и диагностирует изменения со стороны органов ЖКТ (жировой гепатоз, нарушение желчеоттока, дисбиоз кишечника), ухудшение состояния кожных покровов (сухость кожи, высыпания, образование и рост папиллом), половых органов (снижение половой функции у мужчин, нарушение менструального цикла у женщин) или гормонального фона (набор веса, стрии), после чего своевременно направит пациента для обследования к профильным специалистам.
Если человек сидит на диетах, занимается спортом и не худеет, это происходит, как правило, при резком и значительном снижении суточного калоража. Когда организм переходит в режим «выживания», замедляется обмен веществ и начинает работать только на запас. Возможно, здесь имеют место незакрытые дефициты веществ и несоблюдение питьевого режима. Могут быть иные причины, разбираться с которыми поможет врач.
Модификация образа жизни — 80% успеха при желании изменить свое самочувствие и скорректировать вес.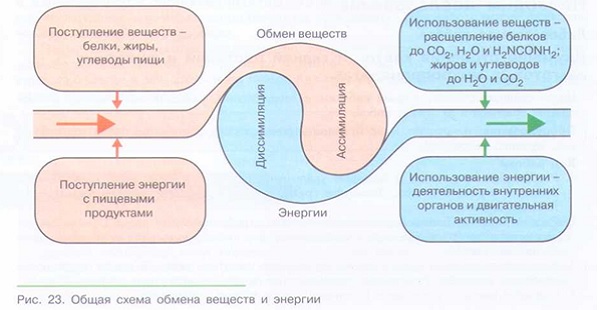 Поэтому очень важно четко следовать основным рекомендациям по соблюдению питьевого режима, дробного питания, закрытию дефицитов витаминов и микроэлементов, физической активности, соблюдению режима сна и бодрствования, исключению из рациона фастфуда, трансжиров. Все эти несложные действия дают гораздо более эффективный и долгосрочный результат, нежели простой подсчет калорий и создание их дефицита.
Поэтому очень важно четко следовать основным рекомендациям по соблюдению питьевого режима, дробного питания, закрытию дефицитов витаминов и микроэлементов, физической активности, соблюдению режима сна и бодрствования, исключению из рациона фастфуда, трансжиров. Все эти несложные действия дают гораздо более эффективный и долгосрочный результат, нежели простой подсчет калорий и создание их дефицита.
Мария Волченкова, нутрициолог компании Best Doctor, клинический психолог, член Российского союза нутрициологов, диетологов и специалистов пищевой индустрии, эксперт по работе с ДНК-тестами
Продукты, ускоряющие обмен веществ, действительно существуют. Их еще называют «мета-бустеры». Во-первых, положительно влияют на скорость обменных процессов высокобелковые продукты. Все потому, что тело должно потратить довольно много энергии на их усвоение. Термический эффект белковых продуктов доходит до 30%. Достаточное количество белка помогает сохранить здоровую мышечную ткань (при снижении веса в том числе). Адекватный уровень мышечной ткани — важный фактор «быстрого метаболизма». Но надо помнить, что при переходе даже на самый адекватный высокобелковый тип питания следует предварительно пройти чек-ап. В частности, при нарушении функции почек обилие высокобелковых продуктов может иметь негативные последствия.
Достаточное количество белка помогает сохранить здоровую мышечную ткань (при снижении веса в том числе). Адекватный уровень мышечной ткани — важный фактор «быстрого метаболизма». Но надо помнить, что при переходе даже на самый адекватный высокобелковый тип питания следует предварительно пройти чек-ап. В частности, при нарушении функции почек обилие высокобелковых продуктов может иметь негативные последствия.
«Разгонять метаболизм» могут продукты, богатые селеном и железом. При дефиците селена или железа снижается функция щитовидной железы. А это, в свою очередь, замедляет метаболизм. Здесь тоже нужно действовать осторожно, потому что при избыточном накоплении и селен, и железо становятся крайне опасны. Например, чтобы закрыть суточную потребность в селене, достаточно съедать 2–3 бразильских ореха в день, но не больше.
Есть и другие метабустеры: чечевица, некоторые виды масла, кофе, корица, имбирь, яблочный уксус, перец чили. Помните, что ни один из этих продуктов не поможет ускорить метаболизм сам по себе: нужны сбалансированное здоровое питание, питьевой режим, качественный сон, управление уровнем стресса и равномерная физическая активность.
Как питаться при ускоренном обмене веществ? А при замедленном?
При замедленном обмене веществ нужно в первую очередь разобраться с причинами такого состояния. Если мы говорим об условно здоровом человеке с эндоморфным соматотипом (соответственно, сниженным коэффициентам энерготорат), часто эффективной оказывается концепция LCHF — рацион с пониженным содержанием углеводов, умеренным — белка и повышенным уровнем полезных жиров (оливковое масло и авокадо, орехи, мелкая жирная морская рыба и так далее). Такой подход подойдет не всем, но в любом случае при эндоморфном соматотипе хорошо работает ограничение углеводов. То есть адекватный низкоуглеводный подход (НУ). Адекватный НУ-подход — это ни в коем случае не диеты вроде Дюкана или Кремлевской и не переход только на белковые продукты. В этих случаях скорее проявятся проблемы с почками, чем ускорится метаболизм. НУ-подход обязательно включает в себя овощи и зелень, большинство злаковых, орехи, полезные жиры, ягоды и фрукты. Вопрос только в количестве.
Вопрос только в количестве.
Есть еще один факт, важный для эндоморфов. У представителей этого соматотипа самые сложные отношения с алкоголем. Если вы не хотите замедлить и без того низкую скорость обменных процессов, постарайтесь полностью отказаться от спиртных напитков, в первую очередь от пива.
Что касается повышенного метаболизма (эктоморфы), главное правило для них— не забывать есть. Приемы пищи должны быть сбалансированы и включать источник белка, «медленных» углеводов, полезных жиров и клетчатки. Есть в данном случае можно и чаще, чем 3 раза в день, добавляя второй завтрак и полдник.
Если метаболизм замедлен, это можно исправить с помощью только образа жизни — активности и системы питания? Или может понадобиться медикаментозная терапия?
Если человек здоров, если у него нет скрытых пищевых непереносимостей, которые, согласно ряду гипотез, могут способствовать набору веса, а значит, замедлению метаболизма, то да. Медикаментозная поддержка нужна там, где замедленный метаболизм обусловлен именно определенными проблемами со здоровьем. Но здесь может быть недостаточно только нормализации питания и адекватного уровня физической активности. Важнейшую роль будут играть качественный сон и управление уровнем стресса. Мы часто недооцениваем эти факторы и совершаем тем самым очень большую ошибку. А ведь нервная система — самый главный «орган», который дирижирует всеми остальными системами, включая гормональную. Поэтому важно контролировать напряжение, отдыхать, расслабляться и заботиться о качестве сна. Кстати, если не можете выбрать — пойти на тренировку или выспаться, однозначно выбирайте второе. Повышайте бытовую активность: припаркуйтесь за квартал до офиса и идите на работу пешком.
Медикаментозная поддержка нужна там, где замедленный метаболизм обусловлен именно определенными проблемами со здоровьем. Но здесь может быть недостаточно только нормализации питания и адекватного уровня физической активности. Важнейшую роль будут играть качественный сон и управление уровнем стресса. Мы часто недооцениваем эти факторы и совершаем тем самым очень большую ошибку. А ведь нервная система — самый главный «орган», который дирижирует всеми остальными системами, включая гормональную. Поэтому важно контролировать напряжение, отдыхать, расслабляться и заботиться о качестве сна. Кстати, если не можете выбрать — пойти на тренировку или выспаться, однозначно выбирайте второе. Повышайте бытовую активность: припаркуйтесь за квартал до офиса и идите на работу пешком.
А если «генетика виновата»? Есть шансы разогнать метаболизм или придется смириться?
При любой генетике человек может набрать существенное количество лишнего веса, заработать сахарный диабет второго типа и инсулинорезистентность. И наоборот: при любой генетике можно иметь здоровый вес без избытка жировой ткани. Даже при низком коэффициенте энерготрат (пониженном метаболизме) можно поддерживать здоровый вес за счет нормализации питания. А вот методы нужно подбирать индивидуально в каждом конкретном случае, потому что мы все отличаемся друг от друга.
И наоборот: при любой генетике можно иметь здоровый вес без избытка жировой ткани. Даже при низком коэффициенте энерготрат (пониженном метаболизме) можно поддерживать здоровый вес за счет нормализации питания. А вот методы нужно подбирать индивидуально в каждом конкретном случае, потому что мы все отличаемся друг от друга.
Читайте также:
Метаболические коррекция — лечение и диагностика нарушений метаболизма в Клинике Expert Clinics
Классическая медицина, как правило, лечит симптомы, в то время как антивозрастная сфокусирована на выявлении и устранении глубинных причин заболеваний.
При интегративном антивозрастном подходе к здоровью организм человека рассматривается как единая система, способная к саморегуляции, особенно если устранить корень нарушений.
Метаболическая коррекция — это тонкая настройка клеточной биохимии, которая способна выровнять состояние здоровья в целом и затормозить развитие многих недугов.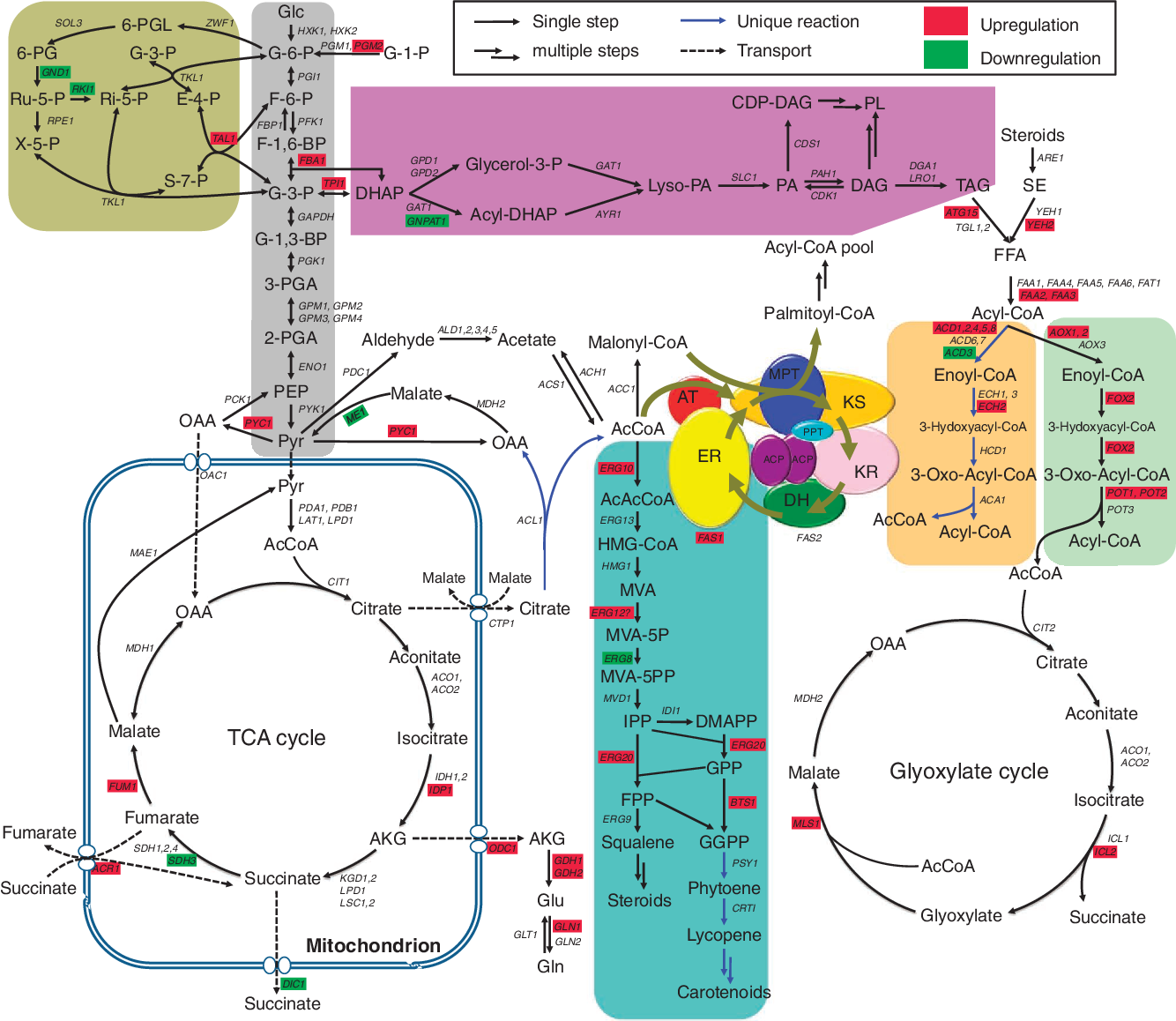
Благодаря персонализированному подходу врачам антивозрастной медицины удается восстановить функции организма на клеточном уровне, во всех тканях и органах.
Перезвоните мне
Основы метаболической коррекции
Научные исследования последних лет доказывают: многие хронические заболевания возникают из-за несбалансированного питания, некачественного сна, токсинов, стресса и определенных дефицитов, мешающих полноценному функционированию организма и истощающих его.
К примеру, нехватка различных нутриентов негативно влияет на метаболические функции организма, которые контролируют все физиологические процессы. В итоге клетки перестают нормально функционировать и повреждаются.
В результате процессы детоксикации становятся неэффективными и замедленными. А это, в свою очередь, приводит к накоплению токсинов в организме, снижению активности иммунной системы, дисбалансу микробиоты, изменению гормонального баланса и снижению функции митохондрий (“энергетических станций” наших клеток).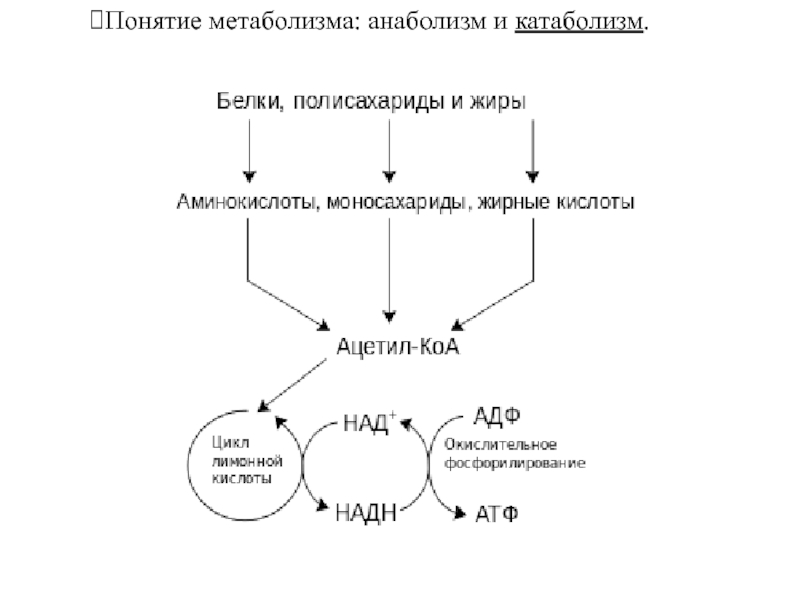
Ученые также доказали, что с возрастом организм человека производит большое количество токсинов — так называемые эндотоксины. При нарушении метаболических путей они перестают выводится.
Подобные изменения ведут к хроническим дегенеративным состояниям, которые сильно подрывают здоровье человека и снижают качество его жизни. “Плохая” наследственность только усиливает эти разрушительные эффекты.
Метаболическая коррекция заключается в использовании методик и персонализированных препаратов в оптимальных формах и дозах. Они регулируют функцию метаболических ферментов, а также устраняют дефициты полезных веществ.
Цели метаболической коррекции
Любая болезнь — результат нарушения нормальной физиологии, которое происходит, когда организм подвергается воздействию внешних или внутренних неблагоприятных факторов.
Суть метаболической коррекции — в поддержании здоровья и улучшении работы всех систем организма. Так, например, нормализуются белковый, углеводный и жировой обмены.
Коррекция метаболических нарушений может быть весьма эффективной для улучшения состояния людей с сахарным диабетом, тяжелыми формами аллергии и аутоиммунными заболеваниями.
Врачи, восстанавливающие метаболические процессы, добиваются сразу нескольких целей:
Предупреждение хронических заболеваний;
Коррекция уже имеющихся болезней;
Активное поддержание здоровья для компенсации генетической предрасположенности к различным недугам.
Повышение качества жизни пациентов.
Работа с пациентами на метаболическом уровне предполагает определенный рацион и применение различных лечебных методик (инфузионные, пероральные, диетотерапия). При грамотном применении они могут стать “первой линией обороны” нашего здоровья.
Записаться на консультацию
Метаболическая коррекция в Expert Clinics
В штате клиники Expert Clinics в Москве — опытные врачи антивозрастной медицины, прошедшие многочисленные обучения как в России, так и за рубежом. Они успешно помогают пациентам выравнивать метаболические процессы организма даже в запущенных случаях.
Они успешно помогают пациентам выравнивать метаболические процессы организма даже в запущенных случаях.
Лечение и диагностика дисфункций метаболизма должна быть комплексной. Для начала необходима полная клиническая оценка, выявляющая проблемы конкретного пациента. Это потребует подробного анамнеза, функциональной диагностики и всестороннего лабораторного тестирования.
Необходимо обследование может выглядеть так:
Генетическое тестирование;
Комплексный анализ крови;
Выявление маркеров воспаления;
Иммунные тесты;
Содержание тяжелых металлов;
Анализ мочи ;
УЗИ внутренних органов;
МРТ, КТ.
После пациенту будет назначена персонализированная схема лечения.
Как показывает обширная практика врачей Expert Clinics, благодаря метаболической коррекции многие пациенты обретают отличное самочувствие, становятся энергичнее и наблюдают ремиссию своих давних болезней.
Записаться
Механизмы реакций метаболизма этоксозепама в гомогенатах печени крыс
Bogatskij, A.V., Andronati, S. A., & Golovenko, N. Ya. (1980). Trankvilizatory (1,4-benzdiazepiny i rodstvennye struktury) [Tranquilizers (1.4-benzdiazepines and relative structures)] Kyiv: Naukova Dumka. [in Ukrainian].
Holovenko, M. Ya., Larionov, V. B., & Pavlovskyi, V. I. (2009). Farmakolohichni vlastyvosti preparatu «Levana IS» [Pharmacological properties of drug “Levana IC”]. Visnyk psykhiatrii ta psykhofarmakoterapii, 2(16), 70–73. [in Ukrainian].
Kravchenko, I. A., Radaeva, I. N., & Zhukova, N. A. (2011). Farmakologicheskaya aktivnost’ novykh proizvodnykh 1,4-benzdiazepina [Pharmacological activity of new 1.4-benzdiazepine derivatives.] Ukrainskyi naukovo-medychnyi molodizhnyi zhurnal, 4, 60–62. [in Ukrainian].
Zhukova, N. O., Holovenko, M. Ya., & Larionov, V. B. (2012). Farmakokinetychnyi profil ta metabolizm 14S-etoksozepamu v orhanizmi myshei pislia vnutrishnovennoho vvedennia [Pharmacological profile and metabolism of 14C-ethoxazepam in mice after intravenous administration] Klinichna farmatsiia, 16(4), 32–38. [in Ukrainian].
Farmakokinetychnyi profil ta metabolizm 14S-etoksozepamu v orhanizmi myshei pislia vnutrishnovennoho vvedennia [Pharmacological profile and metabolism of 14C-ethoxazepam in mice after intravenous administration] Klinichna farmatsiia, 16(4), 32–38. [in Ukrainian].
Pavlovskyi, V. I., Semenishina, K. O., Larionov, V. B., & Zhukova, N. O. (2012). Syntez 14С-etoksozepamu ta vyznachennia yoho osnovnykh fizyko-khimichnykh ta radiolohichnykh pokaznykiv [14C-Ethoxazepam synthesis and determination of its main physics-chemical and radiological parameters]. Farmatsevtychnyi zhurnal, 2, 43–49. [in Ukrainian].
Golovenko, N. Ya., Zin’kovskij, V. G. (1978). Opredelenie trankvilizatorov 1,4-benzdiazepinonovogo ryada i ikh metabolitov v biologicheskikh sredakh [Determination of 1.4-benzdiazepine derivatives and their metabolites in biologic medium] Khimiko-farmacevticheskij zhurnal, 12(1), 3–14. [in Russian].
Todeschini, R., & Consonni, V. (2009) Molecular Descriptors for Chemoinformatics. (Vol. 41 (2 Volume Set)). John Wiley & Sons.
(2009) Molecular Descriptors for Chemoinformatics. (Vol. 41 (2 Volume Set)). John Wiley & Sons.
Bogatskij, A. V., Golovenko, N. Ya., Andronati, S. A., Kolomojchenko, G. Yu., & Zhilina, Z.I. (1978). Uchastie redoks-cepi mikrosom pecheni krys v suzhenii kol’ca 1,4-benzdiazepinov [Participating of rat liver microsome redox-chain in reduction of the ring of 1.4-benzdiazepines]. Doklady Akademii nauk SSSR, 1(234), 215–218. [in Russian].
Выявлены особенности метаболизма у разных народов
Обмен веществ у разных народов и наций протекает по-разному, что, в свою очередь, может пролить свет на причины высокого уровня сердечно-сосудистых заболеваний в некоторых странах – такой вывод был сделан учёными из Имперского колледжа Лондона (Imperial College, London) на основании анализа результатов исследования метаболических маркеров.
В ходе работы было проанализировано 4680 замороженных образцов мочи, взятых у взрослых добровольцев из Великобритании, США, Китая и Японии в рамках международного эпидемиологического проекта INTERMAP, проводившегося ещё в 1997-1999 годах.
Группе учёных из Имперского колледжа удалось провести наиболее полный анализ состава взятых образцов при помощи спектроскопии ядерного магнитного резонанса – отчёт об этом исследовании опубликован в журнале Nature. В ходе анализа удалось выделить метаболические фенотипы тех стран, чьи добровольцы приняли участие в проекте, то есть определить характерные для этих стран особенности обмена веществ.
Относительный уровень продуктов метаболизма играет роль своеобразного маркера, который может свидетельствовать о том, каков риск возникновения тех или иных заболеваний и как это связано с режимом питания, микрофлорой кишечника, экологией и образом жизни человека в целом.
По словам одного из авторов исследования профессора Джереми Николсона (Jeremy Nicholson), анализ позволил выделить несколько устойчивых групп продуктов метаболизма для каждой из четырёх стран. В частности, исследование показало, что у добровольцев из США и Великобритании примерно одинаковый уровень сердечно-сосудистых заболеваний, и при этом их метаболические показатели соответствуют друг другу. По мнению учёных, это свидетельствует о том, что схожий образ жизни человека оказывает одинаковое влияние на его обмен веществ, несмотря на то что США и Великобритания находятся по разные стороны Атлантики.
По мнению учёных, это свидетельствует о том, что схожий образ жизни человека оказывает одинаковое влияние на его обмен веществ, несмотря на то что США и Великобритания находятся по разные стороны Атлантики.
Напротив, у более близких географически Китая и Японии, чьи добровольцы, по всей вероятности, имеют в целом больше родственных генетических связей, чем, к примеру, пёстрое по своему этническому составу население США, характер продуктов метаболизма отличается – свидетельство различий в образе жизни и экологии этих стран. При этом продукты обмена веществ этнических японцев, проживающих в Америке, соответствуют метаболическим отпечаткам американцев, но отличаются от отпечатков «японских» японцев.
По словам Джереми Николсона, данная работа позволит выявить факторы, влияющие на уровень сердечно-сосудистых заболеваний, и даст возможность управлять риском их возникновения. С ним согласен другой участник исследования, профессор Пол Эллиотт (Paul Elliott), который подчёркивает, что человек не может изменить свою ДНК, но способен повлиять на факторы, определяющие характер обмена веществ.
Ссылка на публикацию: MEMBRANA
Лечение заболеваний ЖКТ и нарушений обмена веществ
Нарушение метаболизма – следствие многих причин.
Прогулки по живописным местам крымского побережья и морской чистый воздух эффективно дополняют комплексное лечение желудочно-кишечного тракта в санатории Сакрополь в Крыму.
Для пациентов работает коллектив опытных высококвалифицированных докторов, которые создали на базе современного медицинского оборудования все необходимые условия для отдыха и выздоровления. Вода минеральных источников и озерные сакские грязи прославлены своей чудодейственной силой из глубины веков. Аппликации озерной грязи широко применялись при болезнях кишечника, желудка, пищевода.
Грязелечение ЖКТ в Крыму – традиции и новаторство
Известные столетиями способы применения целебной грязи продолжают давать высокие положительные результаты. Наряду с этим, наши специалисты усовершенствовали методы грязелечения, адаптировали их к современной энтерологии и медицинскому оборудованию. Для каждого пациента врач разрабатывает индивидуальную программу проведения курса терапии.
Лечение ЖКТ сакскими грязями – важный оздоровительный фактор!
Анализ многолетнего опыта применения грязевых аппликаций позволяет констатировать рост положительной динамики в лечении желудочно-кишечных заболеваний. В комплексы процедур входит также прием минеральной воды, кислородных коктейлей, солевых ванн, физиотерапия, ЛФК, диета, массаж.
В санатории желудочно-кишечного профиля Сакрополь сегодня успешно используется новая методика одноразовых аппликаций. Наши схемы проведения грязевых процедур – уникальны в своем роде и назначаются отдельно для каждого пациента.
