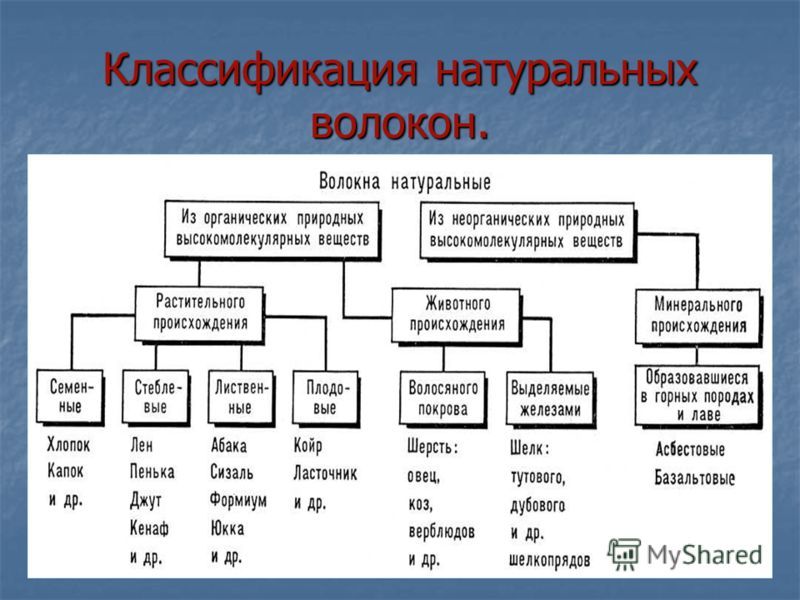
Виды волокон
Волокна являются основой всех современных тканей и материалов. Они различаются по химическому составу, свойствам, строению (структуре).
Классифицируются волокна по двум основным признакам:
- способу получения изделия;
- химическому составу.
С учетом основных классификационных признаков волокна делятся на химические и натуральные. Каждая группа обладает своими преимуществами, особенностями применения и эксплуатационными характеристиками.
Натуральные волокна
К этой категории волокон относятся материалы природного происхождения (минерального, животного, растительного). Самыми известными являются шелк, шерсть, лен и хлопок. Шерстяные и шелковые волокна имеют животное происхождение. Шерсть представляет собой волосяной покров верблюдов, коз, овец и т.д. Отличительной особенностью сырья является неоднородность (в состав входит 4 типа волокон: пух, переходный волос, ость, мертвый волос), высокая степень первоначального загрязнения.
Химические волокна
Из искусственных волокон наиболее известны следующие виды:
- вискоза, мягкая, яркая, гигроскопичная, глянцевая;
- полиамиды (энант, анид, капрон), прочные на растяжение и разрыв;
- полиэфиры (лавсан), обладающие высокой упругостью;
- полиакрилонитрилы (нитрон), мягкие, прочные, превосходящие по теплозащите натуральную шерсть;
- полиуретаны (спандекс) с разрывным удлинением до 800% и низкой гигроскопичностью.

Купить волокна, а также узнать стоимость и возможность доставки вы можете, позвонив по телефонам:
+7 (495) 505 — 6773,
+7 (495) 505 — 6772,
или заполнить форму заявки ниже.
Основные типы волокон мебельных тканей | Полезная информация
Состав тканей определяют текстильные волокна, из которых они состоят. Они могут быть натуральными и химическими.
Натуральные волокна бывают растительного и животного происхождения.
Хлопок — натуральное волокно растительного происхождения, используется в ткачестве и прядении. Хлопок применяется как в сочетании с другими волокнами, так и в самостоятельном прядении. Хлопок — прочный, мягкий, легко отстирывающийся материал (если его предварительно замочить). Однако под прямыми лучами солнца хлопок теряет прочность. Мебельная промышленность широко использует хлопок с добавлением синтетических нитей для упрочнения ткани. Соотношение натуральных и искусственных волокон разнообразно в различных тканях — от 90/10 до 10/90 процентов. Хлопок применяется в производстве обивочных тканей мебели. Его используют как в набивных тканях, так и в шенилах, микрофибре, флоках, велюрах. Он может быть как в составе ткани-основы, так и в составе ворса. Наличие хлопка в обивочных тканях — значительно повышает гигиенические показатели ткани (ткань обладает гигроскопичностью, пропускает воздух).
Хлопок применяется в производстве обивочных тканей мебели. Его используют как в набивных тканях, так и в шенилах, микрофибре, флоках, велюрах. Он может быть как в составе ткани-основы, так и в составе ворса. Наличие хлопка в обивочных тканях — значительно повышает гигиенические показатели ткани (ткань обладает гигроскопичностью, пропускает воздух).
Шерсть относится к классу натуральных волокон животного происхождения, применяется как для самостоятельного прядения и ткачества, так и в сочетании с другими волокнами. Подвержено усадке и уплотнению вплоть до превращения в фетр. Шерсть — идеальный материал для обивки, но, к сожалению, обивочная ткань с добавлением шерсти стоит очень дорого, поэтому редко встречается.
Химические волокна могут быть искусственными или синтетические.
Вискоза принадлежит к классу искусственного легкоокрашивамого и легкосминаемого волокна. Отлично смешивается с другими волокнами, способствует лучшему окрашиванию смесовых тканей.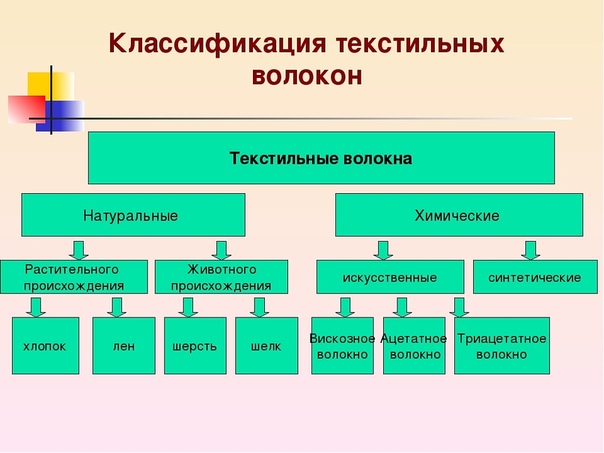 Ткани, в состав которых входит вискоза, огнеустойчивы, очень легко стираются, но при намокании теряют прочность. Вискоза используется в шенилах в смеси с синтетическими волокнами, во флоках — содержится в ткани-основе, в велюрах — в составе ворса.
Ткани, в состав которых входит вискоза, огнеустойчивы, очень легко стираются, но при намокании теряют прочность. Вискоза используется в шенилах в смеси с синтетическими волокнами, во флоках — содержится в ткани-основе, в велюрах — в составе ворса.
Ткани, состоящие из полиэфирных волокон (такие как полиэстер, лавсан и пр.) отличаются высокой прочностью и стойкостью к истираниям, не мнутся, не теряют цвет от солнечных лучей, не боятся органических растворителей. Однако крашению эти полотна поддаются с трудом, они электризуются и достаточно жесткие. Однако современные технологии устраняют эти недостатки химической модификацией. Сегодня полиэстер — распространенное волокно мебельной промышленности. Полиэстер применяется в производстве ворсовых тканей флоков, шенилов, жаккардов, велюров. Полиэстер усиливает устойчивость хлопчатобумажного волокна к воздействию на разрыв. Полиэстер не редко используется как самостоятельное волокно, без смешивания с натуральными волокнами. Современная технология обработки дошла до того уровня, что синтетическое волокно стало гигроскопичным.
Современная технология обработки дошла до того уровня, что синтетическое волокно стало гигроскопичным.
К полиамидным волокнам относятся нейлон, капрон и пр. Они обладают прочными, эластичными свойствами, устойчивы к истиранию и многократным изгибам, стойко переносят действие химических реагентов. Полиамиды практически не впитывают влагу. Данное волокно широко используют при изготовлении флоков. Полиамидные волокна имеют некоторые недостатки — повышенную электризуемость, невысокие термо- и светостойкость.
Мы перечислили основные типы волокон, используемые в производстве обивочных тканей, Однако, существуют еще несколько типов волокон, свойства которых позволяют добиться лучшего эффекта — полипропилен, акрил, спандекс.
Включенный в состав ткани полипропилен увеличивает ее водонепроницаемость, делает ее более легкой. Получаемый из производных нефти акрил часто смешивают с более дорогими волокнами для удешевления ткани. Лайкра или спандекс используются для увеличения драпируемости и несминаемости.
Для улучшения свойств мебельной ткани ее, как правило, производят из нескольких видов волокон. Синтетическая нить присутствует в большем или меньшем количестве во всех мебельных тканях. Она делает ткань более износоустойчивой, долговечной, прочной, придает устойчивость окраске. Современные мебельные ткани с синтетическими нитями обладают целым рядом достоинств: они «дышат», гипоаллергенны.
Типы оптических волокон от производителя
На заводе «Москабель-Фуджикура» по требованию заказчика возможно изготовление кабеля с любыми типами оптических волокон. Используется оптоволокно производства компании «Fujikura Ltd» (Япония) или от других производителей при необходимости.
Оптические волокна соответствуют рекомендациям Международного Союза Электросвязи Сектора телекоммуникационных стандартов ITU-T: G.651, ITU-T G.652.D (Е3), G.655, ITU-T G.657.A1 и другим международным стандартам.
| Типы оптических волокон в условных обозначениях оптического кабеля | Общая характеристика оптических волокон |
|---|---|
Е3 / G. 652.D+G.657.A1 652.D+G.657.A1 | Одномодовое оптическое волокно с низким пиком воды, оптимизированное на длине волны 1310, 1550, 1625 и 1383 нм по рекомендации ITU-T G.652.D с уменьшенным радиусом изгиба по рекомендации ITU-T G.657.A1 |
| Е3 / G.652.D | Одномодовое оптическое волокно с низким пиком воды, оптимизированное на длине волны 1310, 1550 и 1383 нм по рекомендации ITU-T G.652.D |
| Е5 / G.655 | Одномодовое оптическое волокно с ненулевой смещенной дисперсией, оптимизированное на длине волны 1550 и 1625 нм ITU-T G.655 |
| G.657.A1 | Одномодовое оптическое волокно с уменьшенным радиусом изгиба по рекомендации ITU-T G.657.A1 имеет дополнительные требования в части потерь на изгибе радиусом 15 мм |
| G.657.A2 | Одномодовое оптическое волокно с уменьшенным радиусом изгиба по рекомендации ITU-T G. 657.A2 имеет дополнительные требования в части потерь на изгибе радиусом 7,5 мм 657.A2 имеет дополнительные требования в части потерь на изгибе радиусом 7,5 мм |
| ОМ1/ М2 | Многомодовое оптическое волокно 62,5/125 с коэффициентом широкополосности на длине волны 850нм 200 МГц/км |
| ОМ2/ М1 | Многомодовое оптическое волокно 50/125 с коэффициентом широкополосности на длине волны 850нм 500 МГц/км по рекомендации ITU-T G.651.1 |
| ОМ3/ М3 | Многомодовое оптическое волокно 50/125 с коэффициентом широкополосности на длине волны 850нм 1500 МГц/км по рекомендации ITU-T G.651.1 |
| ОМ4/ М4 | Многомодовое оптическое волокно 50/125 с коэффициентом широкополосности на длине волны 850нм 3500 МГц/км по рекомендации ITU-T G.651.1 |
Выбирая оптическое волокно для узкоспециализированных задач необходимо проанализировать общие и специальные параметры кабеля, чтобы обеспечить наиболее полное соответствие эксплуатационных характеристик техническим требованиям.
Общие характеристики
Характерны для всех типов оптоволокон независимо от материала изготовления сердечника, оптической и защитных оболочек:
Число передаваемых световых волн (модов): одномодовый и многомодовый;
Параметры преломления световой волны (профиль): ступенчатый, градиентный, специальный;
Тип дисперсии: стандартная, со сдвигом нуля, с ненулевым смещением, с нулевым пиком воды.
Параметры многомодового оптоволокна
Кроме перечисленных выше общих характеристик многомодовое оптическое волокно (ОВ) делится на классы по материалу изготовления сердечника и оптической оболочки, которые влияют на их диаметры:
А1 – стекло / стекло, диаметр сердечника 50, 62, 5, 85, 100 мкм, диаметр оболочки 125, 125, 125, 140 мкм соответственно;
А2 – стекло / стекло, 200 / 240 мкм;
А3 – стекло / полимер, 200 / 280 мкм;
А4 – полимер / полимер, 980 / 1000 мкм.

Для коммуникационных сетей используются первых 2 типа, в особенности 50/125 и 62,5/125. Однако, учитывая большой коэффициент затухания сигнала в многомодовых кабелях, максимальная дальность прокладки не превышает 2-5 км. Кабели на основе пластиковых оптоволокон используются на абонентском уровне. Они имеют самые низкие характеристики передачи сигнала, но существенно проще в прокладке, к примеру имеют минимальный относительно диаметра радиус изгиба. При передаче сигнала через эти кабели допускается использование светодиодов, что существенно уменьшает диапазон пика воды – параметра отражения с максимальным затуханием светового импульса.
Параметры одномодового оптоволокна
Одномодовые кабели состоят из одного оптоволоконного сердечника, который изготавливается из кварцевого стекла, а у некоторых марок легируется различными материалами, существенно изменяя рабочие параметры конечного изделия. Диаметр оптической оболочки составляет 125 мкм при диаметре сердечника 7-9 мкм.
Одномодовые кабели осуществляет передачу только одного сигнала. В зависимости от материала легирования сердечника для передачи применяются электромагнитные импульсы с разной длиной волны, от видимого спектра до УФ. Дальность передачи может составлять до нескольких десятков до сотен километров соответственно.
Для подбора необходимого типа оптоволокна обращайтесь по телефону в шапке сайта, наши специалисты проведут консультацию и помогут оформить заявку.
Типы волоконно-оптических кабелей
Характеристики и типы оптического волокна
Является наиболее широко используемым одномодовым оптическим волокном в телекоммуникациях.
Одномодовое ступенчатое волокно с несмещенной дисперсией служит основополагающим компонентом оптической телекоммуникационной системы и классифицируется стандартом G.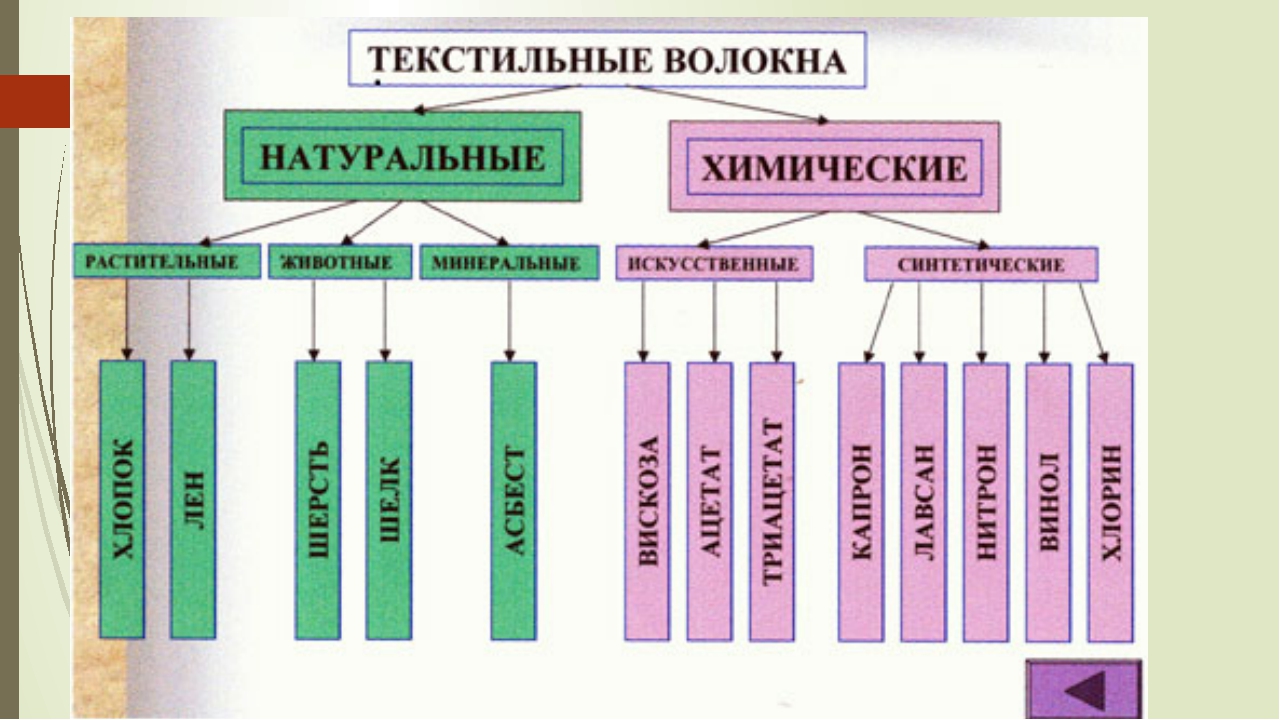 652. Наиболее распространенный вид волокна, оптимизированный для передачи сигнала на длине волны 1310 нм. Верхний предел длины волны L-диапазона составляет 1625 нм. Требования на макроизгиб — радиус оправки 30 мм.
652. Наиболее распространенный вид волокна, оптимизированный для передачи сигнала на длине волны 1310 нм. Верхний предел длины волны L-диапазона составляет 1625 нм. Требования на макроизгиб — радиус оправки 30 мм.
Стандарт разделяет волокна на четыре подкатегории A, B, C, D.
Волокно G.652.B соответствует требованиям, необходимым для передачи информационных потоков уровня до STM 64 в соответствии с Рекомендациями G.691 и G.692, и уровня STM 256, согласно G.691 и G.959.1.
Волокна G.652.C и G.652.D позволяют осуществлять передачу в расширенном диапазоне длин волн 1360-1530 нм и обладают пониженным затуханием на «пике воды» («пик воды» разделяет окна прозрачности в полосе пропускания одномодовых световодов в диапазонах 1300 нм и 1550 нм). В остальном аналогичны G. 652.A и G.652.B.
652.A и G.652.B.
G.652.A/B — эквивалент OS1 (классификация ISO/IEC 11801), G.652.C/D – эквивалент OS2.
Использование волокна — G.652 при более высоких скоростях передачи на расстояния более 40 км приводит к несоответствию эксплуатационных качеств со стандартами для одномодового волокна, требует усложнения оконечной аппаратуры.
G.655 — Одномодовое волокно с ненулевой смещенной дисперсией(NZDSF)
Одномодовое волокно с ненулевой смещенной дисперсией NZDSF оптимизировано для передачи не одной длины волны, а сразу нескольких длин волн (мультиплексного волнового сигнала WDM и высокоплотного волнового сигнала DWDM). Волокно защищено двойным акрилатным покрытием СРС, обеспечивающим высокую надежность и работоспособность. Наружный диаметр покрытия равен 245 мкм.
Волокно с ненулевой смещенной дисперсией (NZDSF) предназначено для применения в магистральных волоконно-оптических линиях и глобальных сетях связи, использующих DWDM-технологии. В этом волокне поддерживается ограниченный коэффициент хроматической дисперсии во всем оптическом диапазоне, используемом в волновом мультиплексировании (WDM).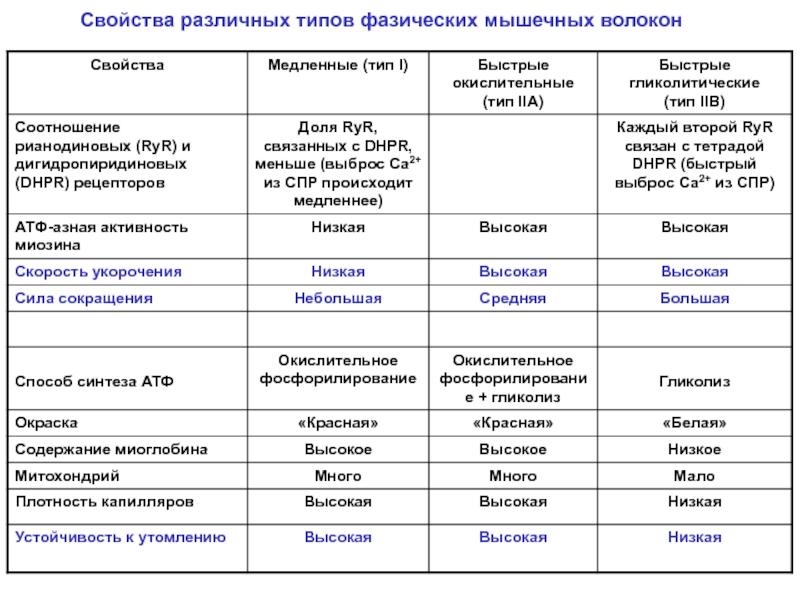 Волокна NZDSF оптимизированы для использования в диапазоне волн от 1530 нм до 1565 нм.
Волокна NZDSF оптимизированы для использования в диапазоне волн от 1530 нм до 1565 нм.
Рекомендации разделяют волокна на три подкатегории — А, В, С, которые различаются по значениям коэффициента поляризационной модовой дисперсии, хроматической дисперсии и рабочему диапазону.
Оптические волокна категории G.655.А обладают параметрами, обеспечивающими их применение в одноканальных и многоканальных системах с оптическими усилителями (Рекомендации G.691, G.692, G.693) и в оптических транспортных сетях (Рекомендация G.959.1). Рабочие длины волн и дисперсия в волокне данной подкатегории ограничивают мощность входного сигнала и их применение в многоканальных системах.
