Анатомия трапеции и шейных мышц — Упражнения — Фитнес
Спортсмену необходимо изучать мышцы спины для того, чтобы правильно тренировать их. Что представляют собой данные мышцы?
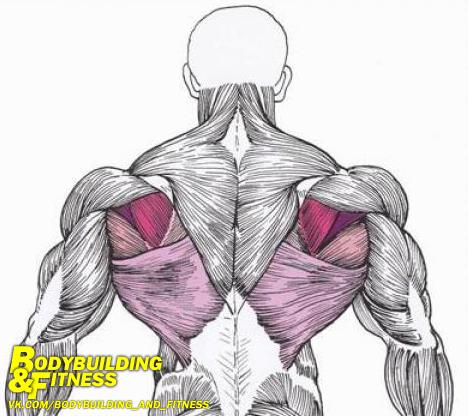
Мышцы трапеции и шейные мышцы прикрепляются к скелету плечевого пояса и состоят из двух слоев. Они занимают верхнее положение в спине и в области шеи. Данные мышцы напоминают фигуру в геометрии – трапецию. Именно поэтому многие атлеты именно так называют данные мышцы. Они разделяются на несколько частей:
— верхнюю;
— среднюю;
— нижнюю.
Мышцы имеют пучки сухожилия. Они представляют собой площадку в виде ромба. Мышцы сходятся в лопатке. Трапеция имеет достаточно большой размер.
Функции трапециевидной мышцы
Данная мышца выполняет множество функций, но в основном она отвечает за движение лопатки, ее поднятие и вращение руками. Функции:
— сокращение мышц способствует приближению лопатки к позвоночнику;
— сокращение верхнего и нижнего волокна способствует вращению костей;
— лопатка поднимает пучки в верхней части мышц;
— мышцы сокращаются с двух сторон и помогают разгибать шею, что помогает наклонять голову в разные стороны;
— данная мышца помогает поворачивать голову в другую сторону.
Как накачать трапециевидную мышцу?
Трапеция находится в спинном отделе наверху. По ее состоянию можно определить, занимается ли человек какими-либо физическими нагрузками. При тренировке трапециевидной мышцы начинает увеличиваться шея. Ее мышцы становятся мощнее, плотнее и рельефнее. Для того, чтобы получить мышцы спины больше, не требуется много усилий.
Есть некоторые специальные упражнения, помогающие получить данный эффект. Тренировать мускулатуру можно с помощью поднятия и опускания штанги или гантелей с определенным весом. Нижнюю часть спины тренируют за счет разведения и сведения лопаток вместе. Не рекомендуется спортсменам тренировать трапециевидную мышцу отдельно от всех. Это может повлечь сужению плеч. Необходимо все мышцы качать вместе для получения желаемого телосложения.
Синдром трапециевидной мышцы снять с помощью мануальной терапии в Москве
Синдром трапециевидной мышцы
Боль в трапециевидной мышце не всегда ограничивается только областью мышцы.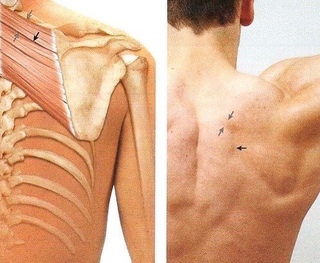
Переутомление и перегрузка открывают дорогу болезни. Установлено, что боль в трапециевидной мышце обусловлена триггерными точками. По мнению Трэвелл и Симонс — авторов книги о миофасциальном синдроме, триггерные точки трапециевидной мышцы встречаются гораздо чаще, чем у других мышц [Дж. Трэвелл и Д. Симонс «Миофасциальные боли и дисфункции». Том I. С. 353].
Но причина боли трапециевидной мышцы не всегда лежит только в физической плоскости. Эта мышца, как ни одна другая, подвержена влиянию эмоциональных факторов. Однако о психосоматике трапециевидной мышцы и миофасциальном синдроме мы поговорим чуть ниже, в разделе «Симптомы…», а сейчас — анатомия.
Анатомия трапециевидной мышцы
Анатомия трапециевидной мышцы предполагает деление мышцы на три части: верхнюю, среднюю и нижнюю. Верхнюю часть, обычно, называют трапециевидной мышцей шеи, а среднюю и нижнюю – трапециевидной мышцей спины. Но, сразу поясним, что это разделение не официальное – для документов, а разговорное – для удобства использования в речи. Вообще, трапециевидная мышца – это одна из наиболее крупных мышц.
Функции трапециевидной мышцы
Функции трапециевидной мышцы обеспечивают движения и статику плеча, лопатки и шеи. Например, мы задействуем эту мышцу, если хотим расправить плечи и выпрямить шею или, когда сводим лопатки вместе и запрокидываем голову или, когда двигаем плечами вверх-вниз и вперёд-назад. Во время ходьбы мы размахиваем руками, и мышца работает в динамике, а, если мы сидим за компьютером — в статике. И даже, когда мы просто стоим, опустив руки — мышца тоже работает, чтобы обеспечить антигравитационный эффект. Кстати, именно для того, чтобы снять напряжение и разгрузить трапециевидную мышцу, мы, машинально, складываем руки на груди или кладём их в карманы.
Говоря об антигравитационной функции трапециевидной мышцы, становится ясно, почему, работая за столом, нужно следить, чтобы локти не находились на весу — иначе вес рук будет вызывать перегрузку.
«Шейная» функция трапециевидной мышцы обеспечивает повороты и наклоны головы. Поэтому экран монитора и телевизора должны располагаться прямо перед нами. Это тоже предотвратит развитие боли и патологии. И, кстати, привычка придерживать телефон ухом, тоже служит причиной боли трапециевидной мышцы.