Показания к лечению ЖКТ в нашем санатории г. Саки:
- Хронический гастрит
- Колит, энтероколит
- Холецистит
- Нарушения функций печени
- Панкреатит
- Дискинезия желчного пузыря и выводящих путей
- Язва желудка и 12-перстной кишки
- Период реабилитации после операции на желудке
- Сахарный диабет 2 степени
Результаты лечения ЖКТ сакскими грязями в санатории «Сакрополь»:
- ускорение микроциркуляции клеточного обмена
- насыщение тканей кислородом, снабжение питательными веществами
- выведение шлаков
- нормализация липидного обмена (снижение риска инсульта, инфаркта)
- уменьшение уровня сахара в крови
- улучшение работы сердечно-сосудистой системы
- устранение болевого синдрома
- восстановление обменных процессов, снижение веса
- повышение качества здоровья и жизни
Нарушение метаболизма – проблема века
Проблема нарушений обмена веществ на сегодняшний день становится все более острой для большинства современных людей. Повышенному риску подвергаются в основном жители мегаполисов с легкодоступным фаст-фудом и ведущие напряженный график работы. Процессы метаболизма выполняют незаменимые функции для работы всего организма.
Основные причины нарушения обмена веществ в организме:
- Неправильное питание (голодание, переедание, перенасыщение углеводной пищей)
- Злоупотребление курением, алкоголем
- Стрессы, длительные депрессии
- Гиподинамия или чрезмерные физические нагрузки
Противопоказания:
- Хронические заболевания органов пищеварения в стадии обострения.
- Тяжелые формы хронического эзофагита, рефлюкс-эзофагит с выраженными кардиальными проявлениями, рефлекторной стенокардией, сочетание рефлюкс-эзофагита с грыжей пищеводного отверстия диафрагмы, подлежащей хирургическому лечению.
- Рубцовые сужения пищевода, привратника, двенадцатиперстной кишки с нарушением их проходимости.
- Стриктуры желчных путей.
- Язвенная болезнь желудка и двенадцатиперстной кишки, осложненная стенозом привратника, повторными кровотечения, пенетрацией язвы в течение последних 8-10 месяцев, подозрение на малигнизацию язвы.
- Болезнь Золлингера-Эллисона.
- Осложнения после операции на желудке (незаживающий послеоперационный рубец, свищи, синдром приводящей петли, послеоперационный панкреатит, демпинг-синдром тяжелой степени, атония культи желудка, обострение послеоперационного гастрита, холецистита, гепатита, тяжелая форма постваготомической диареи.
- Гиперпластические изменения слизистой оболочки желудка ( при невозможности исключить злокачественные перерождения).
- Полипоз желудка (2 полипа и более), одиночные кровоточащие полипы желудка, аденоматозные полипы на широком основании с размерами 1 см и более с тенденцией к росту в течение последнего года.
- Желчнокаменная болезнь при наличии частых и длительных тяжелых приступов, протекающих с нарушением оттока желчи или признаками активной инфекции (повышение температуры тела и др.), эмпиема желчного пузыря.
- Гепатит А.
- Хронический активный гепатит.
- Все формы желтухи.
- Цирроз печени.
- Остаточные явления после перенесенного вирусного гепатита с признаками активности процесса, патологическими отклонениями функциональных проб печени, в том числе уровня АСТ и АЛТ, постгепатитной гипербилируинемии (выше 21 ммоль/л), выраженной гепатомегалией (нижний край печени выступает на 3 см и более из-под реберной дуги по правой срединно-ключичной линии, воспалением желчного пузыря и желчных путей.
- Энтероколит с выраженным нарушением всасывания (истощение).
- Хроническая дизентерия.
- Неспецифический язвенный колит.
- Хронический язвенный колит с обширным язвенным или эрозивным процессами в прямой и сигмовидной кишке.
- Кровоточащий геморрой.
- Полипоз кишечника ( 2 полипа и более), одиночные кровоточащие или аденоматозные полипы на широком основании размером 1 см и более с тенденцией к росту.
- Тяжелые формы хронических панкреатитов в течение 2 месяцев после обострения, а также осложненного воспалительными кистами или свищами, значительном снижении
внешнесекреторной функции железы с выраженным нарушением пищеварения и общего состояния.
Происхождение нарушений метаболизма не ограничивается этими причинами. Ученые продолжают изучать этимологию обменных процессов и внедряют новые способы борьбы с опасными заболеваниями: сахарный диабет, ожирение, анорексия, подагра и т.п.
У нас вы получите квалифицированную помощь эндокринологов, гастроэнтерологов, физиотерапевтов на высоком профессиональном уровне. Пациентам назначают сеансы грязелечения по специально составленной программе с учетом особенностей заболевания. В комплексе процедур для эффективного выздоровления рекомендован массаж, лечебная гимнастика, диетическое питание, правильный режим.
Модель пациента
Возрастная категория: взрослые
Класс болезней XI: болезни органов пищеварения
Группа заболеваний: болезни пищевода, желудка и 12п кишки, неинфекционный колит, другие болезни кишечника, брюшины, печени, желчного пузыря, желчевыводящих путей и поджелудочной железы, другие болезни органов пищеварения.
Код по МКБ-10: К 20, К21.0, К 21.9, К22.1, К25, К26, К28, К29.3, К29.4, К29.5, К29.8, К29.9, К58.9, К66.0, К73.0, К7.1, К73.8, К81.1, К83.0, К86.1, К91.1, К91.5
Фаза: хроническая
Стадия: ремиссии
Осложнение: без осложнений
Условия оказания: санаторно-курортные и амбулаторно-курортные
Лечение из расчета 21 день
ПРОГРАММА ЛЕЧЕНИЯ (СКАЧАТЬ)РЕКОМЕНДУЕМЫЕ ПЛАТНЫЕ ПРОЦЕДУРЫ
При противопоказаниях к общему грязелечению лечащим врачом назначается или одна процедура общего воздействия:
Где полечить заболевания ЖКТ в Крыму?
В течение многих поколений миллионы людей живут, употребляя различную пищу – вареную, сырую, комбинируя её в различных пропорциях. Если мы едим нечто такое, что не годится для нас, несовместимо с потребностями и балансом нашего организма, мы страдаем. Боли, спазмы – это ни что иное, как предупреждение об опасном недуге. Однако благодаря поразительной терпимости нашего организма, наказание может наступить не сразу. Но через много дней, месяцев или даже лет все равно наступит. Это возмездие природы за нарушение её законов. Появляются заболевания желудочно-кишечного тракта – гастрит, язвенная болезнь, колит, ожирение, подагра, холецистит и другие.
Лечение этих болезней начинаются от советов друзей, назначений врачами амбулаторного или стационарного лечения и вершиной оздоровления является санаторно-курортное лечение.
Одним из санаториев Крыма по лечению желудочно-кишечных заболеваний является Санаторий Сакрополь в городе Саки. В комплексе лечения используется минеральная вода скважины № 4323 (3-7м) в г. Саки под названием «Крымская хрустальная» — гидрокарбонатнохлоридная натриевая. В санатории лечебная база европейского уровня, квалифицированный персонал.
С целью увеличения продолжительности ремиссии больных рекомендуется направлять на санаторно-курортное лечение в самое опасное для рецидива болезни время – весной и осенью. Здесь же в санатории можно провести курсовое лечение ожирения, подагры, снизиться вес без ущерба деятельности желудочно-кишечного тракта.
Лечебный климат, забота врачей и уникальные процедуры позволят восстановить утраченное здоровье и качество жизни.
ᐉ Санатории с лечением НАРУШЕНИЯ ОБМЕНА ВЕЩЕСТВ 🔥️ Цены на 2021 год • Путевка.ком
Одним из основных процессов, который происходит непрерывно для поддержания нормальной жизнедеятельности организма, является обмен веществ, то есть метаболизм. Выделяют две эго стадии:
катаболизм – распад сложных химических веществ (аминокислот, липидов, углеводов) на простые производные;
анаболизм – репликация органических соединений из элементарных частиц.
На этапе распада выделяется энергия, а при синтезе она, наоборот, затрачивается, причем в разном количестве. Во всех этих химических реакциях принимают участие ферменты, на молекулярном уровне выступая в роли катализаторов и снижая энергозатратность процессов.
Интенсивность метаболизма зависит от множества факторов, в том числе от температуры окружающей среды (зимой он замедляется), количества накопленной энергии. У каждого человека, в зависимости от пола, возраста, образа жизни скорость его индивидуальна. Но симптомы сбоев в обменных процессах у всех схожи – повышается утомляемость, снижается скорость реакций на внешние раздражители, ухудшается память, скачет давление, наблюдаются резкие перепады настроения, развивается сердечная аритмия, гормональные и другие проблемы.
При лечении заболеваний нарушения обмена веществ применяются следующие процедуры: Парафинолечение, Душ Шарко, Лечебная физкультура (ЛФК), Магнитотерапия , Ванны радоновые, Амплипульстерапия, Ванна гальваническая, Ванна радоновая сухая, Ванны йодобромные, Ванны с морской водой, Ванны хвойные , Ванны углекислые и сероводородные , Ванны хлоридно-натриевые (или солевые) , Гидромассаж, Гальванизация и электрофорез, Гальваногрязелечение, Гирудотерапия , Дарсонвализация, Душ гидролазерный, Душ циркулярный , Индуктотермия , Лазеротерапия , Массаж антицеллюлитный, Массаж лечебно-профилактический , Микроволновая резонансная терапия (МРТ, КВЧ), Озокеритотерапия , Озонотерапия, Питье минеральных вод, Прессотерапия (аппаратный лимфодренажный массаж, компрессионный массаж, пневмомассаж) , Талассотерапия, Терренкуры, Ультразвуковая терапия и ультрафонорез лекарственных средств, Электросон (нейросон, электроанальгезия).
Лазерная терапия
Лазерная терапия
Лазерные методы лечения занимают одно из ведущих мест в списке самых перспективных направлений современной медицины. При наружном применении лечение лазером происходит путемвоздействия излучающего терминала на определенные зоны и точки тела. Свет проникает сквозь ткани на большую глубину и стимулирует обмен веществ в пораженных тканях, активизирует заживление и регенерацию, происходит общая стимуляция организма в целом. При внутривенной лазеротерапии, через тонкий световой проводник, который вводится в вену, лазерный луч воздействует на кровь. Внутрисосудистое действие низкоинтенсивнымизлучением позволяет воздействовать на всю массу крови. Это приводит к стимуляции кроветворения, усилению иммунитета, повышению транспортной функции крови, а так же способствует усилению метаболизма. Такое лечение совершенно безболезненно и комфортно.Существует четкая схема сочетания лазеротерапии с медикаментозной терапией, в соответствии с которой, достигается наилучший лечебный эффект.
Лазерная терапия эффективна и показана в следующих случаях:
— При заболеваниях сердечно — сосудистой системы (ИБС,стенокардия, вегетососудистая дистония, артериальная гипертензия).
— При заболеваниях периферических сосудов (облитерирующие поражения сосудов конечностей, флебит, тромбофлебит,варикозное расширение вен).
— При заболеваниях органов дыхания (пневмония, плеврит, бронхит, бронхиальная астма, туберкулез легких).
— При заболеваниях органов желудочно – кишечного тракта(гастрит, язвенная болезнь желудка и двенадцатиперстной кишки, панкреатит, холецистит, колит).
— При заболеваниях органов мочеполовой системы: почек (острые и хронические пиелонефриты, мочекаменная болезнь), цистит, простатит, уретрит, ослабление половой функции.
— При гинекологических заболеваниях (послеродовой эндометрит,неспецифические сальпингиты, бартолинит).
— При заболеваниях нервной системы (невралгии, невриты, травмынервных стволов и сплетений, последствия черепно – мозговойтравмы, последствия инсульта, последствия нейроинфекции).
— При ЛОР заболеваниях (отит, гайморит, фронтит, ринит,ангина, тонзиллит, ларингит, трахеит).
— При заболеваниях опорно-двигательного аппарата (артриты, артрозы, остеохондроз, миозит, ушибы мягких тканей,периоститы).
— При вялогранулирующих ранах, перитонитах, фурункулах, карбункулах, ей,послеоперационных и посттравматических рубцах, ожогах.
— При заболеваниях кожи (дерматит, нейродермит, герпес, эрозии, язвы слизистых оболочек).
В среднем курс лечения рассчитан на 10 сеансов. В некоторых случаях для закрепления достигнутого эффекта процедура повторяется через полгода.
Флебология
Всем знакомы внешние проявления варикозной болезни вен нижних конечностей — в просторечии варикоза, — набухшие синие узловатые сосуды на ногах. Варикозное расширение вен причиняет значительные неудобства, болезненность, и в конце концов – это просто неэстетично! Какие же есть способы избавиться от этой напасти? На данном этапе развития медицины их всего три: хирургический (удаление вен путем операционного вмешательства), лазерная коагуляция и флебосклерозирующая терапия. За исключением хирургического, процедуры болезненной и применяющейся в крайних и глубоко запущенных случаях, остальные способы давно и успешно применяются для лечения в нашем Центре.
ФЛЕБОСКЛЕРОЗИРУЮЩАЯ ТЕРАПИЯ
Самый распространенный способ лечения варикоза во всем мире. Быстрое достижение хорошего эстетического результата, безболезненность, возможность амбулаторного применения, быстрая реабилитация делают данный вид лечения привлекательным как для врачей, так и для пациентов.
ТЕХНИКА ФЛЕБОСКЛЕРОЗИРУЮЩЕГО ЛЕЧЕНИЯ
При внутривенном введении препаратов, так называемых детергентов, на 8-10 день наступает облитерация варикозной вены, при этом сразу после процедуры необходима немедленная эластическая компрессия – специальные сдавливающие повязки, способствующие «слипанию» варикозной вены и предназначенные для круглосуточного ношения в течение 10-14 дней.
Выбор концентрации флебосклерозирующего препарата определяется диаметром варикозной вены.
Внутрисосудистое введение препаратов практически безболезненное, а сам способ лечения является безопасным, эффективным и радикальным.
ЛАЗЕРНАЯ КОАГУЛЯЦИЯ
Применяется при лечении ретикулярного варикоза (сеть мелких сосудиков на ногах), когда технически невозможно сделать укол в вену диаметром 0,1 -1 мм. При воздействии лазера часть расширенных сосудов исчезает непосредственно в процессе лечения уже во время первого сеанса, но для удаления большинства сосудов может потребоваться до 3-4 сеансов с 4- недельными интервалами.
И, само собой, при лазерном лечении никаких эластичных повязок не требуется.
Таким образом, оптимальный эффект отмечается при сочетанном применении склеротерапии и лазера для обработки сосудов различного диаметра. Ну а какой способ выбрать — решат вместе с Вами высококвалифицированные врачи-флебологи нашего Центра после осмотра и, в случае необходимости, проведения диагностических исследований.
Лазерная очистка крови и сосудов
Предлагаем Вам очень действенную и эффективную методику – ВНУТРИСОСУДИСТОЕ ЛАЗЕРНОЕ ОБЛУЧЕНИЕ КРОВИ (ВЛОК).Данный метод основан на введении ввену через иглу световода, через который протекающая кровь облучается лазерным лучом. При этом происходят активация дыхательных ферментов, увеличение насыщения гемоглобина кислородом, стимуляция выработки форменных элементов крови, улучшение ее микроциркуляции и снижение вязкости, усиление бактерицидных свойств крови. Как общее следствие – детоксикация организма, возрастание его устойчивости к инфекциям.
Показания к применению ВЛОК:
— Хронические заболевания печени и почек
— Хронические бронхиты, пневмонии, бронхиальная астма
— Ишемическая болезнь сердца, постинфарктные состояния
— Заболевания кожи ( псориаз, нейродермит, фурункулез)
— Лекарственная и другие виды аллергий
— Хронические, в т.ч. вирусные, инфекции
Клинически доказана высокая эффективность ВЛОК при следующих заболеваниях:
— Тромбооблитерирующие заболевания артерий конечностей
— Острые и хронические тромбофлебиты, флеботромбозы
— Язвенная болезнь желудка и 12- перстной кишки
— Сахарный диабет
— Хронический панкреатит
— Ревматоидный артрит
— Бесплодие, аднексит
Лечение с помощью ВЛОК может проводиться в сочетании с лекарственной терапией, причем под влиянием лазеротерапии повышается чувствительность к медикаментам, что дает возможность снижения дозировки лекарств и соответственно, уменьшения их побочного воздействия.
Длительность клинического эффекта.
После первого курса ВЛОК результат лечения сохраняется 4 и более месяцев. После повторного курса лазеротерапии ремиссия увеличивается до 6-8 месяцев.
Аппаратное вытяжение позвоночника
Осевое вытяжение и вибрационно-механическое воздействие на мышечно-связочный аппарат позвоночника и суставов , проводимое на аппарате для дозированного вытяжения «ОРМЕД- 3 М» в сочетании с лазерной терапией дает замечательный эффектпри лечении остеохондроза шейного, плечевого и поясничного отделов позвоночника, радикулита, ишиаса, люмбаго, плече- лопаточного периартрита, межпозвонковых грыж, межреберной невралгии.
границ | СЛАВА: Генератор структур вероятных метаболитов цитохрома P450 на основе прогнозируемых участков метаболизма
Введение
Метаболизм отвечает за создание метаболитов с другими физико-химическими и фармакологическими свойствами по сравнению с исходной родительской молекулой. Метаболизм ксенобиотиков, в частности, имеет прямое отношение к человеку, особенно в том, что касается, например, разработки лекарств, косметики и агрохимикатов.Фактически предполагается, что метаболизм является основным путем выведения подавляющего большинства всех ксенобиотиков (Kirchmair et al., 2015). Однако метаболизм может также приводить к образованию фармакологически активных метаболитов, а также токсичных метаболитов (Testa et al., 2012).
Семейство ферментов цитохрома P450 (CYP) играет важную роль в метаболизме ксенобиотиков, особенно в образовании метаболитов первого поколения, из которых примерно 60% формируются из CYP (Testa et al., 2012).Важность CYP для открытия лекарств очевидна из наблюдения, что многие лекарства метаболизируются с помощью CYP; общие оценки варьируются от 50% (Di, 2014) до 80% (Testa et al., 2012). Подробный мета-анализ метаболитов более 1000 различных субстратов ксенобиотиков, проведенный Testa et al., Показал, что 40% всех метаболитов образованы CYP, включая значительную часть всех токсичных или высокореактивных метаболитов (Testa et al. , 2012).
Известно 57 ферментов CYP человека, большинство из которых в основном участвует в эндогенном метаболизме.Подсемейства CYP2 и CYP3 в основном отвечают за метаболизм ксенобиотиков (Testa et al., 2012), а ключевыми изоферментами CYP для метаболизма лекарств являются CYP3A4, 3A5, 2D6, 2C8, 2C9, 2C19, 1A1, 2B6 и 2E1 (Di, 2014). Среди изоферментов CYP, метаболизирующих ксенобиотики, карманы связывания сильно различаются; в некоторых случаях связывающий карман одного изофермента является очень гибким и может вмещать широкий спектр субстратов с широко варьирующимися размерами (Kirchmair et al., 2015).
Вычислительные методы могут внести значительный вклад в прогнозирование метаболизма ксенобиотиков, поскольку их можно использовать для быстрого прогнозирования, которое может сфокусировать экспериментальные аспекты процесса разработки лекарств.Такой эффект фокусировки является экономичным и эффективным по времени (Kirchmair et al., 2015).
Одним относительно хорошо разработанным аспектом компьютерного прогнозирования метаболизма ксенобиотиков является идентификация метаболически лабильных положений атомов, также известных как сайты метаболизма (SoMs) (Kirchmair et al., 2012). Способность предсказать SoM важна, потому что знание положения атома в молекуле, при котором, вероятно, может происходить реакция метаболизма, обычно дает химику хорошее представление о последующей структуре метаболита.Помимо ряда коммерческих предложений, несколько свободно доступных инструментов, таких как SMARTCyp (Olsen et al., 2019), SOMP (Rudik et al., 2015), Xenosite (Zaretzki et al., 2013) и FAME 2 (Šícho et al., al., 2017), могут прогнозировать SoM с высокой точностью (Tyzack, Kirchmair, 2018). FAME 2, который используется в настоящей работе для прогнозирования SoM, представляет собой инструмент на основе машинного обучения, недавно разработанный нашей группой. Модели классификаторов дополнительных деревьев для FAME 2, основанные на наборе двумерных круговых дескрипторов, были разработаны специально для прогнозирования SoM метаболических реакций, катализируемых семейством ферментов CYP у людей.FAME 2 имеет высокую точность, достигая на независимом испытательном наборе коэффициента корреляции Мэтьюза 0,57 и площади под кривой рабочих характеристик приемника (AUC) 0,91.
В отличие от in silico SoM предсказания, вычислительное предсказание структур метаболитов отстает в отношении точности предсказания. В целом в существующих методах прогнозирования структур метаболитов ксенобиотиков преобладают подходы, основанные на правилах. Существует ряд хорошо зарекомендовавших себя коммерческих инструментов для прогнозирования структуры метаболитов, в том числе Meteor Nexus (Lhasa Ltd.), основанное на правилах программное обеспечение для прогнозирования метаболитов (Marchant et al., 2008). Meteor Nexus предлагает три различных метода рассуждения для определения приоритета множества генерируемых метаболитов. Текущий метод рассуждений по умолчанию — это оценка SoM, которая сравнивает SoM, определенную правилом реакции, с экспериментальными данными, чтобы присвоить оценки прогнозируемым метаболитам. Другие основанные на правилах вычислительные инструменты включают TIMES (LMC; Mekenyan et al., 2004), который использует эвристический алгоритм для создания возможных метаболических карт, и MetabolExpert (CompuDrug; Darvas, 1987).
В дополнение к коммерческим инструментам прогнозирования структуры метаболитов, растет число свободно доступных опций. Опять же, многие из доступных вариантов основываются в первую очередь на наборе правил реакции для создания структур возможных метаболитов. Один хорошо известный подход, который существует уже некоторое время, — это SyGMa (Ridder and Wagener, 2008), который в этой работе используется в качестве эталонного метода. SyGMa предсказывает метаболиты, используя основанные на знаниях правила реакции, некоторые из которых были получены из общеизвестных знаний о реакциях метаболизма, а некоторые были разработаны с использованием базы данных Metabolite (База данных MDL Metabolite, Elsevier, 2001), всего 144 правила реакции, охватывающие оба фаза I и фаза метаболизма II.Прогнозируемые метаболиты ранжируются по эмпирическим оценкам вероятности, рассчитанным на основе доли предсказанных метаболитов, произведенных конкретным правилом реакции, которые соответствуют зарегистрированным метаболитам в базе данных. Используя все 144 правила реакции фазы I и фазы II до трех последовательных стадий реакции, SyGMa смог предсказать 68% всех известных метаболитов в тестовой выборке. Что касается ранжирования, SyGMa заняла 45% известных метаболитов в наборе тестов в первой десятке. Авторы дополнительно исследовали потенциальную полезность SyGMa для прогнозирования CYP-опосредованного метаболизма, оценив его эффективность на наборе 127 одноэтапных CYP-опосредованных. реакции.Используя только 118 правил реакции фазы I, которые включают, но не являются специфическими для CYP-опосредованных реакций, SyMGa смог предсказать 84% всех известных метаболитов, образованных CYP, и предсказать 66% известных метаболитов в пределах трех лучших предсказанных метаболитов. . Однако проприетарный характер набора данных, который использовался для вывода правил реакции SyGMa и проверки метода, не говоря уже о текущей недоступности набора данных, препятствует воспроизводимости результатов, а также дальнейшему использованию моделей, полученных на основе данных.
Недавнее бесплатное программное обеспечение, предназначенное для прогнозирования метаболитов из нескольких источников и семейств ферментов, — это BioTransformer (Djoumbou-Feunang et al., 2019), которое в этой работе используется в качестве второго эталонного метода. BioTransformer — это комплексный инструмент для прогнозирования метаболитов, который содержит модуль прогнозирования метаболитов CYP (в дополнение к четырем другим модулям прогнозирования метаболитов). BioTransformer предсказывает образующиеся CYP метаболиты, используя подход, основанный на знаниях, в сочетании со встроенным прогнозированием избирательности CYP CypReact (Tian et al., 2018), инструмент, основанный на машинном обучении, в качестве предшественника прогноза метаболитов. Помимо первоначального прогнозирования специфичности изоформ CYP, в основе модуля прогнозирования метаболитов CYP450 компании BioTransformer лежит метод, основанный на правилах, правила реакции которого частично основаны на метаболических реакциях в MetXBioDB (Djoumbou-Feunang et al., 2019), свободно доступном база данных реакций метаболизма, созданная в контексте разработки Биотрансформатора. В текущей версии BioTransformer прогнозируемые метаболиты не ранжируются.BioTransformer также предлагает возможность определения метаболитов на основе масс-спектрометрических данных. На тестовом наборе данных из 60 родительских молекул с в общей сложности 180 известных метаболитов модуль прогнозирования метаболитов CYP450 компании BioTransformer достиг отзыва 0,90 и точности 0,46.
Еще один свободно доступный инструмент для прогнозирования метаболитов — MetaTox (Rudik et al., 2017), который охватывает как фазу I, так и фазу II метаболизма и объединяет прогноз класса реакции и реагирующего атома для прогнозирования метаболитов.Кроме того, программное обеспечение с открытым исходным кодом Toxtree (Patlewicz et al., 2008) содержит модуль прогнозирования метаболизма под названием «SMARTCyp — Cytochrome P450-Mediated Drug Metabolism», который прогнозирует SoM с помощью SMARTCyp (Rydberg et al., 2010), а затем применяет небольшой набор правил реакции на предсказанные SoM для предсказания метаболитов.
Общим для всех современных подходов к прогнозированию метаболитов является то, что они по-прежнему сталкиваются с проблемой комбинаторного взрыва прогнозов, в частности, при рассмотрении нескольких поколений метаболитов (Judson, 2014).Для предсказателей структуры метаболитов нет ничего необычного в том, что они создают несколько страниц, полных предсказанных метаболитов, что часто и не без оснований критикуется, особенно специалистами по метаболизму. Ключ к решению этой проблемы заключается в разработке подходов для точного ранжирования метаболитов в соответствии с их актуальностью с точки зрения скорости метаболизма и биологических свойств. Ряд методов пытается справиться с огромным количеством предсказанных метаболитов, ранжируя их предсказания в соответствии с различными подходами.