Оптические волокна категории G.655.B аналогичны G.655.А. Но в зависимости от рабочей длины волны и дисперсионных характеристик мощность входного сигнала может быть выше, чем для G.655.А. Требования в части поляризационной модовой дисперсии обеспечивают функционирование систем уровня STM-64 на расстоянии до 400 км.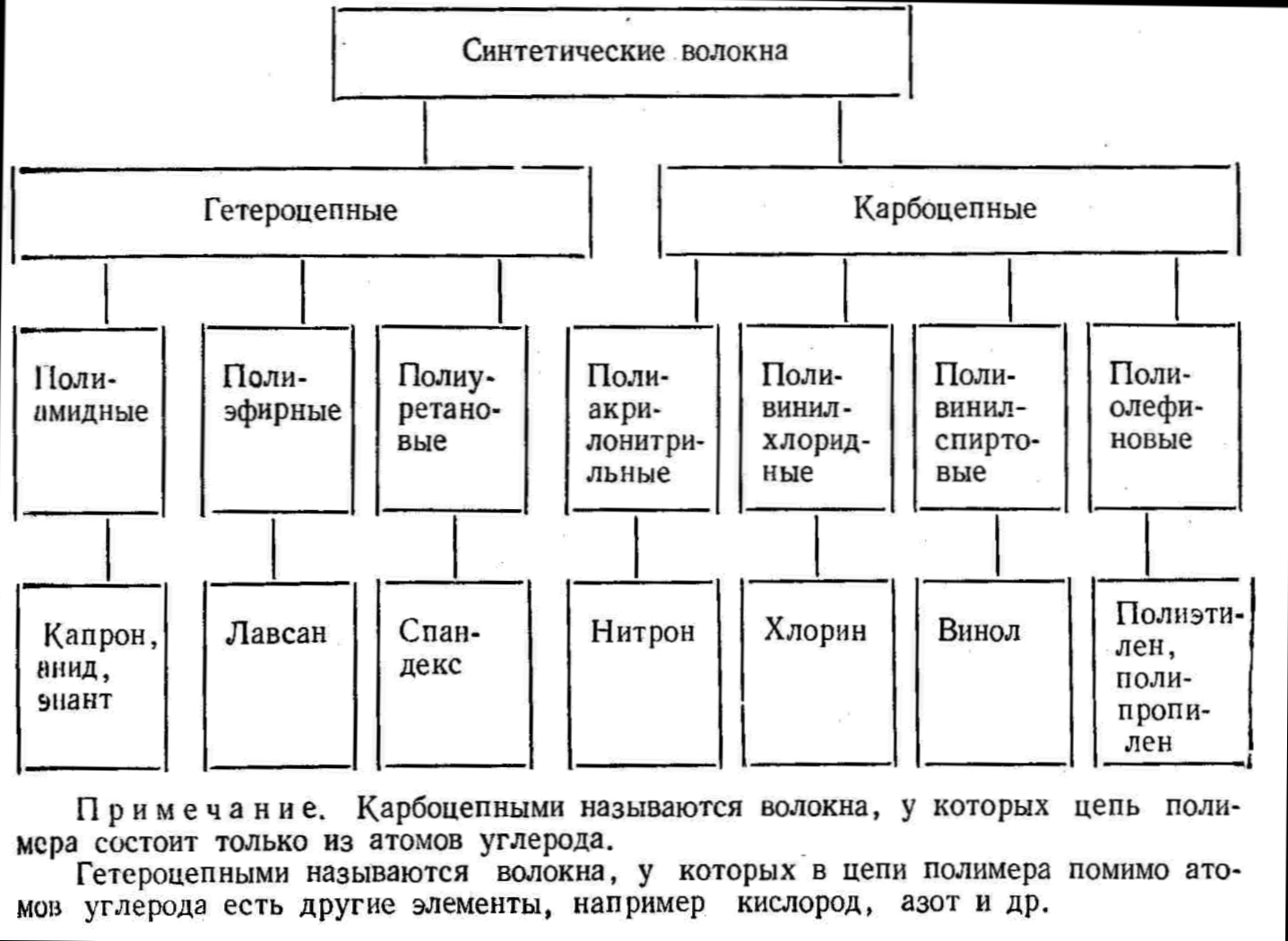
Категория волокон G.655.C подобна G.655.B, однако более строгие требования в части поляризационной модовой дисперсии позволяют использовать на данных оптических волокнах системы уровня STM-256 (Рекомендация G.959.1) или же увеличивать дальность передачи систем STM-64.
G.657 — Одномодовое волокно с уменьшенными потерями на изгибах с малыми радиусами
Оптическое волокно повышенной гибкости версии G.657 находит широкое применение в оптических кабелях для прокладки в сетях многоэтажных домов, офисов и т.д. Волокно G.657.A по своим оптическим характеристикам полностью идентично стандартному волокну G.652.D и в то же время имеет вдвое меньший допустимый радиус при укладке – 15 мм. Волокно G.657.В применяется на ограниченных расстояниях и обладает особо малыми потерями на изгибах.
Одномодовые оптические волокна характеризуются малым уровнем потерь на изгибах, предназначены в первую очередь для сетей FTTH многоквартирных зданий, а их преимущества особенно очевидны на ограниченном пространстве. Работать с волокном стандарта G.657, можно практически как с медножильным кабелем.
Работать с волокном стандарта G.657, можно практически как с медножильным кабелем.
Две подкатегории: A и B, которые различаются диаметром сердцевины и работоспособностью при изгибах.
Для волокон типа G.657.A он составляет от 8,6 до 9,5 мкм, а для волокон типа G.657.B — от 6,3 до 9,5 мкм.
Нормы потерь на макроизгибах существенно ужесточены, поскольку этот параметр для G.657 является определяющим:
• Десять витков волокна подкатегории G.657.A, намотанного на оправку радиусом 15 мм, не должны увеличивать затухание более чем на 0,25 дБ при длине волны 1550 нм. Один виток того же волокна, намотанного на оправку диаметром 10 мм, при условии, что остальные параметры не изменены, не должен увеличивать затухание более чем на 0,75 дБ.
• Десять витков подкатегории G.657.B на оправке диаметром 15 мм, не должны увеличивать затухание более чем на 0,03 дБ при длине волны 1550 нм. Один виток на оправке диаметром 10 мм — более чем на 0,1 дБ, один виток на оправке диаметром 7,5 мм — более чем на 0,5 дБ.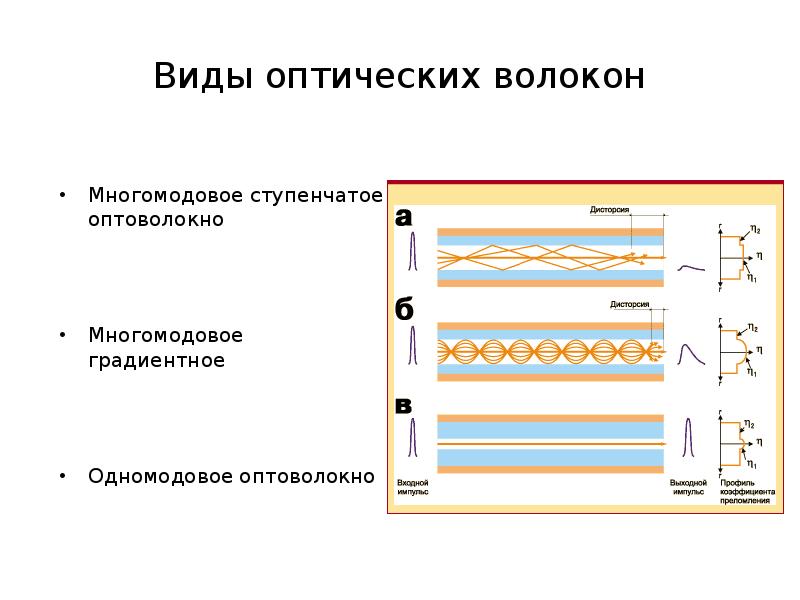
Рекомендация: ITU G.657.А определяет приоритет совместимости со стандартными волокнами по отношению к функциональности (с ITU-T G.652D).
Рекомендация: ITU-T G.657.В делает упор на нечувствительность к изгибам, а не на соответствие требованиям стандартов G.652.
Международной организацией по стандартизации (ISO) и Международной электротехнической комиссией (IEC) был опубликован стандарт ISO/IEC 11801 – «Информационные технологии — структурированные кабельные системы для помещений заказчика»
Стандарт задает структуру и требования к реализации универсальной кабельной сети, а также требования к производительности отдельных кабельных линий.
В стандарте для линий Gigabit Ethernet оптические каналы различаются по классам (аналогично категориям медных линий). OF300, OF500 и OF2000 поддерживают приложения оптического класса на расстояниях до 300, 500 и 2000 м.
|
Класс канала |
Затухание ММ-канала (дБ/Км) |
Затухание SM-канала (дБ/Км) |
||
|
850 нм |
1300 нм |
1310 нм |
1.550 нм |
|
|
OF300 |
2.55 |
1.95 |
1.80 |
1.80 |
|
OF500 |
3.25 |
2.25 |
2.00 |
2.00 |
|
OF2000 |
8.50 |
4.50 |
3.50 |
3.50 |
Кроме классов каналов, во втором издании этого стандарта определены три класса ММ-волокна — OM1, OM2 и OM3 — и один класс SM-волокна — OS1. Эти классы дифференцируются по затуханию и коэффициенту широкополосности.
|
Класс волокна |
Диаметр сердцевины, мкм |
Коэффициент широкополосности при насыщающем возбуждении, МГц х км |
Коэффициент широкополосности при лазерном возбуждении, МГц х км |
|
|
850 нм |
1.300 нм |
850 нм |
||
|
OM 1 |
50 или 62.5 |
200 |
500 |
N/A. |
|
OM 2 |
50 |
500 |
500 |
N/A. |
|
OM 3 |
50 |
1.500 |
500 |
2.000 |
Рекомендации по выбору типа волокна
Все линии короче 275 м могут работать по протоколу 1000Base-Sx. Длину до 550 м, можно обеспечить, используя протокол 1000Base-Lx совместно со смещенным вводом светового луча (Mode Conditioning).
|
Класс канала |
Fast Ethernet |
GigaBit Ethernet |
10 GigaBit Ethernet |
|
|
100 Base T |
1000 Base SX |
1000 Base LX |
10GBase-SR/SW |
|
|
OF300 |
OM1 |
OM2 |
OM1* , OM2* |
OM3 |
|
OF500 |
OM1 |
OM2 |
OM1 *, OM2 * |
OS1 (OS2) |
|
OF2000 |
OM1 |
— |
OM2 Plus, ОМЗ |
OS1 (OS2) |
*) Mode Conditioning
Многомодовое волокно класса OM4 характеризуется минимальным коэффициентом широкополосности 4700 МГц x км при длине волны 850 нм (по сравнению с 2000 МГц х км волокна типа OM3) и является результатом оптимизации характеристик волокна ОМ3, обеспечивающих возможность достижения скорости передачи данных 10 Гб/с на расстоянии 550 метров. Новый сетевой стандарт IEEE 802.3ab 40 и 100 Гигабит Ethernet отметил, что новый тип многомодового волокна ОМ4 позволяет передать 40 и 100 Гигабит Ethernet на расстоянии до 150 метров. Волокна класса OM4 планируется использовать в будущем с оборудованием 40Gbps и наиболее широко при оборудовании ЦОД.
OM 1 и OM2 – Стандартные многомодовые волокна с сердцевиной 62,5 и 50 микрон соответсвенно.
Кабели, патчкорды и пигтейлы с многомодовыми волокнами типов ОМ1 62,5/125мкм и ОМ2 50/125мкм уже давно применяются в СКС для обеспечения передачи данных с высокой скоростью и на относительно большие расстояния, которые требуется в магистралях. Наиболее важными функциональными параметрами ММ-волокна является затухание (attenuation) и коэффициент широкополосности (bandwidth). Оба параметра определяются для длин волн 850 нм и 1300 нм, на которых работает большая часть активного сетевого оборудования.
Является специально разработанным многомодовым оптическим волокном применяемое для сетей Gigabit и 10 Gigabit Ethernet, существует только с размером сердцевины 50 микрон.
OM4 – Оптическое многомодовое волокно с сердцевиной 50 микрон «лазер-оптимизированное» нового поколения.
Многомодовое волокно типа ОМ4 – в настоящее время полностью соответствует современным стандартам волокон, предусмотренных для центров обработки данных и групп серверов следующего поколения. Оптическое волокно ОМ4 может быть использовано для более протяжённых линий в сетях передачи данных нового поколения с высочайшей производительностью передачи данных. Это волокно представляет собой результат дальнейшей оптимизации характеристик волокна ОМ3, позволяющего придать волокну характеристики, обеспечивающие возможность достижения скорости передачи данных 10 Гб/с на расстоянии 550 метров. Волокна типа OM4 характеризуются повышенной эффективной минимальной модальной полосой пропускания 4700 МГц км при длине волны 850 нм (по сравнению с 2000 МГц км волокна типа OM3).
Группа продуктовЯзык: Валюта: МенюРекомендованная статья дБи — усиление изотропной антенны Бюллетень E-mail |
|
ЗАЖИМ ДЫМОХОДА OK-43T13 Нетто: 19.49 EUR ПОДВЕСНОЙ ШКАФ RACK EPRADO-R19-9U/450 Нетто: 106.10 EUR СКРЫТАЯ КАМЕРА AHD, HD-CVI, HD-TVI, CVBS APTI-H50YK-37 2Mpx / 5Mpx 3.7 mm APTI Нетто: 38.10 EUR AHD, HD-CVI, HD-TVI, PAL-КАМЕРА APTI-H50PV2-28W 2Mpx / 5Mpx 2.8 mm Нетто: 22.58 EUR IP-КАМЕРА IPC-HDW1431S-0280B-S4 — 4 Mpx 2.8 mm DAHUA Нетто: 108.58 EUR AHD, HD-CVI, HD-TVI, PAL-КАМЕРА APTI-H50PV2-28W 2Mpx / 5Mpx 2.8 mm Нетто: 22.58 EUR БЛОК ПИТАНИЯ POE POE-48/NX 24 W Нетто: 9.35 EUR АНТИВАНДАЛЬНАЯ КАМЕРАAHD, HD-CVI, HD-TVI, PAL APTI-H50V3-2812W 2Mpx / 5Mpx 2.8 … 12 mm Нетто: 46.40 EUR ВИДЕОТРАНСФОРМАТОР TR-1D*P2 Нетто: 2.23 EUR | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Виды синтетических волокон
Выделяют три основных вида синтетических волокон, которые используют для изготовления нитей для фильтровальных тканей:- полиэфирное волокно;
- полиамидное волокно;
- полипропиленовое.
Из полиэфирных нитей производят такие фильтровальные ткани:
- ткань полиэфирная артикул 86033,
- ткань ТЛФ-5 артикул 56190.
Из полиамидных нитей изготавливают ткань полиамидную артикул 56035, так же полиамид входит в состав ткани ТТФ-11 артикул 2409.
Широкое распространение получили полипропиленовые нити, из них ткут многие виды фильтровальных тканей, такие как:
- ткань фильтровальная КС-34;
- ткань кислостойкая КС-34;
- ткань полипропиленовая ТФП-92М артикул 56306.
Полиэфирные волокна — синтетические волокна, формуемые из расплава полиэтилентерефталата. Превосходят по термостойкости большинство натуральных и химических волокон: при 180°С они сохраняют прочность на 50%. Загораются полиэфирные волокна с трудом и гаснут после удаления источника огня; при контакте с искрой и электродугой не обугливаются.
Полиэфирные волокна сравнительно атмосферостойки. Они растворяются в фенолах, частично (с разрушением) — в концентрированной серной и азотной кислотах; полностью разрушаются при кипячении в концентрированных щелочах. Обработка паром при 100°С из-за частичного гидролиза полимера вызывает снижение прочности волокна (0,12% за 1 ч).
Полиэфирные волокна устойчивы к действию ацетона, четырёххлористого углерода, дихлорэтана и др. растворителей, микроорганизмов, моли, плесени, коврового жучка.
Устойчивость к истиранию и сопротивление многократным изгибам полиэфирных волокон ниже, чем у полиамидных волокон, а ударная прочность выше. Прочность при растяжении полиэфирных волокон выше, чем у других типов химических волокон.
Полиэфирное волокно обладает большим количеством достоинств, но так же имеет недостатки. Такие как трудность крашения обычными методами, сильная электризуемость, склонность к пиллингу, жёсткость изделий — во многом устраняются химической модификацией полиэтилентерефталата, например диметилизофталатом, диметиладипинатом (эти соединения вводят в реакционную смесь на стадии синтеза полиэтилентерефталата).
Техническая нить из полиэфирных волокон имеет широкую сферу использования. Ее применяют при изготовлении транспортёрных лент, приводных ремней, верёвок, канатов, парусов, рыболовных сетей и тралов, бензо- и нефтестойких шлангов, электроизоляционных и фильтровальных материалов, в качестве шинного корда. Полиэфирные волокна успешно применяют в медицине (синтетические кровеносные сосуды, хирургические нити). Из моноволокна делают сетки для бумагоделательных машин, щётки для хлопкоуборочных комбайнов, струны для ракеток и т.д. Текстильная нить идёт на изготовление трикотажа, тканей типа тафты, крепов и др. Методом «ложной крутки» получают высокообъёмную пряжу типа кримплен и мэлан. Штапельное полиэфирное волокно применяют в смеси с шерстью, хлопком или льном. Из таких смесей вырабатывают костюмные, пальтовые, сорочечные, плательные ткани, гардинно-тюлевые изделия и др. В чистом или смешанном виде полиэфирные волокна используют для производства искусственного меха, ковров. Войлок из полиэфирных волокон по важнейшим характеристикам превосходит войлок из натуральной шерсти. Торговые названия полиэфирных волокон: лавсан (СССР), терилен (Великобритания), дакрон (США), тетерон (Япония), элана (ПНР), тергаль (Франция), тесил (ЧССР) и др.
Полиамидное волокно
Полиамидное волокно — синтетические волокна, формуемые из расплавов или растворов полиамидов. Обычно для производства полиамидных волокон используют линейные алифатические полиамиды с молекулярной массой от 15 000 до 30 000 (чаще всего поликапроамид и полигексаметиленадипинамид).
С конца 60-х гг. 20 в. налажен выпуск полиамидных волокон из ароматических полиамидов, обладающих высокой термостойкостью. Технологический процесс получения полиамидных волокон включает три основных этапа: синтез полимера, формование волокна и его текстильную обработку. Полиамидные волокна характеризуются высокой прочностью при растяжении, отличной стойкостью к истиранию и ударным нагрузкам. Устойчивы к действию многих химических реагентов, хорошо противостоят биохимическим воздействиям, окрашиваются многими красителями. Максимальная рабочая температура волокон из алифатических полиамидов 80—150°С, волокон из ароматических полиамидов — 350—600°С.Полиамидные волокна растворяются в концентрированных минеральных кислотах, феноле, крезоле, трихлорэтане, хлороформе и др. Полиамидные волокна малогигроскопичны, что является причиной их повышенной электризуемости. Они плохо устойчивы к термоокислительным воздействиям и действию света, особенно ультрафиолетовых лучей. Для устранения этих недостатков в полиамиды вводят различные стабилизаторы.
Полиамидные волокна используются в производстве товаров широкого потребления, шинного корда, резинотехнических изделий, фильтровальных материалов, рыболовных сетей, щетины, канатов и др. Большое распространение получили текстурированные (высокообъёмные) нити из полиамидных волокон. Полиамидные волокна выпускают в виде непрерывных нитей или штапельных волокон во многих странах под следующими торговыми названиями: волокна из поликапролактама — капрон (СССР), найлон-6 (США), перлон (ФРГ), дедерон (ГДР), амилан (Япония) и др.: волокна из полигексаметиленадипинамида — анид (СССР), найлон-6,6 (США), родиа-найлон (ФРГ), ниплон (Япония) и др.; волокна из ароматических полиамидов — номекс (США).
Полипропиленовое волокно
Полипропиленовое волокно — синтетическое волокно, формуемое из расплава полипропилена. Полипропиленовое волокно по эластичности, устойчивости к двойным изгибам, как правило, превосходит полиамидные волокна, но уступает им по стойкости к истиранию. Обладает хорошими теплоизоляционными свойствами, имеет высокую стойкость к действию кислот, щелочей, органических растворителей.
Термо- и светостойкость полипропиленовых волокон сравнительно невысоки и в значительной мере определяются эффективностью вводимых в них стабилизаторов. Филаментное полипропиленовое волокно и моноволокно используют для изготовления нетонущих канатов, сетей, фильтровальных и обивочных материалов; штапельное полипропиленовое волокно — для выпуска ковров, одеял, тканей для верхней одежды, трикотажа, фильтровальных материалов. Текстурированное (высокообъёмное)полипропиленовое волокно находит применение главным образом в производстве ковров. Полипропиленовое волокно выпускается под различными торговыми названиями: геркулон (США), ульстрен (Великобритания), найден (Япония), мераклон (Италия) и др.
Цена:
Типы шерстяных волокон, руно и его элементы
Шерстяные волокна по внешним признакам делятся на следующие типы: пух, ость, переходный волос, мертвый, сухой, кроющий волос и песига (см. рис. 2).
Пух — самые тонкие волокна, наиболее извитые, образующие у грубошерстных овец нижний, более короткий ярус шерстного покрова (подшерсток). Тонина пуха от 15 до 30 мк. Шерстный покров тонкорунных овец состоит только из пуховых волокон.
Корни волокон пуха располагаются в коже пучками, потому что луковицы этих корней залегают гнездами. В коже тонкорунных овец встречается от 5 до 15 и более корней в одном пучке. Вследствие этого наблюдается слияние двух и более корней в общее волосяное влагалище с одним выходным отверстием. У пуха корни залегают в верхней части кожного слоя.
По внутреннему (гистологическому) строению пух овечьей шерсти состоит из чешуйчатого и коркового слоев. Чешуйчатый слой кольцевидный. Сердцевина у пуха овечьей шерсти отсутствует.
Пух разных видов шерсти различается по тонине, извитости, длине и гистологическому строению.
Пух является наиболее распространенным типом волокон, встречаясь во всех видах шерсти наряду с другими типами волокон или целиком образуя шерстный покров (у тонкорунных овец).
В технологическом отношении пуховые волокна являются наиболее ценными.
Ость — мало извитые, иногда почти прямые, толстые, грубые волокна. Тонина ости в отдельных случаях 30-35 мк, чаще от 40 до 80, встречаются волокна остевые до 120-140 мк. Ость обычно длиннее пуха, образует верхний ярус шерстного покрова. Корни остевых волокон располагаются в коже поодиночке, без всякой закономерности. Луковицы ости залегают глубже, чем у пуха, достигая границы с подкожной клетчаткой.
Ость состоит из трех слоев клеток: чешуйчатого, коркового и сердцевинного. Чешуйчатый слой некольцевидный. Сердцевинный слой бывает различным, обычно сердцевина увеличивается по мере утолщения остевых волокон. Встречается ость с прерывистой сердцевиной.