Другой вариант, который на сегодняшний день в основном реализован в коммерческих инструментах, заключается в использовании прогноза SoM в качестве предварительного шага для уменьшения количества генерируемых метаболитов. Коммерческие инструменты для прогнозирования метаболитов, которые включают прогнозирование SoM, включают ADMET Predictor (SimulationsPlus), который прогнозирует SoM и соответствующие структуры метаболитов для девяти изоформ CYP, и StarDrop (Optibrium; Tyzack et al., 2016), чей модуль «Метаболизм P450» прогнозирует SoMs с помощью квантово-механического моделирования и отображает структуры метаболитов, соответствующие прогнозируемым SoM.Кроме того, META Ultra (MultiCASE Inc .; Klopman et al., 1994) предсказывает SoMs и метаболиты, а MetaSite (Cruciani et al., 2005) был программным обеспечением для прогнозирования селективности SoM и изоформ CYP, которое теперь также предсказывает структуры метаболитов.
Немногие свободно доступные методы прогнозирования метаболитов сочетают информацию о прогнозируемых SoM с набором правил. MetaTox прогнозирует классы реакций и реагирующие атомы (то есть, в принципе, SoM) отдельно для каждой родительской молекулы, а затем объединяет прогнозы для генерации метаболитов.Вероятность образования метаболита рассчитывается на основе прогнозируемых вероятностей класса реакции и SoM, прогнозируемого с помощью метода SOMP (Rudik et al., 2015). Однако при валидации MetaTox эффективность предсказания класса реакции и предсказания реагирующего атома рассматривается отдельно, без оценки предсказания самих структур метаболита, и неясно, как именно предсказания класса реакции и реагирующего атома объединяются для генерации метаболита. структура (Рудик и др., 2017). С другой стороны, очевидно, что прогнозирование SoM используется непосредственно в качестве предварительного фильтра перед применением правил реакции в модуле SMARTCyp Toxtree. Однако валидация этого метода не была опубликована.
Что касается доступности наборов правил для предсказания структуры метаболитов, существует несколько существующих свободно доступных наборов правил реакции, описанных в легкодоступном, машиночитаемом формате, таком как SMIRKS, язык преобразования реакции в системе Daylight.Одним из источников правил реакции CYP является модуль SMARTCyp Toxtree, который использует 16 правил реакции и делает SMIRKS бесплатно доступными как часть исходного кода. Большой выбор правил реакции предоставляется в свободно доступном пакете SyGMA Python. Правила реакции четко разделены на правила фазы I и правила фазы II; однако нет никаких указаний на то, какие из 118 правил реакции фазы I конкретно описывают реакции, опосредованные CYP. Кроме того, эти правила были получены из закрытого и больше не распространяемого набора данных.BioTransformer предлагает большое количество специфичных для CYP правил биотрансформации в формате SMIRKS, а также дополнительные ограничения для каждого правила как часть своей базы знаний о реакции.
В этой работе мы представляем многосторонний подход к предсказанию метаболитов, образованных семейством ферментов CYP у людей. Применительно к FAME мы называем этот подход GLORY. Одним из фундаментальных аспектов GLORY является новая, легко интерпретируемая база правил для метаболизма CYP, которая была разработана исключительно на основе научной литературы и базовых знаний химии, не полагаясь на какой-либо набор данных о метаболических реакциях.Кроме того, мы исследовали эффект от использования прогноза SoM в качестве предварительного фильтра для позиций, в которых разрешено применять правила реакции, а также как часть нового подхода к ранжированию прогнозируемых метаболитов. Таким образом, GLORY имеет два режима: MaxCoverage, который ориентирован исключительно на отзыв, и MaxEfficiency, который больше ориентирован на точность. Кроме того, мы проверили GLORY на новом высококачественном, вручную подобранном наборе данных, который представлен в дополнительном материале.
Результаты и обсуждение
Два ключевых аспекта лежат в основе GLORY, целью которой является прогнозирование метаболитов в контексте человеческого метаболизма, опосредованного CYP: правила реакции и прогнозируемые SoM.Что касается аспекта, основанного на правилах, GLORY использует правила реакции для преобразования родительских молекул в их возможные метаболиты. С этой целью мы разработали набор правил, полностью основанный на научной литературе, чтобы гарантировать, что набор правил не зависит от какого-либо конкретного набора данных о метаболизме. Информация о CYP-опосредованных реакциях из литературы была объединена с базовыми знаниями химии для разработки SMIRKS для описания каждого типа реакции. В некоторых случаях, например, для О-деарилирования, для одного типа реакции требовалось несколько СМИРК, в результате чего для 61 типа реакции, присутствующего в нашей коллекции, было всего 73 СМИРКа (дополнительная таблица 1).Мы дополнительно используем простое бинарное различие между обычными и необычными типами реакций, которые подробно обсуждались и отличались друг от друга в Guengerich (2001), и это различие мы смогли экстраполировать на CYP-опосредованные реакции, обнаруженные в других местах в литературе (см. Методы для подробностей). Мы не используем коэффициенты встречаемости, рассчитанные на основе заданного набора данных, для ранжирования прогнозируемых метаболитов из-за ограниченного размера, качества и доступности существующих наборов данных.Из нашей коллекции 61 типа реакции CYP 22 были определены как общие.
Второй ключевой аспект GLORY — это использование вероятностей SoM, предсказываемых FAME 2 для каждого тяжелого атома в молекуле, чтобы (i) снизить частоту ложноположительных прогнозов при сохранении приемлемой скорости восстановления и (ii) повысить рейтинг предсказанные метаболиты. Чтобы снизить частоту ложноположительных прогнозов, была исследована возможность использования жесткого отсечения на основе вероятностей SoM.Это ограничение использовалось, чтобы определить, в каких атомных позициях разрешалось применять правила. В контексте GLORY мы назвали этот подход, в котором прогнозирование SoM используется в качестве предварительного фильтра, режимом MaxEfficiency. Напротив, мы обозначаем подход, в котором вероятности SoM используются для ранжирования метаболитов, полученных для всех положений в молекуле, независимо от вероятности SoM, режимом MaxCoverage. Разница в рабочем процессе между двумя режимами показана на Рисунке 1.
Рисунок 1 .Рабочий процесс для GLORY с указанием разницы между режимом MaxCoverage и режимом MaxEfficiency.
Наборы данных
Чтобы выбрать порог вероятности SoM для режима максимальной эффективности и разработать приоритетную оценку для ранжирования предсказанных метаболитов, был создан большой набор справочных данных путем объединения данных метаболизма CYP, извлеченных из DrugBank (Wishart et al., 2018) и MetXBioDB. MetXBioDB — это недавно опубликованная база данных метаболических реакций, субстраты которой в основном состоят из ксенобиотиков, а также включают несколько липидов стеролов и первичные метаболиты млекопитающих, и данные о реакциях которой взяты из научной литературы, а также из общедоступных баз данных (Djoumbou-Feunang et al. ., 2019). Кроме того, на основе научной литературы был составлен вручную подобранный высококачественный набор данных для проверки GLORY. Этот набор тестовых данных содержит 29 родительских молекул и в общей сложности 81 метаболит, что в среднем дает 2,79 метаболитов на родительскую молекулу. Важно отметить, что все родительские соединения, которые присутствуют в наборе тестовых данных, были удалены из набора контрольных данных до проведения какого-либо анализа. В общей сложности набор справочных данных содержит 848 исходных молекул и 1588 метаболитов, в среднем 1.87 метаболитов на родительскую молекулу. Можно было сделать прогнозы для 847 из 848 молекул в наборе справочных данных (одна молекула не может быть успешно обработана с помощью FAME 2; подробности см. В разделе «Методы»).
Режим максимальной эффективности: выбор порогового значения для создания структуры метаболита на основе вероятности SoM
Чтобы определить влияние предварительного фильтра, основанного на прогнозировании SoM, на прогнозирование предпочтительно только наиболее релевантных метаболитов и уменьшение количества ложноположительных прогнозов, мы попробовали несколько различных пороговых значений для вероятности SoM, которая должна быть достигнута хотя бы одним задействованным атомом. в реакции (как определено СМИРКАМИ реакции).Для каждого тяжелого атома в молекуле FAME 2 сообщает вероятность от 0 до 1, соответствующую доле деревьев классификатора дополнительных деревьев, которые предсказывают, что конкретный атом является SoM. Порог принятия решения в FAME 2 относительно того, считается ли атом достаточно вероятным, чтобы быть SoM, чтобы быть обозначенным как таковой, было определено обученной моделью как 0,4 (Šícho et al., 2017).
Мы исследовали влияние различных пороговых значений вероятности SoM, используя эталонный набор данных, и выбрали пороговое значение, которое будет использоваться в режиме максимальной эффективности, на основе этих результатов.В частности, мы изучили влияние ограничений вероятности SoM на точность и отзывчивость, которые определяются следующим образом:
Вызов = TP / (TP + FN) Точность = TP / (TP + FP), где TP — количество истинно положительных предсказаний, FP — количество (предполагаемых) ложноположительных предсказаний, а FN — количество ложно отрицательных предсказаний. Другими словами, функция отзыва измеряет долю известных метаболитов, которые были воспроизведены данным методом, а прецизионность измеряет долю всех прогнозируемых метаболитов, представленных в наборе данных.
Здесь стоит отметить, что количество ложных срабатываний и обозначение прогноза как ложного срабатывания особенно зависит от набора данных, который используется для сравнения. Многие метаболиты, которые образуются у человека, еще не открыты, или их структура еще не выяснена. Поскольку даже набор данных самого высокого качества ограничен доступными экспериментальными данными, реальность такова, что различие между реальным ложноположительным предсказанием и истинно положительным предсказанием еще неизвестного или несформированного метаболита может оказаться невозможным.Тем не менее, с этой оговоркой, мы оцениваем наш метод на основе имеющихся данных, включая предполагаемые ложные срабатывания.
Целью режима MaxEfficiency является использование ограничения вероятности SoM для прогнозирования метаболитов с повышенной точностью по сравнению с режимом без ограничения (то есть режим MaxCoverage). В то же время, однако, мы не хотели слишком многим жертвовать с точки зрения запоминания, поскольку все еще важно предсказать фактические метаболиты молекулы, даже при уменьшении количества предполагаемых ложноположительных прогнозов.
Для целей прогнозирования метаболитов мы обнаружили, что использование порога принятия решения FAME 2, равного 0,4, в качестве порогового значения для вероятности SoM, привело к относительно низкому отзыву, равному 0,65 (особенно по сравнению с отзывом 0,83, достигнутым в режиме MaxCoverage, как будет обсуждаться ниже). позже в этой работе). Следовательно, несмотря на повышенную точность, обеспечиваемую пороговым значением 0,4, было определено, что это пороговое значение слишком сильно снижает достигнутый отзыв. Поэтому мы дополнительно протестировали более низкие пороги вероятности SoM (Таблица 1).Наблюдая за компромиссом между точностью и отзывом с отсечками в диапазоне от 0,4 до 0,1 и сравнивая их с режимом MaxCoverage, мы определили, что вероятность отсечения SoM 0,2, что дает точность 0,19 и все еще высокий отзыв 0,75. лучший компромисс. Таким образом, на основе результатов, показанных в этом разделе, было зафиксировано ограничение вероятности SoM, равное 0,2 для режима максимальной эффективности. Обратите внимание, что хотя все значения точности, показанные в таблице 1, довольно низки, точность GLORY с использованием порога вероятности SoM сопоставима с точностью существующих методов прогнозирования структуры метаболитов (см. Ниже результаты на наборе тестовых данных).
Таблица 1 . Влияние различных пороговых значений вероятности SoM на точность и отзывчивость для всего набора контрольных данных.
Разработка приоритетного балла для ранжирования прогнозируемых метаболитов для режима MaxCoverage
Чтобы ранжировать предсказанные метаболиты для конкретной молекулы, мы разработали рейтинг приоритета для каждого предсказанного метаболита на основе вероятности SoM атомов, участвующих в трансформации, и того, является ли тип реакции обычным или нет.В частности, учитывается вероятность SoM, рассчитанная FAME 2 для всех атомов в исходной молекуле, которые участвуют в реакции, как определено SMIRKS, и максимальная вероятность SoM среди этих атомов затем включается в оценку, как показано на рисунке 2. . Оценка приоритета рассчитывалась по простой формуле:
. прогнозируемый метаболит = P × F, где P — максимальная вероятность SoM из атомов в исходной молекуле, которые были сопоставлены примененным преобразованием, а F — коэффициент в зависимости от того, был ли тип реакции обозначен как обычный или необычный.В случае, если один и тот же прогнозируемый метаболит возник в результате нескольких преобразований, использовалась максимальная оценка приоритета по всем преобразованиям, приводящим к этому прогнозированию. Оценка более высокого приоритета предназначена для указания более высокой вероятности того, что прогноз верен. Для всех необычных типов реакций F = 1. Фактор F для общих типов реакций влияет на раннее обогащение прогнозов. В частности, раннее обогащение улучшается, когда общим типам реакции придается больший вес в баллах, чем необычным типам, т.е.е. F общий > 1 (рисунок 3). На основе анализа кривых рабочих характеристик приемника (ROC) и площади под кривыми ROC (AUC) для изменения F обычного , показанного на рисунке 3, с коэффициентом 5, в результате чего AUC составляет 0,90, был выбран. Все последующие результаты, основанные на ранжировании предсказанных метаболитов, поэтому использовали F общий = 5 при вычислении оценки приоритета, и поэтому оценка приоритета может варьироваться от 0 до 5.
Рисунок 2 . Иллюстрация определения максимальной вероятности SoM для всех тяжелых атомов в исходной молекуле, которые соответствуют правилу реакции, на примере правила реакции N-деалкилирования (общий тип реакции; коэффициент F = 5). Эта максимальная вероятность используется для расчета рейтинга приоритета продукта.
Рисунок 3 . Кривые рабочих характеристик приемника (ROC) для всего эталонного набора данных из 848 соединений с 1588 известными метаболитами с различными значениями фактора, используемыми для общих типов реакций при вычислении оценки приоритета для каждого метаболита.Обратите внимание, что коэффициент 1 означает, что только вероятность SoM (то есть максимальная вероятность SoM для всех атомов, которые соответствуют SMIRKS) влияет на оценку приоритета предсказанного метаболита, независимо от типа реакции. Также обратите внимание, что кривую ROC можно рассчитать, несмотря на отсутствие «истинно отрицательных» прогнозов в целом (все прогнозируемые метаболиты являются «положительными» прогнозами). Чтобы построить кривую ROC, количество ложных срабатываний (FPR) рассчитывается для каждого порогового значения. В каждой точке предсказания с оценками ниже порогового значения считаются «отрицательными» предсказаниями, а предсказания с оценками выше порогового значения считаются «положительными» предсказаниями.Следовательно, количество «истинно отрицательных» прогнозов и, следовательно, FPR можно рассчитать для каждой точки кривой ROC.
Сравнение производительности на новом, вручную отобранном тестовом наборе
Эффективность режимов MaxEfficiency и MaxCoverage программы GLORY оценивалась на специально отобранном тестовом наборе из 29 исходных молекул, всего 81 метаболита. Эта оценка включает сравнение с BioTransformer и SyGMa, а также анализ того, насколько хорошо работают аспекты оценки и ранжирования различных подходов.В частности, мы использовали модуль CYP450 BioTransformer и реакции метаболизма фазы I SyGMa (SyGMa не имеет специального модуля для метаболизма CYP, но метаболизм фазы I в значительной степени осуществляется ферментами CYP) для сравнения.
Анализ режима максимальной эффективности
Режим максимальной эффективностиGLORY был разработан для решения проблемы низкой точности, вызванной большим количеством предполагаемых ложноположительных прогнозов метаболитов. Эта общая проблема избытка прогнозов хорошо задокументирована для инструментов прогнозирования метаболитов (Judson, 2014).Однако, как упоминалось выше, важно отметить, что определение прогнозов как ложных срабатываний особенно зависит от набора данных.
Как описано ранее, в режиме максимальной эффективности используется ограничение на основе вероятностей SoM, которые FAME 2 предсказывает для каждого тяжелого атома, чтобы ограничить места в молекуле, в которых разрешено применять правила реакции. Это пороговое значение вероятности SoM было установлено на 0,2 на основе анализа контрольного набора данных; тем не менее, мы также исследуем влияние различных пороговых значений вероятности SoM, используя набор высококачественных тестовых данных, чтобы получить более полную картину того, сколько можно получить с помощью подхода, основанного на отсечении.
Как и ожидалось, использование прогнозов SoM для ограничения применения правил реакции к определенным позициям действительно предполагает компромисс между точностью и отзывом (рис. 4). Отзыв измеряет долю известных метаболитов, которую метод смог воспроизвести, а прецизионный измеряет долю всех предсказанных метаболитов, которые на самом деле являются известными метаболитами (определения см. В предыдущем разделе). Чем больше вероятность SoM, необходимая для присутствия среди атомов, участвующих в преобразовании, тем ниже отклик, но тем выше точность, измеренная по всему набору тестовых данных.Кроме того, чем больше предельное значение вероятности SoM, тем больше родительских молекул, для которых невозможно сделать прогноз метаболитов. Без такого ограничения и даже до порогового значения вероятности SoM, равного 0,2, метаболиты могут быть предсказаны для всех родительских молекул в тестовом наборе данных. Однако при пороговом значении вероятности SoM, равном 0,3, метаболиты для двух исходных молекул не прогнозируются, и это число увеличивается до трех для порогового значения 0,4 (дополнительная таблица 2). В этом случае количество затронутых молекул невелико, но составляет примерно 10% от размера тестового набора данных.В целом, по мере увеличения порогового значения общее количество прогнозируемых метаболитов резко снижается (дополнительная таблица 2).
Рисунок 4 . Точность (часть прогнозов, которые являются истинно положительными) и отзыв (часть известных метаболитов, которые прогнозируются) варьируются в зависимости от порогового значения для прогнозируемой вероятности SoM FAME 2. Отсечка вероятности SoM, равная 0,4, соответствует порогу принятия решения, используемому в FAME 2. Отсечка вероятности SoM, выбранная для режима максимальной эффективности GLORY, была равна 0.2.
К сожалению, как показано на Рисунке 4, при использовании пороговых значений вероятности 0,1 или выше при небольшом увеличении точности наблюдается значительное уменьшение отзыва. При более внимательном рассмотрении скоростей восстановления на одну родительскую молекулу, мы видим, что режим максимальной эффективности GLORY (с использованием выбранного порогового значения 0,2, как описано выше) может предсказать, по крайней мере, половину известных метаболитов для 72% родительских молекул в наборе тестовых данных, поскольку по сравнению с 83% для SyGMa и 79% для BioTransformer (рис. 5).Режим максимальной эффективности GLORY может предсказать все известные метаболиты для 41% исходных молекул в наборе тестовых данных, в отличие от 45% для SyGMa и 38% для BioTransformer. С другой стороны, количество предполагаемых ложноположительных результатов на одну родительскую молекулу находится в пределах того же диапазона, что и для SyGMa и BioTransformer (рис. 6). Используя режим MaxEfficiency, большинство родительских молекул имеют менее 10 предполагаемых ложных срабатываний, что также относится к BioTransformer, но не совсем к SyGMa (однако, как упоминалось выше, база правил SyGMa также включает правила для фазы I, не опосредованной CYP. реакции).
Рисунок 5 . Гистограммы скорости восстановления известных метаболитов с разбивкой по исходному соединению: (A) GLORY в режиме MaxCoverage, (B) GLORY в режиме MaxEfficiency, (C) SyGMa, (D) BioTransformer. Например, коэффициент восстановления 0,5 означает, что для x% всех исходных молекул была предсказана по крайней мере половина всех зарегистрированных метаболитов из набора тестовых данных.
Рисунок 6 . Гистограммы количества предполагаемых ложноположительных прогнозов: (A) GLORY в режиме MaxCoverage, (B) GLORY в режиме MaxEfficiency, (C) SyGMa, (D) BioTransformer.Эти гистограммы используют интервалы, закрытые вправо.
На основании этих результатов кажется, что использование предсказанных вероятностей SoM FAME 2 в качестве жесткого ограничения для предсказания метаболитов может оказаться недостаточным для многих случаев использования. Однако прогнозы SoM полезны не только для жесткого ограничения, а именно для ранжирования прогнозируемых метаболитов, как будет показано в следующем разделе.
Сравнение режима MaxCoverage с SyGMa и BioTransformer
Ни SyGMa, ни BioTransformer не используют прогноз региоселективности в качестве предварительного фильтра перед применением правил реакции.То же самое верно и для режима MaxCoverage, который использует только прогнозирование SoM для оценки и ранжирования прогнозируемых метаболитов. Таким образом, мы сравнили SyGMa и BioTransformer с режимом GLORY MaxCoverage с точки зрения запоминания, точности и способности ранжировать предсказанные метаболиты.
Высокая степень запоминания важна для любого варианта использования предсказателя структуры метаболитов, но тем более для приложений, в которых чрезвычайно важно не пропустить какие-либо физически существующие метаболиты, например, при попытке идентифицировать метаболиты на основе Данные МС.Режим GLORY MaxCoverage показывает хорошие результаты с точки зрения отзыва: отзыв составляет 0,83 по сравнению с 0,74 и 0,72 для SyGMa и BioTransformer, соответственно, по всему набору тестовых данных (Таблица 2). Более пристальный взгляд на отзыв, разбитый на уровень скорости восстановления известных метаболитов для каждой родительской молекулы, показывает, что GLORY может предсказать все известные метаболиты для 62% родительских молекул, тогда как SyGMa и BioTransformer достигают только 45% и 38%. , соответственно, в этом плане (рисунок 5). Количество родительских молекул, для которых GLORY может предсказать, по крайней мере, половину известных метаболитов, составляет 90%, по сравнению с 83% для SyGMa и 79% для BioTransformer (рис. 5).
Таблица 2 . Результаты оценки для режимов MaxCoverage и MaxEfficiency в SyGMa, BioTransformer и GLORY на отобранном вручную наборе тестовых данных.
Точность может быть полезным показателем для измерения того, насколько хорошо метод может контролировать количество предполагаемых ложноположительных прогнозов. Точность предсказания метаболитов в наборе тестовых данных была низкой по всем направлениям, при этом BioTransformer достиг наивысшей точности из трех инструментов — 0,17. SyGMa почти не отставала с 0.15, а режим GLORY MaxCoverage еще больше отставал с точностью всего 0,08 (Таблица 2). Снова разбив это на чуть более подробный обзор, мы видим, что и BioTransformer, и SyGMa всегда дают менее 25 предполагаемых ложных срабатываний на одну родительскую молекулу и, для большинства родительских молекул, менее 15 предполагаемых ложных срабатываний или даже, в случае Биотрансформатора менее 10 (рис. 6). С другой стороны, GLORY в режиме MaxCoverage часто дает столько прогнозов на одну родительскую молекулу, что в наборе тестовых данных имеется до 53 предполагаемых ложноположительных результатов на одну родительскую молекулу и в среднем относительно большое количество предполагаемых ложноположительных прогнозов по сравнению с другими. два инструмента (рисунок 6).
В случае низкой точности, наблюдаемой для SyGMa, важно отметить, что набор правил SyGMa не специфичен для CYP-опосредованного метаболизма, а скорее охватывает метаболизм фазы I в целом. Это может указывать на то, что SyGMa могла бы достичь более высокой точности, если бы использовались только правила, специфичные для CYP.
Модуль прогнозирования CYP450BioTransformer, который имеет наивысшую точность из всех трех методов, использует прогнозирование изоформ в качестве предварительного фильтра. Только соответствующие реакции для предсказанной метаболизирующей изоформы (ов) CYP применяются к родительской молекуле, что может способствовать наблюдаемой точности.
Хотя точность режима MaxCoverage (а также SyGMa и BioTransformer) оказалась низкой, а высокие уровни ложноположительных прогнозов в целом проблематичны, в случае предсказателей структуры метаболитов низкая точность проблематична только в том случае, если нет способа различать истинно положительные и предполагаемые ложноположительные предсказанные метаболиты. Это различие может быть достигнуто с помощью хорошо работающего ранжирования прогнозируемых метаболитов, что позволяет избежать необходимости уменьшать общее количество прогнозируемых метаболитов.Следовательно, важно, чтобы инструмент прогнозирования метаболитов мог ранжировать прогнозируемые метаболиты с точки зрения вероятности появления.
GLORY оценивает свои предсказанные метаболиты частично на основе максимальной вероятности SoM всех атомов, участвующих в реакции, а также принимает во внимание тип реакции (см. Выше для более подробного описания оценки приоритета). SyGMa использует эмпирические оценки вероятности, рассчитанные на основе процента всех прогнозов для каждого правила реакции, найденных в наборе обучающих данных.Таким образом, система подсчета очков SyGMa полностью полагается на прекращенный набор данных Metabolite. Оценки, полученные с помощью GLORY или SyGMa, можно использовать для ранжирования предсказанных метаболитов для данного исходного соединения с точки зрения вероятности их появления. Текущая версия BioTransformer, с другой стороны, не оценивает и не ранжирует свои прогнозы.
Мы сравнили возможности ранжирования в режиме MaxCoverage в GLORY и в SyGMa. SyGMa смогла предсказать известный метаболит в трех верхних ранжированных позициях для 69% родительских молекул в тестовом наборе данных, тогда как режим GLORY MaxCoverage предсказал известный метаболит в пределах трех верхних прогнозов для 76% родительских молекул (таблица 2). .