Остевые волокна в шерстном покрове находятся в смеси с пухом; не бывает шерсти, состоящей целиком из одной ости. Остевые волокна входят в шерстный покров полугрубошерстных и грубошерстных овец наряду с другими типами волокон.
Ость менее ценное в технологическом отношении волокно по сравнению с пухом. В различных видах шерсти ость различается как по техническим свойствам, так и по гистологическому строению.
Переходный (промежуточный) волос — волокно, среднее между остью и пухом, толще пуха, но тоньше ости. Тонина переходных волокон колеблется от 30 до 50-52 мк. Длина переходного волоса мало отличается от длинного пуха и средней ости. Поэтому их трудно иногда различить.
По гистологическому строению переходный волос ближе стоит к пуху, чем к ости. Сердцевина в нем чаще отсутствует или развита слабее, чем в ости, напоминая островки (прерывистая сердцевина).
Типичный переходный волос образует шерстный покров полутонкорунных овец (цигайской, куйбышевской, помесных тонкорунно-грубошерстных и других пород). В большинстве видов грубой шерсти переходный волос встречается обычно в меньшем количестве, чем ость и пух. Но в шерсти таких грубошерстных овец, как тушинская, балбас, сараджинская, переходного волоса содержится значительное количество. Особенно много переходного волоса в полугрубой помесной шерсти тонкорунно-грубошерстных овец.
По технологической ценности переходный волос стоит ближе к пуху.
Мертвый волос — очень грубое, ломкое, обычно короткое, почти прямое остевое волокно тониной до 160 мк.
По гистологическому строению мертвый волос близок к толстой ости. Чешуйчатый слой некольцевидный. Чешуйки мертвого волоса имеют вид узких пластинок, расположенных по 6-8 шт. по окружности волокна.
Мертвый волос имеет сильно развитую сердцевину. На поперечных срезах волокон мертвого волоса с трудом распознается очень тонкий корковый слой. Такое сильное развитие сердцевины служит одной из основных причин малой крепости его на разрыв. В силу пористого сердцевинного слоя мертвый волос практически не поддается окрашиванию. Мертвый волос в шерстяных изделиях плохо удерживается, быстро разрушается, выпадает из пряжи и ткани, вследствие этого сильно понижается качество изделия.
Сухой волос — это грубая ость, отличающаяся большой жесткостью и сухостью наружных концов, меньшим блеском, хрупкостью в верхней части волоса. Чем больше разница в косицах шерсти между длиной ости и пуха, тем больше в такой шерсти сухого волоса. Тонина волокна — от 50 до 140 мк.
По гистологическому строению сухой волос занимает промежуточное положение между остью и мертвым волосом.
В верхней части сухой волос напоминает мертвый, а в средней и нижней части, находящейся вместе с переходными и пуховыми волокнами, — ость.
Сухой волос встречается в шерсти большинства грубошерстных пород овец, являясь спутником мертвого полоса (у курдючных овец), а у отдельных пород овец большое количество сухого волоса не сопровождается мертвым (волошские, черкасские).
В технологическом отношении сухой волос приближается к ости.
Кроющий волос — прямой, жесткий, с сильным Плеском, короткий, длиной обычно 3-5 см. По строению приближается к ости. Корни кроющего волоса расположены наклонно, поэтому один волос прикрывает другой. Встречается кроющий волос на голове, конечностях и хвосте животного. На туловище животного кроющий волос из-за наклонного расположения не дает возможности остригать его, и поэтому он не входит в шерстную продукцию, получаемую с овец.
Песига (собачий волос) — шерстяные волокна, встречающиеся у тонкорунных и у некоторых полутонкорунных ягнят и выделяющиеся среди других волокон большой длиной, толщиной и меньшей извитостью. Некоторые ягнята рождаются с такими волокнами, а в дальнейшем в течение первого года жизни песига заменяется типичными пуховыми шерстяными волокнами. Встречается поэтому песига в тонком и полутонком поярке (шерсти с ягнят) и в шерсти с молодняка (годичного возраста) тонкорунных овец.
волокна | Источник питания
Клетчатка — это вид углеводов, который организм не может переваривать. Хотя большинство углеводов расщепляется на молекулы сахара, клетчатка не может быть расщеплена на молекулы сахара, и вместо этого она проходит через организм непереваренной. Клетчатка помогает регулировать потребление сахара организмом, помогая контролировать чувство голода и уровень сахара в крови.
Детям и взрослым для хорошего здоровья требуется не менее 20–30 граммов клетчатки в день, но большинство американцев получают только около 15 граммов в день.Отличные источники — цельные фрукты и овощи, цельнозерновые продукты и бобы.
Клетчатка бывает двух видов, каждая из которых полезна для здоровья:
- Растворимая клетчатка, растворяющаяся в воде, может помочь снизить уровень глюкозы, а также снизить уровень холестерина в крови. Продукты, содержащие растворимую клетчатку, включают овсянку, орехи, бобы, чечевицу, яблоки и чернику.
- Нерастворимая клетчатка, которая не растворяется в воде, может помочь пище проходить через пищеварительную систему, обеспечивая регулярность и предотвращая запоры.Продукты с нерастворимой клетчаткой включают пшеницу, цельнозерновой хлеб, цельнозерновой кускус, коричневый рис, бобовые, морковь, огурцы и помидоры.
Лучшими источниками клетчатки являются цельнозерновые, свежие фрукты и овощи, бобовые и орехи.
Несколько советов по увеличению потребления клетчатки:
- Ешьте цельные фрукты вместо фруктовых соков.
- Замените белый рис, хлеб и макароны коричневым рисом и цельнозерновыми продуктами.
- На завтрак выбирайте злаки, в качестве первого ингредиента которых используются цельнозерновые продукты.
- Закуска из сырых овощей вместо чипсов, крекеров или плиток шоколада.
- Заменяйте мясо фасолью или бобовыми овощами два-три раза в неделю в чили и супах.
Клетчатка и болезни
Клетчатка снижает риск развития различных состояний, включая болезни сердца, диабет, дивертикулярную болезнь и запоры. Несмотря на эти преимущества, клетчатка, вероятно, мало влияет на риск рака толстой кишки.
Болезнь сердца
Высокое потребление пищевых волокон было связано с более низким риском сердечно-сосудистых заболеваний в ряде крупных исследований, которые проводились на людях в течение многих лет.(16) В Гарвардском исследовании, в котором приняли участие более 40 000 медицинских работников-мужчин, исследователи обнаружили, что высокое общее потребление пищевых волокон связано с 40-процентным снижением риска ишемической болезни сердца. (17) Связанное с этим Гарвардское исследование медсестер дало очень похожие результаты. (18)
Более высокое потребление клетчатки также связано с более низким риском метаболического синдрома, сочетания факторов, повышающих риск развития сердечных заболеваний и диабета. Эти факторы включают высокое кровяное давление, высокий уровень инсулина, избыточный вес (особенно в области живота), высокий уровень триглицеридов и низкий уровень холестерина ЛПВП (хорошего).Несколько исследований показывают, что более высокое потребление клетчатки может иметь защитные преимущества от этого синдрома. (19,20)
Сахарный диабет 2 типа
Диеты с низким содержанием клетчатки и высоким содержанием пищевых продуктов, вызывающие внезапное повышение уровня сахара в крови, могут повысить риск развития диабета 2 типа. Оба гарвардских исследования — медсестер и мужчин-медиков — показали, что этот тип диеты более чем вдвое увеличивает риск диабета 2 типа по сравнению с диетой с высоким содержанием зерновых волокон и низким содержанием продуктов с высоким гликемическим индексом.(21-23) Диета с высоким содержанием зерновых волокон была связана с более низким риском диабета 2 типа.
Другие исследования, такие как Исследование здоровья чернокожих женщин (24) и Европейское проспективное исследование рака и питания в Потсдаме, показали аналогичные результаты.
Прочтите о том, что вы можете сделать, чтобы предотвратить диабет 2 типа.
Дивертикулярная болезнь
Дивертикулит, воспаление кишечника, является одним из наиболее распространенных возрастных заболеваний толстой кишки в западном обществе.Среди мужчин-медиков, участвовавших в долгосрочном последующем исследовании, употребление пищевых волокон, особенно нерастворимых, было связано с примерно на 40% снижением риска дивертикулярной болезни. (25)
Клетчатка и запор
Запор — самая распространенная желудочно-кишечная жалоба в Соединенных Штатах, а потребление клетчатки, по-видимому, облегчает и предотвращает запор.
Клетчатка в пшеничных и овсяных отрубях считается более эффективной, чем клетчатка из фруктов и овощей.Эксперты рекомендуют увеличивать потребление клетчатки постепенно, а не внезапно, и, поскольку клетчатка поглощает воду, потребление напитков следует увеличивать по мере увеличения потребления клетчатки.
Рак толстой кишки
Исследования в значительной степени не смогли показать связь между клетчаткой и раком толстой кишки. Одно из них — исследование Гарварда, в котором за 16 лет наблюдали более 80 000 медсестер — показало, что пищевые волокна не сильно связаны со снижением риска рака толстой кишки или полипов (предшественников рака толстой кишки).(26)
Следуя по научному следу: История о клетчатке и раке толстой кишкиПоскольку наука — это такой динамичный процесс, вы никогда не можете точно сказать, куда он вас приведет. Выводы, которые когда-то казались логичными и достаточно убедительными, могут быть пересмотрены — или полностью отменены — по мере того, как по определенной теме проводится все больше и больше исследований. Одним из примеров этого является взаимосвязь между клетчаткой и раком толстой кишки.
Примерно 30 лет назад высокое потребление клетчатки регулярно рекомендовалось как один из способов снизить риск рака толстой кишки.Эта рекомендация в значительной степени основана на наблюдениях, что в странах с высоким потреблением клетчатки, как правило, уровень заболеваемости раком толстой кишки ниже, чем в странах с низким потреблением клетчатки.
Но такие описательные исследования не дают наиболее точной информации. Хотя они часто являются хорошей отправной точкой для начала научного путешествия, они позволяют взглянуть только на большие группы людей. В описательных исследованиях, как правило, не учитываются все факторы, которые могут объяснить различия в уровне заболеваемости.Потребление клетчатки действительно может иметь какое-то отношение к различиям в заболеваемости раком толстой кишки, но эти различия могут также включать многие другие факторы, которые различаются между странами, включая другие факторы питания или образа жизни.
Когда исследования, которые могут принимать во внимание такие вещи на индивидуальном уровне, начали рассматривать проблему клетчатки и рака толстой кишки, картина стала гораздо менее ясной. Ряд исследований методом случай-контроль показали, что высокое потребление клетчатки связано с более низким риском рака толстой кишки, но многие этого не сделали.Учитывая эти колеблющиеся результаты, а также поскольку исследования методом случай-контроль не являются оптимальным способом оценки потребления пищи, поскольку они полагаются на воспоминания участников о том, что они ели в прошлом, потребовались дополнительные исследования с использованием более совершенных методов. Между тем, многие медицинские работники по-прежнему регулярно рекомендуют высокое потребление клетчатки людям, пытающимся снизить риск рака толстой кишки.
Только после того, как появились результаты когортных исследований, эта рекомендация начала терять свою поддержку. Поскольку когортные исследования наблюдают за группой людей с течением времени, их результаты, как правило, сильнее, чем результаты исследований методом случай-контроль, особенно когда речь идет о таких вещах, как диета и рак толстой кишки.Большинство этих когортных исследований показало, что потребление клетчатки очень мало связано с раком толстой кишки.
Такие выводы были дополнительно подтверждены результатами рандомизированных исследований — типов исследований, которые многие считают золотым стандартом исследований. В этих исследованиях группа людей случайным образом распределялась в одну из двух групп. Одна группа была на диете с высоким содержанием клетчатки, а другая группа придерживалась диеты с низким содержанием клетчатки. Через 3–4 года две группы сравнили, и не было обнаружено различий в частоте полипов толстой кишки — доброкачественных новообразований, которые могут перерасти в рак.Конечно, полипы толстой кишки не являются раком, но поскольку считается, что все виды рака толстой кишки начинаются с полипов, это убедительное доказательство того, что потребление клетчатки не имеет прямой связи с раком толстой кишки.
В этом случае путь открытий вел от широко распространенной веры в четкую связь между клетчаткой и раком толстой кишки к признанию вероятности того, что между ними нет прочной связи. Таким образом, это отличный пример того, как часто могут развиваться исследования. То, что может начаться как четкая связь, основанная на результатах обширных описательных исследований, может постепенно разрушиться по мере того, как все более качественные исследования раскрывают истинную природу отношений.Однако учтите, что слабые отношения сложно исключить совсем. Дальнейшие исследования могут продемонстрировать слабое влияние клетчатки на рак толстой кишки, хотя такое открытие не изменит вывода о том, что необходимо искать другие средства для предотвращения рака толстой кишки.
Рак груди
Крупномасштабное исследование 2016 г. (27), проведенное учеными из Гарварда T.H. Школа общественного здравоохранения Чана показала, что более высокое потребление клетчатки снижает риск рака груди, предполагая, что потребление клетчатки в подростковом и раннем взрослом возрасте может быть особенно важным.
- Женщины, которые в подростковом и юношеском возрасте потребляют больше продуктов с высоким содержанием клетчатки, включая овощи и фрукты, могут иметь значительно более низкий риск рака груди, чем те, кто в молодом возрасте ест меньше клетчатки.
Список литературы
16. Перейра М.А., О’Рейли Э., Аугустссон К. и др. Пищевые волокна и риск ишемической болезни сердца: объединенный анализ когортных исследований. Арк Интерн Мед. . 2004; 164: 370-6.
17. Римм Э. Б., Аскерио А., Джованнуччи Э, Шпигельман Д., Штампфер М. Дж., Виллетт В.Потребление овощей, фруктов и зерновых волокон и риск ишемической болезни сердца среди мужчин. ЯМА . 1996; 275: 447-51.
18. Браун Л., Рознер Б., Уиллетт У. С., Сакс Ф. М.. Эффекты пищевых волокон по снижению холестерина: метаанализ. Ам Дж. Клин Нутр . 1999; 69: 30-42.
19. Маккеун Н.М., Мейгс Дж.Б., Лю С., Уилсон П.В., Жак П.Ф. Согласно исследованию Framingham Offspring, потребление цельного зерна благоприятно связано с метаболическими факторами риска диабета 2 типа и сердечно-сосудистых заболеваний. Ам Дж. Клин Нутр . 2002; 76: 390-8.
20. Маккеун Н.М., Мейгс Дж.Б., Лю С., Зальцман Э., Уилсон П.В., Жак П.Ф. Углеводное питание, инсулинорезистентность и распространенность метаболического синдрома в Framingham Offspring Cohort. Уход за диабетом . 2004; 27: 538-46.
21. Fung TT, Hu FB, Pereira MA, et al. Потребление цельного зерна и риск диабета 2 типа: проспективное исследование на мужчинах. Ам Дж. Клин Нутр . 2002; 76: 535-40.
22. Лю С., Виллетт В.С., Штампфер М.Дж. и др. Проспективное исследование пищевой гликемической нагрузки, потребления углеводов и риска ишемической болезни сердца у женщин в США. Ам Дж. Клин Нутр . 2000; 71: 1455-61.
23. Шульце М.Б., Лю С., Римм Э.Б., Мэнсон Дж. Э., Виллетт В. К., Ху Ф. Б.. Гликемический индекс, гликемическая нагрузка, потребление пищевых волокон и частота диабета 2 типа у женщин молодого и среднего возраста. Ам Дж. Клин Нутр . 2004; 80: 348-56.
24.Кришнан С., Розенберг Л., Зингер М. и др. Гликемический индекс, гликемическая нагрузка, потребление зерновых волокон и риск диабета 2 типа у чернокожих женщин в США. Арк Интерн Мед. . 2007; 167: 2304-9.
25. Алдури У.Х., Джованнуччи Э.Л., Рокетт Х.Р., Сэмпсон Л., Римм Э.Б., Уиллетт У. Проспективное исследование типов пищевых волокон и симптоматической дивертикулярной болезни у мужчин. Дж Нутрь . 1998; 128: 714-9.
26. Fuchs CS, Giovannucci EL, Colditz GA, et al. Пищевые волокна и риск колоректального рака и аденомы у женщин. N Английский язык J Med . 1999; 340: 169-76.
27. Фарвид М.С., Элиассен А.Х., Чо Э., Ляо Х, Чен В.Й., Виллетт В. Потребление пищевых волокон у молодых людей и риск рака груди. Педиатрия 2016: 137 (3).
Условия использования
Содержание этого веб-сайта предназначено для образовательных целей и не предназначено для предоставления личных медицинских консультаций. Вам следует обратиться за советом к своему врачу или другому квалифицированному поставщику медицинских услуг с любыми вопросами, которые могут у вас возникнуть относительно состояния здоровья.Никогда не пренебрегайте профессиональным медицинским советом и не откладывайте его обращение из-за того, что вы прочитали на этом веб-сайте. Nutrition Source не рекомендует и не одобряет какие-либо продукты.
Fast-Twitch Vs. Типы медленно сокращающихся мышечных волокон + советы по тренировкам
Хотите развить выносливость? А что насчет власти? Нужно ли разбивать мечты о том, чтобы стать звездным бомбардиром или марафонцем, если соотношение сокращений не идеальное? Не обязательно. Типы мышечных волокон, нацеленные на разные типы тренировочных программ, могут влиять на спортивные тренировочные цели.
В этой статье мы подробно исследуем два типа мышечных волокон и обсудим, как тренировать каждый тип в соответствии с спортивными целями.
Что такое мышечные волокна?
Скелетные мышцы состоят из отдельных мышечных волокон . И, как и сами мышцы, не все мышечные волокна одинаковы. Существует два типа волокон скелетных мышц, быстро сокращающиеся и медленно сокращающиеся, и каждый из них выполняет разные функции, которые важно понимать, когда дело доходит до программирования движений и упражнений.
Медленно сокращающиеся мышечные волокна устойчивы к усталости и ориентированы на постоянные, более мелкие движения и контроль осанки. Они содержат больше митохондрий и миоглобина и обладают аэробной природой по сравнению с быстро сокращающимися волокнами. Медленно сокращающиеся волокна также иногда называют типом I или красными волокнами из-за их кровоснабжения.
Быстро сокращающиеся мышечные волокна обеспечивают большую и мощную силу, но на меньшую продолжительность и быстро утомляются. Они более анаэробны с меньшим кровоснабжением, поэтому их иногда называют белыми волокнами или типом II.Скелетные мышцы содержат оба типа волокон, но их соотношение может различаться в зависимости от множества факторов, включая функцию мышц, возраст и тренировку.
Скелетные мышцы содержат волокна обоих типов, но их соотношение может различаться в зависимости от множества факторов, включая функцию мышц, возраст и тренировку. Если вы специалист по спортивным достижениям, важно помнить о различиях между этими двумя типами мышц.
Типы мышечных волокон с медленным сокращением и с быстрым сокращением
Двумя типами волокон скелетных мышц являются медленно сокращающиеся (тип I) и быстро сокращающиеся (тип II).Медленно сокращающиеся мышечные волокна поддерживают упражнения на выносливость на длинные дистанции, такие как марафонский бег, в то время как быстро сокращающиеся мышечные волокна поддерживают быстрые и мощные движения, такие как спринт или поднятие тяжестей.
Сравнительная таблица типов мышечных волокон
Характеристика | Медленно-сокращающийся тип I | Быстрое переключение типа IIA | Fast-Twitch, тип IIX или IIB |
Мероприятия | Марафоны, бег на длинные дистанции, плавание, велоспорт, силовая ходьба, тренировка на выносливость | Пауэрлифтинг, спринт, прыжки, силовые и ловкие тренировки | Пауэрлифтинг, спринт, прыжки, силовые и ловкие тренировки |
Размер мышечного волокна | Малый | Большой | Большой |
Производство сил | Низкая | Высокая | Очень высокий |
Устойчивость к усталости | Медленная | Быстрый | Очень быстро |
Скорость сокращения | Медленная | Быстрый | Очень быстро |
Митохондрии | Высокая | Средний | Низкая |
Капилляры | Высокая | Средний | Низкая |
| Миоглобин | Высокая | Средний | Низкий |
Уровень АТФазы | Низкая | Средний | Высокая |
Окислительная способность | Высокая | Средний | Низкая |
Медленно сокращающиеся мышечные волокна имеют высокие концентрации митохондрий и миоглобина.Хотя они меньше, чем быстросокращающиеся волокна, они окружены большим количеством капилляров (1,2). Эта комбинация поддерживает аэробный метаболизм и устойчивость к усталости , что особенно важно при длительных субмаксимальных (аэробных) нагрузках.
Волокна типа I создают меньшее усилие и на медленнее создают максимальное напряжение (более низкая активность миозиновой АТФазы) по сравнению с волокнами типа II. Но они способны поддерживать более длительные сокращения , что имеет ключевое значение для стабилизации и контроля позы (1,2).
Помните:
- Мелкие мышечные волокна
- Низкое, медленное усилие
- Утомляемость медленнее, чем у быстрых сокращений, тип II
- Долгосрочные схватки
- Поддерживает сопротивление усталости при аэробных нагрузках, стабилизации и контроле осанки
Fast-Twitch, тип II
Быстро сокращающиеся мышечные волокна типа II подразделяются на тип IIx и тип IIa.
Как правило, они имеют более низкие концентрации митохондрий, миоглобина и капилляров по сравнению с нашими медленно сокращающимися волокнами, что означает, что они на быстрее утомляются (1,2).
Эти волокна большего размера также создают большую и более быструю силу , что является важным соображением для силовых активностей (1,2).
Тип IIX (также известный как Тип IIB ) волокна производят наибольшую силу , но невероятно неэффективны из-за их высокой активности миозиновой АТФазы, низкой окислительной способности и сильной зависимости от анаэробного метаболизма (1,2 ).
Волокна типа IIA , также известные как промежуточных мышечных волокон, представляют собой смесь типа I и типа IIx со сравнимым натяжением. могут использовать как аэробные, так и анаэробные энергетические системы. , эти волокна обладают более высокой окислительной способностью и утомляются медленнее, чем тип IIx (1,2).
Помните:
- Крупные мышечные волокна
- Большая и быстрая сила
- Усталость быстрее, чем у медленных I
- Два типа: Тип II x и Тип II a
- Тип IIx обеспечивает наибольшую силу, но неэффективен (очень быстро устает)
- Тип IIa представляет собой смесь мышечных волокон типа I и типа IIx (утомляемость медленнее, чем у типа IIx)
- Краткосрочные контракты
- Поддерживает работу с электропитанием
Какой у вас тип мышечных волокон?
Итак, теперь, когда мы рассмотрели различные типы, вам интересно, что вы за тип? Какие мышцы есть у ваших рук, бицепсов, ног, груди и рук?
Вы и ваши мышцы не состоят из одного типа мышечных волокон. Все из ваши мышцы представляют собой смесь быстро сокращающихся и медленно сокращающихся мышечных волокон типов (1).
Будет ли у вас больше типа I или типа II, зависит от вашего уровня активности и возраста .
Уровень активности
Люди, не занимающиеся спортом, имеют баланс типов волокон примерно 50/50.
Когда вы начнете смотреть на высококвалифицированных и успешных спортсменов, могут начаться некоторые различия.
Силовые атлеты имеют более высокое соотношение быстро сокращающихся волокон (например, у спринтеров 70-75% типа II), тогда как у спортсменов на выносливость больше медленно сокращающихся волокон (например, у марафонцев / бегунов на длинные дистанции 70-80% типа I) (2).
Конечно, тип мышечных волокон — не единственный фактор успеха спортсмена! Есть много других переменных, которые переводят спортсмена из хорошего в класс .
Возраст
Возраст также является фактором для наших мышечных волокон.
Старение вызывает потерю на безжировой мышечной массы , с уменьшением на наших быстро сокращающихся волокон , особенно типа IIx, но также наблюдается увеличение на наших медленных волокон (2-4).
Напомним, что быстросокращающиеся волокна больше по размеру, чем медленно сокращающиеся, и являются метаболически эффективными волокнами. Таким образом, потеря мышечной массы может способствовать возрастным метаболическим дисфункциям , изменениям состава тела , даже повышенному риску падений (2-5).
Тренировки сопротивления помогут справиться с этим упадком .
Тренировка мышечных волокон типа I и типа II
Типы волокон можно изменять с помощью упражнений.
Мышечные волокна типа I можно развить с помощью тренировок на выносливость , например, с низким сопротивлением, частым повторением или длительными и низкоинтенсивными. (Как видно из этапов 1 и 2 OPT ™)
Мышечные волокна типа II можно развить с помощью силовых тренировок .
Тренировка с отягощениями увеличивает размер мышечных волокон как I, так и II типа. Более сильный рост (т.е. гипертрофия) происходит в волокнах типа II и увеличивает количество актиновых и миозиновых нитей. Это приводит к повышенной способности генерировать силу (2).
Быстросокращающиеся волокна также могут задействовать медленно сокращающиеся волокна. : тренировки на выносливость с интервалами высокой интенсивности могут быть эффективными для улучшения аэробной силы (2,6).
Сужение во время тренировочных программ (уменьшение объема и интенсивности), может также улучшить силу и мощность волокон типа IIA без снижения производительности типа I. (9).
В одном исследовании изучались изменения мышечных волокон при подготовке бегунов-любителей к марафону. После 13 недель увеличения пробега и трехнедельного цикла сужения не только улучшились функции волокон типа I и типа IIa, но и типа IIa продолжили значительно улучшаться во время цикла сужения (9).
Если вы заинтересованы в обучении спортсменов, в том, чтобы стать тренером по силовой и физической подготовке или получить подготовку в качестве специалиста по повышению производительности, свяжитесь с нами!
3 спортивных блога, которые стоит посетить
Список литературыClark M, Lucett S, McGill E, Montel I и Sutton B (редакторы).(2018). NASM Essentials of Personal Fitness Training (6 th ed). Берлингтон, Массачусетс: Jones & Bartlett Publishing.
- Пауэрс СК и Хоули Э. (2012). Физиология упражнений: теория и применение к фитнесу и производительности, (8 th Edition). Нью-Йорк, штат Нью-Йорк: Макгроу Хилл.
- Акасаки Ю., Оучи Н., Изумия Ю., Бернардо Б., ЛеБрассер Н. и Уолш К. (2013). Гликолитическое восстановление быстро сокращающихся мышечных волокон противодействует неблагоприятным возрастным изменениям в составе тела и обмене веществ.Ячейка старения 13: 80-91. DOI: 10.1111 / acel.12153
- Наричи М.В., Маффулли Н. (2010). Саркопения: характеристики, механизмы и функциональное значение. Британский медицинский бюллетень 95: 139-159. DOI: 10.1093 / bmb / ldq008
- Стюарт Калифорния, член парламента Маккарри, Марино А, Южный Массачусетс, Хауэлл МЭА, Лэйн А.С., Рэмси М.В. и Стоун М.Х. (2013) Доля медленно сокращающихся волокон в скелетных мышцах коррелирует с чувствительностью к инсулину. Журнал клинической эндокринологии и метаболизма 98: 5, 2027-2036. DOI: http: // dx.doi.org/10.1210/jc.2012-3876
- Ванхатало А., Пул, округ Колумбия, ДиМенна Ф.Дж., Бейли С.Дж. и Джонс А.М. (2011). Рекрутмент мышечных волокон и медленный компонент поглощения O2: постоянная скорость работы или спринтерские упражнения. Американский журнал физиологии — регуляторная, интегративная и сравнительная физиология. 300 : 3, 700-707. DOI: 10.1152 / ajpregu.00761.2010
- McGill E, Montel I. (Редакторы). (2019). Основы спортивной подготовки NASM (2-е изд.). Берлингтон, Массачусетс: Jones & Bartlett Publishing.
- Barh R (редактор). (2012). Руководство МОК по спортивным травмам. Чичестер, Западный Сассекс: Wiley-Blackwell / Jophn Wiley & Sons Ltd.
- Трапп С., Харбер М., Крир А., Галлахер П., Сливка Д., Минчев К. и Уитсетт Д. (2006). Адаптация отдельных мышечных волокон при марафонских тренировках. Журнал прикладной физиологии, 101: 3, 721-727. DOI: 10.1152 / japplphysiol.01595.2005
рев. 7 / 18-1 / 15
видов продуктов питания и пользы для здоровья
Каковы некоторые рекомендации по увеличению количества пищевых волокон?
Это руководство содержит основную информацию, которая поможет вам увеличить количество клетчатки в вашем рационе.Клетчатка — важное диетическое вещество для вашего здоровья. Большинство продуктов, содержащих клетчатку, также являются хорошими источниками витаминов, минералов и антиоксидантов, которые приносят много пользы для здоровья. Дипломированный диетолог может провести углубленное обучение по вопросам питания, чтобы помочь вам разработать личный план действий.
Что такое клетчатка?
Клетчатка — это структурная часть растительной пищи, такой как фрукты, овощи и злаки, которую наш организм не может переваривать или расщеплять. Есть два вида клетчатки: растворимая и нерастворимая.
- Растворимая клетчатка : растворяется в воде с образованием липкого геля. Он может замедлить прохождение пищи из желудка в кишечник. Примеры включают сушеные бобы, овес, ячмень, бананы, картофель и мягкие части яблок и груш.
- Нерастворимые волокна : часто называют «грубыми кормами», потому что они не растворяются в воде. Он удерживает воду, что помогает производить более мягкий и объемный стул, что помогает регулировать дефекацию. Примеры включают цельные отруби, цельнозерновые продукты, орехи, кукурузу, морковь, виноград, ягоды и кожуру яблок и груш.
Какие еще функции выполняет волокно?
Исследования показали, что диета, богатая клетчаткой, имеет множество преимуществ для здоровья, в том числе следующие:
- Снижает уровень холестерина: Было показано, что растворимая клетчатка снижает уровень холестерина, связываясь с желчью (состоящей из холестерина) и выводя ее из организма. Это может помочь снизить риск сердечных заболеваний.
- Лучше регулирует уровень сахара в крови: Еда с высоким содержанием клетчатки замедляет переваривание пищи в кишечнике, что может помочь предотвратить быстрое повышение уровня сахара в крови.
- Контроль веса: Диета с высоким содержанием клетчатки может помочь вам дольше оставаться сытым, что предотвращает переедание и чувство голода между приемами пищи.
- Может предотвратить рак кишечника: Нерастворимая клетчатка увеличивает объем и скорость движения пищи по кишечному тракту, что сокращает время накопления вредных веществ.
- Запор: Запор часто можно облегчить, увеличив количество клетчатки или грубых кормов в вашем рационе. Клетчатка помогает регулировать дефекацию, втягивая воду в толстую кишку, делая стул более мягким и объемным.Это действие способствует лучшей регулярности.
Сколько клетчатки мне нужно есть?
Академия питания и диетологии рекомендует потреблять около 25-35 граммов общей клетчатки в день, из которых 10-15 граммов — растворимой клетчатки или 14 граммов клетчатки на 1000 калорий. Этого можно достичь, выбрав 6 унций зерен (3 или более унций из цельного зерна), 2½ стакана овощей и 2 стакана фруктов в день (из расчета 2000 калорий в день). Однако с возрастом потребности в клетчатке снижаются.Для людей старше 70 лет рекомендуемая норма для женщин составляет 21 грамм, а для мужчин — 30 граммов клетчатки в день.
Примечание. Диета с высоким содержанием клетчатки может повлиять на усвоение и эффективность некоторых лекарств. Поговорите со своим врачом о том, какие лекарства принимать с осторожностью и когда их принимать. Клетчатка также связывается с определенными питательными веществами и выводит их из организма. Чтобы этого избежать, старайтесь потреблять 20-35 граммов клетчатки в день. При соблюдении диеты с высоким содержанием клетчатки обязательно выпивайте не менее восьми стаканов жидкости каждый день.
Советы по увеличению количества пищевых волокон в вашем рационе:
- Добавляйте клетчатку в свой рацион медленно. Слишком большое количество клетчатки сразу может вызвать спазмы, вздутие живота и запор.
- Добавляя клетчатку в свой рацион, обязательно пейте достаточное количество жидкости (не менее 64 унций или 8 чашек в день), чтобы предотвратить запоры.
- Выбирайте продукты, у которых в качестве первого ингредиента указано цельнозерновые , а не обогащенная мука. Цельнозерновая мука — это цельнозерновая мука, а пшеничная — нет.
- Выбирайте цельнозерновой хлеб с 2-4 граммами пищевых волокон на ломтик.
- Выбирайте злаки, содержащие не менее 5 граммов пищевых волокон на порцию.
- Выбирайте сырые фрукты и овощи вместо сока и ешьте кожуру.
- Попробуйте альтернативные варианты клетчатки, такие как цельнозерновая крупа, кускус из цельной пшеницы, киноа, булгур, зародыши пшеницы, семена чиа, семена конопли, чечевичные макароны и макароны эдамаме.
- Попкорн — цельнозерновой. Подавайте его с низким содержанием жира без масла, чтобы получить более здоровую закуску.
- Добавляйте отруби в супы, крупы, выпечку, соус для спагетти, фарш и запеканки. Также отруби хорошо смешиваются с апельсиновым соком.
- Используйте сушеный горох, фасоль и бобовые в основных блюдах, салатах или гарнирах, таких как рис или макароны.
- Добавляйте сухофрукты в йогурт, хлопья, рис и кексы.
- Попробуйте коричневый рис и цельнозерновые макаронные изделия.
Добавки клетчатки
Добавки с клетчаткоймогут быть вариантом, если вы не можете получить достаточное количество клетчатки из своего рациона.Добавки с клетчаткой можно использовать для нормализации как запора, так и диареи. Проконсультируйтесь с врачом, прежде чем начинать принимать какие-либо добавки. Внимательно читайте этикетки на волокне.
- Выпейте не менее 8 унций жидкости вместе с добавкой. Прием некоторых пищевых добавок с клетчаткой без достаточного количества жидкости может вызвать набухание клетчатки и вызвать удушье и запор.
- Некоторые пищевые добавки с клетчаткой, которые следует учитывать, — это Benefiber® (пшеничный декстрин), Metamucil® (псиллиум), Konsyl® (псиллиум), Citrucel® (метилцеллюлоза), Fiberco® (SmartFiber, полученный из целлюлозы) и FiberChoice® (инулин).Шелуха подорожника и гуаровая камедь — растворимые волокна.
- Подумайте о том, чтобы вести дневник питания и отслеживать, сколько клетчатки вы едите в течение обычного дня.
- Используйте таблицу содержания клетчатки в этом раздаточном материале в качестве руководства для достижения своей цели по высокому содержанию клетчатки или обратитесь на сайт www.NAL.usda.gov/fnic для получения дополнительной информации о содержании пищевых волокон в пище.
Содержание клетчатки в обычных пищевых продуктах
Хлеб
- Бублик из цельной пшеницы
- Размер порции: 3 1/2 дюйма
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Светло-белый / пшеничный
- Размер порции: 2 ломтика
- Всего клетчатки (граммы): 1
- Растворимая клетчатка (граммы): следы
- Пита-цельнозерновой
- Размер порции: 7 дюймов
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
- Пумперникель
- Размер порции: 1 ломтик
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Цельная пшеница
- Размер порции: 1 ломтик
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): следы
- Рожь
- Размер порции: 1 ломтик
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Двойное волокно
- Размер порции : 1 ломтик
- Всего клетчатки (граммы): 5
- Растворимая клетчатка (граммы): 2
Злаки
- Хлопья отрубей
- Размер порции: 3/4 стакана
- Всего клетчатки (граммы): 5
- Растворимая клетчатка (граммы): следы
- Cheerios ™
- Размер порции: 1 1/4 стакана
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
- Овсянка
- Размер порции: Приготовленная 1 чашка
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 2
- Fiber One ™
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 14
- Растворимая клетчатка (граммы): 1
- Все отруби ®
- Размер порции: 2/3 чашки
- Всего клетчатки (граммы): 13
- Растворимая клетчатка (граммы): 1
- Каши ® От сердца к сердцу ®
- Размер порции: 3/4 стакана
- Всего клетчатки (граммы): 5
- Растворимая клетчатка (граммы): 1
Зерна
- Ячмень
- Размер порции: 1/2 чашки приготовленной
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
- Коричневый рис
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): следы
- Макаронные изделия из цельной пшеницы
- Размер порции: Готовые 1/2 чашки
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Квиноа
- Размер порции: 1/2 чашки приготовленной
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Паста из чечевицы
- Размер порции: Готовые 1/2 чашки
- Всего клетчатки (граммы): 6
- Растворимая клетчатка (граммы): 2
- Паста Эдамаме
- Размер порции: Приготовленные 1/2 чашки
- Всего клетчатки (граммы): 6
- Растворимая клетчатка (граммы): 3
Бобовые и крахмалистые овощи
- Бобы гарбанзо
- Размер порции: 1/2 чашки
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
- Фасоль
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 6
- Растворимая клетчатка (граммы): 3
- Чечевица
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 5
- Растворимая клетчатка (граммы): 1
- Картофель (с кожурой)
- Размер порции: 1 средний
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Картофель сладкий
- Размер порции: Готовый 1/2 чашки
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 2
- Сквош (зима)
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 2
- Зеленый горошек, приготовленный
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
- Лима фасоль
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 7
- Растворимая клетчатка (граммы): 3
- Кукуруза, приготовленная
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): следы
Орехи и семена
- Миндаль
- Размер порции: 1/4 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Арахис
- Размер порции: 1/4 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Семена подсолнечника
- Размер порции: 1/4 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Грецкие орехи
- Размер порции: 1/4 стакана
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): следы
- Льняное семя (молотое)
- Размер порции: 1/8 стакана или 2 столовые ложки
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 2
- Семена чиа
- Размер порции: 1/8 стакана или 2 столовые ложки
- Всего клетчатки (граммы): 10
- Растворимая клетчатка (граммы): 7
- Семена конопли
- Размер порции: 1/8 стакана или 2 столовые ложки
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
Фрукты
- Яблоко с кожурой
- Размер порции: 1 средний
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Банан
- Размер порции: 1 средний
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Черника
- Размер порции: 1 чашка
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): следы
- Грейпфрут
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 1
- Растворимая клетчатка (граммы): 1
- Апельсин
- Размер порции: 1 средний
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 2
- Груша с кожурой
- Размер порции: 1 средний
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 2
- Чернослив
- Размер порции: 3
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Клубника
- Размер порции: 1 чашка
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 1
Овощи некрахмалистые
- Брокколи
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Брюссельская капуста
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 4
- Растворимая клетчатка (граммы): 2
- Капуста (зеленая)
- Размер порции: 1 стакан, свежий
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Морковь
- Размер порции: 1/2 чашки, приготовленные
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Цветная капуста
- Размер порции: 1/2 чашки, приготовленные
- Всего клетчатки (граммы): 1
- Растворимая клетчатка (граммы): следы
- Зеленые бобы
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Капуста
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 3
- Растворимая клетчатка (граммы): 1
- Шпинат
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 2
- Растворимая клетчатка (граммы): 1
- Кабачки (кабачки)
- Размер порции: 1/2 стакана
- Всего клетчатки (граммы): 1
- Растворимая клетчатка (граммы): 1
Как читать этикетку продукта
Этикетки для пищевых продуктов стандартизированы U.Закон о национальной маркировке и образовании правительства США (NLEA). Для большинства продуктов требуются этикетки с питанием и список ингредиентов, чтобы вы могли сделать лучший выбор для здорового образа жизни. Ознакомьтесь с этикеткой продукта ниже. Определите общее количество клетчатки в этом продукте или попросите своего диетолога или поставщика медицинских услуг показать вам, как читать этикетки на пищевых продуктах и применять эту информацию для ваших личных нужд. Чтобы продукт имел маркировку «с высоким содержанием клетчатки», он должен содержать 5 или более граммов пищевых волокон на порцию.
Классификация типов волокон скелетных мышц человека | Физиотерапия
Скелетные мышцы человека состоят из разнородного набора типов мышечных волокон. 1–3 Этот диапазон типов мышечных волокон обеспечивает широкий спектр возможностей, которые демонстрируют мышцы человека. Кроме того, мышечные волокна могут адаптироваться к изменяющимся требованиям, изменяя размер или состав волокон. Эта пластичность служит физиологической основой для многочисленных физиотерапевтических вмешательств, направленных на увеличение силы или выносливости пациента.Изменения в составе волокон также могут быть частично ответственны за некоторые нарушения и инвалидность, наблюдаемые у пациентов, потерявших физическую форму из-за длительного бездействия, иммобилизации конечностей или денервации мышц. 2 За последние несколько десятилетий количество доступных методов классификации мышечных волокон увеличилось, что привело к появлению нескольких систем классификации. Цель этого обновления — предоставить базовые знания, необходимые для чтения и интерпретации исследований скелетных мышц человека.
Типы мышечных волокон можно описать с помощью гистохимических, биохимических, морфологических или физиологических характеристик; однако классификации мышечных волокон по разным методикам не всегда совпадают. 1 Следовательно, мышечные волокна, которые могут быть сгруппированы вместе с помощью одного метода классификации, могут быть отнесены к разным категориям с использованием другого метода классификации. Для понимания методов классификации мышечных волокон необходимо базовое понимание структуры и физиологии мышц.
Обзор анатомии и физиологии мышечных волокон
Мышечные волокна состоят из функциональных единиц, называемых саркомерами. 3 Внутри каждого саркомера находятся миофибриллярные белки миозин (толстая нить) и актин (тонкая нить). Взаимодействие этих двух миофибриллярных белков позволяет мышцам сокращаться (рис. 1). 4 Несколько методов классификации различают волокна на основе различных структур миозина (изоформ) или физиологических возможностей. 1,2,5 Молекула миозина состоит из 6 полипептидов: 2 тяжелых цепей и 4 легких цепей (2 регуляторных и 2 щелочных). Регуляторная и щелочная легкая цепь связаны с каждой из тяжелых цепей. Тяжелые цепи содержат миозиновые головки, которые взаимодействуют с актином и позволяют мышцам сокращаться (рис. 1). 4 Тяжелая цепь миозина в области головы также содержит сайт связывания аденозинтрифосфата (АТФ) и служит ферментом (аденозинтрифосфатаза [АТФаза]) для гидролиза АТФ в аденозиндифосфат (АДФ) и неорганический фосфат (P I ) , который обеспечивает энергию, необходимую для сокращения мышц.Тонкая нить состоит из актина и двух регуляторных белков, тропонина и тропомиозина. 3 Когда мышечное волокно получает стимул в виде потенциала действия, Ca 2+ высвобождается из саркоплазматической сети. Затем кальций связывается с тропонином и через тропомиозин открывает сайт связывания миозина на молекуле актина (рис. 1). 4 В присутствии АТФ головка миозина связывается с актином и тянет тонкую нить вдоль толстой нити, позволяя саркомеру укорачиваться.Пока присутствуют Ca 2+ и АТФ, миозиновые головки будут прикрепляться к молекулам актина, притягивать актин, высвобождать и снова присоединяться. Этот процесс известен как циклическое переключение мостов. Скорость, с которой может происходить поперечный мостиковый цикл, ограничена в основном скоростью, с которой АТФаза миозиновой головки может гидролизовать АТФ.
Рисунок 1
Регуляторная функция тропонина и тропомиозина. Тропонин — это небольшой глобулярный белок с 3 субъединицами (TnT, TnI, TnC).(A) Состояние покоя: тропомиозин в условиях покоя блокирует активные участки актина, предотвращая связывание актина и миозина. (B) Сокращение: когда тропонин связывается с Ca 2+ , он претерпевает конформационные изменения и вытягивает тропомиозин из блокирующего положения на актиновом филаменте, позволяя миозиновым головкам образовывать поперечные мостики с актином. От Ploughman SA, Smith DL. Физиология упражнений для здоровья, фитнеса и работоспособности . Бостон, Массачусетс: Аллин и Бэкон; 1997: 433.Авторские права 1997 г. принадлежат Allyn & Bacon. Перепечатано / адаптировано с разрешения.
Рисунок 1
Регуляторная функция тропонина и тропомиозина. Тропонин — это небольшой глобулярный белок с 3 субъединицами (TnT, TnI, TnC). (A) Состояние покоя: тропомиозин в условиях покоя блокирует активные участки актина, предотвращая связывание актина и миозина. (B) Сокращение: когда тропонин связывается с Ca 2+ , он претерпевает конформационные изменения и вытягивает тропомиозин из блокирующего положения на актиновом филаменте, позволяя миозиновым головкам образовывать поперечные мостики с актином.От Ploughman SA, Smith DL. Физиология упражнений для здоровья, фитнеса и работоспособности . Бостон, Массачусетс: Аллин и Бэкон; 1997: 433. Авторские права 1997 г. принадлежат Allyn & Bacon. Перепечатано / адаптировано с разрешения.