Чтобы посмотреть на общее качество оценки, а также на ранжирование способности SyGMa по сравнению с GLORY, мы построили кривые ROC для каждого метода, используя оценку каждого предсказанного метаболита, а также ранг каждого предсказанного метаболита для данной молекулы. Ранговый анализ лучше соответствует реальному варианту использования, в котором желательно расставить приоритеты для прогнозируемых метаболитов для конкретной родительской молекулы, а не для всего набора данных [обратите внимание, что SyGMa первоначально оценивалась только с точки зрения ранжирования для каждого родителя молекула (Риддер, Вагенер, 2008)].Однако мы дополнительно использовали основанную на оценках кривую ROC для визуализации эффективности оценки приоритета GLORY по всему набору тестовых данных. Чтобы лучше сравнить кривые ROC, ложноотрицательные результаты были включены в кривые ROC и, таким образом, в рассчитанные AUC путем добавления этих молекул к набору точек данных и искусственного присвоения им балла 0 или ранга 1000, в зависимости от обстоятельств. для этой оценки.
Хотя значения AUC низкие, частично из-за включения ложноотрицательных точек данных в кривые ROC, кривые ROC показывают гораздо лучшее раннее обогащение для GLORY, чем для SyGMa (рис. 7).SyGMa не ставит известный метаболит в лучшую позицию для любой родительской молекулы в наборе тестовых данных (таблица 2), что отражено на кривой ROC. Это приличное раннее обогащение с помощью GLORY, которое подтверждается значением топ-3, является весьма обнадеживающим результатом, указывающим на то, что наиболее вероятные прогнозы ближе к началу ранжированного списка, чем предполагаемые ложноположительные прогнозы.
Рисунок 7 . Кривые ROC по всему набору тестовых данных сравнивают подходы оценки (A), и (B) SyGMa с режимом GLORY MaxCoverage.Для лучшего сравнения двух методов ложноотрицательные результаты были включены в кривую ROC путем присвоения этим точкам данных балла 0 или ранга 1000, в зависимости от обстоятельств.
Одно из возможных объяснений того, почему SyGMa плохо работает с точки зрения оценки, может заключаться в том, что его схема оценки была получена из соотношений встречаемости в базе данных Metabolite и поэтому оптимизирована для прогнозирования метаболитов в этом конкретном наборе данных. Хотя база данных Metabolite была большой, авторы SyGMa сообщают, что база данных, тем не менее, была смещена в сторону соединений с одним известным метаболитом, и постулируют, что многие профили метаболитов были неполными (Ridder and Wagener, 2008).Наш вручную подобранный набор тестовых данных состоит из родительских молекул с метаболитами, которые были опубликованы с 2014 года, в то время как SyGMa была разработана с использованием версии Metabolite 2001 года, поэтому мы предполагаем, что перекрытие, если оно есть, между набором обучающих данных SyGMa и нашим тестовым набором данных невелико. Без доступа к набору данных, который использовался для разработки методологии оценки SyGMa, остается неясным, насколько хорошо типы реакций, которые приводят к метаболитам в тестовом наборе данных, были представлены в их наборе обучающих данных.В связи с этим, дополнительным недостатком подхода SyGMa, основывающего свой подход к оценке на базе данных метаболических реакций, является то, что, поскольку правила реакции могут быть включены только в том случае, если база данных содержит достаточно примеров определенного типа реакции для расчета оценки вероятности, что более необычно. типы реакций или типы реакций, которые по какой-то причине недостаточно хорошо представлены в базе данных, могут отсутствовать в базе правил SyGMa (Ridder and Wagener, 2008).
Есть несколько других различий в методологии между GLORY и SyGMa, которые могут способствовать разнице в производительности.Во-первых, SyGMa не предсказывает конкретно CYP-опосредованный метаболизм, а скорее метаболизм фазы I в целом, что означает, что он может предсказывать другие метаболиты фазы I, которые просто не присутствуют в тестовом наборе данных, потому что они не образуются CYP. Во-вторых, в текущей реализации пакета Python, который использовался для этой проверки, SyGMa, похоже, не требует, чтобы его предсказанные метаболиты имели определенный минимальный размер. В отличие от GLORY, который не выводит потенциальный метаболит, если в нем меньше трех тяжелых атомов, SyGMa предсказывает несколько метаболитов (по всему набору тестовых данных) только с одним или двумя тяжелыми атомами.
Время вычисления
Время работы GLORY было измерено на рабочей станции, оснащенной восемью процессорами Intel (R) Core (TM) i7-4790, 32 ГБ оперативной памяти и операционной системой Linux. Для тестового набора данных общее время выполнения (с использованием восьми ядер) составило 4,6 мин в режиме MaxCoverage и 4,3 мин в режиме MaxEfficiency (усредненное по трем прогонам). В среднем время вычисления на молекулу, необходимое для прогнозирования метаболитов, составляло 10,9 с для режима MaxCoverage и 10,3 с для режима MaxEfficiency (усредненное по трем прогонам).
Методы
Разработка сборника преобразований
Набор преобразований, определенных СМИРКС и представляющих типы реакций, был собран на основе известных CYP-опосредованных реакций, обнаруженных в литературе (подробности см. В дополнительных материалах). SMIRKS были определены как максимально общие, ограничиваясь разумной химией реакции, как указано в литературе и общепринятыми химическими знаниями. Следовательно, если реакция была обнаружена в литературе, но было неясно, как эта реакция применима к другим молекулам, помимо предоставленного примера, реакция исключалась из коллекции.Так было для большинства реакций с участием больших кольцевых систем, а также слияния колец и сжатия колец. В частности, из нашей коллекции были исключены следующие типы реакций: реакции, которые оказались одноэлементными, реакции с участием более чем двух конденсированных колец, не являющихся частью основной цепи стероида, слияния колец, сжатия колец, реакции, в которых субстрат или продукт представляет собой радикал, и реакции, конкретно указанные, были обнаружены только в случае изоферментов CYP растений.
Некоторые СМИРКИ, используемые для описания преобразований, были взяты из модуля Toxtree SMARTCyp. Однако большая часть СМИРКОВ была разработана специально для GLORY. При разработке выражений СМИРКС было уделено внимание включению как можно меньшего количества атомов в явное отображение, поскольку вероятности SoM учитывались для всех атомов в отображении.
Каждый тип реакции был обозначен как «обычный» или «необычный». По возможности этот ярлык присваивался в соответствии с классификацией реакций Генгерихом в его обзоре CYP-опосредованных реакций 2001 г. (Guengerich, 2001), который четко разделил реакции на эти две категории.Если тип реакции не был описан в этой публикации, была выбрана метка «общий» или «необычный» на основе экстраполяции (на основе эмпирического сходства с типами реакций, представленными в публикации).
Наша коллекция правил реакции CYP состоит из 61 типа реакции. В некоторых случаях для описания одного и того же типа реакции требовалось несколько преобразований, что приводило в общей сложности к 73 преобразованиям в наборе определенных реакций. Полный список типов реакций и их СМИРК можно найти в дополнительной таблице 1.
Программа прогнозирования метаболитов
Прогнозирование структуры метаболитов включает применение правил реакции во всех соответствующих положениях. Соответствующие позиции определяются самим правилом реакции и, в случае режима максимальной эффективности, вероятностью SoM, предсказанной для каждого тяжелого атома. В режиме MaxCoverage вероятности SoM также используются для оценки предсказанных метаболитов.
Прогнозирование SoM с FAME 2
Прогнозы SoM были выполнены с использованием программного обеспечения FAME 2 (Šícho et al., 2017), который включал предварительную обработку молекул. Обученная модель circCDK_ATF_6, которая показала лучшую среднюю производительность во время независимой проверки набора тестов в Šícho et al. (2017), использовался для прогнозирования SoM в GLORY.
Применение преобразований
Превращения родительских молекул в предсказанные метаболиты на основе определенных строк SMIRKS были выполнены с использованием Ambit-SMIRKS [Kochev et al., 2018; Библиотека Java Ambit-SMARTS, версия 3.1.0. http: // ambit.sourceforge.net/smirks.html (по состоянию на 4 октября 2017 г.)]. Некоторые преобразования могут привести к созданию нескольких продуктов. Продукты, содержащие менее трех тяжелых атомов, не входят в набор прогнозируемых метаболитов, генерируемых GLORY.
Когда прогнозирование SoM используется в качестве предварительного фильтра, правило преобразования применяется только в определенном месте в родительской молекуле, если один из задействованных тяжелых атомов прогнозируется как SoM с вероятностью, превышающей определенный порог (подробнее см. информацию об этом пороге).
Оценка предсказанных метаболитов
Оценка предсказанных метаболитов была основана на предсказаниях вероятности SoM и на том, был ли тип реакции определен как обычный или необычный. Каждому атому в родительской молекуле была присвоена вероятность того, что он является SoM от FAME 2. При применении преобразований, определенных SMIRKS, Ambit-SMIRKS сопоставляет реагирующую часть определенного преобразования с любым совпадающим набором атомов в родительской молекуле. В рамках этого сопоставления была рассчитана максимальная вероятность SoM и использована для оценки прогнозируемого метаболита, полученного в результате этого конкретного преобразования и сопоставления.
Для каждого прогнозируемого метаболита оценка приоритета рассчитывается путем умножения максимальной вероятности SoM в отображении на коэффициент F в зависимости от того, был ли тип реакции классифицирован как «обычный» или «необычный». Таким образом, оценки приоритета для прогнозируемых метаболитов варьируются от 0 до F обычный . Чем выше оценка, тем более вероятным считается предсказанный метаболит. Дополнительные сведения о выборе значений для F см. В разделе «Результаты».
Если несколько преобразований данной исходной молекулы приводят к одной и той же структуре метаболита, оценка приоритета рассчитывается отдельно в каждом случае, и сохраняется наивысшая оценка. Чтобы вычислить максимальные значения k и кривые ROC на основе ранжирования, необходимо было ранжировать предсказанные метаболиты для каждой исходной молекулы на основе их оценок приоритета. Если разные метаболиты одного и того же исходного соединения имеют одинаковую оценку приоритета, то они получают одинаковый рейтинг. В случае ничьей пропускаются одно или несколько ранговых номеров в соответствии с количеством связанных прогнозов, следующих за равным рангом.Например, если наивысший балл составляет 2,5 и оба прогнозируемых метаболита имеют этот балл, то обоим этим метаболитам присваивается ранг 1, ни одному из прогнозируемых метаболитов не присваивается ранг 2, а прогнозируемым (ым) метаболитам (-ам) присваивается следующий балл. наивысший балл присваивается 3 место.
Программный вывод
Прогнозируемые метаболиты предоставляются в виде файла SD со следующей информацией для каждого прогнозируемого метаболита: ранг (из всех прогнозируемых метаболитов для конкретной родительской молекулы), оценка приоритета, название реакции, а также InChI, SMILES и идентификатор родительского молекула.Если несколько преобразований привели к одному и тому же продукту, сообщается оценка наивысшего приоритета и соответствующее название реакции. Если входные данные состоят из нескольких молекул, идентификатор родительской молекулы устанавливается в соответствии с положением молекулы в упорядоченном списке входных молекул (т. Е. Ее положением во входном файле).
Создание эталонного набора данных
Контрольный набор данных был создан путем объединения данных метаболизма CYP из DrugBank и MetXBioDB. Общий размер объединенного набора справочных данных, не включая информацию о метаболизме для любой из родительских молекул, содержащихся в вручную подобранном наборе тестовых данных, составляет 848 родительских молекул и 1588 метаболитов (в среднем 1.87 метаболитов на родительскую молекулу).
Набор данных DrugBank
База данных DrugBank (DrugBank, версия 5.1.2. Https://www.drugbank.ca/ [доступ 14 января 2019 г.]) была загружена с веб-сайта. Помимо базы данных в формате XML, структуры всех молекул, как исходных, так и метаболитов, были загружены в формате SD с веб-сайта (группа препаратов «Все» для исходных молекул).
Любая исходная молекула или молекула метаболита, не имеющая доступной структуры, игнорировалась.Одно исходное соединение (ID банка DrugBank: DB09327) было проигнорировано, поскольку его SMILES содержал два компонента, из которых нельзя было однозначно идентифицировать основной компонент. Учитывались все доступные поколения реакций метаболизма при условии, что реакция была аннотирована как опосредованная одним или несколькими изоферментами CYP. Ферменты для реакций, перечисленные в DrugBank, не имеют какой-либо очевидной информации о видах, поэтому все они были предположительно человеческими и, следовательно, значимыми для этого набора данных.
Для всех реакций, опосредованных CYP, реагент считался исходной молекулой, а продукт считался метаболитом первого поколения этой конкретной родительской молекулы.Любой метаболит с тем же InChI, игнорируя информацию о стереохимии, поскольку его родительская молекула была удалена из набора метаболитов для этой родительской молекулы. В окончательный набор данных были включены только те родительские молекулы, которые содержат хотя бы один действующий метаболит.
Наконец, шесть родительских молекул, которые также присутствуют в вручную подобранном наборе тестовых данных, были удалены из набора данных DrugBank перед любой оценкой вместе с соответствующей информацией о метаболизме. Этими исходными соединениями были бупропион, тиклопидин, имипрамин, ифосфамид, бозентан и оланзапин.
После предварительной обработки, включая удаление совпадений с вручную подобранным набором тестовых данных, набор данных DrugBank содержал в общей сложности 364 родительских молекулы и 702 метаболита, в среднем 1,93 метаболита на родительскую молекулу в наборе данных.
Набор данных MetXBioDB
CYP-опосредованные реакции человека были извлечены из набора данных MetXBioDB (MetXBioDB, версия 1.0. Https://bitbucket.org/djoumbou/biotransformerjar/src/master/database/ [доступ 11 января 2019 г.]).Поскольку единственная структурная информация, представленная в MetXBioDB, находится в форме InChIs и InChIKeys, любой субстрат или продукт без указанного InChI не может рассматриваться. Отсутствие InChI характерно только для одной из 1468 CYP-опосредованных реакций человека в MetXBioDB.
Информация о стереохимии была удалена путем создания InChI без слоя стереохимии, в результате чего в общей сложности было получено 751 CYP, исходное соединение человека. Из них 259 также присутствуют в наборе данных DrugBank. Для этих перекрывающихся родительских соединений 512 из 569 метаболитов DrugBank также находятся в MetXBioDB, а MetXBioDB имеет еще 93 метаболита для этих перекрывающихся исходных соединений.
Восемь исходных соединений (оланзапин, бупропион, метоклопрамид, бозентан, имипрамин, тиклопидин, ифосфамид и атомоксетин) из вручную подобранного набора тестовых данных также присутствовали в наборе данных MetXBioDB, только два из которых (метоклопрамид и атомоксетин) также присутствовали в набор данных DrugBank. Эти исходные соединения и соответствующие данные о метаболизме были удалены из набора данных MetXBioDB.
После предварительной обработки, включая удаление пересечения с вручную подобранным набором тестовых данных, набор данных MetXBioDB содержал в общей сложности 743 родительских молекулы и 1385 метаболитов, в среднем 1.86 метаболитов на родительскую молекулу в наборе данных.
Слияние наборов данных DrugBank и MetXBioDB
Набор данных DrugBank и набор данных MetXBioDB были объединены в базовый набор данных путем прямого объединения информации о родителях и метаболитах. Все сравнения молекул проводились с использованием InChI, созданных без информации о стереохимии. Для любой родительской молекулы, которая присутствовала как в наборах данных DrugBank, так и в наборах данных MetXBioDB, что имело место для 259 родительских молекул, наборы метаболитов из обоих наборов данных были объединены без учета стереохимии, чтобы получить окончательный набор метаболитов для этой родительской молекулы в эталонный набор данных.
Создание вручную подобранного набора тестовых данных
Новый набор данных для тестирования GLORY был вручную собран из научной литературы. Данные были взяты из публикаций по метаболизму, которые были найдены в двух журналах: Xenobiotica и Drug Metabolism and Disposition . Рассматриваемые временные рамки: с января 2014 г. по июнь 2018 г. для Xenobiotica и с января 2014 г. по июнь 2017 г. для Метаболизм и утилизация лекарственных средств .
Было выбранопубликаций, и содержащаяся в них информация о метаболизме была включена в набор данных, если выполнялись следующие критерии:
1. Публикация должна содержать рисунок, изображающий схему метаболизма и включающий химические структуры исходного соединения и метаболитов первого поколения.
2. Данные о метаболизме должны быть экспериментально определены на человеческом источнике (т.е. на людях, человеческих клетках или рекомбинантных человеческих ферментах CYP).Если некоторые, но не все данные были получены от людей, любые нечеловеческие метаболиты в схеме метаболизма были исключены из набора данных.
3. По крайней мере, для 75% всех метаболитов человека первого поколения, представленных в схеме метаболизма (обратите внимание, что любой метаболит, который изображен как просто промежуточный продукт, не рассматривается), должны быть удовлетворены следующие два критерия. Во-первых, необходимо знать тип фермента (ов), ответственного за образование метаболита. Для этого достаточно знать, образуется ли этот метаболит CYP.Во-вторых, должна быть известна точная химическая структура метаболита, включая взаимосвязь всех атомов. Есть одно исключение из этого правила: если известно, что метаболит не образует CYP, то точная структура не имеет значения, и метаболит все равно считается.
На основе этих критериев были найдены и включены в набор данных 29 схем метаболизма, содержащих по крайней мере один человеческий CYP-образованный метаболит первого поколения с полностью определенной структурой. Для этих 29 родительских молекул в общей сложности имеется 81 метаболит, которые удовлетворяют критериям (первое поколение, человеческий, CYP, полностью определенная структура) для включения в набор данных.Обратите внимание, что в набор данных включены только метаболиты первого поколения. Также обратите внимание, что промежуточные продукты, как показано на схеме метаболизма, не включены в набор данных. Вместо этого используется первый метаболит, не являющийся промежуточным звеном пути.
УЛЫБКИ для метаболитов были получены с использованием ChemSpider (ChemSpider. Http://www.chemspider.com/ [доступ 13 февраля 2019 г.]). Согласованность стереохимической информации между родителями и их метаболитами сохранялась.
Проверка предсказателей структуры метаболита
Прогнозируемые метаболиты сравнивались с известными метаболитами из эталонных и тестовых наборов данных с использованием их InChI.InChI, используемые для этого сравнения, были сгенерированы без информации о стереохимии с использованием CDK (Willighagen et al., 2017; Chemistry Development Kit, версия 2.0. Https://cdk.github.io/ [доступ 3 ноября 2017 г.]).
Во время валидации предсказанный метаболит альдегида считался эквивалентным соответствующей карбоновой кислоте, потому что есть свидетельства того, что некоторый процент метаболита альдегида действует как промежуточное соединение, которое далее окисляется до карбоновой кислоты, не покидая активного центра фермента CYP (Bell -Парих и Генгерих, 1999).
В случае одной родительской молекулы в наборе справочных данных невозможно сделать никаких прогнозов, поскольку родительская молекула содержит бор. FAME 2 не может делать прогнозы для молекул, содержащих бор, потому что борсодержащие молекулы не присутствовали в наборе данных, используемом для обучения модели.
Прогнозы SyGMa выполнялись на Python с использованием пакета SyGMa Python (SyGMa, версия 1.1.0) и RDKit (RDKit: Open-Source Cheminformatics, версия 2017_03_01, 2017). Использовали только набор правил реакции фазы I и применяли один цикл реакции.
Прогнозы BioTransformer были выполнены с использованием режима CYP450 программы BioTransformer (BioTransformer, версия 1.0.8. Https://bitbucket.org/djoumbou/biotransformerjar/src/master/ [доступ 5 февраля 2019 г.]). BioTransformer запускался индивидуально для каждого исходного соединения с использованием одного ввода SMILES.
Кривые ROC были построены с использованием пакета ROCR R (Sing et al., 2005; ROCR, версия 1.0-7, 2015). Когда к кривой добавлялись ложноотрицательные точки данных, этим точкам данных присваивался балл 0 или ранг 1000, соответственно, в зависимости от того, представляла ли кривая ROC баллы или ранги.
Выводы
Мы разработали GLORY, новый инструмент для прогнозирования структур человеческих метаболитов, образованных CYP. GLORY включает в себя две ключевые идеи: основанный на литературе сборник правил CYP-опосредованных реакций и предсказание SoM, которое особенно успешно использовалось для разработки нового подхода к оценке предсказанных метаболитов.
Для GLORY мы разработали новую коллекцию из 73 правил реакций, описывающих 61 тип реакции для CYP-опосредованного метаболизма. При разработке этой коллекции мы уделяли приоритетное внимание воспроизводимости нашего набора правил и поэтому основывали правила на научной литературе, а не на каком-либо наборе данных.Помимо самих правил, каждый тип реакции был обозначен как обычный или необычный, опять же на основании научной литературы, а не набора данных.
Кроме того, мы разработали оценку приоритета для прогнозируемых метаболитов на основе прогнозируемых вероятностей SoM и простого, основанного на литературе различия между обычными и необычными типами реакций. Следовательно, ни наш набор правил, ни наш подход к оценке не основаны непосредственно на каком-либо наборе данных о метаболических реакциях, что отличает наш подход от других инструментов, например SyGMa, который использует правила реакции и коэффициенты встречаемости, полученные из собственного набора данных, и BioTransformer, правила которого были в некоторой степени на основе свободно доступного набора данных.
GLORY имеет два режима: MaxEfficiency, который использует прогнозирование SoM в качестве предварительного фильтра для позиций в молекуле, в которых разрешены реакции, и MaxCoverage, который не использует предварительный фильтр, а вместо этого фокусируется на высоком уровне отзыва и точном ранжировании предсказанные метаболиты. Использование прогноза SoM в качестве предварительного фильтра, то есть в режиме максимальной эффективности, не работает так хорошо, как можно было бы ожидать, с точки зрения уменьшения количества предполагаемых ложноположительных прогнозов при сохранении высокой скорости восстановления сообщенных метаболитов.Однако, разрабатывая оценку приоритета для прогнозируемых метаболитов с использованием прогноза SoM в сочетании с простым двоичным различием между обычными и необычными типами реакций, мы можем ранжировать метаболиты, прогнозируемые в режиме MaxCoverage, в той степени, в которой GLORY может предсказать хотя бы один известный метаболит. в трех лучших ранжированных позициях для 76% молекул в независимом тестовом наборе при достижении отзыва 0,83. Режим GLORY MaxCoverage превосходит SyGMa и BioTransformer с точки зрения отзыва и превосходит SyGMa с точки зрения ранжирования (BioTransformer в настоящее время не оценивает свои прогнозы метаболитов).Одним из вариантов использования режима MaxCoverage может быть, например, идентификация метаболитов по данным масс-спектрометрии.
Наряду с набором правил реакции, мы предоставляем новый, вручную подобранный набор тестовых данных для бесплатного использования в качестве набора данных для тестирования. Кроме того, GLORY находится в свободном доступе в качестве веб-сервера по адресу https://acm.zbh.uni-hamburg.de/glory/.
Важно отметить, что концепция GLORY такова, что ее можно расширить для прогнозирования метаболитов, образованных ферментами, не принадлежащими к семейству CYP.Ферменты, на которые можно распространить этот подход, в принципе ограничены только объемом доступных данных и охватом соответствующих метаболических реакций инструментами прогнозирования SoM.
Доступность данных
В данном исследовании были проанализированы общедоступные наборы данных. Эти данные можно найти здесь: https://bitbucket.org/djoumbou/biotransformerjar/ и https://www.drugbank.ca/.
Примечание автора
Веб-сервис GLORY доступен по следующему адресу: https: // acm.zbh.uni-hamburg.de/glory/.
Авторские взносы
CdBK и JK: концептуализация; CdBK, CS и JK: методология; CdBK, CS, MŠ, NK и NJ: разработка программного обеспечения; CdBK: проверка; JK и DS: ресурсы; CdBK: курирование данных; все авторы: написание — подготовка оригинального проекта; CdBK: визуализация; DS, NJ и JK: наблюдение; JK: администрирование проекта; DS и JK: привлечение финансирования.
Финансирование
CS и JK поддерживаются Deutsche Forschungsgemeinschaft (DFG, Немецкий исследовательский фонд) — проект номер KI 2085 / 1-1.DFG — это немецкая организация, финансирующая исследования, поддерживающая научные, инженерные и гуманитарные исследования. JK также поддерживается Бергенским исследовательским фондом (BFS) [BFS2017TMT01]. BFS предоставляет гранты на исследования и поддержку исследовательской деятельности в Университете Бергена (UiB) и Университетской больнице Хаукеланда (HUS), а также в других норвежских исследовательских учреждениях, которые сотрудничают с учреждениями в Бергене. Фонд также предоставляет гранты для поддержки исследований в UiB и HUS на стыке фундаментальных и клинических исследований.MŠ и DS поддерживаются Министерством образования, молодежи и спорта (MEYS) — номера проектов MSMT № 20-SVV / 2017, LM2015063 и RVO: 68378050-KAV-NPUI. MEYS отвечает за государственное управление в сфере образования, за разработку политики в области образования, молодежи и спорта, а также за международное сотрудничество в этих областях. Это также центральный административный офис, отвечающий за исследования и разработки, и один из основных государственных органов по координации и финансированию исследований в Чешской Республике.
Заявление о конфликте интересов
NJ является основателем и совладельцем Ideaconsult Ltd.и является техническим менеджером компании с 2009 года. Н.К. работает в Ideaconsult Ltd. по совместительству.
Остальные авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Крейга Нокса, Майкла Уилсона и Дэвида Вишарта из Университета Альберты, AB, Канада, и OMx Personal Health Analytics Inc., AB, Канада за отправку нам структур метаболитов в базе данных DrugBank до их бесплатного размещения. доступно онлайн.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fchem.2019.00402/full#supplementary-material
Дополнительные таблицы. Правила реакции и дополнительные результаты оценки (DOCX).
Дополнительный лист данных 1. Набор тестовых данных, содержащий SMILES для исходных молекул и метаболитов, а также ссылки на публикации (CSV).