Набор мышечных волокон
Первоначально целые мышцы классифицировались как быстрые или медленные в зависимости от скорости сокращения. 3 Это разделение также соответствовало морфологическим различиям: быстрые мышцы выглядели белыми у некоторых видов, особенно птиц, а медленные — красными.Покраснение является результатом большого количества миоглобина и высокого содержания капилляров. 3 Повышенное содержание миоглобина и капилляров в красных мышцах способствует большей окислительной способности красных мышц по сравнению с белыми мышцами. Гистологический анализ показывает, что существует корреляция между активностью миозиновой АТФазы и скоростью укорачивания мышц. 6 Этот гистохимический анализ привел к первоначальному разделению мышечных волокон на тип I (медленный) и тип II (быстрый). В настоящее время типирование мышечных волокон осуществляется тремя различными методами: гистохимическим окрашиванием на миозин-АТФазу, идентификацией изоформ тяжелой цепи миозина и биохимической идентификацией метаболических ферментов.
Окрашивание миозиновой АТФазой
У человека скорость гидролиза миозин-АТФазой быстрых волокон в 2–3 раза выше, чем у медленных. 7 Однако гистохимическое окрашивание миозин-АТФазы, которое широко используется для классификации мышечных волокон, не позволяет оценить скорость гидролиза миозин-АТФазы. 1 Волокна разделяются исключительно на основе интенсивности окрашивания из-за различий в чувствительности pH, а не из-за относительной скорости гидролиза АТФаз. 1 Достижения в технике гистохимического окрашивания, используемой для оценки миозин-АТФазы, привели к 7 признанным типам мышечных волокон человека (рис. 2). 1 Первоначально волокна определялись как типы I, IIA или IIB. 1,5 Совсем недавно были идентифицированы типы IC, IIC, IIAC и IIAB, которые обладают промежуточными характеристиками окрашивания миозиновой АТФазой. Самое медленное волокно, тип IC, имеет характеристики окрашивания, больше похожие на волокна типа I, тогда как самое быстрое волокно, тип IIAC, окрашивает больше, чем тип IIA.Волокна типа IIAB имеют промежуточные характеристики окрашивания между волокнами типа IIA и IIB. Поскольку эти разграничения основаны на качественном анализе окрашенных волокон, остается возможность, что в будущем будет идентифицировано больше типов волокон. Таким образом, 7 типов мышечных волокон человека, идентифицированные гистохимическим окрашиванием миозин-АТФазы (от самого медленного до самого быстрого): типы I, IC, IIC, IIAC, IIA, IIAB и IIB (рис. 2). 1,3,5 Эти деления основаны на интенсивности окрашивания при разных уровнях pH, и поэтому любое данное волокно может быть сгруппировано по-разному разными исследователями.Кроме того, не во всех исследованиях используются все 7 типов волокон. Некоторые исследователи относят все мышечные волокна к исходным 3 типам волокон.
Рисунок 2
Сравнение 3 различных классификаций типов волокон скелетных мышц: гистохимическое окрашивание на миозин-аденозинтрифосфатазу (mATPase), идентификация тяжелых цепей миозина и биохимическая идентификация метаболических ферментов. Примечание: у людей MHCIIb теперь более точно обозначается как MHCIIx / d. Знаки вопроса указывают на слабую корреляцию между схемами классификации биохимических и тяжелых цепей миозина или мАТФазных волокон.
Рисунок 2
Сравнение 3 различных классификаций типов волокон скелетных мышц: гистохимическое окрашивание на миозин-аденозинтрифосфатазу (mATPase), идентификация тяжелых цепей миозина и биохимическая идентификация метаболических ферментов. Примечание: у людей MHCIIb теперь более точно обозначается как MHCIIx / d. Знаки вопроса указывают на слабую корреляцию между схемами классификации биохимических и тяжелых цепей миозина или мАТФазных волокон.
Идентификация тяжелой цепи миозина
Идентификация различных изоформ тяжелой цепи миозина также позволяет классифицировать волокна по типу (рис.2). 1 Различные волокна на основе миозин-АТФазы соответствуют различным изоформам тяжелой цепи миозина. 1,8 Это неудивительно, потому что тяжелые цепи миозина содержат сайт, который служит АТФазой. Тот факт, что каждое мышечное волокно может содержать более одной изоформы тяжелой цепи миозина, объясняет существование типов волокон миозин-АТФазы, отличных от чистых волокон типа I, типа IIA и типа IIB. Хотя геном человека содержит по крайней мере 10 генов тяжелых цепей миозина, только 3 из них экспрессируются в мышцах конечностей взрослого человека. 1 Изоформы тяжелой цепи миозина могут быть идентифицированы с помощью иммуногистохимического анализа с использованием антител к антимиозину или с помощью электрофоретического разделения додецилсульфат натрия и полиакриламидного геля (SDS-PAGE). 5
Три изоформы миозина, которые были первоначально идентифицированы, были MHCI, MHCIIa и MHCIIb, и они соответствовали изоформам, идентифицированным при окрашивании миозиновой АТФазой как типы I, IIA и IIB, соответственно. 1,3,5 Смешанные волокна человека почти всегда содержат изоформы тяжелой цепи миозина, которые являются «соседями» (т.е. MHCI и MHCIIa или MHCIIa и MHCIIb). 2 Следовательно, волокна гистохимической миозин-АТФазы типа IC, IIC и IIAC коэкспрессируют гены MHCI и MHCIIa в разной степени, тогда как волокна типа IIAB коэкспрессируют гены MHCIIa и MHCIIb. 1 Из-за своей количественной природы идентификация изоформ тяжелой цепи миозина с помощью электрофоретического разделения отдельных волокон (метод SDS-PAGE), вероятно, представляет собой лучший метод типирования мышечных волокон. Электрофоретическое разделение позволяет определять относительные концентрации различных изоформ тяжелой цепи миозина в смешанном волокне. 5,8
Один момент, касающийся изоформ тяжелой цепи миозина человека и идентификации типа волокна, может сбить с толку кого-то, кто пытается читать исследовательскую литературу в этой области. У мелких млекопитающих присутствует четвертая изоформа тяжелой цепи миозина, MHCIIx или MHCIId, которая имеет промежуточную скорость сокращения между изоформой MHCIIa и MHCIIb. 9 Основываясь на нескольких типах доказательств, вплоть до уровня анализа ДНК, то, что первоначально было идентифицировано у людей как MHCIIb, фактически гомологично MHCIIx / d мелких млекопитающих. 2,5,9 В результате то, что у людей называется MHCIIb, на самом деле является MHCIIx / d, и люди не экспрессируют самую быструю изоформу тяжелой цепи миозина (MHCIIb). 5 Поскольку номенклатура типов волокон гистохимической миозин-АТФазы была разработана с использованием мышц человека, волокна типа IIB, которые, как мы теперь знаем, соответствуют изоформе тяжелой цепи миозина MHCIIx / d, вряд ли будут переименованы в тип IIX. 1 Следовательно, в зависимости от автора, человеческие волокна типа IIB на основе гистохимической миозин-АТФазы могут быть связаны либо с изоформами MHCIIb, либо с MHCIIx / d.Важно помнить, что в мышцах конечностей человека присутствуют только 3 изоформы тяжелой цепи миозина (от самой медленной до самой быстрой): MHCI, MHCIIa и MHCIIx / d (ранее ошибочно определялись как MHCIIb). 1 Люди не экспрессируют самую быструю изоформу тяжелой цепи миозина, MHCIIb. 9 В оставшейся части статьи мы будем ассоциировать MHCIIx / d у людей с гистохимическим волокном типа IIB на основе миозин-АТФазы.
Биохимический
Третья схема классификации, которая часто используется для классификации мышечных волокон, объединяет информацию о гистохимии миозин-АТФазы мышечных волокон и качественной гистохимии определенных ферментов, которые отражают энергетический метаболизм волокна (рис.2). 2 Гистохимическое типирование миозин-АТФазы волокон используется для классификации мышечных волокон как типа I или типа II, которые, как известно, соответствуют медленным и быстрым мышечным волокнам, соответственно. 2 Анализируемые ферменты отражают метаболические пути, которые являются аэробными / окислительными или анаэробными / гликолитическими. 5 Этот метод классификации позволяет выделить 3 типа волокон: быстро сокращающиеся гликолитические (FG), быстро сокращающиеся окислительные (FOG) и медленно сокращающиеся окислительные (SO). 2,3 Хотя существует хорошая корреляция между волокнами типа I и SO, корреляции между волокнами типа IIA и FOG и типа IIB и волокон FG более разнообразны. 3,10 Следовательно, волокна типа IIB не всегда зависят в первую очередь от анаэробного / гликолитического метаболизма, а волокна типа IIA не всегда зависят в первую очередь от аэробного / окислительного метаболизма. 5 Хотя, как правило, волокна на конце континуума типа I зависят от аэробного / окислительного энергетического метаболизма, а волокна на конце континуума типа IIB зависят от анаэробного / гликолитического метаболизма, корреляция недостаточно сильна для типа IIB. и FG или тип IIA и FOG должны использоваться как взаимозаменяемые. 2,5
Легкие миозиновые цепи
Легкие цепи молекулы миозина также существуют в различных изоформах, медленных и быстрых, которые влияют на сократительные свойства мышечного волокна. 3,11 Мышечные волокна, гомогенные для изоформы тяжелой цепи миозина (т.е. чистое волокно), могут быть гетерогенными в отношении изоформ легкой цепи миозина, хотя в целом быстрых изоформ тяжелой цепи миозина ассоциируются с быстрыми изоформами тяжелой цепи миозина. изоформ легкой цепи миозина и медленных изоформ тяжелой цепи миозина связываются с медленными изоформами легкой цепи миозина . 2,5,12 Существуют убедительные доказательства того, что дополнительные белки в мышечных волокнах экспрессируются вместе, так что различные «быстрые» белки экспрессируются друг с другом, а различные «медленные» белки экспрессируются друг с другом, что предполагает «волокно. специфическая для типа программа экспрессии генов ». 2,11,12
Классификация моторных агрегатов
Хотя мы обсуждали типы волокон, истинной функциональной единицей нервно-мышечной системы является двигательная единица. 13,14 Двигательная единица — это альфа-мотонейрон (происходящий из спинного мозга) и все мышечные волокна, которые он иннервирует. На основании гистохимии миозин-АТФазы и качественной гистохимии ферментов, которые отражают энергетический метаболизм волокна, все мышечные волокна двигательной единицы имеют схожие характеристики. 15 Двигательные единицы можно разделить на группы в зависимости от сократительной способности и утомляемости мышечных волокон. 3,14 В зависимости от скорости сокращения двигательные единицы классифицируются как медленно сокращающиеся (S) или быстро сокращающиеся (F). 14 Двигатели F подразделяются на быстро сокращающиеся, устойчивые к утомлению (FR), быстро сокращающиеся, средние при утомляемости (Fint) и быстро сокращающиеся утомляющие (FF). 16,17
Моторная единица / пластичность мышечного волокна
Независимо от схемы классификации, используемой для группировки мышечных волокон, есть неопровержимые доказательства того, что мышечные волокна — и, следовательно, двигательные единицы — не только изменяются в размере в ответ на требования, но также могут преобразовываться из одного типа в другой. 2,18,19 Эта пластичность сократительных и метаболических свойств в ответ на стимулы (например, тренировка и реабилитация) позволяет адаптироваться к различным функциональным требованиям. 2 Преобразования волокон между типом IIB и типом IIA являются наиболее распространенными, но преобразования типа I в тип II возможны в случаях тяжелого разрушения или повреждения спинного мозга (SCI). 2,20
Существует меньше доказательств превращения волокон типа II в волокна типа I при тренировках или реабилитации, потому что только исследования, в которых используются денервированные мышцы, которые хронически активируются с помощью электростимуляции, последовательно демонстрируют, что такое преобразование возможно. 21
Изменения типов мышечных волокон также ответственны за некоторую потерю функции, связанную с нарушением кондиционирования. 2 Эксперименты на животных, включающие подвешивание задних конечностей, которое разгружает мышцы задних конечностей, и наблюдения за людьми и крысами после воздействия микрогравитации во время космического полета продемонстрировали переход от медленных к быстрым типам мышечных волокон. 2 Кроме того, многочисленные исследования животных и людей с ТСМ продемонстрировали переход от медленных к быстрым волокнам. 2,20 Было показано, что у людей ослабление тренированности (то есть снижение использования мышц по сравнению с ранее высоким уровнем активности) приводит к такому же медленному превращению в быстрое со сдвигом от MHCIIa к MHCIIx / d и, возможно, MHCI к MHCIIa . 2 Существует также сопутствующее снижение ферментов, связанных с аэробно-окислительным метаболизмом. 2 Таким образом, уменьшение использования скелетных мышц может привести к преобразованию типов мышечных волокон из медленного в быстрое направление.
Интересно, что некоторая потеря работоспособности мышц (например, снижение выработки силы) из-за старения, по-видимому, происходит не только из-за преобразования мышечных волокон из одного типа в другой, но в основном из-за избирательной атрофии определенных групп населения. типов мышечных волокон. 22,23 С возрастом происходит прогрессирующая потеря мышечной массы и максимального потребления кислорода, что приводит к снижению работоспособности мышц и, предположительно, к некоторой потере функции (например, снижению способности выполнять повседневную деятельность), наблюдаемой в пожилые люди. 1,22,23
Потеря мышечной массы, связанная с возрастом, в первую очередь связана с уменьшением общего количества волокон как типа I, так и типа II и, во-вторых, из-за преимущественной атрофии волокон типа II. 22,24 Атрофия волокон типа II приводит к увеличению доли мышечной массы медленного типа в старых мышцах, о чем свидетельствует более медленное время сокращения и расслабления в старых мышцах. 25,26 Кроме того, потеря альфа-мотонейронов с возрастом приводит к некоторой реиннервации «покинутых» мышечных волокон соседними двигательными единицами, которые могут быть другого типа. 22,27 Это может способствовать преобразованию типа волокна, поскольку повторно иннервируемые мышечные волокна приобретают свойства новой «родительской» двигательной единицы. 3,22 Недавние данные о старых мышцах предполагают, что может происходить преобразование типа волокон, потому что у пожилых людей наблюдается гораздо большая коэкспрессия тяжелой цепи миозина по сравнению с молодыми людьми. 28 Было обнаружено, что более старые мышцы имеют больший процент волокон, которые совместно экспрессируют MHCI и MHCIIa (28,5%) по сравнению с более молодыми мышцами (5–10%). 28
К счастью, физиотерапевтические вмешательства могут повлиять на типы мышечных волокон, что приведет к улучшению работы мышц. В контексте этого обновления физиотерапевтические вмешательства можно в общих чертах разделить на те, которые предназначены для повышения устойчивости пациента к утомлению, и те, которые предназначены для увеличения выработки силы пациентом.В течение некоторого времени было известно, что тренировки, которые предъявляют высокие метаболические требования к мышцам (тренировка на выносливость), увеличивают окислительную способность всех типов мышечных волокон, в основном за счет увеличения количества митохондрий, аэробных / окислительных ферментов и капилляризации тренированная мышца. 29,30 Использование системы классификации, основанной на метаболических ферментах, может привести к переходу от FG к FOG мышечным волокнам без обязательного преобразования изоформ тяжелой цепи миозина. 2
Состав тяжелой цепи миозина в мышечном волокне может измениться при тренировке на выносливость. 19 Внутри волокон типа II происходит преобразование из IIB в IIA, при этом экспрессируется больше MHCIIa за счет MHCIIx / d. 2,19 Следовательно, процентное содержание чистых волокон типа IIB уменьшается, а процентное содержание волокон типа IIAB и чистого типа IIA увеличивается. Отсутствуют доказательства того, что волокна типа II превращаются в тип I при тренировке на выносливость, 19 , хотя, похоже, наблюдается увеличение популяции волокон смешанного типа I и IIA. 2 Исследователи обнаружили, что волокна типа I у людей становятся быстрее при упражнениях на выносливость и медленнее при нарушении кондиционирования. 31,32 Это изменение скорости сокращения происходит не из-за преобразования типов волокон, а скорее из-за изменений изоформ легкой цепи миозина из медленных изоформ в быстрые и из быстрых изоформ в медленные, соответственно. 31,32 Поскольку это изменение скорости мышечного сокращения не происходит за счет изменения миозиновой АТФазы, оно не может быть обнаружено гистохимическим типированием волокон. 2 Переход от медленных к быстрым изоформ легкой цепи миозина позволяет медленным волокнам сокращаться со скоростью, достаточно быстрой для данного упражнения (например, бега, езды на велосипеде), сохраняя при этом эффективные свойства использования энергии. 30 Таким образом, адаптация мышечных волокон к упражнениям на выносливость зависит от типа волокон, хотя окислительная способность всех волокон увеличивается. Волокна типа I могут становиться быстрее за счет преобразования легкой цепи миозина, тогда как волокна типа II превращаются в более медленные, более окислительные типы.
Высокоинтенсивная тренировка с отягощениями (например, тренировка с высокой нагрузкой и малым количеством повторений) приводит к изменениям типа волокон, аналогичным тем, которые наблюдаются при тренировках на выносливость, хотя гипертрофия мышц также играет важную роль в увеличении силы. 33 Первоначальное увеличение выработки силы с помощью программ высокоинтенсивных тренировок с отягощениями в значительной степени опосредовано нервными факторами, а не видимой гипертрофией мышечных волокон у взрослых без патологии или нарушений. 34 Даже в этом случае изменения в мышечных белках, таких как тяжелые цепи миозина, действительно начинаются после нескольких тренировок, но видимая гипертрофия мышечных волокон не проявляется до тех пор, пока тренировка не будет проводиться в течение более длительного периода времени (> 8 недель). 33
Большинство исследователей обнаружили, что высокоинтенсивные тренировки с отягощениями достаточной продолжительности (> 8 недель) вызывают увеличение состава MHCIIa и соответствующее уменьшение состава MHCIIx / d. 35–37 Во многих исследованиях высокоинтенсивных тренировок с отягощениями исследователи также сообщали о сопутствующем увеличении состава MHCI, 37 , хотя некоторые исследователи не сообщают об изменениях в составе MHCI. 38,39 Как тренировки на выносливость, так и тренировки с отягощениями приводят к аналогичному снижению коэкспрессии тяжелых цепей миозина, так что присутствует большее количество «чистых» волокон. 40 Хотя тенденции преобразования типов волокон аналогичны для тренировок на выносливость и силовых тренировок, различия в физиологических изменениях, которые происходят с каждым типом упражнений, также важны. Тренировка на выносливость увеличивает окислительную способность мышц, тогда как тренировка для увеличения выработки силы достаточной интенсивности и продолжительности способствует гипертрофии мышечных волокон за счет увеличения объема сократительных белков в волокнах.
Знание различий между типами волокон скелетных мышц человека позволяет клиницистам более полно понять морфологические и физиологические основы эффективности физиотерапевтических вмешательств, таких как тренировки на выносливость и тренировки с отягощениями.Кроме того, эти знания также предлагают некоторое объяснение изменений в мышцах, которые происходят с возрастом, нарушением условий, иммобилизацией и денервацией мышц. Такие знания полезны для оптимального проектирования программ реабилитации, направленных на снижение морфологии и физиологии мышц.
Список литературы
1Старон
RS
.Типы волокон скелетных мышц человека: описание, развитие и распределение
.Можно использовать Physiol
.1997
;22
:307
—327
,2Петте
D
,Старон
РС
.Переходные типы волокон скелетных мышц млекопитающих
.Int Rev Cytol
.1997
;170
:143
—223
,3МакКомас
AJ
.Скелетные мышцы: форма и функции
. Шампейн, штат Иллинойс: Human Kinetics,1996
. 4Пахарь
SA
,Смит
DL
.Физиология упражнений для здоровья, фитнеса и работоспособности
.Бостон, Массачусетс
:Allyn & Bacon
,1997
:433
,5Петте
D
,Peuker
H
,Staron
RS
.Влияние биохимических методов на анализ отдельных мышечных волокон
.Acta Physiol Scand
.1999
;166
:261
—277
,6Бараны
м
.АТФазная активность миозина коррелировала со скоростью укорачивания мышц
.J Gen Physiol
.1967
;50
:197
—218
.7Тейлор
AW
,Эссен
B
,Saltin
B
.Миозин-АТФаза в скелетных мышцах здоровых мужчин
.Acta Physiol Scand
.1974
;91
:568
–570
,8Фрай
AC
,Allemeier
CA
,Staron
RS
.Корреляция между процентом площади типа волокна и содержанием тяжелой цепи миозина в скелетных мышцах человека
.Eur J Appl Physiol Occup Physiol
.1994
;68
:246
–251
,9Хильбер
К
,Galler
S
,Gohlsch
B
,Pette
D
.Кинетические свойства изоформ миозиновой цепи в отдельных волокнах скелетных мышц человека
.FEBS Lett
.1999
;455
:267
—270
.10Хамалайнен
N
,Петле
D
.Паттерны изоформ миозина в волокнах скелетных мышц млекопитающих
.Microsc Res Tech
.1995
;30
:381
—389
.11Талмадж
RJ
,Рой
РР
,Эджертон
ВР
.Типы и функции мышечных волокон
.Curr Opin Rheumatol
.1993
;5
:695
—705
.12Jostarndt-Fogen
К
,Puntschart
A
,Hoppeler
H
,Billeter
R
.Волоконно-специфическая экспрессия быстрых и медленных мРНК незаменимых легких цепей миозина в тренированных скелетных мышцах человека
.Acta Physiol Scand
.1998
;164
:299
—308
,13Берк
РЭ
.Комментарий о наличии моторного блока «Тип
». В кн .: Башня БД, изд.Основные нейронауки
. Нью-Йорк, Нью-Йорк: Raven Press,1975
.Нервная система
.;том 1.