Дополнительный лист данных 2. Контрольный набор данных, содержащий SMILES (с информацией о стереохимии), InChI (без информации о стереохимии), а также идентификаторы DrugBank и MetXBioDB для родительских молекул и метаболитов (JSON).
Сокращения
AUC — площадь под кривой рабочей характеристики приемника; CYP, цитохром P450; ROC, рабочая характеристика приемника; SoM, место метаболизма.
Сноски
Список литературы
Белл-Парих, Л. К., и Генгерих, Ф.П. (1999). Кинетика катализируемого цитохромом P450 2E1 окисления этанола до уксусной кислоты через ацетальдегид. J. Biol. Chem. 274, 23833–23840. DOI: 10.1074 / jbc.274.34.23833
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cruciani, G., Carosati, E., De Boeck, B., Ethirajulu, K., Mackie, C., Howe, T., et al. (2005). MetaSite: понимание метаболизма в цитохромах человека с точки зрения химика. J. Med. Chem. 48, 6970–6979. DOI: 10.1021 / jm050529c
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дарвас, Ф. (1987). «Metabolexpert: экспертная система для прогнозирования метаболизма веществ», в QSAR in Environmental Toxicology — II , ed K. L.E. Kaiser, (Dordrecht: Springer), 71–81.
Google Scholar
Джумбу-Феунанг, Ю., Фьямончини, Дж., Хиль-де-ла-Фуэнте, А., Грейнер, Р., Манах, К., и Вишарт, Д. С. (2019). BioTransformer: комплексный вычислительный инструмент для прогнозирования метаболизма малых молекул и идентификации метаболитов. J. Cheminform. 11: 2. DOI: 10.1186 / s13321-018-0324-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джадсон, П. Н. (2014). «Основанные на знаниях подходы для прогнозирования участков и продуктов метаболизма», в Drug Metabolism Prediction , ed J. Kirchmair, Methods and Principles in Medicinal Chemistry (Weinheim: Wiley-VCH), 293–318.
Google Scholar
Кирхмайр, Дж., Геллер, А. Х., Ланг, Д., Кунце, Дж., Теста, Б., и Уилсон, И. Д. (2015). Прогнозирование метаболизма лекарств: эксперимент и / или расчет? Nat. Rev. Drug Disc. 14, 387–404. DOI: 10.1038 / nrd4581
CrossRef Полный текст | Google Scholar
Кирчмайр, Дж., Уильямсон, М. Дж., Тайзак, Дж. Д., Тан, Л., Бонд, П. Дж., Бендер, А. и др. (2012). Вычислительное прогнозирование метаболизма: сайты, продукты, динамика и механизмы фермента S.A.R., P450. J. Chem. Сообщить. Модель. 52, 617–48. DOI: 10.1021 / ci200542m
CrossRef Полный текст | Google Scholar
Клопман, Г., Димаюга М. и Талафус Дж. (1994). МЕТА. 1. Программа для оценки метаболического преобразования химических веществ. J. Chem. Сообщить. Комп. Sci. 34, 1320–1325.
PubMed Аннотация | Google Scholar
Кочев Н., Аврамова С., Желязкова Н. (2018). Амбит-СМИРКС: программный модуль для представления реакций, поиска реакций и преобразования структур. J. Cheminform. 10:42. DOI: 10.1186 / s13321-018-0295-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маршан, К.А., Бриггс, К. А., Лонг, А. (2008). In silico Инструменты для обмена данными и знаниями о токсичности и метаболизме: Derek для Windows, Meteor и Vitic. Toxicol. Механизмы Методы . 18, 177–187. DOI: 10.1080 / 15376510701857320
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мекенян, О. Г., Димитров, С. Д., Павлов, Т. С., Вейт, Г. Д. (2004). Системный подход к моделированию метаболизма в вычислительной токсикологии. I. Среда эвристического моделирования TIMES. Curr. Фармацевтический дизайн . 10, 1273–1293. DOI: 10.2174 / 1381612043452596
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Олсен, Л., Монтефиори, М., Тран, К. П., и Йоргенсен, Ф. С. (2019). SMARTCyp 3.0: усовершенствованный сервер прогнозирования места метаболизма цитохрома P450. Биоинформатика . DOI: 10.1093 / биоинформатика / btz037. [Epub перед печатью].
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Патлевич, Г., Jeliazkova, N., Safford, R.J, Worth, A.P, and Alexiev, B. (2008). Оценка реализации схемы классификации Крамера в программном обеспечении Toxtree. SAR QSAR Environ. Res. 19, 495–524. DOI: 10.1080 / 10629360802083871
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Риддер, Л., и Вагенер, М. (2008). SyGMa: сочетание экспертных знаний и эмпирической оценки в прогнозировании метаболитов. Chem. Med. Chem . 3, 821–832.DOI: 10.1002 / cmdc.200700312
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рудик А., Дмитриев А., Лагунин А., Филимонов Д., Поройков В. (2015). SOMP: веб-сервер для in silico прогнозирования участков метаболизма для соединений, подобных лекарствам. Биоинформатика 31, 2046–2048. DOI: 10.1093 / биоинформатика / btv087
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рудик А.В., Беженцев В.М., Дмитриев А.В., Дружиловский Д.С., Лагунин А.А., Филимонов Д.А. и др. (2017). MetaTox: веб-приложение для прогнозирования структуры и токсичности метаболитов ксенобиотиков. J. Chem. Сообщить. Модель. 57, 638–642. DOI: 10.1021 / acs.jcim.6b00662
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ридберг, П., Глориам, Д. Э., Зарецки, Дж., Бренеман, К., и Олсен, Л. (2010). SMARTCyp: 2D-метод прогнозирования метаболизма лекарств, опосредованного цитохромом p450. ACS Med.Chem. Lett. 1, 96–100. DOI: 10,1021 / мл100016x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шихо, М., де Брюн Копс, К., Сторк, К., Свозил, Д., и Кирчмайр, Дж. (2017). FAME 2: простая и эффективная модель машинного обучения региоселективности цитохрома P450. J. Chem. Сообщить. Модель. 57, 1832–1846. DOI: 10.1021 / acs.jcim.7b00250
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Testa, B., Pedretti, A., and Vistoli, G.(2012). Реакции и ферменты метаболизма лекарств и других ксенобиотиков. Диск с наркотиками. Сегодня 17, 549–560. DOI: 10.1016 / j.drudis.2012.01.017
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тиан С., Джумбу-Феунанг Ю., Грейнер Р. и Вишарт Д. С. (2018). CypReact: программный инструмент для прогнозирования реагентов in silico для ферментов цитохрома P450 человека. J. Chem. Сообщить. Модель. 58, 1282–1291. DOI: 10.1021 / acs.jcim.8b00035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тайзак, Дж.Д., Хант П. А., Сегалл М. Д. (2016). Прогнозирование региоселективности и лабильности метаболизма цитохрома p450 с использованием квантово-механического моделирования. J. Chem. Сообщить. Модель. 56, 2180–2193. DOI: 10.1021 / acs.jcim.6b00233
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тайзак, Дж. Д., и Кирчмайр, Дж. (2018). Вычислительные методы и инструменты для прогнозирования метаболизма цитохрома P450 для открытия лекарств. Chem. Биол. Дизайн лекарств 93, 377–386.DOI: 10.1111 / cbdd.13445
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виллигхаген, Э. Л., Мэйфилд, Дж. У., Альварссон, Дж., Берг, А., Карлссон, Л., Джелязкова, Н., и др. (2017). The Chemistry Development Kit (CDK) v2.0: типирование атомов, описание, молекулярные формулы и поиск субструктур. J. Cheminform. 9:33. DOI: 10.1186 / s13321-017-0220-4
CrossRef Полный текст | Google Scholar
Wishart, D. S., Feunang, Y. D., Guo, A.К., Ло, Э. Дж., Марку, А., Грант, Дж. Р. и др. (2018). DrugBank 5.0: крупное обновление базы данных DrugBank за 2018 год. Nucleic Acids Res. 46, D1074 – D1082. DOI: 10.1093 / nar / gkx1037
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Зарецки Дж., Мэтлок М. и Свамидасс С. Дж. (2013). XenoSite: точное прогнозирование CYP-опосредованных участков метаболизма с помощью нейронных сетей. J. Chem. Сообщить. Модель. 53, 3373–3383. DOI: 10.1021 / ci400518g
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Общая схема клеточного метаболизма.Клетка является термодинамически …
Гузун, Р., Камбре, Т., Багур, Р., Гричин, А., Уссон, Ю., Варикмаа, М. Анманн, Т., Тепп, К., Тимохина, Н., Шевчук, И., Чекулаев, В., Буше, Ф., Дос Сантос, П., Шлаттнер, У., Валлиман, Т., Кузнецов, А.В., Дзея П., Алиев М., Сакс В. «Модульная организация сердечного энергетического метаболизма: преобразование энергии, передача и регулирование с обратной связью», Acta Physiol (Oxf), 2015, 213, 1, 84-106. http://www.ncbi.nlm.nih.gov/pubmed/24666671. Абстрактный. Чтобы удовлетворить высокие клеточные потребности, энергетический метаболизм сердечных мышц организован точным и скоординированным функционированием внутриклеточных энергетических единиц (ICEU).ICEU представляют собой структурные и функциональные модули, объединяющие множественные потоки в сайтах генерации АТФ в митохондриях и утилизации АТФ миофибриллярными, саркоплазматическим ретикулумом и АТФазами ионного насоса сарколеммы. Роль ICEU заключается в повышении эффективности векторного внутриклеточного переноса энергии и точной настройке окислительного синтеза АТФ, поддержании стабильных уровней метаболитов для адаптации к потребностям внутриклеточной энергии посредством динамической системы компартментализованных сетей переноса фосфорилов.Одним из ключевых элементов в регуляции распределения потока энергии и обратной связи является избирательная проницаемость внешней мембраны митохондрий (MOM), которая представляет собой узкое место в переносе и микрокомпартментализации аденин-нуклеотида и других энергетических метаболитов. Основываясь на экспериментальных и теоретических аргументах (математическое моделирование), мы описываем регуляцию митохондриального синтеза АТФ в ICEU, позволяя нагрузке на сердце линейно коррелировать с потреблением кислорода, обеспечивая условия метаболической стабильности, передачи сигналов и синхронизации.Особое внимание было уделено взаимосвязи структура-функция в развитии ICEU и роли взаимодействия митохондрий с белками цитоскелета, такими как тубулин, в регуляции проницаемости MOM в ответ на энергетические метаболические сигналы, обеспечивающие регуляцию митохондриального дыхания. Особое внимание было уделено важности метаболизма креатина для гомеостаза сердечной энергии.
Интеграция клеточного метаболизма в многомасштабную модель всего тела
Abstract
Клеточный метаболизм непрерывно превращает огромное количество внешних соединений в эндогенные метаболиты и является ключевым элементом физиологии человека.Многогранная физиологическая роль метаболической сети, выполняющей каталитические преобразования, может быть полностью понята только с точки зрения всего тела, когда одновременно рассматриваются причинно-следственные связи метаболических состояний отдельных клеток, окружающей ткани и всего организма. Здесь мы представляем подход, основанный на анализе динамического баланса потоков, который позволяет интегрировать метаболические сети на клеточном уровне в стандартизированные физиологически обоснованные фармакокинетические модели на уровне всего тела.Чтобы оценить наш подход, мы интегрировали сетевую реконструкцию гепатоцита человека в ткани печени физиологической фармакокинетической модели взрослого человека. Полученная многомасштабная модель была использована для исследования терапии гиперурикемии, детоксикации аммиаком и парацетамол-индуцированной токсичности на системном уровне. Конкретные модели одновременно объединяют несколько уровней биологической организации и предлагают механистическое понимание патологии и лечения. Представленный подход может в будущем поддержать понимание механизмов диагностики и разработки лекарств.
Сведения об авторе
Клеточный метаболизм — ключевой элемент физиологии человека. В идеале метаболическую сеть необходимо рассматривать в контексте окружающей ткани и организма, поскольку различные уровни биологической организации взаимно влияют друг на друга. Чтобы механистически описать взаимодействие между внутриклеточным пространством и внеклеточной средой, мы здесь интегрируем модель метаболической сети в масштабе генома HepatoNet1 на клеточном уровне в физиологически обоснованные фармакокинетические модели на уровне всего тела.Полученная многомасштабная модель позволяет количественно описать метаболическое поведение в контексте разрешенных во времени профилей концентрации метаболитов в организме и окружающей ткани печени. Модель была применена к трем тематическим исследованиям, охватывающим фундаментальные аспекты медицины и фармакологии: прием лекарств, идентификация биомаркеров и токсичность, вызванная лекарствами. В частности, наш многомасштабный подход способствует лучшему количественному пониманию действия лекарств и влияния метаболических нарушений на уровне организма на основе представления клеточного метаболизма в масштабе генома.Вычислительные модели, подобные представленной, включают различные аспекты физиологии человека и, следовательно, могут в значительной степени поддержать рациональные подходы в медицинской диагностике и разработке фармацевтических препаратов в будущем.
Образец цитирования: Krauss M, Schaller S, Borchers S, Findeisen R, Lippert J, Kuepfer L (2012) Интеграция клеточного метаболизма в многомасштабную модель всего тела. PLoS Comput Biol 8 (10): e1002750. https://doi.org/10.1371/journal.pcbi.1002750
Редактор: Джейсон А. Папин, Университет Вирджинии, Соединенные Штаты Америки
Поступила: 12.06.2012; Одобрена: 6 сентября 2012 г .; Опубликовано: 25 октября 2012 г.
Авторские права: © 2012 Krauss et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: MK, SS, JL и LK выражают признательность Федеральному министерству образования и исследований Германии за финансовую поддержку гранта № 0315747 (Виртуальная печень). MK, SS, JL и LK являются сотрудниками Bayer Technology Services GmbH, компании, разрабатывающей PK-Sim и MoBi. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: MK, SS, JL и LK являются сотрудниками Bayer Technology Services GmbH, компании, разрабатывающей PK-Sim® и MoBi®.
Введение
Метаболизм человека является неотъемлемой частью физиологии всего тела, и его дисфункция играет ключевую роль во многих системных заболеваниях. Частыми симптомами метаболических заболеваний являются изменения в экзометаболизме [1], [2], которые обычно следуют за изменениями распределения внутриклеточного потока [3]. Чтобы связать диагностические наблюдения на уровне организма, сопровождающие определенные заболевания, со структурными нарушениями на клеточном уровне, необходимо механистическое понимание корреляций генотип-фенотип [3], [4].Адекватные аналитические методы для системного рассмотрения лежащих в основе процессов все еще отсутствуют. Однако такие многомасштабные подходы необходимы для понимания очень сложной и взаимосвязанной структуры биологических сетей и взаимодействия с окружающим организмом [3], [5], [6].
В последние годы были разработаны подходы к моделированию, описывающие биологические процессы на разных уровнях физиологической организации, на основе множества расходящихся математических формализмов [5], [7], [8], [9].На уровне всего тела физиологически обоснованное фармакокинетическое (PBPK) моделирование количественно описывает абсорбцию, распределение, метаболизм и выведение (ADME) эндогенных и экзогенных соединений в организмах млекопитающих [10], [11], [12], [13] ]. В отличие от классического фармакокинетического (PK) / фармакодинамического (PD) моделирования [14], модели PBPK нацелены на механистическое представление процессов, связанных с ADME. Структурно модели PBPK состоят из компартментарных представлений всех соответствующих тканей и сосудистой системы.В частности, модели PBPK основаны на большом количестве предшествующей анатомической и физиологической информации, а также на общих моделях распределения, так что большинство параметров модели могут быть либо получены из коллекций баз данных, интегрированных в программное обеспечение для моделирования, либо они могут быть выведены из физико-химии соединение [15], [16], [17], [18], [19]. Следовательно, даже несмотря на то, что модели PBPK содержат более сотни обыкновенных дифференциальных уравнений и несколько сотен переменных, количество независимых параметров, которые необходимо корректировать во время разработки модели, невелико (обычно менее 10, см. Также Материалы и методы).Процессы, связанные с ADME, могут быть автоматически определены количественно на основе параметров, определяемых составом, что позволяет получить подробное представление о переносе массы в различных тканях. Модели PBPK ранее использовались для механистического анализа фармакокинетики лекарственных средств [20], фармакогеномики [21], экстраполяции видов [22] или анализа редких нежелательных явлений [23].
Для анализа на клеточном уровне реконструкции метаболической сети являются важным инструментом восходящей системной биологии.Клеточный метаболизм собирает множество вышестоящих регуляторных событий на различных уровнях клеточной организации, таких как транскриптом и метаболом, представляющих важную угловую точку в физиологии организма. Метаболические сети обычно описываются стехиометрическими матрицами, и распределения внутриклеточного потока являются неотъемлемыми переменными в таких моделях. Первые стехиометрические модели метаболизма человека в масштабе генома включали общие коллекции метаболической биохимии в человеческих клетках [24], [25].Недавние модели явно учитывают сетевую структуру в определенных тканях, что позволяет впервые рассматривать метаболические модели в конкретном контексте физиологии человека [4], [26].
Хотя модели метаболической сети применимы для исследования экспериментов in vitro, экспериментов с более или менее четко определенными условиями среды, их недостаточно для рассмотрения метаболизма in vivo , когда клетка встроена в постоянно меняющуюся среду. окружающих тканей и организма.Следовательно, метаболизм человека можно полностью понять только с помощью интегративного анализа, который одновременно учитывает контекст всего тела. Это позволяет, в частности, количественно оценить клеточные граничные условия и влияние на внутриклеточные состояния и процессы. Несколько подходов к объединению моделей, охватывающих разные уровни биологической организации, были описаны ранее [8], [27], [28]. Что касается метаболических сетей, анализ динамического баланса потоков (dFBA) был использован для соединения стехиометрических моделей метаболизма с динамическими моделями микробных пакетных культур и интегрированных омикс-сетей [29], [30], [31].
Здесь мы применяем dFBA для описания метаболических сетей человека в контексте моделей PBPK всего тела (рисунок S1 в тексте S1). Этот подход позволяет представить метаболизм человека при одновременном учете количественной доступности веществ на уровне организма (рис. 1 А). Мы в качестве примера используем HepatoNet1 [26], модель метаболизма в печени человека в масштабе генома, чтобы проанализировать специфические ответы сети перед лицом зависимых от времени профилей концентрации в ткани печени.Следуя этому подходу, мы исследуем три примера приложений (рисунок 1 B). Во-первых, мы используем многомасштабную модель PK / PD для анализа распределения и терапевтического эффекта аллопуринола при лечении гиперурикемии. Во втором примере мы рассматриваем влияние нарушения метаболизма аммиака на уровни в плазме крови, чтобы продемонстрировать способность методов идентифицировать биомаркеры, специфичные для патологических изменений метаболического состояния [25], [32], [33]. Наконец, мы применяем наш подход к анализу токсического воздействия парацетамола на функцию печени.
Рис. 1. Схема «галстук-бабочка», иллюстрирующая рассмотрение метаболических сетей в контексте всего тела.
( A ) Схематическое изображение многомасштабного подхода. Модели PBPK используются для количественного описания тканевой доступности экзогенных и эндогенных соединений на уровне организма. Модели PBPK связаны со стехиометрическими сетями с помощью обменных курсов, рассчитываемых с помощью dFBA. Можно моделировать как потребление, так и образование метаболитов, а также регулирующие эффекты.( B ) Возможные области применения, иллюстрирующие широкую применимость подхода: (i) многомасштабное моделирование PK / PD, (ii) количественная идентификация специфических для болезни или индивидуальных биомаркеров и (iii) анализ токсичности, вызванной лекарственными средствами.
https://doi.org/10.1371/journal.pcbi.1002750.g001
Результаты
Модель муфты
МоделиPBPK описывают процессы, лежащие в основе распределения соединения в организме, на основе предшествующей физиологической информации и общих моделей распределения.Органы в моделях PBPK обычно подразделяются на дополнительные компартменты, такие как сосудистое, интерстициальное и внутриклеточное пространство [12], [34]. Основные дифференциальные уравнения в этих разделах описывают поглощение, секрецию, образование и потребление конкретного соединения, представляя уравнения общего баланса массы [10]. Напротив, стехиометрические модели описывают механизмы внутри клетки в гораздо более тонком пространственном масштабе, обеспечивая более детальное понимание внутриклеточных процессов с особым акцентом на клеточной биохимии.Таким образом, интуитивно понятной точкой соприкосновения обоих формализмов моделей является внутриклеточное пространство, где модели PBPK количественно описывают профили концентрации во времени эндогенных или экзогенных соединений, которые, в свою очередь, представляют собой субстраты или продукты метаболических сетей на молекулярном уровне.
Чтобы связать распределение эндогенных и экзогенных соединений на уровне организма с метаболическими сетевыми структурами и, таким образом, с конкретным ферментативным процессом на клеточном уровне, стехиометрическая сеть была встроена в динамическую модель всего тела путем пошаговой дискретизации модели. .С этой целью можно предположить, что функциональная адаптация метаболизма, количественно определяемая распределением внутриклеточного потока и скоростью внеклеточного обмена, будет быстрой по отношению к окружающим процессам распределения в масштабе всего тела. Следовательно, распределения потоков остаются постоянными в каждом временном интервале [29], [30], [31]. В нашем случае выбранный временной интервал составлял 1 шаг / мин. Следовательно, для определенного распределения внеклеточных концентраций в данный момент времени внутриклеточное устойчивое состояние (т.е.е. равновесие), а анализ баланса потоков (FBA) может применяться для оценки распределений потоков [31].
Следуя логике проверки сети, используемой в HepatoNet1 [26], мы здесь применили цели для конкретных случаев, такие как максимизация производства аммиака или максимизация производства мочевой кислоты, для количественной оценки скорости внеклеточного обмена с учетом определенного набора граничных условий. Примечательно, что распределение внутриклеточного потока, имеющее биологическое значение, вряд ли может быть идентифицировано с использованием этих функциональных целей, поскольку они скорее оценивают макроскопическое поведение клетки.Напротив, нижележащее пространство потоков оценивается качественно. В нашем подходе соединение в модели PBPK может действовать либо как регуляторный модификатор, либо как субстрат ферментативной реакции в метаболической сети. Поэтому мы рассмотрели два различных способа связывания моделей PBPK и стехиометрических сетевых моделей: (1) непрямое связывание, когда концентрации соединения в модели PBPK оказывают регулирующее влияние на активность фермента, которая количественно оценивается на клеточном уровне («прямая связь» ), тем самым ограничивая потоки через эту специфическую реакцию и (2) прямое связывание, когда нарушенные метаболические процессы (например, ингибированные ферменты) итеративно влияют на доступность вещества в модели PBPK, напрямую влияя на соответствующий баланс массы (« обратная связь ») .В обоих случаях внутриклеточная концентрация соединения ограничивает метаболическое состояние в базовой сетевой структуре [35]. Это также влияет на последующие последующие события, поскольку катаболические или анаболические продукты, образующиеся во внутриклеточной метаболической сети, снова распределяются на уровне всего организма. Это централизованное рассмотрение метаболизма как ключевого компонента физиологии человека можно рассматривать как схему «песочные часы» или «галстук-бабочку» (рис. 1 A) [36], [37]. В частности, ферментативная блокада приводит к накоплению вышележащего субстрата, истощению нисходящего продукта и потенциальной активации альтернативных путей.Подробности косвенной и прямой связи будут объяснены ниже.
Непрямая связь.
Непрямое связывание используется для моделирования регуляторных модификаций, таких как лекарственное ингибирование метаболических ферментов (уравнение 1, 2). В этом случае ингибирование пораженного фермента не влияет на концентрацию действующего препарата. Таким образом, можно описать относительное изменение ферментативной активности и, таким образом, изменение внутриклеточного потока в метаболической сети как следствие разрешенных во времени внутриклеточных концентраций лекарственного средства в конкретном типе ткани.Законы скорости ингибирования, используемые для кинетики Михаэлиса-Ментен, используются для ограничения определенного внутриклеточного потока: (1) (2) Здесь концентрация лекарственного средства в определенный момент времени в модели PBPK соответствует концентрации ингибитора I (t ) . Относительная активность фермента ( relE (t) ) представляет собой разрешенное во времени соотношение скорости ингибированной реакции v (t) и скорости реакции без влияния v 0 ( I = 0 мкМ ). Затем relE (t) используется для количественной оценки эффекта ингибирования ферментов в метаболической сети.Дополнительное ограничение для этапа FBA, налагаемое на конкретный поток в метаболической сети, определяется как: (3) В уравнении 3 v E (t) соответствует потоку через затронутый фермент на каждом временном шаге, который равен определяется как произведение невозмущенного потока v E (0) , рассчитанного с помощью FBA, и относительной активности фермента relE (t) . Следовательно, набор ограничений изменяется со временем, так что скорости внеклеточного обмена, идентифицированные с помощью FBA, также динамически развиваются.Это позволяет, с одной стороны, количественно оценить влияние внеклеточных нарушений, вызванных соединением, на метаболические состояния. С другой стороны, обменные потоки с временным разрешением, определенные с помощью FBA, могут влиять на баланс масс модели PBPK, поэтому необходимо учитывать прямую связь (см. Раздел ниже).
Прямая муфта.
Метаболический клиренс и процессы образования моделируются путем прямого связывания, поскольку они напрямую влияют на баланс массы модели PBPK.Таким образом, прямое связывание описывает влияние активных процессов на клеточном уровне на распределение соединений в масштабе всего тела. С этой целью рассматривается цикл обновления обратной связи, который состоит из следующих этапов (рис. 2): (1) сначала рассчитываются клиренс и производительность по законам кинетической скорости модели PBPK. (2) Скорости впоследствии используются как верхние границы для FBA, чтобы (3) идентифицировать внутриклеточное распределение потока, количественно определяя скорости клеточного обмена. (4) Эти обменные курсы затем используются в качестве темпов очистки и добычи в модели PBPK, чтобы интегрировать следующий временной шаг.(5) Это приводит к новым уровням концентрации в конце временного шага, которые затем снова используются для расчета зазора и производительности для следующей итерации (1).
Рис. 2. Схематическое изображение петли обратной связи, используемой для прямого соединения.
Распределения потоков, рассчитанные FBA, используются для корректировки зазоров и дебитов в модели PBPK. После моделирования одного временного шага в модели PBPK, новые ставки очистки ограничивают следующий шаг FBA.
https: // doi.org / 10.1371 / journal.pcbi.1002750.g002
Хотя обменные курсы в модели PBPK и метаболической сети идентичны для эталонного метаболического состояния, эти скорости будут отличаться, если наложены внутренние и внешние возмущения в сети. Без какого-либо нарушения ферментных реакций в метаболической сети скорость очистки или продукции, рассчитанная с помощью FBA, достигнет верхнего предела (шаг 2). В этом случае показатели связанной модели идентичны показателям только модели PBPK. Напротив, скорости, рассчитанные с помощью FBA, будут уменьшаться, если, например, активность ферментов нарушена из-за ограниченной функциональности или структурных сетевых ошибок (уравнение 3).Следовательно, показатели связанной модели уменьшаются, и это впоследствии влияет на концентрации в масштабе всего тела (шаг 4).
Лечение аллопуринолом
В качестве первого примера мы проанализировали лекарственное действие аллопуринола при лечении гиперурикемии с помощью многомасштабной модели PK / PD. Метаболизм пуринов обеспечивает большое количество мишеней для лекарств [38], [39], причем мочевая кислота является конечным продуктом распада в организме человека. Таким образом, количественное моделирование эффекта лекарств, влияющих на этот важнейший метаболический путь, дает ценную информацию для разработки лекарств.
В клинической практике высокие уровни мочевой кислоты в плазме (выше 470 мкМ) называются гиперурикемией, которая может быть результатом врожденных ошибок метаболизма пуринов и даже чаще из-за нарушения выведения мочевой кислоты почками, что рассматривается ниже. Гиперурикемия может привести к таким заболеваниям, как подагра, когда мочевая кислота откладывается в тканях, особенно в суставах [40]. В настоящее время используется препарат против гиперурикемии — аналог пурина аллопуринол [41], который снижает выработку мочевой кислоты за счет ингибирования ксантиноксидазы.Этот фермент окисляет гипоксантин до ксантина, а затем ксантин до мочевой кислоты [42]. Сам аллопуринол окисляется ксантиноксидазой до оксипуринола, который также ингибирует ксантиноксидазу как активный метаболит (рис. S2 в тексте S1). Поскольку метаболизм аллопуринола до оксипуринола очень быстрый, а выведение оксипуринола очень медленное, оксипуринол играет значительную роль в ингибировании ксантиноксидазы [41].
Для оценки ингибирующей концентрации аллопуринола и его активного метаболита оксипуринола была разработана модель связанного PBPK (см. Материалы и методы).Экспериментально измеренные концентрации в плазме после перорального приема 200 мг аллопуринола [43] были рассмотрены для идентификации и точной настройки четырех физиологических параметров, описывающих абсорбцию и клиренс обоих соединений, а также четырех физико-химических параметров аллопуринола и оксипуринола (Идентификация параметров раздела в тексте S1, таблица S1 в тексте S1). Примечательно, что эти параметры распределены в двух независимых моделях PBPK, которые связаны реакцией клиренса.Например, был проведен анализ чувствительности для оценки влияния каждого параметра на модель (рисунок S3 в тексте S1). Полученная в результате комбинированная модель одновременно описывала PK аллопуринола и оксипуринола с превосходной точностью (рис. 3 A). Создав рабочую модель для однократного дозирования аллопуринола, на следующем этапе была предсказана PK при многократном введении (рис. 3 B). При моделировании профилей концентрации аллопуринола и оксипуринола в диапазоне времени 35 дней стало очевидно, что аллопуринол не накапливается в организме человека, а всегда полностью разлагается перед следующим применением.Напротив, оксипуринол больше не может быть полностью удален из организма, что приводит к средней концентрации оксипуринола около 45 мкМ.
Рисунок 3. Снижение выработки мочевой кислоты после многократного введения аллопуринола.
( A ) Моделируемые профили концентрации аллопуринола и оксипуринола в венозной плазме и внутри печени согласуются с экспериментальными данными PK [43]. ( B ) Прогнозирование профилей концентрации аллопуринола и оксипуринола в венозной плазме и внутри печени после многократного дозирования на основе модели однократного применения.( C ) Относительная ферментативная активность ксантиноксидазы (XO) после ингибирования однократной дозой аллопуринола. ( D ) Относительная ферментативная активность XO после ингибирования многократным введением аллопуринола. ( E ) Смоделированные профили концентрации мочевой кислоты в венозной плазме и внутри печени после однократного приема аллопуринола. ( F ) Смоделированные профили концентрации мочевой кислоты в венозной плазме и внутри печени после многократного введения аллопуринола.Показаны экспериментально измеренные концентрации венозной плазмы, определяющие гиперурикемическое состояние (*) и здоровое мочеиспускание (**) [48] после лечения аллопуринолом.
https://doi.org/10.1371/journal.pcbi.1002750.g003
На следующем этапе была создана модель PBPK для мочевой кислоты, которая позволила описать изменения концентрации мочевой кислоты как механистическое следствие ингибирования аллопуринола в печени. метаболизм пуринов (таблица S2 в тексте S1). Концентрация мочевой кислоты в плазме у здоровых мужчин составляет около 302 мкМ (мужчины: 302 ± 60 мкМ, женщины: 234 ± 52 мкМ), тогда как у пациентов с подагрой наблюдается гораздо более высокая концентрация, приблизительно 480 мкМ [44].Эта физиологическая информация была использована для определения трех параметров устойчивого состояния клиренса и скорости производства мочевой кислоты на модели всего тела здорового, а также подагрического индивидуума мужского пола (таблица S2 в тексте S1). Предполагалась одинаковая скорость продукции мочевой кислоты как у здоровых, так и у больных подагрой, поскольку нарушение почечной экскреции считается физиологической причиной гиперурикемии в данном случае.
Чтобы окончательно связать восходящее распределение аллопуринола и оксипуринола в масштабе всего тела с последующим ингибирующим эффектом на ксантиноксидазу в метаболической сети печени, было использовано непрямое связывание для моделирования эффекта PD как одной, так и нескольких доз аллопуринола ( Рисунок 3 C, D).Максимальное увеличение выработки мочевой кислоты считалось целевой функцией, и возникающая проблема FBA дополнительно ограничивалась ферментативным ингибированием, вызванным лекарственным средством, а также скоростью образования мочевой кислоты, оцененной в модели PBPK. Значения IC50 аллопуринола (13,4 мкМ [45]) и оксипуринола (15,6 мкМ [46]) были использованы для оценки констант Ki ингибирования фермента с помощью уравнения Ченга-Прусоффа (уравнения S1 – S6 в тексте S1 ) [47]. Взятые вместе, общая многомасштабная модель включала две связанные модели PBPK всего тела аллопуринола и оксипуринола, метаболизма в печени и последующую модель PBPK всего тела мочевой кислоты.
После успешного создания многомасштабной модели было смоделировано развитие уровня мочевой кислоты у пациентов с подагрой, отслеживающих терапевтический успех лечения подагры после однократного или многократного приема аллопуринола (рис. 3 E, F). Перед лечением аллопуринолом средний пациент с подагрой в устойчивом гиперурикемическом состоянии имел венозную плазменную концентрацию 476 мкМ. После однократного применения аллопуринола концентрация мочевой кислоты стала снижаться. Однако прерывание терапии или несоблюдение пациентом режима лечения привело к восстановлению концентрации мочевой кислоты, что типично для гиперурикемического состояния (рис. 3 E).Только при обычном приеме многократных доз аллопуринола непрерывное снижение концентрации мочевой кислоты могло быть достигнуто при концентрации в венозной крови 146 мкМ. В частности, прогнозируемая концентрация мочевой кислоты после нескольких дозировок была близка к наблюдаемому диапазону мочевой кислоты in vivo, (рис. 3 F) [48], [49], хотя модель PBPK была создана на основе однократного дозирования аллопуринола. Это правильное предсказание нашей модели ясно подчеркивает прогностические возможности связанной модели.
Детоксикация аммиака
В качестве второго примера мы проанализировали патогенез нарушений цикла мочевины на клеточном уровне и его влияние на концентрацию аммиака в плазме на уровне организма. В частности, мы стремились оценить диагностические маркеры конкретного состояния здоровья или прогрессирования заболевания. Печеночный метаболизм аминокислот и детоксикация аммиака играют важную роль в организме человека. До 95% аммиака, метаболизируемого в гепатоцитах, разлагается до мочевины, которая впоследствии выводится из организма, в то время как около 5% метаболизируются до глутамина и около 1% — до аланина [26].Нарушение метаболизма аммиака приводит к снижению выведения аммиака и тем самым вызывает гипераммониемию [50]. Следствием гипераммониемии является повышение концентрации аммиака в головном мозге — так называемая печеночная энцефалопатия, которая может вызвать спутанность сознания, летаргию, дезориентацию, а в тяжелых случаях — кому и смерть [51], [52]. Дисфункция печени при метаболизме аммиака может быть вызвана заболеваниями печени или врожденными нарушениями метаболизма (IEMs), например Нарушения цикла мочевины (UCD), которые могут иметь летальные последствия без адекватного лечения и диеты [53].Нарушения способности детоксикации аммиака вызывают прямые последующие изменения концентраций метаболитов в крови, что делает детоксикацию аммиаком основным примером для идентификации биомаркеров, специфичных для заболевания.
У человека аммиак образуется в результате расщепления аминокислот в печени или интенсивных мышечных упражнений [54]. Помимо эндогенного аммиака, экзогенный аммиак также попадает в организм с пищей. Всего в организме ежедневно вырабатывается около 17 г аммиака.Выведение через почки составляет около 13 г в день, в то время как 4 г в день метаболизируются в печени. Кроме того, во время нарушенной детоксикации аммиака после UCD скорость синтеза глутамина и аланина увеличивается в 4-6 раз [53], [55].
В качестве первого шага к исследованию нарушения детоксикации аммиака с использованием подхода многомасштабного сочетания была создана модель аммиака PBPK (Таблица S3 в тексте S1). Для определения скорости образования и потребления аммиака в рамках модели PBPK учитывалась равновесная концентрация 29 мкМ в венозной плазме, что является нормальным уровнем для здоровых людей [48].Три параметра модели, описывающие выработку и выделение аммиака, были идентифицированы с использованием вышеуказанной физиологической информации: была оценена общая скорость производства аммиака 0,694 мкмоль / л / мин, а также макроскопическая скорость клиренса печени и почек 0,163 мкмоль / л / мин и 0,530 мкмоль / л. / мин соответственно. Затем модель PBPK аммиака и метаболическая сеть были объединены с помощью прямого связывания. В частности, модель PBPK была смоделирована для одного временного шага для расчета новых концентраций и соответствующей скорости выведения из печени, которую затем можно было использовать в качестве новой верхней границы на следующем этапе FBA.Максимизация производства аммиака использовалась в качестве целевой функции, которая ограничивалась доступностью субстрата, скоростями обмена, рассчитанными с помощью модели PBPK, а также ферментативной недостаточностью, сопровождающей UCD. При постоянной концентрации аммиака 29,02 мкМ в венозной крови клетка печени показала внутриклеточную концентрацию аммиака 25,78 мкМ (рис. 4 A). В метаболической сети гепатоцита был рассчитан поток поглощения аммиака 0,163 мкмоль / л / мин, в то время как скорости продуцирования мочевины, глутамина и аланина были равны 0.070 мкмоль / л / мин, 0,008 мкмоль / л / мин и 0,002 мкмоль / л / мин, соответственно (рис. 4 B). Потребности в глюкозе и кислороде согласуются с предыдущими результатами [26].
Рисунок 4. Патогенез нарушения цикла мочевины по концентрации аммиака в плазме и скорости метаболического обмена.
( A ) Смоделированные профили концентрации аммиака в венозной плазме и внутри печени во время развития нарушения цикла мочевины. Черная пунктирная линия представляет собой общее снижение активности орнитин-транскарбамилазы.( B, C ): результирующие обменные потоки, рассчитанные с помощью FBA во время развития нарушения цикла мочевины. (B) Скорость притока в печень трех субстратов — аммиака, кислорода и глюкозы. (C) Скорость оттока трех продуктов — мочевины, аланина и глутамина.
https://doi.org/10.1371/journal.pcbi.1002750.g004
С помощью комбинированной многомасштабной модели мы смоделировали патогенез нарушения цикла мочевины (UCD), что привело к снижению способности детоксикации аммиака.Патогенез UCD в качестве примера предполагали как линейное снижение активности фермента орнитин-транскарбамилазы (OTC), ведущее к полному нарушению фермента (рис. 4 A) [56]. Одновременно скорость продукции глутамина и аланина увеличивается в четыре раза по сравнению с номинальной производительностью глутамина и аланина [57]. Следовательно, в то время как ферментативная активность ОТС снижается, максимальная активность ферментов, синтезирующих глутамин и аланин, увеличивается. Предполагается, что продукция глутамина и аланина увеличится через 6 часов после начала UCD, предполагая задержку из-за инициации транскрипции и трансляции.
С связанной моделью и описанными выше ограничениями, моделирование было выполнено во временном диапазоне 21 день. Через 24 часа начинает развиваться UCD, что привело к снижению производительности мочевины через 30 часов. В то же время потоки глутамина и аланина начали расти, а скорость поглощения аммиака стала снижаться. В новом устойчивом состоянии (через 6,5 дней) скорость поглощения аммиака составляла 0,072 мкмоль / л / мин (рис. 4 B), а скорости продуцирования глутамина и аланина составляли 0,033 мкмоль / л / мин и 0.007 мкмоль / л / мин соответственно (Рисунок 4 C). Концентрация аммиака в новой венозной крови составляла 33,99 мкМ, а концентрация в печени — 30,27 мкМ (рис. 4 A).
Вышеупомянутая модель представляет собой изображение среднего пациента. Поскольку модель, однако, структурно включает множество потенциальных причин межличностной изменчивости на большом уровне механистической детализации, ее также можно использовать для анализа патогенеза в масштабе популяции. Такие индивидуальные различия могут включать физиологию [58], экспрессию белка [20] или даже питание [59].Чтобы в качестве примера описать влияние индивидуальной вариабельности, 100 человек были смоделированы на основе случайно измененных показателей продуктивности и клиренса соответственно (при условии 10% стандартного нормального распределения относительно среднего пациента, рисунок 5 A, таблица S4 в тексте S1) . Распределение концентраций аммиака у здоровых и больных людей вместе с их совокупными суммами подчеркивает индивидуальную изменчивость во время патогенеза UCD (Рисунок 5 B). Выполнение теста Колмогорова-Смирнова показало, что распределения в здоровом и больном состоянии значительно различаются (p <0.001), что делает концентрацию аммиака количественным биомаркером безрецептурного дефицита. Путем моделирования популяции результаты демонстрируют, что простое рассмотрение отдельных лиц может вызвать вводящие в заблуждение диагнозы, когда необходимо исследовать определенные подгруппы пациентов, такие как люди с ожирением, пожилые или больные. Только популяционное моделирование всеобъемлющих механистических моделей может стать возможным механически различать различные, потенциально противоточные факторы, такие как высокая производительность аммиака и низкая скорость очистки.
Рис. 5. Определение влияния индивидуальной изменчивости на развитие нарушения цикла мочевины.
( A ) Смоделированные профили концентрации аммиака в венозной плазме у 100 человек во время развития нарушения цикла мочевины (отдельные профили и среднее значение). ( B ) Распределение концентраций аммиака, а также совокупные суммы у здоровых и больных людей значительно различаются (p <0,001).
https: // doi.org / 10.1371 / journal.pcbi.1002750.g005
Отравление парацетамолом
В нашем последнем примере рассматривается токсичность, вызванная лекарствами, путем устранения последующих эффектов дозирования лекарств на метаболические функции на клеточном уровне. Здесь мы выбрали передозировку парацетамола, одну из наиболее частых причин печеночной недостаточности [60] и отравлений [61]. Парацетамол обычно считается ингибитором синтеза простагландинов и широко используется для уменьшения боли (болеутоляющее) и лихорадки (жаропонижающее) [62].В более высоких дозах парацетамол может вызывать тяжелые гепатотоксические эффекты, приводящие к острой печеночной недостаточности и некрозу печени [63]. Парацетамол метаболизируется тремя основными путями: (i) глюкуронирование, (ii) сульфатирование и (iii) N-гидроксилирование цитохромом P450 2E1 (CYP2E1) [64]. Соответствующими метаболитами трех путей являются (i) глюкуронид парацетамола (PG), (ii) сульфат парацетамола (PS) и (iii) N-ацетил-п-бензохинонимин (NAPQI). В частности, NAPQI, продукт гидроксилирования, важен для рассмотрения токсичности парацетамола, поскольку он очень реактивен и токсичен [63].В терапевтических дозах NAPQI почти сразу детоксифицируется конъюгацией глутатиона (GSH). После реакции с GSH NAPQI далее разлагается в кишечнике и почках и выводится в виде парацетамола, цистеина (AC) и меркаптуровой кислоты [65]. Парацетамол метаболизируется первично в PG и PS. Однако при более высоких дозах пути синтеза PG и PS становятся насыщенными, вызывая выработку большего количества NAPQI [64]. В этом случае GSH почти полностью истощается (до 80%) за счет детоксикации NAPQI, так что избыток NAPQI накапливается в печени и организме.Затем свободный NAPQI ковалентно связывается с белками, образуя белковые аддукты, которые считаются одной из причин гепатотоксичности парацетамола [63]. В совокупности метаболическое воздействие передозировки парацетамолом описывается сниженной активностью трех ферментов N-10-тетрагидрофолатдегидрогеназы (THFDH, до 25%), глутаматдегидрогеназы (GDH, до 25%) и митохондриальной АТФ-синтетазы (ATPS). , до 60%), соответственно, и истощение GSH до 80% [66].
Для количественной оценки метаболического воздействия нарушений активности ферментов на функциональность всей печени было использовано непрямое связывание модели PBPK парацетамола и HepatoNet1 для определения влияния передозировки парацетамола на большое количество функциональных целей печени.HepatoNet1 ранее был проверен с 123 физиологическими функциями, которые представляют собой важные задачи для метаболизма печени [26]. В частности, 67 из 123 представленных целей были протестированы с тремя конкретными наборами внеклеточных метаболитов, которые поэтому использовались в качестве основного набора для проверки сети [26]. Чтобы количественно оценить влияние парацетамол-индуцированной печеночной недостаточности, мы проверили степень снижения значения каждой из 67 целевых функций во время передозировки парацетамолом, таким образом количественно оценив устойчивость печеночной сети к внешним воздействиям [67].
Мы начали наш анализ с разработки моделей PBPK исходного лекарственного средства парацетамола, трех метаболитов (таблица S5 в тексте S1). Первым был рассмотрен пероральный прием 1 г парацетамола. Восемь параметров модели были идентифицированы для модели парацетамола, по три для каждой модели PG и PS и четыре для модели NAPQI путем сравнения компьютерных симуляций с соответствующими экспериментально измеренными концентрациями в венозной крови для всех четырех соединений [65] (Идентификация параметров раздела в Текст S1, таблица S5 в тексте S1).Примечательно, что параметры либо характеризуют физико-химию соединений, либо описывают физиологию людей, так что неявно включены предшествующие знания (см. Материалы и методы). Более того, четыре модели сильно связаны между собой лежащими в основе балансами масс, что значительно снижает системную степень свободы. Поскольку метаболизм NAPQI в AC не может быть определен количественно из-за отсутствия литературной информации, предполагалось, что концентрация AC эквивалентна концентрации NAPQI.После настройки параметров моделирование парацетамола, PG, PS и NAPQI PBPK описало экспериментальные данные с превосходным соответствием (рис. 6 A, таблица S6 в тексте S1). Впоследствии была предсказана PK четырех соединений после летальной дозы парацетамола 15 г (фиг. 6B, таблица S7 в тексте S1).
Рис. 6. Влияние терапевтической дозы и токсической передозировки парацетамола на функциональность печени.
( A ) Смоделированные профили концентрации парацетамола и трех его метаболитов в венозной плазме после приема парацетамола в дозе 1 г согласуются с экспериментальными данными PK [65].( B ) Прогнозирование профилей концентрации парацетамола и трех его метаболитов в венозной плазме после дозы парацетамола 15 г на основе модели применения 1 г. ( C, D ) Относительная ферментативная активность трех нарушенных ферментов THFDH, GDH, ATPS и истощение GSH также реализуются как ингибирование фермента после применения 1 г (C) и 15 г (D) парацетамола, соответственно. ( E, F ) Влияние доз парацетамола 1 г (E) и 15 г (F), соответственно, на функциональность печени в трех различных временных точках.Полосы представляют собой уменьшение максимальных значений каждой целевой функции, которая претерпевает изменение своего максимального значения.
https://doi.org/10.1371/journal.pcbi.1002750.g006
Чтобы исследовать изменения метаболических функций после передозировки парацетамола, сначала была проведена FBA для определения оптимального значения каждой целевой функции и количественной оценки всех потоков. у здорового, нелеченного референтного индивидуума. Впоследствии ингибирование ферментов парацетамолом и его активными метаболитами было реализовано в качестве дополнительных ограничений на значения потока.Для каждого ингибированного фермента значение потока фиксировалось как произведение значения в здоровом состоянии и оставшейся относительной активности фермента (уравнение 6). Значения K i были рассчитаны исходя из предположения, что максимальная концентрация NAPQI после летальной дозы парацетамола в 15 г индуцирует максимальное ингибирование фермента, как описано выше. Для расчета значений K i для парацетамола и всех трех метаболитов, концентрации субстрата были приняты равными K m (уравнения S7 – S12 в тексте S1).Следовательно, разрешенные во времени относительные активности ферментов (рис. 6 C, D) могут быть рассчитаны следующим образом: (6) Затем для каждой целевой функции с этими дополнительными ограничениями был проведен FBA. Поскольку NAPQI в природе не продуцируется в печени, молекулярное взаимодействие экзогенного NAPQI с эндогенным GSH не может быть реализовано в HepatoNet1 без структурных модификаций сети. Следовательно, продукция NAPQI как экзогенного соединения моделируется в модели PBPK, как описано выше. Истощение GSH считалось довольно феноменологическим из-за непрямого связывания, тем самым вызывая снижение активности фермента, которое связано посредством непрямого связывания с внутриклеточной концентрацией NAPQI.Ни размер пула метаболитов GSH, ни регуляторные эффекты после истощения GSH нельзя механистически рассматривать в стехиометрических моделях, таких как HepatoNet1. Следовательно, все реакции, продуцирующие GSH (таблица S8 в тексте S1) в HepatoNet1, были идентифицированы и ингибированы, как описано выше, так что реакции потребления GSH ограничены. Изменение функциональности печени рассчитывали как разницу между максимальными значениями целевых функций в здоровом состоянии и в случае передозировки парацетамолом.Были рассмотрены три различных момента времени, которые включали время пиковых концентраций парацетамола (t max, парацетамол ) и NAPQI (t max, NAPQI ), соответственно, а также время минимальных концентраций через 24 часа после введения лекарства (t 24 ч ) (Рисунок 6 E, F). Из 67 рассмотренных целевых функций 20 целевых функций претерпели изменение после применения 1 г парацетамола по сравнению с необработанным состоянием, в то время как 24 целевые функции претерпели изменение после применения 15 г парацетамола.Почти все оптимальные значения были на уровне контрольных значений через 24 часа для дозы парацетамола 1 г, в то время как все затронутые цели оставались сильно сниженными через 24 часа для дозы 15 г.
Затем мы проанализировали, изменился ли поток, лежащий в основе различных оптимальных значений, во время применения разных доз парацетамола. Это могло бы объяснить, почему токсическая доза парацетамола влияет на большее количество объективных функций, чего следует ожидать с точки зрения устойчивости сети [67]. В качестве примера целевой функции, которая затрагивается только после токсической дозы, продукция олеата показала большую устойчивость к метаболическому эффекту парацетамола, поскольку многие активные потоки, лежащие в основе этой целевой функции, остались неизменными после применения 1 г парацетамола (54 .8%) (Рисунок 7 A). Точно так же максимальное значение цели было лишь немного уменьшено после применения 15 г парацетамола. Напротив, введение парацетамола показало больший эффект на максимальное значение продукции цистеина, поскольку максимальное значение уже уменьшилось после применения 1 г парацетамола по сравнению с необработанным контрольным состоянием. Более того, только 11,1% потоков, лежащих в основе этой целевой функции, остались неизменными после введения парацетамола, и многие новые потоки стали активными, предполагая, что изменение маршрута [68] использовалось для компенсации ингибирования нарушенного метаболического пути (Рисунок 7 B).
Рис. 7. Сравнение двух конкретных функций печени (выработка олеата и цистеина соответственно).
Сравниваются только потоки, отличные от нуля хотя бы в одном из трех случаев. Указывается количество потоков, которые остаются на своих исходных значениях (=), становятся меньше или выше (<,>) или отличны от нуля (новые) после нанесения 1 г и 15 г парацетамола. ( A ) Изменения флюсов после применения 1 г и 15 г парацетамола для производства олеата.( B ) Изменения потоков после применения 1 г и 15 г парацетамола для производства цистеина.
https://doi.org/10.1371/journal.pcbi.1002750.g007
Обсуждение
Здесь мы интегрируем метаболические сети в масштабе генома, включающие тысячи биохимических реакций, в модели PBPK для анализа метаболизма на уровне человеческого организма. Подход основан на dFBA [31], который используется для моделирования стационарных стехиометрических метаболических сетей на клеточном уровне в сочетании с моделями PBPK на основе ODE.Этот полунепрерывный подход использует скорости метаболического обмена в качестве входных данных на уровне организма и, наоборот, концентрации лекарств в масштабе всего тела для расчета верхних границ активности ферментов в метаболической сети.