14Берк
РЭ
.Возвращение к понятию «типы двигателей
.».Prog Brain Res
.1999
;123
:167
—175
,15Берк
РЭ
,Levine
PN
,Zajac FE
III
.Двигательные единицы млекопитающих: физиолого-гистохимическая корреляция трех типов у кошек gastrocnemius
.Наука
.1971
;174
:709
—712
,16Берк
РЭ
.Типы двигательных единиц трехглавой мышцы бедра кошки
.Дж. Физиол
.1967
;193
:141
—160
,17Sieck
GC
,Prakash
YS
.Морфологические адаптации нервно-мышечных соединений зависят от типа волокна
.Можно использовать Physiol
.1997
;22
:197
—230
,18Гроссман
EJ
,Рой
RR
,Talmadge
RJ
и др. .Влияние неактивности на состав тяжелой цепи миозина и размер волокон камбаловидной мышцы крысы
.Мышечный нерв
.1998
;21
:375
—389
,19Ricoy
JR
,Encinas
AR
,Cabello
A
и др. .Гистохимическое исследование типов мышечных волокон широкой мышцы бедра у спортсменов
.Дж. Физиол Биохим
.1998
;54
:41
—47
.20Рой
руб. ,Talmadge
RJ
,Hodgson
JA
, et al. .Дифференциальная реакция быстрых мышц-разгибателей и сгибателей задних конечностей на упражнения у взрослых кошек с спинномозговой системой
.Мышечный нерв
.1999
;22
:230
—241
,21Экен
т
,Гундерсен
К
.Электростимуляция, напоминающая нормальную активность двигательных единиц: воздействие на денервированные быстрые и медленные мышцы крысы
.Дж. Физиол
.1988
;402
:651
—669
.22Ру
MR
,Рис
CL
,Вандервурт
AA
.Возрастные изменения функции моторного блока
.Мышечный нерв
.1997
;20
:679
–690
,23Портер
ММ
,Вандервурт
AA
,Lexell
J
.Старение мышц человека: структура, функции и адаптивность
.Scand J Med Sci Sports
.1995
;5
:129
—142
.24Лекселл
Дж
,Тейлор
CC
,Sjostrom
M
.В чем причина старческой атрофии
? Общее количество, размер и пропорция различных типов волокон, изученных во всей широкой латеральной мышце широкой мышцы бедра, у мужчин в возрасте от 15 до 83 лет.J Neurol Sci
.1988
;84
:275
—294
.25Наричи
МВ
,Бордини
M
,Cerretelli
P
.Влияние старения на функцию приводящей мышцы большого пальца человека
.J Appl Physiol
.1991
;74
:1227
–1281
,26Харридж
SD
,Крайгер
А
,Стенсгаард
А
.Сила, активация и размер разгибателей колена у очень пожилых людей после силовых тренировок
.Мышечный нерв
.1999
;22
:831
–839
,27Камень
G
,Sison
SV
,Du
CC
,Patten
C
.Разрядка моторных единиц у пожилых людей во время сокращений с максимальным усилием
.J Appl Physiol
.1995
;79
:1908
—1913
,28Андерсен
JL
,Terzis
G
,Kryger
A
.Повышение степени коэкспрессии изоформ тяжелой цепи миозина в волокнах скелетных мышц очень старого образца
.Мышечный нерв
.1999
;22
:449
—454
.29Holloszy
JO
,Стенд
FW
.Биохимические адаптации мышц к упражнениям на выносливость
.Анну Рев Физиол
.1976
;38
:273
—291
,30Фитинги
правая
,Видрик
JJ
.Мышечная механика: адаптация с упражнениями
.Exerc Sport Sci Rev
.1996
;24
:427
—473
,31Ларссон
л
,Li
XP
,Berg
HE
,Frontera
WR
.Влияние отмены функции опоры на сократительную способность и состав изоформ миозина в отдельных клетках скелетных мышц человека
.Арка Пфлюгерса
.1996
;432
:320
—328
.32Видрик
JJ
,Trappe
SW
,Blaser
CA
и др. .Изометрическая сила и максимальная скорость сокращения отдельных мышечных волокон у элитных бегунов-мастеров
.Am J Physiol
.1996
;271
(2 балла 1
):C666
—C675
.33Kraemer
WJ
,Флек
SJ
,Эванс
WJ
.Силовые тренировки: физиологические механизмы адаптации
.Exerc Sport Sci Rev
.1996
;24
:363
—397
,34МакАрдл
WD
,Катч
FI
,Катч
VL
.Основы физиологии упражнений
. Филадельфия, Пенсильвания: Леа и Фебигер,, 1994,
. 35Старон
RS
,Карапондо
DL
,Kraemer
WJ
и др. .Адаптация скелетных мышц на ранней стадии тренировок с отягощениями у мужчин и женщин
.J Appl Physiol
.1994
;76
:1247
–1255
,36Kraemer
WJ
,Patton
JF
,Gordon
SE
и др. .Совместимость высокоинтенсивных силовых и выносливых тренировок с гормональной адаптацией и адаптацией скелетных мышц
.J Appl Physiol
.1995
;78
:976
—989
.37Старон
RS
,Малики
ES
,Леонарди
MJ
и др. .Гипертрофия мышц и быстрое преобразование типов волокон у женщин, тренирующихся с отягощениями
.Eur J Appl Physiol Occup Physiol
.1990
;60
:71
–79
,38Адамс
GR
,Hather
BM
,Baldwin
KM
,Dudley
GA
.Состав тяжелых цепей миозина скелетных мышц и тренировки с отягощениями
.J Appl Physiol
.1993
;74
:911
–915
,39Хаккинен
К
,Ньютон
RU
,Гордон
SE
и др. .Изменения морфологии мышц, электромиографической активности и характеристик производства силы во время прогрессивных силовых тренировок у молодых и пожилых мужчин
.J Gerontol A Biol Sci Med Sci
.1998
;53
:B415
—B423
.40Уильямсон
DL
,Godard
MP
,Porter
DA
и др. .Прогрессивные тренировки с отягощениями снижают коэкспрессию тяжелых цепей миозина в отдельных мышечных волокнах у пожилых мужчин
.J Appl Physiol
.2000
;88
:627
—633
.© 2001 Американская ассоциация физиотерапии
Клетчатка — Типы, источники пищи, польза для здоровья и цельное или очищенное зерно — Питание: наука и повседневное применение
Пищевые волокна определены Советом по пищевым продуктам и питанию Института медицины как «неперевариваемые углеводы и лигнин, которые являются естественными и неповрежденными в растениях.«Клетчатка играет важную роль в обеспечении структуры и защиты растений, а также играет важную роль в питании человека.
Целлюлоза — это один тип волокна. Химическая структура целлюлозы показана на рисунке ниже с нашим упрощенным изображением рядом с ней. Вы можете видеть, что у целлюлозы есть длинные цепи глюкозы, похожие на крахмал, но они сложены друг с другом, и эти стопки соединены водородными связями. Особые связи между этими глюкозными единицами в клетчатке не перевариваются ферментативно в пищеварительном тракте, и поэтому клетчатка проходит непереваренной в толстую кишку или толстую кишку.
Рисунок 4.22. Химическая структура целлюлозы и упрощенная иллюстрация целлюлозы.
Вам может быть интересно, какая польза от клетчатки, если мы не можем ее переваривать. Однако он не проходит через пищеварительный тракт как отходы. Вместо этого он выполняет множество функций на своем пути, и они способствуют нашему здоровью. Давайте рассмотрим различные типы клетчатки, где мы находим их в продуктах питания и какую пользу они приносят!
Типы волокнаЦельные растительные продукты содержат множество различных типов молекул, которые подпадают под определение клетчатки.Один из способов классификации типов волокон — их растворимость в воде. Цельные растительные продукты содержат смесь как растворимой, так и нерастворимой клетчатки, но некоторые из них являются лучшими источниками одной, чем другой.
- Растворимое волокно — Эти волокна растворяются в воде, образуя вязкий гель в желудочно-кишечном тракте, который помогает замедлить пищеварение и усвоение глюкозы. Это означает, что включение растворимой клетчатки в еду помогает предотвратить резкие скачки сахара в крови, вместо этого способствует более постепенному повышению уровня глюкозы в крови.Диета с высоким содержанием растворимой клетчатки также может помочь снизить уровень холестерина в крови, поскольку растворимая клетчатка связывает холестерин и желчные кислоты (которые содержат холестерин) в желудочно-кишечном тракте. Растворимая клетчатка также хорошо ферментируется, поэтому она легко переваривается бактериями в толстом кишечнике. Пектины и камеди являются распространенными типами растворимых волокон, а хорошие источники пищи включают овсяные отруби, ячмень, орехи, семена, бобы, чечевицу, горох и некоторые фрукты и овощи. (Добавки с клетчаткой подорожника, такие как Metamucil, состоят в основном из растворимой клетчатки, поэтому, если вы когда-либо размешивали ложку этого вещества в стакане воды, вы видели вязкую консистенцию, характерную для растворимой клетчатки.)
- Нерастворимое волокно — Эти волокна обычно не растворяются в воде и не являются вязкими. Некоторые из них ферментируются бактериями в толстом кишечнике, но в меньшей степени, чем растворимые волокна. Нерастворимые волокна помогают предотвратить запоры, поскольку они создают более мягкий и объемный стул, который легче удалить. Лигнин, целлюлоза и гемицеллюлоза являются распространенными типами нерастворимых волокон, а источники пищи включают пшеничные отруби, овощи, фрукты и цельнозерновые продукты.
Поскольку клетчатка обеспечивает структуру растений, клетчатку можно найти во всех цельных растительных продуктах , включая цельнозерновые (например, овсянку, ячмень, рис и пшеницу), бобы, орехи, семена, цельные фрукты и овощи.
Рисунок 4.23. Чаша овсянки с черникой и семечками.
Это блюдо содержит клетчатку из овсяных хлопьев, черники и семян подсолнечника.
Когда пища рафинирована, части растения удаляются, и во время этого процесса теряются клетчатка и другие питательные вещества . Например, клетчатка теряется при переходе от целого свежего апельсина к апельсиновому соку. Целый апельсин содержит около 3 граммов клетчатки, тогда как стакан апельсинового сока практически не содержит клетчатки.Волокно также теряется при измельчении зерна. Об этом мы поговорим чуть позже.
Взгляните на список продуктов ниже, чтобы увидеть разнообразие продуктов, которые содержат пищевые волокна.
Продукты питания | Размер стандартной порции | калорий в стандартной порции | Пищевые волокна в стандартной порции (г) |
Измельченная пшеничная крупа, готовая к употреблению (разная) | 1–1 мкл | 155-220 | 5.0-9,0 |
Хлопья из пшеничных отрубей, готовые к употреблению (разные) | ¾ чашка | 90-98 | 4,9-5,5 |
Чечевица вареная | ½ стакана | 115 | 7,8 |
Черная фасоль, приготовленная | ½ стакана | 114 | 7.5 |
Жареные бобы консервированные | ½ стакана | 107 | 4,4 |
Авокадо | ½ стакана | 120 | 5,0 |
Груша сырая | 1 средний | 101 | 5,5 |
Груша сушеная | ¼ чашка | 118 | 3.4 |
Яблоко, с кожицей | 1 средний | 95 | 4,4 |
Малина | ½ стакана | 32 | 4,0 |
Овощная смесь, приготовленная из замороженных | ½ стакана | 59 | 4.0 |
Картофель, запеченный, с кожурой | 1 средний | 163 | 3,6 |
Тыквенные семечки, целые, жареные | 1 унция | 126 | 5,2 |
Семена чиа сушеные | 1 столовая ложка | 58 | 4.1 |
Ядра подсолнечника, обжаренные в сухом виде | 1 унция | 165 | 3,1 |
Миндаль | 1 унция | 164 | 3,5 |
Крекеры ржаные вафельные простые | 2 вафли | 73 | 5.0 |
Булгур, приготовленный | ½ стакана | 76 | 4,1 |
Попкорн воздушный | 3 чашки | 93 | 3,5 |
Спагетти из цельной пшеницы, приготовленные | ½ стакана | 87 | 3.2 |
Лебеда, приготовленная | ½ стакана | 111 | 2,6 |
Таблица 4.4. Обычные продукты, перечисленные со стандартным размером порции, а также калорийность и клетчатка в стандартной порции.
Хотя вы можете получить клетчатку из добавок, цельные продукты являются лучшим источником, потому что клетчатка поставляется в упаковке с другими необходимыми питательными веществами и фитонутриентами.
Польза пищевых волокон для здоровьяДиета с высоким содержанием клетчатки имеет множество преимуществ, в том числе:
- Помогает предотвратить запор. Многие волокна (но в основном нерастворимые) помогают сделать стул более мягким и объемным, от которого потом легче избавиться.
- Помогает поддерживать здоровье пищеварительной системы и кишечника. Пищевые волокна способствуют здоровью пищеварительной системы благодаря своей роли в элиминации и ферментации, а также положительно влияют на микробиоту кишечника.Поскольку клетчатка обеспечивает более объемный стул, это помогает поддерживать мышцы пищеварительного тракта в тонусе и силе, что помогает предотвратить геморрой и дивертикулы.
- Снижает риск сердечно-сосудистых заболеваний. Было показано, что более высокое потребление клетчатки улучшает липиды крови за счет снижения общего холестерина, триглицеридов и холестерина низкой плотности («плохой холестерин», связанный с более высоким риском сердечно-сосудистых заболеваний), и увеличения холестерина высокой плотности («хорошего холестерина», связанного с с меньшим риском сердечно-сосудистых заболеваний).Более высокое потребление клетчатки также связано с более низким кровяным давлением и уменьшением воспаления.
- Снижает риск диабета 2 типа. Было показано, что более высокое потребление клетчатки (особенно вязких или растворимых волокон) замедляет переваривание и всасывание глюкозы, улучшая метаболизм глюкозы. Диета с высоким содержанием клетчатки также может снизить риск диабета за счет уменьшения воспаления.
- Снижает риск колоректального рака. Дополнительные доказательства подтверждают идею о том, что более высокое потребление клетчатки снижает риск колоректального рака, хотя исследователи не уверены, почему.Одна из гипотез состоит в том, что пищевые волокна сокращают время прохождения (время, необходимое для прохождения пищи через пищеварительный тракт), тем самым подвергая клетки желудочно-кишечного тракта воздействию канцерогенов из пищи в течение более короткого времени.
- Помогает поддерживать здоровую массу тела. Исследования показали взаимосвязь между повышенным потреблением пищевых волокон и более низкой массой тела. Механизмы этого неясны, но, возможно, продукты с высоким содержанием клетчатки более сытны и, следовательно, дольше удовлетворяют людей с меньшим количеством калорий.Продукты с высоким содержанием клетчатки также, как правило, более богаты питательными веществами по сравнению со многими обработанными продуктами, которые более калорийны.
Перед уборкой все зерна представляют собой цельнозерновые. Они содержат все семя (или ядро) растения. Это семя состоит из трех съедобных частей: отрубей, зародыша и эндосперма. Семя также покрыто несъедобной шелухой, которая защищает семя.
Рисунок 4.24. Пшеница растет в поле.
- отруби — это внешняя оболочка семени. Он содержит антиоксиданты, витамины группы В и клетчатку.
- Эндосперм — безусловно, самая большая часть семян и обеспечивает энергию в виде крахмала для поддержки воспроизводства. Он также содержит белок и небольшое количество витаминов и минералов.
- Зародыш — это зародыш семени — часть, которая может прорасти в новое растение.Он содержит витамины группы B, белок, минералы, такие как цинк и магний, и полезные жиры.
Рисунок 4.25. Анатомия зерна пшеницы
включает отруби, эндосперм и зародыш.
Диетические рекомендации для американцев определяют цельнозерновые и рафинированные зерна следующим образом:
“ Цельнозерновые —Зерновые и зерновые продукты, полученные из цельного зерна зерна, обычно называемого ядром, которое состоит из отрубей, зародыша и эндосперма.Если ядро треснуло, раздавлено или расслоено, оно должно сохранять те же относительные пропорции отрубей, зародышей и эндосперма, что и исходное зерно, чтобы его можно было назвать цельным зерном. Многие, но не все цельнозерновые продукты также являются источниками пищевых волокон ».
Цельнозерновые продукты включают такие продукты, как ячмень, кукуруза (цельнозерновая мука и попкорн), овес (включая овсянку), рожь и пшеницу. (Более полный список цельнозерновых продуктов можно найти в Совете по цельнозерновым продуктам.)
“ Очищенные зерна —Зерновые и зерновые продукты с удаленными отрубями и зародышами; любой зерновой продукт, не являющийся цельнозерновым.Многие очищенные зерна содержат мало клетчатки, но обогащены тиамином, рибофлавином, ниацином и железом и обогащены фолиевой кислотой ».
Рафинированное зерно включает такие продукты, как белый рис и белая мука. По данным Совета по цельнозерновым продуктам, «переработка зерна удаляет около четверти белка из зерна и от половины до двух третей или более десятка питательных веществ, оставляя зерно лишь тенью от его первоначального состояния».
Рафинированное зерно часто обогащено витаминами и минералами, что означает, что некоторые питательные вещества, потерянные в процессе очистки, возвращаются обратно после обработки.Однако многие витамины и минералы не добавляются обратно, как и клетчатка, белок и полезные жиры, содержащиеся в цельнозерновых продуктах. В таблице ниже вы можете увидеть различия в основных питательных веществах между цельнозерновой мукой, очищенной пшеничной мукой и обогащенной пшеничной мукой.
Рисунок 4.26. Содержание питательных веществ в очищенной пшенице и обогащенной пшенице по сравнению с цельнозерновой мукой.
Поскольку цельные зерна обладают большей плотностью питательных веществ, MyPlate и диетические рекомендации рекомендуют, чтобы по крайней мере половина наших злаков была цельнозерновой.Тем не менее, текущие данные показывают, что, хотя большинство американцев в целом едят достаточно злаков, они едят слишком много очищенных злаков и недостаточно цельного зерна, как показано на рисунке из Руководства по питанию:
Рисунок 4.27. Среднее цельное и очищенное зерно Потребление в эквивалентах унций в день по возрастным и половым группам, по сравнению с диапазоном рекомендуемого суточного потребления цельного зерна и предельными значениями для очищенного зерна.
Искать цельнозерновые продукты в продуктовом магазине может быть непросто, поскольку маркировка на лицевой стороне упаковки предназначена для маркетинга и продажи продуктов.Такие слова, как «цельнозерновой» и «мультизерновой» на передней стороне упаковки создают впечатление, что продукт представляет собой цельнозерновой продукт, хотя на самом деле цельнозерновых продуктов может быть очень мало.
Цвет хлеба тоже может быть обманчивым. Продукты из рафинированного зерна могут иметь карамельный цвет, чтобы они больше походили на цельнозерновые.
Чтобы определить, является ли продукт хорошим источником цельного зерна, лучше всего посмотреть список ингредиентов на панели «Пищевая ценность». В ингредиентах должно быть указано цельное зерно в качестве первого ингредиента (например,г., 100% цельнозерновой), и за ним не должно следовать кучка очищенных зерен, таких как обогащенная пшеничная мука.
Знакомство с названием цельнозерновых продуктов поможет вам их идентифицировать. Общие разновидности включают пшеницу, ячмень, коричневый рис, гречку, кукурузу, рожь, овес и дикий рис. Менее известные разновидности включают теф, амарант, просо, киноа, черный рис, черный ячмень и полбу.
Большинство, но не все цельнозерновые продукты являются хорошим источником клетчатки, и это одно из преимуществ выбора цельного зерна.Имейте в виду, что некоторые продукты, например пшеничные отруби, инулин или целлюлоза, добавляют дополнительную клетчатку в качестве отдельного ингредиента. Это увеличивает количество граммов клетчатки на этикетке Nutrition Facts и может сделать продукт хорошим источником клетчатки, но это не значит, что это хороший источник цельного зерна. Фактически, это может быть продукт, сделанный в основном из очищенных зерен, поэтому в нем все равно будут отсутствовать другие питательные вещества, которые содержатся в цельнозерновых продуктах, и они могут не иметь такой же пользы для здоровья. Таким образом, просто взглянуть на клетчатку на этикетке «Пищевая ценность» — не лучший показатель того, сделан ли продукт из цельного зерна.
Кроме того, некоторые продукты на 100% состоят из цельной пшеницы, но не являются хорошим источником клетчатки из-за небольшого размера порции. Ярлык хлеба ниже является примером этого. Первый ингредиент — это «цельнозерновая мука каменного помола», в которой не указана рафинированная мука, но она все еще содержит только 2 г клетчатки и 9% суточной нормы. Но, конечно, это по-прежнему способствует дневному потреблению клетчатки, и если вы сделаете себе бутерброд с двумя ломтиками хлеба, это обеспечит 18% дневной нормы.
Рисунок 4.28. Пример 100% цельнозернового хлеба с пищевой ценностью и списком ингредиентов.
ВИДЕО: «Чтение этикеток и цельное зерно», Тамберли Пауэлл, YouTube (24 сентября 2018 г.), 7:46 минут.
гибридных волокон превращаются в разные типы волокон в созревающих мышцах мышей — FullText — Cells Tissues Organs 2013, Vol. 198, № 3
Абстрактные
Роль гибридных волокон в качестве промежуточных продуктов в превращениях типов волокон до конца не изучена.В некоторых случаях гибриды представляют собой явно переходные волокна, переходящие от одного типа к другому, но в других они представляют собой фенотипически стабильные волокна в нормальных мышцах. В текущем исследовании нашей целью было понять судьбу гибридных волокон в переходах между типами волокон, которые происходят во время созревания мышц. Предыдущие исследования показали высокую долю гибридных волокон во время постнатального развития, но лишь немногие проследили судьбу этих волокон после отлучения от груди. Мы количественно определили пропорции гибридных волокон в трех различных скелетных мышцах мышей от только что отлученных от груди до 6-месячных мышей.Гибридные волокна были очень распространены в мышцах brachioradialis (BR) и tibialis anterior (TA) недавно отлученных от груди мышей, где они составляли 50 и 40% волокон соответственно. Эти гибриды неуклонно снижались примерно до 15-30% в течение следующих нескольких месяцев. В камбаловидной мышце доля гибридов не изменилась, но составляла примерно 20% волокон. Уменьшение гибридов IIX / IIB произошло в результате различных процессов в BR и TA. В BR уменьшение совпало с увеличением волокон типа IIX.В ТА количество гибридов IIX / IIB обратно коррелировало с долей волокон IIB. Эти закономерности показывают, что роль гибридных волокон как промежуточных звеньев в развитии мышц является сложной. Некоторые гибридные волокна в созревающих мышцах представляют собой переходные типы волокон, в то время как другие фенотипически стабильны. Более того, судьба переходных волокон может отличаться среди сходных типов волокон в разных мышцах.