Наш подход объединяет хорошо зарекомендовавшие себя подходы к компьютерному моделированию из различных биологических масштабов. В масштабе организма стандартизованные модели PBPK, которые обычно используются при разработке фармацевтических препаратов, обеспечивают обобщенное описание распределения веществ в организме человека.На клеточном уровне реконструкции метаболических сетей представляют собой основные строительные блоки восходящей системной биологии, которые описывают фундаментальную клеточную биохимию. Таким образом, наш подход к объединению обеспечивает общую основу для широкого диапазона возможных приложений. Примечательно, что ко всем этим приложениям можно обращаться без необходимости дальнейшего курирования или модификации модели.
Чтобы проиллюстрировать наш подход, мы в качестве примера интегрируем HepatoNet1 [26], модель метаболизма человеческого гепатоцита в масштабе генома, в ткань печени стандартизированной модели PBPK [11], [69].Таким образом, как и в действительности, процессы метаболизма на уровне организма являются результатом биохимической реакции внутри гепатоцита. Чтобы проиллюстрировать широкую применимость нашего подхода, представлены три тематических исследования, посвященных типичным медицинским и фармацевтическим вопросам.
В первом примере применения мы исследуем многомасштабный подход PK / PD, в котором косвенное связывание используется для механистического описания фармакологического эффекта пуринового аналога аллопуринола на биосинтез мочевой кислоты.ФК экзогенного лекарственного средства, а также его результирующий нижестоящий PD-эффект на образование эндогенных метаболитов оцениваются количественно, что позволяет провести всестороннюю оценку безопасности и эффективности лекарственного средства. При многократном пероральном приеме 200 мг аллопуринола мы прогнозируем снижение концентрации мочевой кислоты в венозной плазме на 69,3%, что количественно согласуется с клиническими данными [48]. Поскольку соответствующая модель PK / PD была создана на основе данных по однократным дозировкам аллопуринола [43], это точное предсказание долгосрочного терапевтического эффекта убедительно иллюстрирует прогностическую силу нашего подхода.
Второй пример применения — идентификация количественных биомаркеров метаболических нарушений. В дополнение к классической качественной идентификации биомаркеров [70], наш подход предоставляет количественную информацию с точки зрения конкретных профилей концентрации путем моделирования распределения затронутых соединений на уровне организма. Это позволяет механистически описать метаболические нарушения, такие как IEM в плазме крови и других биожидкостях. В качестве иллюстративного тематического исследования для количественной идентификации биомаркеров мы исследуем нарушение детоксикации аммиака, приводящее к увеличению содержания аммиака в плазме крови на 17%.Затем мы моделируем виртуальные популяции здоровых людей и пациентов, варьируя анатомические и физиологические параметры в соответствии с предыдущей статистической информацией [58]. Несмотря на значительный уровень индивидуальной вариабельности в обеих подгруппах, мы демонстрируем, что разница в уровнях аммиака в плазме между обеими подгруппами статистически значима. Примечательно, что структурная сложность моделей PBPK вместе с включенной предшествующей физиологической и анатомической информацией помогает объяснить противоречащее интуиции поведение во время прогрессирования заболевания у отдельных пациентов, поскольку многие соответствующие сопутствующие факторы механистически представлены в самой структуре модели.Это важно в клинической практике, поскольку в дополнение к диагностике основных метаболитов в модели структурно учитываются другие факторы, так что вариабельность нормального здорового состояния может быть механически определена количественно.
Токсикация, вызванная лекарственными средствами, вызванная передозировкой парацетамола, является третьим примером применения. Мы демонстрируем, как парацетамол снижает метаболическую способность, влияя на широкий спектр различных метаболических функций. Специфическое метаболическое воздействие проиллюстрировано для терапевтической (1 г) и токсической (15 г) доз парацетамола.Результаты показывают большее метаболическое влияние на отдельные функции печени после токсической дозы парацетамола. Кроме того, более высокая доза влияет на большее количество метаболических функций (24 функции против 20). Поскольку затронуто значительное количество потоков, мы заключаем, что отравление парацетамолом вызывает изменение маршрута потока, которое используется гепатоцитами для компенсации сетевых возмущений, тем самым обеспечивая устойчивость клеток. Примечательно, что этот эффект, который был показан ранее для микроорганизмов [68], распространяется по всей метаболической сети и может быть исследован только с помощью моделей в масштабе генома.
Описание воздействия соединения на организм и клетку является фундаментальным вопросом фармакодинамики. Классические подходы описывают эту интерференцию с помощью довольно феноменологических моделей [14]. Заменяя клеточное пространство моделями метаболической сети на уровне генома, наш подход описывает клеточные процессы на гораздо более высоком уровне детализации. Для количественной оценки конкретных метаболических состояний мы используем целевые функции, которые использовались ранее для проверки функциональной способности печени [26].Следует отметить, что такие функции не позволяют идентифицировать фактическое распределение внутриклеточного потока, в отличие от показанного ранее для микроорганизмов [71]. Напротив, пространство потока оценивается качественно перед лицом внешних или внутренних возмущений. Однако дополнительное ограничение оптимизации FBA с помощью кинетических скоростей, смоделированных с помощью моделей PBPK, обеспечивает важную передачу физиологической информации между обоими масштабами модели. Если бы было возможно количественно оценить распределение внутриклеточного потока в клетках млекопитающих, можно было бы использовать дополнительные алгоритмы [72] для дальнейшей оценки и характеристики общего пространства потока.Это может также включать рассмотрение дополнительной экспериментальной информации, такой как данные omics [73] или включение нормативной информации [74].
В совокупности представленный подход, объединяющий специфические для органов метаболические сети в модели PBPK, предоставляет множество возможностей для научных исследований, клинических применений и разработки лекарств, как показано на прототипных примерах, описанных выше. Только с помощью таких многомасштабных моделей будет достигнуто механистическое понимание дисфункции органов и этиологии заболевания на системном уровне.Это будет в значительной степени поддержано реконструкцией дальнейших метаболических сетей в масштабе генома, которые станут доступны в будущем [7]. Структурно метаболические сети обеспечивают шаблон, на котором может быть отображена генетическая предрасположенность пациента. Вместе с конкретной физиологической информацией это может когда-нибудь позволить на основе моделей оптимизировать профили риска и пользы в персонализированной медицине.
Материалы и методы
ПБПК моделирование
На уровне всего тела моделирование PBPK количественно описывает все связанные с ADME процессы эндогенных и экзогенных соединений в организмах млекопитающих [10], [11], [12], [13].В отличие от довольно описательного рассмотрения ПК в классических компартментных моделях, модели PBPK включают подробное представление физиологических процессов в организме, основанное на предварительных знаниях и информации. Базовая модельная структура, которая соединяет различные тканевые компартменты и сосудистую систему, основана на общих моделях распределения и количественно определяет массоперенос между различными субкомпартментами. Параметры моделей распределения автоматически выводятся из физико-химии соединений, таких как молекулярная масса или липофильность [15], [16], [17], [18], [19].Параметры, описывающие физиологию организма, такие как объем органов, скорость кровотока или состав ткани, получаются из коллекций физиологических данных и доступны во внутренней базе данных программного обеспечения PBPK. Из-за большого количества предварительной структурной и физиологической информации количество независимых параметров, которые необходимо идентифицировать во время разработки модели, невелико (обычно менее 10). Примечательно, что существует четкое разделение между физиологическими параметрами, которые относятся к организму, в котором распределено соединение, и конкретными параметрами соединения, которые конкретно описывают свойства самого вещества.Таким образом, параметры соединения необходимо настраивать только в узком диапазоне, поскольку уже имеется литературная информация. Напротив, физиологические параметры необходимо определять индивидуально.
Создание моделей PBPK с помощью PK-Sim и MoBi
Все модели PBPK, рассмотренные в этом исследовании, были построены с помощью программных инструментов PK-Sim и MoBi, для которых академические лицензии доступны бесплатно (раздел информация о программном обеспечении в тексте S1) и которые были подробно объяснены ранее [11], [ 69].Модели PBPK были созданы для исходных веществ, а также для всех метаболитов. Параметры, специфичные для соединения, использовались в каждом случае для параметризации базовой структуры модели PBPK. Во всех моделях мы рассматривали средних людей. Антропометрическая информация, касающаяся возраста, веса и роста, дополнительно определяет выбор физиологических параметров, как это предусмотрено в программном обеспечении. Это позволяет конкретную параметризацию модели PBPK, поскольку модель содержит только несколько независимых параметров, как описано выше.
Специфические для конкретного случая процессы метаболизма и клиренса, представленные кинетикой Михаэлиса-Ментен или реакциями первого порядка и согласно литературной информации, определяются дополнительно. Для соответствующих кинетических параметров настройка параметров важна, поскольку (1) обычно доступно меньше литературных данных и (2) эти параметры часто имеют значительную чувствительность, хотя общее динамическое поведение моделей PBPK является устойчивым (рисунок S3 в тексте S1).
Для примеров лечения аллопуринолом и отравления парацетамолом, модели PBPK в PK-Sim экспортируются в MoBi, где они дополнительно модифицируются.Модели PBPK исходного лекарственного средства и его метаболитов связаны таким образом, что распределение всех соединений описывается одновременно. В этом случае скорость реакции клиренса в модели исходного лекарственного средства устанавливается в качестве входных данных в модель метаболита.
Стехиометрические модели сети
Стехиометрические модели предоставляют математически формальный способ уловить основную биохимию клеточного метаболизма в аналитическую структуру. Предполагая установившееся состояние системы, все внутриклеточные метаболиты могут быть сбалансированы в линейных системах уравнений, которые обычно недоопределены, поскольку они охватывают гораздо больше неизвестных скоростей реакции, чем линейные независимые уравнения баланса массы.Распределения потоков можно идентифицировать со стехиометрическими моделями с помощью FBA [35], [75], [76] 🙁 4) Здесь f · v (,) соответствует целевой функции, которая отражает обоснование клеточной функции, S () обозначает стехиометрическую матрицу метаболической сети (включая m метаболитов и n реакции), а v представляет распределение внутриклеточного потока. Общее пространство для решения ограничено набором дополнительных ограничений ( A · v≤b ), которые представляют, например, доступность подложки.
HepatoNet1
Чтобы проиллюстрировать наш подход, мы здесь в качестве примера используем HepatoNet1, тканеспецифичную сетевую реконструкцию метаболизма печени [26]. HepatoNet1 состоит из 777 метаболитов и 2539 реакций и разделен на шесть внутри- и два внеклеточных компартмента. Его общая сетевая структура была подтверждена путем проверки выполнения 123 биохимических задач, представляющих метаболически возможные функциональные режимы. Следовательно, сеть была создана специально для изучения различных метаболических процессов в печени и обеспечивает структурную платформу для механистических исследований тканеспецифических физиологических функций [26].
Вклад авторов
Задумал и спроектировал эксперименты: MK SS JL LK. Проведены опыты: МК СС ЛК. Проанализированы данные: МК СС СБ РФ ЖЛ ЛК. Написал статью: МК СС СБ РФ ЖЛ ЛК.
Ссылки
- 1. Аллен Дж., Дэйви Х.М., Бродхерст Д., Хилд Дж. К., Роуленд Дж. Дж. И др. (2003) Высокопроизводительная классификация дрожжевых мутантов для функциональной геномики с использованием метаболического следа. Nat Biotechnol 21: 692–696.
- 2. Део Р.К., Хантер Л., Льюис Г.Д., Паре Г., Васан Р.С. и др.(2010) Интерпретация метаболомных профилей с использованием объективных моделей путей. PLoS Comput Biol 6: e1000692.
- 3. Lanpher B, Brunetti-Pierri N, Lee B (2006) Врожденные ошибки метаболизма: переход от менделевских к сложным заболеваниям. Нат Рев Генет 7: 449–460.
- 4. Джерби Л., Шломи Т., Руппин Э. (2010) Вычислительная реконструкция тканеспецифичных метаболических моделей: приложение к метаболизму печени человека. Мол сист Биол 6: 401.
- 5. Duarte NC, Becker SA, Jamshidi N, Thiele I., Mo ML, et al.(2007) Глобальная реконструкция метаболической сети человека на основе геномных и библиомических данных. Proc Natl Acad Sci U S A 104: 1777–1782.
- 6. Кюпфер Л. (2010) К физиологии систем всего тела. Мол сист Биол 6: 409.
- 7. Бордбар А., Фейст А., Усайт-Блэк Р., Вудкок Дж., Палссон Б. и др. (2011) Многотканевая метаболическая сеть в масштабе генома для анализа физиологии систем всего тела. BMC Syst Biol 5: 180
- 8. Hoehme S, Brulport M, Bauer A, Bedawy E, Schormann W. и др.(2010) Прогнозирование и проверка выравнивания клеток вдоль микрососудов как принцип порядка восстановления архитектуры ткани при регенерации печени. Proc Natl Acad Sci U S A 107: 10371–10376.
- 9. Kuepfer L, Lippert J, Eissing T (2012) Многомасштабное механистическое моделирование в фармацевтических исследованиях и разработках. Adv Exp Med Biol 736: 543–561.
- 10. Несторов И. (2007) Фармакокинетические модели, основанные на физиологии всего тела. Мнение эксперта Drug Metab Toxicol 3: 235–249.
- 11. Виллманн С., Липперт Дж., Севестре М., Солоденко Дж., Фойс Ф. и др. (2003) PK-Sim®: физиологически обоснованная фармакокинетическая модель «всего тела». Biosilico 1: 121–124.
- 12. Schmitt W, Willmann S (2004) Фармакокинетическое моделирование на основе физиологии: готово к использованию. Лекарственные открытия сегодня: Технологии 1: 449–456.
- 13. Rowland M, Peck C, Tucker G (2011) Фармакокинетика на основе физиологии в разработке лекарств и нормативной науке.Анну Рев Pharmacol Toxicol 51: 45–73.
- 14. Meibohm B, Derendorf H (1997) Основные концепции фармакокинетического / фармакодинамического (PK / PD) моделирования. Int J Clin Pharm Th 35: 401–413.
- 15. Роджерс Т., Лихи Д., Роуленд М. (2005) Фармакокинетическое моделирование на физиологической основе 1: прогнозирование тканевого распределения от умеренных до сильных оснований. J Pharm Sci 94: 1259–1276.
- 16. Роджерс Т., Роуленд М. (2006) Фармакокинетическое моделирование на физиологической основе 2: прогнозирование тканевого распределения кислот, очень слабых оснований, нейтралов и цвиттерионов.J Pharm Sci 95: 1238–1257.
- 17. Poulin P, Schoenlein K, Theil FP (2001) Прогнозирование жировой ткани: коэффициенты распределения плазмы для структурно не связанных лекарств. J Pharm Sci 90: 436–447.
- 18. Willmann S, Schmitt W, Keldenich J, Dressman JB (2003) Физиологическая модель для моделирования желудочно-кишечного кровотока и всасывания лекарств у крыс. Pharm Res 20: 1766–1771.
- 19. Willmann S, Schmitt W, Keldenich J, Lippert J, Dressman JB (2004) Физиологическая модель для оценки фракционной дозы, поглощенной людьми.J Med Chem 47: 4022-4031.
- 20. Meyer M, Schneckener S, Ludewig B, Kuepfer L, Lippert J (2012) Использование данных экспрессии для количественной оценки активных процессов в физиологически обоснованном фармакокинетическом моделировании. Drug Metab Dispos 40: 892–901.
- 21. Eissing T, Lippert J, Willmann S (2012) Фармакогеномика кодеина, морфина и морфин-6-глюкуронида: модельный анализ влияния активности CYP2D6, активности UGT2B7, почечной недостаточности и ингибирования CYP3A4.Мол Диаг Тер 16: 43–53.
- 22. Вебер О., Уиллманн С., Бишофф Х., Ли В., Вакалопулос А. и др. (2012) Прогнозирование потенциально эффективной дозы для человека BAY 60-5521, мощного ингибитора белка-переносчика эфиров холестерина (CETP), путем масштабирования аллометрических видов и комбинированного фармакодинамического и физиологически обоснованного фармакокинетического моделирования. Br J Clin Pharmacol 73: 219–231.
- 23. Willmann S, Edginton AN, Coboeken K, Ahr G, Lippert J (2009) Риск для новорожденного, находящегося на грудном вскармливании, от лечения кодеином для матери: исследование с количественным механистическим моделированием.Clin Pharmacol Ther 86: 634–643.
- 24. Duarte NC, Becker SA, Jamshidi N, Thiele I., Mo ML, et al. (2007) Глобальная реконструкция метаболической сети человека на основе геномных и библиомических данных. Proc Natl Acad Sci U S A 104: 1777–1782.
- 25. Ма Х, Горянин И. (2008) Реконструкция метаболической сети человека и ее влияние на открытие и разработку лекарств. Drug Discov Today 13: 402–408.
- 26. Гилле С., Беллинг С., Хоппе А., Булик С., Хоффманн С. и др.(2010) HepatoNet1: комплексная метаболическая реконструкция гепатоцита человека для анализа физиологии печени. Мол сист Биол 6: 411.
- 27. Гаммак Д., Гангули С., Марино С., Сеговия-Хуарес Дж, Киршнер Д.Е. (2005) Понимание иммунного ответа при туберкулезе с использованием различных математических моделей и биологических шкал. Мультимасштабная модель Sim 3: 312.
- 28. Hetherington J, Bogle IDL, Saffrey P, Margoninski O, Li L и др. (2007) Решение проблем управления многомасштабными моделями в системной биологии.Comput Chem Eng 31: 962–979.
- 29. Covert MW, Xiao N, Chen TJ, Karr JR (2008) Интеграция моделей метаболизма, регуляции транскрипции и передачи сигнала в Escherichia coli . Биоинформатика 24: 2044–2050.
- 30. Lee JM, Gianchandani EP, Eddy JA, Papin JA (2008) Динамический анализ интегрированных сигнальных, метаболических и регуляторных сетей. PLoS Comput Biol 4: e1000086.
- 31. Махадеван Р., Эдвардс Дж. С., Дойл Ф. Дж. 3-е (2002) Анализ динамического баланса потока диуксического роста в Escherichia coli .Biophys J 83: 1331–1340.
- 32. Шломи Т., Кабили М.Н., Руппин Э. (2009) Прогнозирование метаболических биомаркеров врожденных нарушений метаболизма человека. Мол сист Биол 5: 263.
- 33. Urbanczyk-Wochniak E, Luedemann A, Kopka J, Selbig J, Roessner-Tunali U, et al. (2003) Параллельный анализ транскриптов и метаболических профилей: новый подход в системной биологии. EMBO Rep 4: 989–993.
- 34. Герловски Л.Е., Джейн Р.К. (1983) Фармакокинетическое моделирование на физиологической основе: принципы и приложения.J Pharm Sci 72: 1103–1127.
- 35. Прайс Н.Д., Рид Дж. Л., Палссон Б. (2004) Модели микробных клеток в масштабе генома: оценка последствий ограничений. Nat Rev Microbiol 2: 886–897.
- 36. Csete M, Doyle J (2004) Галстуки-бабочки, метаболизм и болезни. Trends Biotechnol 22: 446–450.
- 37. Ма Х.В., Цзэн А.П. (2003) Структура связности, гигантский сильный компонент и центральная роль метаболических сетей. Биоинформатика 19: 1423–1430.
- 38.Ма Х, Сорокин А, Мазеин А, Сельков А, Сельков Э и др. (2007) Эдинбургская реконструкция метаболической сети человека и ее функциональный анализ. Мол Сист Биол 3: 135
- 39. Уильямс М., Джарвис М.Ф. (2000) Пуринергические и пиримидинэргические рецепторы как потенциальные мишени для лекарств. Biochem Pharmacol 59: 1173–1185.
- 40. Thews G, Mutschler E, Vaupel P (1999) Anatomie, Physiologie, Pathophysiologie des Menschen. Штутгарт: Wissenschaftliche Verlagsgesellschaft.868 с.
- 41. Пачер П. (2006) Терапевтические эффекты ингибиторов ксантиноксидазы: Возрождение через полвека после открытия аллопуринола. Pharmacol Rev 58: 87–114.
- 42. Харрисон Р. (2002) Структура и функция ксантин оксидоредуктазы: где мы сейчас находимся? Free Radic Biol Med 33: 774–797.
- 43. Turnheim K, Krivanek P, Oberbauer R (1999) Фармакокинетика и фармакодинамика аллопуринола у пожилых и молодых людей.Br J Clin Pharmacol 48: 501–509.
- 44. Беккер М.А. (1995) Гиперурикемия и подагра. В: Scriver CR, Beaudet AL, Sly WS, Valle D, редакторы. Метаболические и молекулярные основы наследственного заболевания. 7 изд. Нью-Йорк: Макгроу-Хилл. С. 1192.
- 45. Хуанг XF, Li HQ, Shi L, Xue JY, Ruan BF и др. (2008) Синтез аналогов ресвератрола и оценка их цитотоксической и ингибирующей ксантиноксидазы активности. Chem Biodivers 5: 636–642.
- 46. Fuchs P, Haefeli WE, Ledermann HR, Wenk M (1999) Ингибирование ксантиноксидазы аллопуринолом влияет на надежность метаболических соотношений кофеина в моче как маркеров активности N-ацетилтрансферазы 2 и CYP1A2.Eur J Clin Pharmacol 54: 869–876.
- 47. Cheng Y, Prusoff WH (1973) Связь между константой ингибирования (K1) и концентрацией ингибитора, которая вызывает 50-процентное ингибирование (I50) ферментативной реакции. Biochem Pharmacol 22: 3099–3108.
- 48. Лентнер С., редактор (1981) Научные таблицы Гейги. Вест Кэдвелл, Нью-Джерси
- 49. Wishart DS, Knox C, Guo AC, Eisner R, Young N, et al. (2009) HMDB: база знаний о метаболоме человека.Nucleic Acids Res 37: D603-610.
- 50. Kanamori K, Ross BD, Chung JC, Kuo EL (1996) Тяжесть гипераммонемической энцефалопатии коррелирует с уровнем аммиака в мозге и насыщением глутамин синтетазы in vivo. Журнал Neurochem 67: 1584–1594.
- 51. Онг Дж. П., Аггарвал А., Кригер Д., Исли К. А., Карафа М. Т. и др. (2003) Корреляция между уровнем аммиака и тяжестью печеночной энцефалопатии. Am J Med 114: 188–193.
- 52. Lockwood AH (2004) Уровни аммиака в крови и печеночная энцефалопатия.Metab Brain Dis 19: 345–349.
- 53. Клеппе С., Миан А., Ли Б. (2003) Расстройства цикла мочевины. Варианты лечения Curr Neurol 5: 309–319.
- 54. Eriksson LS, Broberg S, Bjorkman O, Wahren J (1985) Метаболизм аммиака во время физических упражнений у человека. Clin Physiol 5: 325–336.
- 55. Тофтенг Ф., Хауэрберг Дж., Хансен Б.А., Педерсен С.Б., Йоргенсен Л. и др. (2006) Стойкая артериальная гипераммонемия увеличивает концентрацию глутамина и аланина в головном мозге и коррелирует с внутричерепным давлением у пациентов с фульминантной печеночной недостаточностью.J Cereb Blood Flow Metab 26: 21–27.
- 56. Бен-Ари З., Далал А., Морри А., Питлик С., Зингер П. и др. (2010) Дефицит орнитин-транскарбамилазы (OTC) у взрослых, выявленный диетой Аткинса. J Hepatol 52: 292–295.
- 57. Маэстри NE, McGowan KD, Brusilow SW (1992) Концентрация глутамина в плазме: руководство по лечению нарушений цикла мочевины. J Pediatr 121: 259–261.
- 58. Виллманн С., Хон К., Эдгинтон А., Севестре М., Солоденко Дж. И др.(2007) Разработка модели популяции всего тела на основе физиологии для оценки влияния индивидуальной изменчивости на фармакокинетику лекарств. J Pharmacokinet Pharmacodyn 34: 401–431.
- 59. Willmann S, Thelen K, Becker C, Dressman JB, Lippert J (2010) Прогнозирование на основе механизма растворения и абсорбции в зависимости от размера частиц: фармакокинетика цилостазола у собак. Eur J Pharm Biopharm 76: 83–94.
- 60. Ларсон А.М., Полсон Дж., Фонтана Р.Дж., Дэверн Т.Дж., Лалани Э. и др.(2005) Острая печеночная недостаточность, вызванная ацетаминофеном: результаты многоцентрового проспективного исследования в США. Гепатология 42: 1364–1372.
- 61. Слышал KJ (2008) Ацетилцистеин при отравлении парацетамолом. N Engl J Med 359: 285–292.
- 62. Hinz B, Cheremina O, Brune K (2008) Ацетаминофен (парацетамол) является селективным ингибитором циклооксигеназы-2 у человека. FASEB J 22: 383–390.
- 63. Hinson JA, Roberts DW, James LP (2010) Механизмы некроза печени, индуцированного ацетаминофеном.Handb Exp Pharmacol 369-405.
- 64. Vermeulen NP, Bessems JG, Van de Straat R (1992) Молекулярные аспекты гепатотоксичности, вызванной парацетамолом, и ее профилактика на основе механизмов. Drug Metab Rev 24: 367–407.
- 65. Шинода С., Аояма Т., Аояма Ю., Томиока С., Мацумото Ю. и др. (2007) Фармакокинетика / фармакодинамика анальгезии парацетамолом у японских пациентов с хронической болью. Биол Фарм Булл 30: 157–161.