© 2013 S. Karger AG, Базель
Введение
Скелетные мышцы состоят из сотен и тысяч отдельных мышечных волокон, которые существуют в нескольких типах, что определяется экспрессией альтернативных изоформ тяжелой цепи миозина (MHC) и других миофибриллярных белков [Schiaffino and Reggiani, 2011 ].В мышцах конечностей мелких млекопитающих экспрессируются четыре изоформы MHC и обеспечивают континуум скоростей сокращения от самой медленной до самой быстрой в следующем порядке: I → IIA → IIX → IIB [Schiaffino and Reggiani, 1996, 2011]. За последние несколько десятилетий мы стали лучше понимать разнообразие и сложность отдельных типов волокон. Типы волокон, которые когда-то определялись просто как «быстрые» или «медленные», на самом деле являются частью континуума типов волокон, которые трудно классифицировать по точным категориям [Staron and Pette, 1993; Caiozzo et al., 2003; Медлер и Майклес, 2003; Медлер и др., 2004; Zhang et al., 2010; Скьяффино, Реджиани, 2011]. Во время развития волокна претерпевают основные переходы в экспрессии изоформ MHC, при этом изоформы развития обмениваются на взрослые MHCs [Di Maso et al., 2000; Agbulut et al., 2003; Либер, 2010]. Даже полностью дифференцированные скелетные мышцы взрослых сохраняют способность адаптироваться к множеству внешних стимулов благодаря фенотипической пластичности [Pette and Staron, 2001; Скьяффино, Реджиани, 2011].В самых драматических случаях отдельные волокна могут полностью трансформироваться из одного типа волокна в другой. Этот сложный процесс требует значительного сдвига в экспрессии MHC и других миофибриллярных белков, а также селективного протеолиза и удаления существующих изоформ. Во время этого процесса переключения типа волокна волокна временно становятся гибридами, коэкспрессируя две или более изоформ MHC в одном и том же волокне.
Смена типов волокон особенно важна в периоды дифференцировки и развития мышц.На раннем этапе эмбриональные и неонатальные MHCs экспрессируются за исключением взрослых изоформ, и эти изоформы продолжают экспрессироваться вместе со взрослыми MHCs в течение первых недель после рождения [Di Maso et al., 2000; Agbulut et al., 2003]. Взрослые изоформы MHC присутствуют в скелетных мышцах к моменту рождения, но переходы в типе волокон продолжаются по мере роста и созревания мышц [Wigston and English, 1992; Agbulut et al., 2003]. В этот постнатальный период часто встречаются гибридные волокна, содержащие смеси онтогенетических и взрослых изоформ MHC [Wigston and English, 1992; Di Maso et al., 2000]. В мышцах мышей экспрессия изоформ MHC в процессе развития заканчивается к 21 дню постнатального развития, но некоторые сдвиги в пропорциях изоформ MHC взрослых продолжаются в течение нескольких месяцев [Agbulut et al., 2003]. В камбаловидной мышце крысы (SOL) гибриды, экспрессирующие как быстрые, так и медленные MHCs, значительно уменьшаются в течение первого года развития, так как большинство этих волокон трансформируются в чистые медленные волокна [Wigston and English, 1992]. В мышином SOL некоторый уровень трансформации происходит за тот же период времени, но гораздо менее драматичен [Wigston and English, 1992].
В то время как гибридные волокна могут указывать на такие переходы типа волокна [Talmadge et al., 1995, 1999; Ди Масо и др., 2000; Unguez et al., 2000; Williamson et al., 2000], во многих случаях гибридные волокна сохраняются как стабильные фенотипы волокон [Stephenson, 2001; Caiozzo et al., 2003; Асеведо и Риверо, 2006 год; Glaser et al., 2010]. В мышцах человека доля гибридных волокон непостоянна, но в среднем около 15-20% гибридных волокон является нормальным явлением [Andersen et al., 1999; Williamson et al., 2000; Андерсен, 2003]. В мышцах крыс гибридные волокна особенно распространены, при этом многие мышцы состоят на 50-75% из гибридных волокон [Caiozzo et al., 2003]. У мышей общее распределение типов волокон ниже, чем у крыс, с сильным перекосом в сторону самого быстрого типа волокон IIB. Тем не менее, некоторые мышцы мышей содержат 25-50% их гибридов [Glaser et al., 2010; Zhang et al., 2010]. В мышцах обоих этих грызунов наиболее распространенными типами гибридных волокон являются гибриды IIX / IIB, которые составляют примерно 70-90% всех гибридных волокон [Caiozzo et al., 2003; Glaser et al., 2010; Zhang et al., 2010].
В текущем исследовании мы использовали анализ отдельных волокон для точного определения гибридных типов волокон в трех мышцах после отлучения. Мы определили содержание изоформы MHC в отдельных волокнах brachioradialis (BR), tibialis anterior (TA) и SOL мышц от только что отлученных мышей от животных в возрасте приблизительно 6 месяцев. Наша цель состояла в том, чтобы определить, какую роль гибридные волокна играют в созревании выбранных мышей в течение нескольких недель после отъема.Наша гипотеза заключалась в том, что доля гибридных волокон будет снижаться, поскольку они превращаются в чистые типы волокон в процессе созревания скелетных мышц. BR и TA были выбраны, потому что мы ранее идентифицировали эти мышцы как экспрессирующие высокие пропорции гибридных волокон [Zhang et al., 2010]. SOL был выбран потому, что он содержит популяцию гибридов, коэкспрессирующих изоформы MHC типа I и IIA, и был в центре внимания предыдущих исследований мышечного развития мышей [Wigston and English, 1992; Agbulut et al., 2003]. Вкратце, мы обнаружили, что доля гибридных волокон значительно снизилась в течение первых нескольких недель после отъема в мышцах BR и TA, но что доля гибридов была стабильной в SOL в течение того же периода времени. Гибриды IIX / IIB, которые были потеряны во время созревания мышц, имели разные судьбы в мышцах BR и TA. В BR большинство переходных гибридов были преобразованы в волокна IIX, тогда как в TA гибридные волокна преимущественно переходили в волокна IIB.
Материалы и методы
Животные
Мышей (C57BL / 6J) были приобретены в лаборатории Джексона (Бар-Харбор, штат Мичиган, США) и содержались в соответствии с утвержденным IACUC протоколом в Университете Буффало. Их содержали в клетках с 12-часовым 12-часовым циклом свет-темнота и обеспечивали пищей и водой ad libitum. Пометы содержались со своими матерями до достижения 21-дневного возраста, когда они были разделены с однополыми однопометниками в их собственные клетки. Мышей забивали в разном возрасте от 23 до 210 дней.Волокна собирали у 12 мышей самцов или самок.
Анализ отдельных волокон
Мышей умерщвляли воздействием CO 2 и немедленно собирали мышцы. Целые мышцы переносили в ледяной расслабляющий раствор (50% глицерин, 2 мМ EGTA, 1 мМ MgCl 2 , 4 мМ АТФ, 10 мМ имидазол и 100 мМ KCl) и хранили при -20 ° C. Отдельные волокна выделяли в холодном расслабляющем растворе с помощью тонких пинцетов под стереомикроскопом. В этом исследовании было проанализировано в общей сложности 2587 волокон (818 SOL, 723 BR и 1046 TA), при этом от каждого животного было отобрано примерно 100 волокон.Каждое волокно помещали в микроцентрифужную пробирку, содержащую ~ 30 мкл буфера для образцов, содержащего 8 М мочевину, 2 М тиомочевину, 50 мМ трис-основания, 75 мМ дитиотреитол, 3% додецилсульфат натрия (SDS) и 0,004% бромфенолового синего, pH 6,8 [ Blough et al., 1996; Zhang et al., 2010]. Образцы нагревали до 65 ° C в течение 15 мин и наносили 10 мкл каждого образца на гель. Электрофорез в SDS-полиакриламидном геле (SDS-PAGE) использовали с Hoeffer SE 600 для разделения отдельных изоформ MHC. Рассасывающиеся гели состояли из 12% акриламида (200: 1 акриламид: метилен- бис- акриламид), 12% глицерина, 0,1%.675 М трис-основание (pH 8,8) и 0,1% SDS. Укладываемые гели состояли из 4% акриламида (20: 1 акриламид: метилен- до -акриламид), 0,125 М трис-основания (pH 6,8) и 0,1% SDS. Рабочий буфер содержал 0,192 М глицина, 25 мМ трис-основания, 0,1% SDS и 0,08% 2-меркаптоэтанола. Гели запускали при постоянном токе 20 мА в течение примерно 41 ч при 8 ° C. В конце эксперимента гели фиксировали в 50% метаноле, который содержал формальдегид (0,037%), чтобы повысить чувствительность процедуры окрашивания серебром.Гели оставляли фиксироваться в этом растворе в течение ночи, а затем промывали в течение 1 ч в деионизированной воде перед окрашиванием серебром. Затем гели окрашивали аммиачным серебром [Wray et al., 1981; Medler et al., 2007].
Высушенные гели сканировали с высоким разрешением, и относительные доли изоформ MHC в отдельных волокнах определяли из цифровых изображений с использованием денситометрии (Image J 1.42q, NIH). Серийные разведения стандартов, содержащих известные пропорции изоформ MHC, использовали для установления того, что концентрации были линейной функцией плотности образца.
Сегментная экспрессия изоформ MHC
Отдельные волокна исследуемых мышц имели в среднем около 2–3 мм в длину после того, как они были отделены от всей мышцы. Мы разрезали эти отдельные волокна на 3 приблизительно равных длины и поместили каждый сегмент в отдельную пробирку с буфером для образцов мочевины. Для этого анализа мы сосредоточили внимание на TA и BR волокнах недавно отлученных от груди животных, потому что мы уже определили, что эти мышцы содержат большое количество гибридов IIX / IIB.Образцы разделяли с помощью SDS-PAGE, как описано выше, и определяли конкретные изоформы MHC в каждом сегменте. У идентифицированных гибридов мы использовали денситометрию для количественной оценки относительных пропорций изоформ IIX и IIB для выявления различий в экспрессии MHC по длине отдельных волокон.
Статистический анализ
Пропорции волокон в мышцах были проанализированы как функция возраста животного (количество дней от рождения) с использованием регрессионного анализа. Модели энергетической регрессии соответствовали данным, потому что спады гибридных волокон в TA и BR были явно криволинейными, и эти модели соответствуют данным лучше, чем простая линейная регрессия.Мы также использовали линейную регрессию для проверки других корреляций между пропорциями типов волокон (IIX, IIB и IIX / IIB), а также корреляций между типами волокон и возрастом или массой животных. Относительные пропорции изоформ IIX и IIB в гибридах IIX / IIB также анализировали как функцию возраста развития.
В дополнение к использованию регрессии для отслеживания пропорций гибридов как непрерывной функции возраста, мы также оценили различия в гибридных волокнах между недавно отлученными от груди мышцами и мышцами зрелых мышей.Средние доли конкретных изоформ MHC в этих мышцах сравнивали с помощью t-теста. Критические значения p для значимых различий были скорректированы с поправкой Бонферрони, чтобы установить экспериментальную частоту ошибок 0,05.
Результаты
Пропорции типов волокон в мышцах TA, BR и SOL
При наших параметрах геля и рабочих условиях четыре изоформы MHC мигрировали в порядке I> IIB> IIA> IIX (рис. 1). Эта картина отличается от результатов, полученных для MHC грызунов в других исследованиях [Talmadge and Roy, 1993; Di Maso et al., 2000; Caiozzo et al., 2003; Glaser et al., 2010] в том, что изоформы IIA и IIX имеют обратный порядок миграции. Как мы указывали ранее, результатом является лучшее разделение изоформ IIA и IIX [Zhang et al., 2010]. В мышцах TA и BR подавляющее большинство волокон были гибридами IIX, IIB или IIX / IIB (сумма этих типов волокон составила 97,4 ± 0,93% и 95,9 ± 1,38% в TA и BR соответственно; рис. ). Около 2% волокон были гибридами IIA / IIX в обеих мышцах, за которыми следовало небольшое количество волокон других типов (рис.2). В SOL доминировали волокна типа I и IIA, за которыми следовали гибриды I / IIA. Примерно 4% волокон были IIX, 4% были гибридами IIX / IIB, за ними следовало около 2% гибридов IIA / IIX (рис. 2).
Рис. 1
Типы мышиного волокна, идентифицированные с помощью SDS-PAGE белков из отдельных волокон. Используя описанные условия SDS-PAGE, изоформы MHC мигрируют как I> IIB> IIA> IIX. Показанные здесь одиночные волокна включают несколько различных типов волокон, в том числе несколько гибридов IIX / IIB.Типы выведенных волокон указаны над каждой полосой в геле.
Рис. 2
Общее распределение типов волокон в мышцах TA, BR и SOL, усредненное для всех мышей. a В TA преобладающей изоформой была IIB, за которой следовали IIX и IIX / IIB. b В BR волокна типов IIB, IIX и IIX / IIB присутствовали примерно в равных пропорциях. c В SOL изоформы типа I и IIA присутствовали в аналогичных пропорциях, и волокна I / IIA были преобладающим гибридным типом волокон.В каждой мышце присутствовало небольшое количество других типов волокон.
Пропорция гибридных волокон в зависимости от возраста мыши
Относительные пропорции гибридных волокон определяли примерно из 100 волокон от каждой взятой пробы мышцы (диапазон 71–170 волокон; в среднем 112 ± 26 волокон). Общее количество всех гибридов было объединено в индекс полиморфизма (общая доля всех гибридных волокон [Caiozzo et al., 2003]), который использовался для последующего статистического анализа. В мышцах TA и BR индекс полиморфизма был самым высоким у только что отлученных мышей, а затем снижался по криволинейной схеме, которая со временем выравнивалась.В качестве альтернативы возрасту мышей в качестве независимой переменной мы использовали массу животных, поскольку эта переменная является прямым индикатором роста животных (рис. 3). Для мышц TA и BR снижение количества гибридных волокон соответствовало силовой регрессии. Для ТА-мышц наклон регрессии был отрицательным и значимым при использовании возраста (p <0,0008) и массы (p <0,0033) в качестве независимой переменной. Для BR наклон не был значимым для возраста мышей (p <0,078), но был отрицательным и значимым для массы мышей (p <0.028). Для SOL не было значительного изменения пропорции гибридов в зависимости от возраста или массы мыши (рис. 3).
Рис. 3
Изменения пропорций гибридных волокон в зависимости от массы тела мыши. a TA-мышца показала значительное криволинейное уменьшение гибридных волокон с увеличенной массой (p <0,0033). b BR-мышца аналогичным образом показала криволинейное уменьшение пропорций гибридных волокон с ростом (p <0,028). c Пропорции гибридных волокон в мышце SOL со временем оставались стабильными и составляли 15-20%.
Судьба гибридных волокон в мышцах TA и BR
Учитывая значительное сокращение гибридных волокон по мере созревания в мышцах TA и BR, возникает актуальный вопрос: что случилось с этими волокнами? Так как подавляющее большинство гибридов в этих мышцах были IIX / IIB, мы сосредоточили внимание на различиях в пропорциях этих волокон у только что отлученных от матери мышей (рис. 4). В ТА-мышцах доля гибридов IIX / IIB была значительно ниже у зрелых мышей (34,9% у только что отлученных от груди мышей vs.17,7% у зрелых мышей; р <0,008). Совпадая с этим различием, волокна IIB составляли 60,7% у зрелых мышей по сравнению с 43,3% у только что отлученных мышей, но эта разница не была статистически значимой (рис. 4a; p <0,04; критический p = 0,025 с поправкой на Бонферрони). Однако линейный регрессионный анализ показал, что пропорции гибридных волокон IIB и IIX / IIB обратно коррелировали друг с другом в TA (r 2 = 0,522; p <0,028). Между гибридами IIX и IIX / IIB корреляции не наблюдалось (r 2 = 0.02; р> 0,68). В мышцах BR доля гибридов IIX / IIB также была значительно ниже у зрелых мышей (46,3% у только что отлученных мышей против 26% у зрелых мышей; p <0,01). В мышцах BR доля волокон IIX была значительно выше у зрелых мышей (рис. 4b; 38% у зрелых мышей против 18,9% у только что отлученных от груди мышей; p <0,005), в то время как доля волокон IIB была стабильной. Общий пул волокон IIX, IIB и IIX / IIB составлял 97,4 и 95,9% волокон в TA и BR, соответственно.Эти пропорции не изменились с созреванием (t-тесты, сравнивающие недавно отлученных от взрослых мышей: p> 0,99 и p> 0,98 для TA и BR, соответственно). Эти тенденции предполагают, что гибриды IIX / IIB, потерянные во время созревания, преимущественно преобразовывались в волокна IIB в мышцах TA, но в основном трансформировались в волокна IIX в BR (рис. 5). В обеих мышцах значительная доля стабильных гибридов (~ 20-25%) все еще присутствовала у самых старых исследованных мышей.
Фиг.4
Пропорции типов волокон у только что отлученных от груди мышей по сравнению с молодыми взрослыми мышами. a В мышцах ТА гибриды IIX / IIB значительно выше у только что отлученных мышей, чем у молодых взрослых мышей (p <0,008). Разница в волокнах IIB между только что отлученными от груди и молодыми взрослыми мышами не была статистически значимой (p <0,04). b В мышцах BR гибридов IIX / IIB было значительно меньше у молодых взрослых мышей по сравнению с недавно отлученными от груди мышами (p <0,010). Это меньшее количество гибридов совпадало со значительно более высокой долей чистых волокон IIX у старых мышей (p <0.005).
Рис. 5
Предполагаемая роль гибридов IIX / IIB в переходах типа волокна. Пул гибридов IIX / IIB снижается по мере созревания как в TA, так и в BR. Примерно половина гибридных волокон сохраняется как фенотипически стабильные гибриды, а примерно половина трансформируется в волокна либо IIB (TA), либо IIX (BR). Незначительное количество гибридных волокон также, вероятно, трансформируется в IIX в TA и в IIB в BR.
Пропорции изоформ IIX и IIB в гибридах IIX / IIB
В предыдущем исследовании мы определили, что гибриды IIX / IIB образуют континуум типов волокон в мышцах TA и BR мыши [Zhang et al., 2010]. В текущем исследовании нас интересовало, изменились ли эти пропорции во время созревания мышц, поскольку гибридные волокна превращались в чистые волокна IIX или IIB. В целом, мы снова наблюдали континуум типов волокон, от преимущественно содержащих IIX гибридов до преимущественно содержащих IIB гибридов. В среднем было равное соотношение изоформ IIX и IIB, и они были нормально распределены (рис. 6). В мышцах BR средняя доля изоформы IIX, содержащейся в этих гибридах, значительно увеличивалась в зависимости от возраста мыши (p <0.02), что, возможно, указывает на сдвиг в сторону волокна типа IIX (данные не показаны). Однако доля изоформы IIX все еще была близка к 50% (диапазон 44-60%). Не наблюдалось значительного сдвига в пропорциях изоформ MHC у гибридов IIX / IIB от TA.
Рис. 6
Относительная доля изоформы IIX в отдельных гибридных волокнах IIX / IIB в мышцах BR ( a ) и TA ( b ). Более темные столбцы указывают на большую долю IIX MHC, а более светлые столбцы соответствуют большему количеству IIB.В целом пропорции этих изоформ распределены нормально, при этом самая большая группа гибридов содержит примерно равные количества каждой изоформы.
Распределение изоформы MHC по длине одиночных волокон
Мы разделили отдельные волокна TA и BR мышей только что отлученных от груди мышей на три равных сегмента (конец, середина и конец). Затем определяли содержание изоформы MHC для каждого сегмента с использованием описанной процедуры SDS-PAGE. Мы наблюдали четыре модели содержания MHC в этих волокнах: (1) чистые волокна IIX (рис.7a), (2) чистые волокна IIB (рис. 7b), (3) симметричные гибриды IIX / IIB с аналогичными пропорциями в каждом сегменте (рис. 7c) и (4) асимметричные гибриды IIX / IIB, где пропорции IIX и Изоформы IIB различались по длине волокна (рис. 7e-g). Из 48 исследованных волокон 23% были чистыми IIX, 42% были чистыми IIB, 10% были симметричными гибридами IIX / IIB и 25% были асимметричными гибридами IIX / IIB. Мы количественно оценили относительные пропорции изоформ IIX и IIB в этих сегментах одиночных асимметричных гибридных волокон и обнаружили, что большая часть изоформы IIX была сконцентрирована на одном конце (рис.8). Как правило, наблюдалась градация от большего содержания IIX к одному концу и большей части IIB к другому (рис. 8).
Рис. 7
Пропорции изоформ IIX и IIB в сегментах волокна по длине отдельных волокон. Волокна были разделены на три сегмента [два конца (E и E ‘) и середина (M)]. В чистых волокнах IIX (A) или IIB (B) экспрессия MHC была однородной среди различных участков волокна. Точно так же некоторые из гибридов IIX / IIB демонстрируют симметричные смеси обеих изоформ по длине волокна (C).У 65% гибридов IIX / IIB мы наблюдали асимметричное распределение присутствующих изоформ MHC (D-G).
Рис. 8
Существуют значительные различия в пропорциях MHC среди сегментов одиночных волокон. Относительные пропорции изоформ IIX и IIB из сегментов отдельных волокон (см. Рис. 7) были количественно определены с помощью денситометрии. Среди асимметричных гибридов IIX / IIB значительная концентрация MHC типа IIX была расположена на одном конце отдельных волокон (n = 12 волокон; p <0,0014).