- 66. Hinson JA, Reid AB, McCullough SS, James LP (2004) Гепатотоксичность, вызванная ацетаминофеном: роль метаболической активации, активных форм кислорода / азота и перехода митохондриальной проницаемости.Drug Metab Rev 36: 805–822.
- 67. Вильгельм Т., Бехре Дж, Шустер С. (2004) Анализ структурной устойчивости метаболических сетей. Системы Биол 1: 114.
- 68. Blank LM, Kuepfer L, Sauer U (2005) Крупномасштабный анализ потока 13C раскрывает механистические принципы устойчивости метаболической сети к нулевым мутациям у дрожжей. Геном Биол 6: R49.
- 69. Эйссинг Т., Кюпфер Л., Беккер С., Блок М, Кобокен К. и др. (2011) Программная платформа вычислительной системной биологии для многомасштабного моделирования и симуляции: интеграция физиологии всего тела, биологии болезней и сетей молекулярных реакций.Фронт Физио 2.
- 70. Филлипс К.А., Ван Беббер С., Исса А.М. (2006) Диагностика и разработка биомаркеров: подготовка трубопровода. Nat Rev Drug Discov 5: 463–469.
- 71. Schuetz R, Kuepfer L, Sauer U (2007) Систематическая оценка целевых функций для прогнозирования внутриклеточных потоков в Escherichia coli . Мол Сист Биол 3.
- 72. Wiback SJ, Famili I, Greenberg HJ, Palsson BØ (2004) Выборка Монте-Карло может использоваться для определения размера и формы пространства стационарного потока.J Theor Biol 228: 437–447.
- 73. Shlomi T, Cabili MN, Herrgård MJ, Palsson BØ, Ruppin E (2008) Сетевое предсказание тканеспецифического метаболизма человека. Nat Biotechnol 26: 1003–1010.
- 74. Covert MW, Knight EM, Reed JL, Herrgard MJ, Palsson BO (2004) Интеграция высокопроизводительных и вычислительных данных проясняет бактериальные сети. Природа 429: 92–96.
- 75. Бланк Л.М., Кюпфер Л. (2010) Распределение метаболических потоков: генетическая информация, расчетные прогнозы и экспериментальная проверка.Appl Microbiol Biotechnol 86: 1243–1255.
- 76. Прайс Н.Д., Папин Дж.А., Шиллинг С.Х., Палссон Б.О. (2003) Микробные модели in silico в масштабе генома: подход, основанный на ограничениях. Trends Biotechnol 21: 162–169.
Метаболизм лекарств: ферментные механизмы и ингибирование
Многие потенциальные кандидаты в лекарства в конечном итоге не работают на практике, потому что они слишком эффективно метаболизируются по мере всасывания. Часто проблемой, вызывающей биотрансформацию, является реакция окислительного N-деалкилирования, катализируемая ферментом цитохромом P450.Несколько лет назад в качестве средства преодоления этого «эффекта первого прохождения» мы попытались разработать соединения, которые могли бы ингибировать вовлеченные P450. Мы обнаружили, что простые циклопропиламины, такие как N-бензилциклопропиламины (BCA), чрезвычайно эффективны в качестве ингибиторов P450, а позже показали, что они на самом деле являются субстратами самоубийства для ферментов P450.
Чтобы объяснить эту уникальную активность циклопропиламинов, мы предположили, что помимо удаления атомов водорода (схема 1) или передачи атомов кислорода своим субстратам, ферменты P450 могут также окислять определенные субстраты за счет переноса одного электрона (схема 2).
Добавление метильной группы (R) к циклопропановому кольцу BCA на схеме 1 блокирует N-деалкилирование, но не блокирует активность суицидного субстрата. Таким образом, схема 2 стала привлекательной для объяснения как N-делакилирования обычных аминов, так и суицидной субстратной активности, уникальной для циклопропиламинов. Со временем циклопропильная группа получила широкое распространение в качестве механического зонда для ферментов метаболизма амина и химических модельных систем, а схема 2 стала широко принятым механизмом метаболизма амина.
К сожалению, хотя схема 2 получила большую поддержку со стороны химических модельных систем, сопоставимых доказательств ее участия в ферментативном окислении не было получено. Поэтому несколько лет назад мы вернулись к этой проблеме. В частности, мы хотели получить окончательные доказательства судьбы циклопропильных углеродов, которые удаляются во время N-деалкилирования циклопропиламинов. Мы ожидали, что образование циклических продуктов по сравнению с продуктами с раскрытым кольцом будет различать механизмы HAT и SET.
Используя пероксидазу хрена (HRP) в качестве модельного фермента, окисляющего амин, и N-циклопропил-N-метиланилин (NCNMA) в качестве субстрата, мы продемонстрировали, что SET-окисление NCNMA приводит исключительно к раскрытию цикла циклопропильной группы, что и в данном случае путем внутримолекулярной реакции с фенильным фрагментом с образованием производного хинолина.
Однако, когда мы подвергли NCNMA окислению P450, не было не только раскрытия цикла, но и инактивации суицидных ферментов! Вместо этого единственными продуктами были формальдегид и гидрат циклопропанаона.Очевидно, что в этом случае P450 использует механизм HAT (схема 1).
Затем мы вернулись, чтобы более внимательно изучить взаимодействия BCA-P450, и получили еще один большой сюрприз. BCA окисляется до нитрозосоединения, которое чрезвычайно плотно связывает гемовое железо P450, тем самым инактивируя фермент! Однако количество образованного комплекса меньше количества инактивированного фермента, поэтому должны существовать дополнительные механизмы инактивации фермента. В настоящее время мы пытаемся определить, является ли Схема 2 одной из них.
Публикации
- М. А. Черни и Р. П. Ханзлик, «Катализируемое цитохромом P450 окисление N-бензил-N-циклопропиламина генерирует как гидрат циклопропанона, так и 3-гидроксипропионовый альдегид посредством отщепления водорода, а не переноса одного электрона». Варенье. Chem. Soc., 128, 3346-3354 (2006).
- M. A. Cerny и R. P. Hanzlik, «Циклопропиламиновая инактивация цитохромов P450. Роль метаболических промежуточных комплексов». Arch. Биохим. Биофиз., 436, 265-275 (2005).
- Р. П. Ханзлик, С. П. Гарриман, К. Л. Шаффер, Ю. М. Коэн, Р. А. Тота и М. А. Черни, «Окислительный метаболизм циклопропиламинов: судьба трех атомов углерода и другие интересные наблюдения». Синтез и применение изотопно меченых соединений, 8, 111-114 (2004).
- R. A. Totah и R. P. Hanzlik, «Неокислительное декарбоксилирование производных глицина пероксидазой». Варенье. Chem. Soc., 124, 10000-10001 (2002).
- К. Л.Шаффер, Яков М. Коэн и Р. П. Ханзлик, «Образование циклопропанона во время катализируемого цитохромом P450 N-деалкилирования циклопропиламина». J. Am Chem. Soc., 124, 8268-8274 (2002).
- Р. А. Тота и Р. П. Ханзлик, «Обнаружение промежуточных амино-иона: N-циклопропил против N-карбоксиметилгрупп в качестве репортеров». Варенье. Chem. Soc., 123, 10107-10108 (2001).
- С. Л. Шаффер, М. Д. Мортон и Р. П. Ханзлик, «Ферментативное N-деалкилирование N-циклопропиламина. Судьба циклопропильной группы.»J. Am Chem. Soc., 123, 8502-8508 (2001).
- С. Л. Шаффер, М. Д. Мортон и Р. П. Ханзлик, «Ферментативное N-деалкилирование N-циклопропиламина: необычная судьба циклопропильной группы». J. Am Chem. Soc., 123, 349-350 (2001).
- Х. Чен, М. деГрут, НПЕ Вермёлен и Р.П. Ханзлик, «Окислительное N-деалкилирование п-циклопропил-N, N-диметиланилина. Экстремальный эффект заместителя на реакцию радикальных часов, объясненный расчетами ab initio на радикальный катион» Промежуточные звенья.»J. Org. Chem., 62, 8227-8230 (1997).
Метаболизм встречается с эпигенетикой — курс и конференц-офис
Обзор
EMBL стремится делиться научными достижениями и поддерживать научное взаимодействие на протяжении всей пандемии коронавируса. Мы рады сообщить, что эта конференция станет виртуальной, и приглашаем вас присоединиться к нам онлайн.
Подача тезисов для устных презентаций больше невозможна.
Обзор симпозиума
Метаболизм и эпигенетика неразрывно связаны, играя ключевую роль в развитии, раке, иммунной передаче сигналов и старении.Этот симпозиум объединяет ведущих мировых исследователей и молодых ученых для изучения этой взаимосвязи с особым акцентом на том, как метаболиты и метаболические сети влияют на регуляцию генов, на недавно обнаруженную роль метаболитов в болезнях и как это открывает новые терапевтические возможности.
Мы выделим молекулярные и вычислительные инструменты, позволяющие обнаруживать метаболиты во времени и пространстве, проливая свет на метаболизм и связывая метаболизм с регуляцией структуры хроматина, активностью генов и эпигенетической наследственностью.Мы будем исследовать недавние открытия на границах метаболизма и эпигенетики — сосредоточив внимание на пересечении этих тем — достигнутые in vitro, на модельных организмах, в нечеловеческих условиях и в окружающей среде. Цель нашего междисциплинарного симпозиума — укрепить возникающую связь между метаболизмом и эпигенетикой, вдохновить на трансформационные идеи и помочь наладить сотрудничество на этом рубеже современной биологии.
Темы сеанса
- Метаболизм
- Эпигенетика
- Иммунометаболизм
- Инструменты и системы анализа
- Болезнь
- Хроматин
Что бывшие участники говорят о симпозиуме
« Я был рад открыть для себя новые подходы к метаболизму с эпигенетической и метаболомической точки зрения.Эта конференция повысила мое мнение о потенциале моих исследований. »Д-р Мануэль А. Фернандес-Рохо, руководитель группы регенеративной медицины печени, IMDEA-Food Institute, Испания
« Конференция собрала великих спикеров и многих молодых ученых из самых разных областей. Это вызвало много плодотворных дискуссий, помогло придумать новые идеи и позволило мне взглянуть на мой собственный докторский проект с другой точки зрения. ”Anna Nieborak, Helmholtz Zentrum München, Германия
« Выдающаяся конференция по актуальной теме.Я узнал от экспертов, как метаболические изменения и эпигеном влияют друг на друга. ”Ананд Сваруп, Национальный институт глаз, NIH, Бетесда, США
Динамики
Основные докладчики
Динамики
Асифа Ахтар
Институт иммунобиологии и эпигенетики Макса Планка, Германия
Андреас Бергталер
Исследовательский центр молекулярной медицины (CeMM) Австрийской академии наук, Австрия
Хунбо Чи
Санкт-ПетербургДетская исследовательская больница Джуда
США
Ян Мэйз
Медицинская школа Икана на горе Синай
США
Научные организаторы
Организаторы конференции
Инна Бессельманн
EMBL Heidelberg
Германия
Рейли Палл
EMBL Heidelberg
Германия
Программа
Есть что сказать? Твитнуть #EESMetabolism
- Все выступления будут транслироваться в прямом эфире, с сессиями вопросов и ответов в конце каждой сессии.
- Информация о прямой трансляции и доступ к дискуссионной платформе и цифровым постерам будет предоставлена незадолго до начала мероприятия.
- Доступ к записанным выступлениям будет доступен в течение 2 недель после начала мероприятия.
Все время в программе ниже показано как время в Европе (Берлин, Амстердам, Париж).
Чтобы узнать эквивалентный часовой пояс в вашем регионе, введите Берлин, время и дату программы, а также ваш город в Конвертер часовых поясов.
День 0 — вторник, 16 ноября 2021 г.| Время | Спикер |
| 15:00 — 16:00 | Веб-семинар перед симпозиумом от Dovetail Genomics: Удаление смещения фермента рестрикции ваши данные лигирования близости с MNase (см. аннотацию на вкладке вебинара перед симпозиумом) |
| Время | Спикер |
| 13:00 — 13:25 | Вступительное слово научных организаторов ДОСТУПНО ПО ЗАПРОСУ ПОТОК |
| 13:25 — 14:25 | Цикл Кребса и макрофаги Ключевое выступление: Люк О’Нил — Тринити-колледж Дублин, Ирландия ДОСТУПНО ПО СПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 911 14:25 — 17:50 | Сессия 1: Метаболизм Председатели: Ирен Мигель-Алиага, Теодор Александров |
| 14:30 — 14:55 | Половые различия в межорганных метаболических перекрестных помехах Ирен Мигель-Алиага — Имперский колледж в Лондоне, Великобритания В НАЛИЧИИ ПО ЗАПРОСУ ПОСЛЕ ПОТОКА |
| 14:55 — 15:20 | 91 155 Контроль гликолитического потока в формировании паттерна и времени эмбриона мыши|
| 15:20 — 15:45 | Mitochondlesria as signaling Navdeep Chandel — Северо-Западный университет, США ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА |
| 15:50 — 16:05 | Перерыв |
| 16:05 — 16:20 | Короткий разговор 1: Селективное по возрасту наследование органелл регулирует судьбу стволовых клеток посредством метаболизма Pekka Katajisto — University of Helsinki, Финляндия ДОСТУПНО ПО ЗАПРОСУ ПО ПОТОКУ ЖИВОГО ПОТОКА |
| 16:20 — 16:35 | Короткий разговор 2 : Метаболическое усиление паузы в развитии млекопитающих Вера ван дер Вейден — Институт Макса Планка Molecular Genetics, Германия ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА |
| 16:35 — 16:50 | Короткий разговор 3: Митохондриальное ADP-рибозилирование контролирует ядерное PARP1-индуцированное PARylation и эффективность ингибитора PARP Майкл Хоттигер — Цюрихский университет, Швейцария ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:50 — 17:15 | Комплексы, ассоциированные с ацетилтрансферазой MOF, связывают эпигенетику и метаболизм Asifa Akhtar Institute of Max. Иммунобиология и эпигенетика, Германия ТОЛЬКО LIVE STREAM |
| 17:15 — 17:20 | Transition |
| 17:20 — 17:50 | Панельная дискуссия 1 Председатели: Ирен Мигуэль -Алиага, Теодор Александров |
| 17:50 — 18:20 | Познакомьтесь с докладчиками |
| 18:20 — 18:50 | Встреча с редакторами — панельная дискуссия с Мария Полихрониду — EMBO Press Криста Бледсо — Молекулярная клетка Пунам Бхеда — Nature Communications Арунима Сингх — Nature Methods Christoph Schmitt — Nature Metabolism Mari-Carmen Fernandez-Aguera — Cell Metabolism Carolina Perdigoto — Nature Structural & Molecular Biology George Caputa — Nature Research |
| 18:50 — 19:20 | Социальная программа: быстрые сети |
| Конец дня 1 — Постоянный доступ к цифровым плакатам, сетевым и дискуссионным платформам |
| Время | Спикер |
|---|---|
| 12:30 — 12:50 | Отраслевая сессия |
| 13:00 — 15:50 | Сессия 2: Хроматин Председатели: Яд Гави-Хельм, Андреас Ладурнер |
| 13:05 — 13:30 | Разбор взаимосвязи между хроматином, метаболизмом и экспрессией генов Джейн Меллор — Оксфордский университет, Великобритания A ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 13:30 — 13:55 | Регулирование экспрессии генов в 3D во время эмбрионального развития Яд Гави-Хельм — Институт функциональной геномики Лиона, Франция ТОЛЬКО ПРЯМОЙ ПОТОК |
| 13:55 — 14:10 | Краткий доклад 4: Эволюция гистоновый вариант, участвующий в компартментальной регуляции метаболизма НАД Ива Губерович — Исследовательский институт лейкемии Хосепа Каррераса, Испания ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 14:15 — 14:30 | Перерыв |
| 14:30 — 14:45 | Краткий доклад 5: Swi-Snf и Cys на качелях серы: поддержание метаболического баланса и толерантности к окислительному стрессу посредством ремоделирования хроматина Майкл Черч — Институт медицинских исследований Стоуэрса, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 14:45 — 15:00 | Краткий доклад 6. Мутации IDH показывают влияние ингибирования PARP на репликацию гетерохроматина Juan Manuel Schvartzman — Memorial Sloan Kettering Cancer Center , США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПОТОКА В РЕЖИМЕ РЕАЛЬНОГО ВРЕМЕНИ |
| 15: 00-15: 15 | Краткий доклад 7: Связанные со старением изменения в метаболической зональности печени сопровождаются пространственными сдвигами в транскрипции и доступности хроматина Питер Тессарц — Институт биологии старения им. Макса Планка, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ LIVE STREAM |
| 15:20 — 15:50 | Панельная дискуссия 2 Председатели: Яд Гави-Хельм, Ян Мейз |
| 15:50 — 16:50 | Break and digital стендовая сессия Темы: Обмен веществ, хроматин и заболевания (живые чаты и видеозвонки с докладчиками) |
| 16:50 — 20:00 | Сессия 3: Анализ инструментов и систем Председатели: Янв Якуб Жилич, Эрика Пирс |
| 16:55 — 17:20 | Метаболизм как саморегулируемая система Джошуа Рабиновиц — Принстонский университет, США ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА |
| 17:20 — 17:45 | Использование ландшафтов динамических энхансеров для декодирования фенотипов макрофагов при здоровье и болезнях Кристофер Гласс — Калифорнийский университет, Сан-Диего, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 17:45 — 18:10 | Интеграция метаболизма с системной иммунологией в иммунитет и болезнь Hongbo Chi — St.Детский исследовательский госпиталь Джуд, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 18: 10-18: 35 | Пространственная одноклеточная метаболомика выявляет метаболические состояния клеток Теодор Александров — EMBL Heidelberg, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 18:40 — 18:55 | Перерыв |
| 18:55 — 19:10 | Короткий разговор 8: Анализ метаболома одного эмбриона и анализ транскриптома выявляет динамические события во время раннего развития дрозофилы Хайди Лемпрадл — Институт Ван Андел, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 19:10 — 19:25 | Краткий рассказ 9: Влияние истории питания в ювенильных стадиях на взрослой жизни дрозофилы Tadashi Uemura — Университет Киото, Япония ТОЛЬКО LIVE STREAM |
| 19:25 — 19:30 | Transition |
| 19:30 — 20:00 | Панельная дискуссия 3 Председатели: Ян Якуб ylicearz, Эрика |
| 20:00 — 20:30 | Познакомьтесь с докладчиками из сессий 2 и 3 |
| 20:30 — 21:00 | Социальная программа: микшер виртуального бара |
| Конец дня 2 — Постоянный доступ к цифровым плакатам, сетевым и дискуссионным платформам |
| Время | Спикер | ||
|---|---|---|---|
| 13:00 — 16:25 | Сессия 4: Иммунометаболизм Председатели: Андреас Бергталер, Мэтт Свит | ||
| : 05 — 13:30 | Лидия Линч — Гарвардская медицинская школа, США | ||
| 13:30 — 13:55 | Транскрипция полимеразы III и активность CK2 необходимы для прайминга Т-клеток дендритными клетками Philippe Pierre — Centre d’Immunologie Marseille-Luminy, Франция ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА | ||
| 13:55 — 14:20 | Гистоновые деацетилазы как молекулярные переключатели, контролирующие деление митохондрий и пути защиты хозяина в макрофагах Matt Sweet — Университет Квинсленда, Австралия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПРЯМОГО ПОТОКА | ||
| 14:20 — 14:45 911 58 | Изменение формы митохондрий в Т-клетках Эрика Пирс — Университет Джона Хопкинса, США ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА | ||
| 14:50 — 15:05 | Перерыв | 05 — 15:20 | Краткий доклад 10: Миелоидная трансглутаминаза-2 регулирует противогельминтный иммунитет путем регулирования метаболизма сукцината и аргинина Фиона Хенкель — Технический университет Мюнхена и Центр им. Гельмгольца, Мюнхен, Германия ДОСТУПНО ПО ЗАПРОСУ ПО ЗАПРОСУ STREAM |
| 15: 20- 15:35 | Краткий доклад 11: Неклассическая сигнальная ось ретиноевой кислоты регулирует идентичность гемопоэтических стволовых клеток Катарина Шёнбергер — Институт иммунобиологии и эпигенетики Макса Планка, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПРЯМОГО ПОТОКА | ||
| 15: 35- 15:50 | Shor Обсуждение 12: Исследование эпигенетической регуляции внутриопухолевой метаболической гетерогенности при тройном отрицательном раке груди Марина Фукано — Университет Монреаля, Университет Макгилла, Канада ТОЛЬКО LIVE STREAM | ||
| 15:50 — 15:55 | Transition | ||
| 15:55 — 16:25 | Панельная дискуссия 4 Председатели: Андреас Бергталер, Мэтт Свит | ||
| 16:25 — 17:25 | Перерыв и цифровой стендовая сессия Темы: Инструменты и системный анализ, Иммунометаболизм и эпигенетика (живые чаты и видеозвонки с докладчиками) | ||
| 17:25 — 20:30 | Сессия 5: Болезни Председатели: Анна Брюне, Филипп Пьер | ||
| 17:30 — 17:55 | Регулирование Т-клеточного иммунитета с помощью иммунометаболитов A ndreas Bergthaler — Исследовательский центр молекулярной медицины Австрийской академии наук, Австрия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 17:55 — 18:20 | Моноаминилирование белков в головном мозге: новый механизм нейронного развитие, пластичность и болезнь Ian Maze — Медицинская школа Icahn на горе Синай, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 18: 20- 18:35 | Короткий разговор 13: Визуализация метаболизма и эпигенетические переходы во время дифференцировки стволовых клеток при раке кишечника Мария Родригес Колман — Университетский медицинский центр Утрехта, Нидерланды ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 18: 40-18: 55 | Перерыв | ||
| 18:55 — 19:10 | Краткий разговор 14: Когда метаболизм пирувата встречается с эпитрансмией риптом: важность для синдрома Ли Laurent Le Cam — INSERM Montpellier, France ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 19:10 — 19:25 | Краткий разговор 15: Управление метиловой группой в процессе адаптации на митохондриальную болезнь Анастасия Мармылева — Университет Хельсинки, Финляндия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 19:25 — 19:40 | Короткий разговор 16: Метастазирующие клетки рака молочной железы (CSC) на ацетил-коа для колонизации легких Меган Янг — Швейцарский институт экспериментальных исследований рака, Федеральная политехническая школа Лозанны, Швейцария ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| 19: 40-40 | Краткий доклад 17: Ось пируват-лактат модулирует сердечную гипертрофию и сердечную недостаточность Ахмад Клунтун — Университет штата Юта, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВЫХ ПОТОКОВ | ||
| 19: 55-20:00 | Transition | ||
| 20:00 — 20:30 | Панельная дискуссия 5 Стулья: Анн Брюне, Филипп Пьер | ||
| 20:30 — 21:00 | Познакомьтесь с редакторами и спикерами — Сессия 4 и 5 в отдельных комнатах для обсуждения с | ||
| Конец день 3 — Постоянный доступ к цифровым плакатам, сетевым и дискуссионным платформам |
| Время | Спикер | |
|---|---|---|
| 13:00 — 17:20 | Сессия 6: Эпигенетика Председатели: Джейн Меллор, Андреас Ладурнер | 131163 |
| Лактат-опосредованное гистоновое лизиновое лактилирование: химия, протеом и биохимия Инмин Чжао — Чикагский университет, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | ||
| Транскрипционные, эпигенетические и метаболические регуляторные пути в первые дни развития Ян Якуб Жилич — Университет Копенгагена, Дания ТОЛЬКО ЖИВОЙ ПОТОК | ||
| 13:55 — 14:20 | Биохимическая основа регулируемой метаболитом НАД активации ремоделирования нуклеосом онкогеном ALC1 Андреас Ладурнер — Людв ig Университет Максимилиана в Мюнхене, Германия ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА | |
| 14:25 — 14:40 | Перерыв | |
| 14:40 — 14:55 | Короткий разговор 18: Обмен веществ -эпигенетическая регуляция вкусовых ощущений с помощью диеты Моника Дус — Мичиганский университет, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | |
| 14:55 — 15:10 | Короткий разговор 19: Межпоколенческий гормезис — это регулируется наследственным метилированием 18S рРНК Эрик Грир — Гарвардская медицинская школа, Бостонская детская больница, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | |
| 15:10 — 15:25 | Короткий разговор 20: Перепрограммирование ранних эмбрионов: первая встреча метаболизма и эпигенетики Джессика Испада — UFABC, Бразилия ДОСТУПНО ПО СПРОСУ ПОСЛЕ ЖИЗНИ S TREAM | |
| 15:25 — 15:40 | Краткий доклад 21: Нацеливание на метаболизм ацетил-КоА ослабляет формирование воспоминаний о страхе за счет снижения зависимого от активности ацетилирования гистонов Дези Александер — Университет Пенсильвании, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | |
| 15:40 — 16:40 | Эпигенетическая и метаболическая регуляция старения стволовых клеток Основной доклад: Anne Brunet -Stanford University School of Medicine, USA LIVE ТОЛЬКО ДЛЯ ПОТОКА | |
| 16:40 — 16:50 | Переход | |
| 16:50 — 17:20 | Панельная дискуссия 6 Председатели: Джейн Меллор, Андреас Ладурнер | |
| 17: 20 — 17:35 | Перерыв | |
| 17:35 — 18:05 | Познакомьтесь с докладчиками | |
| 18:05 — 18:35 | Заключительное слово ТОЛЬКО ДЛЯ ЖИВОГО ПОТОКА |
Практическая информация
Регистрационный взнос и подача тезисовРегистрационный взнос (включает доступ ко всем выступлениям, цифровым стендовым докладам и групповым обсуждениям в Интернете, а также помогает нам покрыть наши расходы на проведение мероприятия.Для получения дополнительной информации, пожалуйста, обратитесь к странице часто задаваемых вопросов):
| Академия | 190 евро |
| аспирант | 140 евро |
| Промышленность | 240 евро |