Обсуждение
Переходы типов волокон с созреванием
В текущем исследовании анализ содержания изоформы МНС в отдельных волокнах показал, что в мышцах молодых мышей наблюдаются значительные изменения типа волокон задолго до отлучения от груди.В мышцах BR и TA доля гибридных волокон была близка к 50% вскоре после отъема, но снизилась примерно до половины этого значения в течение следующих нескольких недель и месяцев (рис. 3). В ТА-мышце снижение у гибридов IIX / IIB происходило синхронно с увеличением содержания IIB, тогда как в мышце BR сокращение волокон IIX / IIB сопровождалось увеличением волокон IIX. Наша интерпретация этих данных состоит в том, что около половины гибридов IIX / IIB сохранялись как гибриды в обеих мышцах, но другая половина была преобразована в волокна IIX в BR, в то время как аналогичные пропорции были преобразованы в волокна IIB в TA (рис.5). Эти закономерности показывают, что судьба гибридных волокон при переходах между типами волокон может быть сложной, поскольку одни и те же типы гибридных волокон (IIX / IIB) в разных мышцах могут быть преобразованы в отдельные «чистые» типы волокон (IIX или IIB) в течение одного и того же периода. стадии созревания. Однако не все гибриды в трех исследованных мышцах были переходными, так как доля гибридов в SOL составляла стабильно 20% волокон в мышцах мышей всех возрастов, а 20-25% всех волокон были гибридами в мышцах. BR и TA в возрасте 6 месяцев (рис.3, 4). Эти устойчивые популяции гибридных волокон в мышцах мышей согласуются с обычным появлением гибридных волокон в нормальных мышцах различных видов [Unguez et al., 2000; Lutz et al., 2001; Стивенсон, 2001; Caiozzo et al., 2003; Медлер и др., 2004; Strbenc et al., 2006].
Точная роль гибридных волокон в пластичности скелетных мышц до конца не изучена [Stephenson, 2001; Caiozzo, 2002; Caiozzo et al., 2003], но наши результаты показывают, что они действительно играют роль промежуточных волокон во время переходов типа волокна (рис.5). Любые волокна, испытывающие изменения в экспрессии MHC, должны стать гибридами на какое-то время, поскольку устаревшие изоформы удаляются, а новые включаются в миофибриллы. Эта роль особенно важна во время раннего развития мышц и последующего постнатального созревания, когда происходят драматические сдвиги в типах волокон [Di Maso et al., 2000; Unguez et al., 2000; Strbenc et al., 2006]. Во время внутриутробного развития волокна скелетных мышц экспрессируют только эмбриональную изоформу MHC или в комбинации с другими изоформами [Di Maso et al., 2000; Agbulut et al., 2003; Либер, 2010]. Затем эмбриональный MHC заменяется неонатальным MHC, который затем экспрессируется вместе с изоформами взрослых [Di Maso et al., 2000; Agbulut et al., 2003; Либер, 2010]. К моменту рождения большинство мышечных волокон экспрессируют некоторые уровни эмбриональных и неонатальных изоформ в различных комбинациях друг с другом или со взрослыми изоформами [Di Maso et al., 2000; Unguez et al., 2000; Стрбенц и др., 2006; Wank et al., 2006]. В течение последующих недель после рождения эти изоформы развития быстро теряются, и экспрессия изоформ МНС у взрослых резко возрастает.В мышцах мышей развивающиеся изоформы MHC почти полностью отсутствуют в мышцах ко времени отлучения от груди (день 21) [Agbulut et al., 2003]. Однако наши результаты демонстрируют, что относительные пропорции этих взрослых изоформ MHC продолжают изменяться в течение следующих нескольких недель или месяцев в мышцах мышей. Эта модель непрерывного постепенного перехода к зрелым типам волокон согласуется с аналогичными изменениями в мышцах других видов [Wigston and English, 1992; Unguez et al., 2000; Стрбенц и др., 2006; Medler et al., 2007].
Первые несколько недель после рождения представляют собой период основного роста и дифференциации скелетных мышц [Agbulut et al., 2003; Ванк и др., 2006; Гохин и др., 2008; White et al., 2010]. Эти изменения многогранны и драматичны, затрагивая не только тип волокна, но также изменения мышечной массы, метаболических ферментов, удельного напряжения и связи возбуждения-сокращения [Wank et al., 2006; Гохин и др., 2008; Гудман и др., 2008; White et al., 2010]. Поскольку количество волокон практически фиксировано при рождении, увеличение мышечной массы и изменение типа мышечных волокон должно осуществляться за счет ремоделирования существующих волокон. Увеличение массы происходит по мере того, как большая плотность миофибрилл упаковывается в волокна, и когда волокна растут в длину [Gokhin et al., 2008; Гудман и др., 2008; White et al., 2010]. Увеличиваются не только волокна в размере, но и удельное натяжение, создаваемое каждым волокном, также увеличивается из-за более плотной упаковки миофибрилл внутри каждого волокна [Gokhin et al., 2008; Goodman et al., 2008]. Эти изменения в массе волокна и миофибриллярной упаковке идут рука об руку с переходами типа волокон. Сдвиги в типе волокон требуют увеличения экспрессии и включения новых изоформ, а также удаления устаревших изоформ MHC. В мышцах мышей изоформы MHC взрослых экспрессируются к моменту отлучения от груди, но значительные изменения в экспрессии этих изоформ в отдельных волокнах продолжаются, поскольку они включают новые миофибриллы в свои растущие мышцы. Когда MHC в этих новых миофибриллах отличается от MHC в существующих миофибриллах, результатом является изменение типа волокна.
Сегментарные различия в типах волокон
В предыдущем исследовании мы сравнили содержание MHC в соседних сегментах волокон TA и BR-мышц и обнаружили, что асимметрии были обычным явлением [Zhang et al., 2010]. В текущем исследовании мы обнаружили, что 2/3 гибридов IIX / IIB от только что отлученных мышей обнаруживают асимметрию в содержании изоформы MHC по длине отдельных волокон (рис. 7, 8). Хотя общепринятая точка зрения состоит в том, что отдельные мышечные волокна состоят из волокон одного типа и, следовательно, сокращаются как скоординированная единица, существование асимметрии типов волокон ставит под сомнение это основное предположение.Эдман и др. [1985] обнаружили существенные различия в скорости укорочения среди сегментов в пределах отдельных мышечных волокон лягушки и предположили, что эти различия являются результатом дифференциальной экспрессии изоформ MHC в разных областях волокон. Эти региональные различия в содержании MHC в отдельных волокнах лягушки и их вклад в различия во внутриволоконных сократительных свойствах с тех пор были подтверждены [Edman et al., 1988; Lutz et al., 2001]. Совсем недавно в мышцах жаб in vivo были идентифицированы независимые образцы мышечного напряжения внутри отдельных пучков [Ahn et al., 2003]. Хотя функциональное значение региональных различий в сокращении мышц плохо изучено, мы можем предложить механизм их возникновения.
Мышечные волокна представляют собой огромные клетки, образованные слиянием отдельных миобластов, и часто имеют длину несколько миллиметров [Lieber, 2010]. Присутствие сотен отдельных ядер, наряду с ограничениями по диффузионному расстоянию, означает, что разные миоядерные домены могут содержать разные белковые комплексы друг от друга [Pavlath et al., 1989; Newlands et al., 1998; Allen et al., 1999]. Действительно, ряд исследований продемонстрировал, что экспрессия миозина действительно варьирует по длине в пределах определенных одиночных мышечных волокон [Staron and Pette, 1987; Эдман и др., 1988; Термин и др., 1989; Sakuma et al., 1995; Пойкер и Петт, 1997; Lutz et al., 2001; Zhang et al., 2010]. Хотя в настоящее время неизвестно, насколько распространена такая асимметрия в нормальных мышцах, вполне вероятно, что продольные различия в содержании MHC могут отражать динамические изменения в типе волокон.Например, сегментные различия в экспрессии MHC могут быть связаны с резким увеличением длины мышечных волокон, которое происходит во время фазы быстрого роста мышц после отлучения от груди [White et al., 2010]. В созревающих мышцах SOL крыс региональная асимметрия по типу волокон присутствует в отдельных волокнах в возрасте от 3 до 4 недель, но эти волокна затем становятся более однородными со временем [Sakuma et al., 1995]. В развивающихся мышцах птиц неонатальные MHC экспрессируются на растущих концах волокон, но взрослые MHC обнаруживаются в средних областях мышц [Rosser et al., 1995]. Мы обнаружили, что во многих случаях один конец волокна отличался по содержанию MHC по сравнению с остальной частью волокна (рис. 7, 8). Поскольку мышцы только что отлученных мышей быстро растут на концах [White et al., 2010], одно из объяснений этих различий заключается в том, что изоформа MHC, включенная в миофибриллы в областях роста, может отличаться от MHC в существующих областях волокон. . Необходима дальнейшая работа, чтобы лучше определить роль роста мышц в создании асимметрии волокон.
В заключение, мы обнаружили, что гибридные волокна IIX / IIB составляют примерно 40-50% волокон в TA и BR только что отлученных от груди мышей, но эти гибриды превращаются в чистые волокна типов IIX и IIB в течение последующих недель и месяцев.Следовательно, многие гибриды в этих мышцах являются переходными типами волокон в мышцах созревающих мышей. Многие из гибридов в этих мышцах демонстрируют выраженную асимметрию в содержании MHC по длине, паттерн, который поднимает ряд важных вопросов для регуляции типа мышечных волокон. Мы предполагаем, что региональные асимметрии могут возникать во время быстрого роста длины волокна, когда вновь включенная изоформа MHC может отличаться от существующих. Необходимы дальнейшие исследования, чтобы выяснить роль этих асимметрий MHC во время роста и развития мышц.
Благодарности
Эта работа была поддержана Национальным институтом артрита, опорно-двигательного аппарата и кожных заболеваний, грант RAR-053666A С. Медлеру. Мы искренне ценим проницательные комментарии двух анонимных рецензентов рукописи.
Список литературы
- Асеведо, Л.М., Дж.Л. Риверо (2006) Новое понимание типов волокон скелетных мышц у собак с особым акцентом на фенотипы гибридного миозина. Cell Tissue Res 323: 283-303.
- Agbulut, O., P. Noirez, F. Beaumont, G. Butler-Browne (2003) Изоформы тяжелой цепи миозина в постнатальном развитии мышц.Биологическая ячейка 95: 399-406.
- Ан, А.Н., Р.Дж. Монти, А.А. Biewener (2003) In vivo и in vitro неоднородность изменения длины сегмента в полуперепончатой мышце жабы. J. Physiol 549: 877-888.
- Аллен, Д.Л., Р. Р. Рой, В. Edgerton (1999) Миоядерные домены в адаптации и заболеваниях мышц. Мышечный нерв 22: 1350-1360.
- Андерсен, Дж. Л. (2003) Адаптация типа мышечных волокон в мышцах пожилого человека. Scand J Med Sci Sports 13: 40-47.
- Андерсен, Дж.L., G. Terzis, A. Kryger (1999) Повышение степени коэкспрессии изоформ тяжелой цепи миозина в волокнах скелетных мышц очень старых. Мышечный нерв 22: 449-454.
- Blough, E., E. Rennie, F. Zhang, P. Reiser (1996) Улучшенное электрофоретическое разделение и разрешение тяжелых цепей миозина в скелетных мышцах млекопитающих и птиц.Анал Биохим 233: 31-35.
- Кайоццо, В.Дж. (2002) Пластичность фенотипа скелетных мышц: механические последствия. Мышечный нерв 26: 740-768.
- Кайоццо, В.J., M.J. Baker, K. Huang, H. Chou, Y. Wu, K. Baldwin (2003) Полиморфизм тяжелой цепи миозина с одним волокном: сколько паттернов и какие пропорции? Am J Physiol Regul Integr Comp Physiol 285: R570-R580.
- Ди Мазо, Н.А., В.Дж. Кайоццо, К. Болдуин (2000) Полиморфизм тяжелой цепи миозина из одного волокна во время постнатального развития: модуляция гипотиреозом.Am J Physiol Regul Integr Comp Physiol 278: R1099-R1106.
- Эдман, К.А.П., К. Реджиани, С. Скьяффино, Г. те Кронни (1988) Максимальная скорость укорочения, связанная с изоформным составом миозина в скелетно-мышечных волокнах лягушки. J. Physiol 395: 679-694.
- Эдман К.А.П., К. Реджиани, Г. те Кронни (1985) Различия в максимальной скорости укорочения отдельных мышечных волокон лягушки. J. Physiol 365: 147-163.
- Глейзер, Б., G. You, M. Zhang, S. Medler (2010) На относительные пропорции гибридных волокон не влияют 6-недельные беговые упражнения в скелетных мышцах мышей. Exp Physiol 95: 211-221.
- Гохин Д.С., Гохин С. Уорд, С. Бремнер, Р.Л.Либер (2008) Количественный анализ функционального улучшения скелетных мышц новорожденных у мышей.J Exp Biol 211: 837-843.
- Гудман, К.А., Р. Блазев, Дж. Кемп, Г.М.М. Stephenson (2008) E-C сцепление и сократительные характеристики механически очищенных отдельных волокон молодых крыс во время быстрого роста и созревания. Арка Пфлюгерс 456: 1217-1228.
- Либер Р.Л. (2010) Структура, функция и пластичность скелетных мышц: физиологическая основа реабилитации. Балтимор, Липпинкотт Уильямс и Уилкинс.
- Лутц, Г.J., S.N. Бремнер, М.Дж.Баде, Р.Л. Либер (2001) Идентификация легких цепей миозина в скелетных мышцах Rana pipiens и паттерны их экспрессии вдоль отдельных волокон. J Exp Biol 204: 4237-4248.
- Медлер, С., Лилли Т., Д.Л. Mykles (2004) Полиморфизм волокон в скелетных мышцах американского лобстера, Homarus americanus : континуум между медленно сокращающимися (S-1) и медленно-тоническими (S-2) волокнами.J Exp Biol 207: 2755-2767.
- Медлер, С., Т. Лилли, Дж. Риль, Э. Малдер, Э. Чанг, Д.Л. Mykles (2007) Экспрессия миофибриллярных генов в дифференцирующихся мышцах когтей лобстера. J Exp Zool A Ecol Genet Physiol 307A: 281-295.
- Медлер, С., Д.Л. Mykles (2003) Анализ миофибриллярных белков и транскриптов в скелетных мышцах взрослых американских лобстеров Homarus americanus : переменная экспрессия миозинов, актина и тропонинов в быстрых, медленно сокращающихся и медленных тонических волокнах. J Exp Biol 206: 3557-3567.
- Ньюлендс, С., Л.К. Левитт, К.С. Робинсон, A.B.C. Карпф, В. Ходжсон, Р.П. Уэйд, Э.К. Хардеман (1998) Транскрипция происходит импульсами в мышечных волокнах. Genes Dev 12: 2748-2758.
- Павлат, Г.К., К. Рич, С.Г. Вебстер, Х.М. Blau (1989) Локализация мышечных генных продуктов в ядерных доменах.Природа 337: 570-573.
- Петте, Д., Р.С. Staron (2001) Переходы фенотипических профилей мышечных волокон. Histochem Cell Biol 115: 359-372.
- Пойкер, Х., D. Pette (1997) Количественный анализ мРНК тяжелой цепи миозина и изоформ белка в отдельных волокнах выявил выраженную неоднородность волокон в нормальных мышцах кролика. Eur J Biochem 247: 30-36.
- Россер, B.W.C., D.M. Вальдбиллиг, С. Лово, Дж.Д. Армстронг, Э.Bandman (1995) Экспрессия тяжелых цепей миозина в сужающихся концах волокон скелетных мышц. Анат Рек. 242: 462-470.
- Сакума, К., А. Ямагути, Х. Омори, С. Кацута (1995) Неоднородные изменения типов волокон в камбаловидной мышце развивающейся крысы. Eur J Appl Physiol Occup Physiol 70: 132-137.
- Schiaffino, S., C. Reggiani (1996) Молекулярное разнообразие миофибриллярных белков: регуляция генов и функциональное значение. Physiol Rev 76: 371-423.
- Скьяффино, С., C. Reggiani (2011) Типы волокон в скелетных мышцах млекопитающих. Physiol Rev 91: 1447-1531.
- Staron, R.S., D. Pette (1987) Неоднородная экспрессия миозина вдоль отдельных волокон хронически стимулированных и контралатеральных мышц передней большеберцовой мышцы кролика. Pflügers Arch 409: 67-73.
- Staron, R.S., D. Pette (1993) Континуум чистых и гибридных типов волокон на основе тяжелых цепей миозина в скелетных мышцах крыс. Гистохимия 100: 149-153.
- Стивенсон, Г.М.М. (2001) Гибридные волокна скелетных мышц: редкое или обычное явление? Clin Exp Pharmacol Physiol 28: 692-702.
- Strbenc, M., V. Smerdu, A. Pogacnik, G. Fazarinc (2006) Переходы изоформ тяжелой цепи миозина в скелетных мышцах собак во время постнатального роста.Дж. Анат 209: 149-163.
- Талмадж, Р.Дж., Р.Р. Рой (1993) Электрофоретическое разделение изоформ тяжелой цепи миозина скелетных мышц крысы. J Appl Physiol 75: 2337-2340.
- Талмадж, Р.Дж., Р. Р. Рой, В. Edgerton (1995). Выявление гибридных волокон тяжелой цепи миозина в камбаловидной мышце крыс после перерезки спинного мозга. J Appl Physiol 78: 1256-1265.
- Талмадж, Р.Дж., Р.Р. Рой, В. Edgerton (1999) Сохранение гибридных волокон в камбаловидной мышце крысы после перерезки спинного мозга.Анат Рек. 255: 188-201.
- Термин, А., Р.С. Staron, D. Pette (1989) Изменения изоформ тяжелой цепи миозина во время хронической низкочастотной стимуляции быстрых мышц задних конечностей крысы: исследование одного волокна. Eur J Biochem 186: 749-754.
- Унгес, Г.А., Р.Дж. Талмадж, Р.Р. Рой, Д. Далпонте, В. Edgerton (2000) Четкие переходы изоформ тяжелой цепи миозина в развитие медленных и быстрых мышц задних конечностей кошек. Клетки и ткани органов 167 : 138-152.
- Ванк, В., М.С. Фишер, Б. Вальтер, Р. Бауэр (2006) Рост мышц и тип волокон в мышцах задних конечностей во время постнатального развития у свиней.Клетки и ткани органов 182: 171-181.
- Уайт, Р. Б., А. С. Биеринкс, В.Ф. Ньокки, П.С. Заммит (2010) Динамика роста мышечных волокон во время постнатального развития мышей. BMC Dev Biol 10: 21.
- Вигстон, Д.ЧЕЛЮСТЬ. English (1992) Пропорции типа волокон в камбаловидной мышце млекопитающих во время постнатального развития. Дж. Neurobiol 23: 61-70.
- Уильямсон, Д.Л., М.П. Годар, Д.А. Портер, Д. Костилл, С. Trappe (2000) Прогрессивные тренировки с отягощениями снижают коэкспрессию тяжелых цепей миозина в отдельных мышечных волокнах у пожилых мужчин.J Appl Physiol 88: 627-633.
- Рэй, В., Т. Буликас, В.П. Рэй, Р. Хэнкок (1981) Окрашивание белков в полиакриламидных гелях серебром. Анальный Биохим 118: 197-203.
- Чжан, М.Y., W.J. Zhang, S. Medler (2010) Континуум гибридных волокон IIX / IIB в нормальных мышцах мыши: пропорции изоформ MHC и пространственное распределение внутри отдельных волокон. Am J Physiol Regul Integr Comp Physiol 299: 1582-1591.
Автор Контакты
Доктор Скотт Медлер
Департамент биологии, Государственный университет Нью-Йорка в Фредонии
205 Джуэтт Холл
Фредония, Нью-Йорк 14063 (США)
Электронная почта Скотта[email protected]
Подробности статьи / публикации
Предварительный просмотр первой страницы
Принято: 22 августа 2013 г.
Опубликована онлайн: 2 ноября 2013 г.
Дата выпуска: декабрь 2013 г.
Количество страниц для печати: 10
Количество фигур: 8
Количество столов: 0
ISSN: 1422-6405 (печатный)
eISSN: 1422-6421 (онлайн)
Для дополнительной информации: https: // www.karger.com/CTO
Авторские права / Дозировка препарата / Заявление об ограничении ответственности
Авторские права: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме и любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование или с помощью какой-либо системы хранения и поиска информации, без письменного разрешения издателя. .
Дозировка лекарств: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарств, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Однако ввиду продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю настоятельно рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендованным агентом является новый и / или редко применяемый препарат.
Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, причиненный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
Типы мышечных волокон: Учебник ортопедии Wheeless
— См .: Мышечное меню
— Два основных типа волокон: волокна типа I и типа II;
— у низших млекопитающих и других животных мышцы обычно полностью состоят из волокон типа I или типа II;
— мышца человека состоит из смеси волокон типа I и типа II;
— тип мышечного волокна зависит не от каких-либо внутренних свойств самого волокна, а от двигательного нейрона, снабжающего это конкретное волокно;
— все мышечные волокна, поставляемые одним конкретным аксоном, будут либо типа I, либо типа II;
— Тип I:
— есть существенные различия по типам волокна;
— мышечные волокна I типа богаты ферментами, необходимыми для окислительного метаболизма, и имеют более темный вид;
— при стимуляции у них медленное время сокращения или «подергивания»;
— эти волокна обладают повышенной устойчивостью к усталости;
— волокна типа I хорошо подходят для деятельности, связанной с физическими усилиями, требующими силы и выносливости, энергетический обмен которых зависит от окислительных процессов;
— Волокна типа II:
— волокна типа II получают энергию в результате гораздо более быстрого гликолитического процесса;
— поскольку запасы гликогена истощаются быстрее, чем запасы кислорода, волокна типа II менее подходят для непрерывных видов деятельности и больше подходят для быстрых переменных усилий;
— мышцы, наиболее привыкшие к медленной непрерывной работе, имеют меньший процент мышечных волокон типа II;
— волокна типа II могут быть более подвержены анатомическим изменениям вследствие изменения потребности в энергии, чем волокна типа I;
— волокна типа II имеют тенденцию быть меньше, чем волокна типа I у детей и взрослых, которые не выполняют интенсивных физических упражнений, хотя они увеличиваются в размере при
повторяющихся физических нагрузках на мышцы;
— волокна типа II подразделяются на тип IIa и тип IIb;
— волокна типа IIa содержат смесь гликолитических и окислительных ферментов и показывают промежуточное время сокращения;
— волокна типа IIb (быстрые гликолитические волокна) имеют самый большой размер двигательных единиц, имеют самую высокую скорость сокращения и наиболее подвержены утомлению;
— травма мышцы:
— мышечные волокна типа II более восприимчивы к травмам, чем волокна типа I.