Симптомы синдрома трапециевидной мышцы
Симптомы трапециевидной мышцы носят, в основном, болевой характер, это связано с тем, что синдром трапециевидной мышцы является миофасциальным, по своей сути.
Миофасциальный синдром трапециевидной мышцы – это патология, при которой в мышечной ткани образуются небольшие болезненные участки – триггерные точки. Длительное время они могут никак не проявляться, оставаясь в латентном состоянии.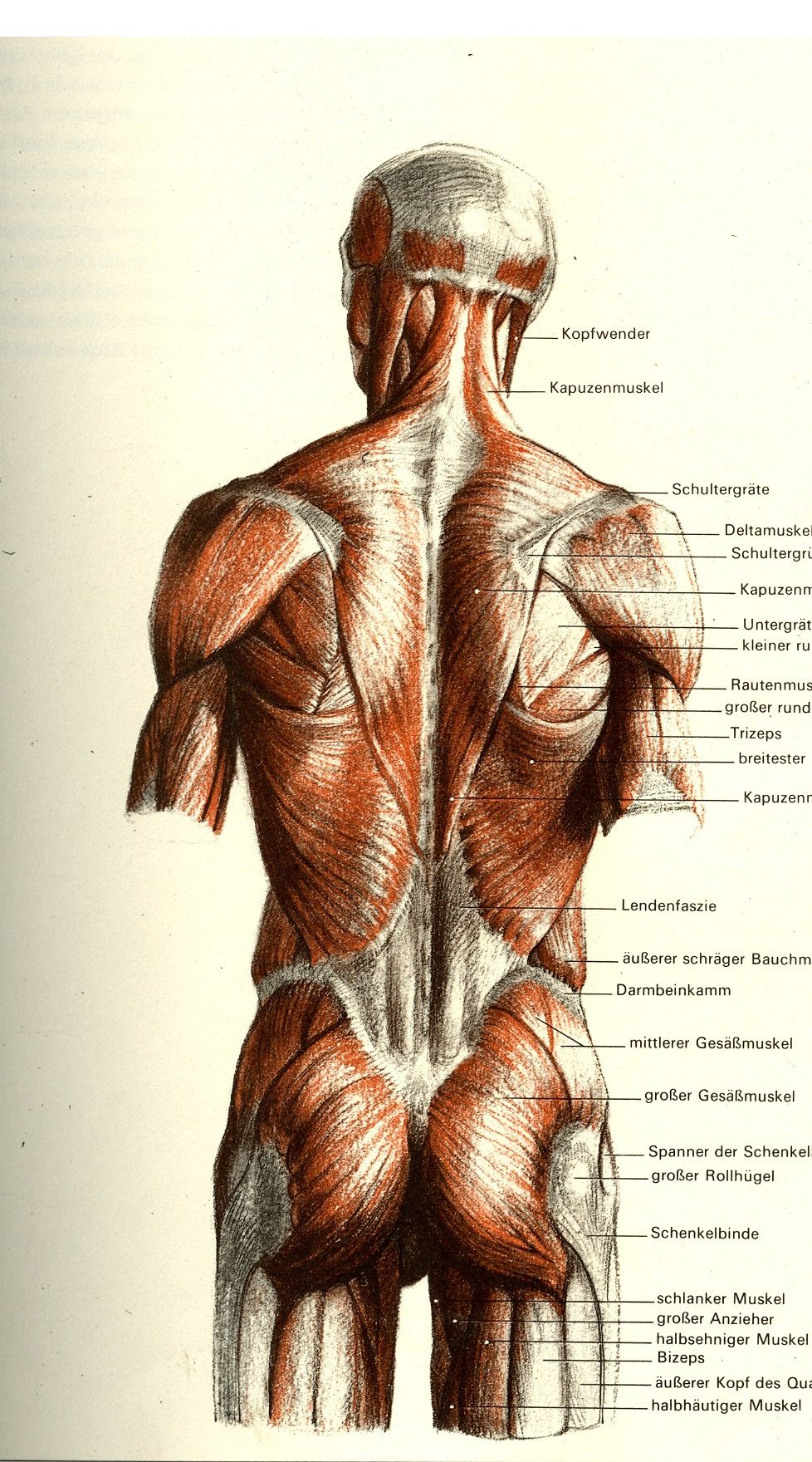 Но стоит их активировать, как тут же возникает боль. Фактором активации может стать неудобная поза, резкое движение, перегрузка, состояние голода, переохлаждение или стресс. Кстати, боль, сопряжённая со стрессом и эмоциями, это и есть психосоматика трапециевидной мышцы.
Но стоит их активировать, как тут же возникает боль. Фактором активации может стать неудобная поза, резкое движение, перегрузка, состояние голода, переохлаждение или стресс. Кстати, боль, сопряжённая со стрессом и эмоциями, это и есть психосоматика трапециевидной мышцы.
Механизм психосоматических реакций проще понять на примере животных. Черепаха, при опасности, втягивает голову в панцирь, а большинство других — просто, вжимают голову в плечи. Тем самым животные защищают своё самое уязвимое место — шею. Нам тоже, эволюция сохранила этот биологический рефлекс. Именно он, в ответ на стресс, вызывает у нас напряжение многих мышц, но, в первую очередь — трапециевидной. Нетрудно догадаться, что у тех людей, которые испытывают постоянную эмоциональную нагрузку, психосоматика трапециевидной мышцы будет только нарастать. Но, вернёмся к триггерным точкам.
Мы уже говорили, что в трапециевидной мышце триггерные точки возникают чаще, чем в других. При этом существует удивительная закономерность.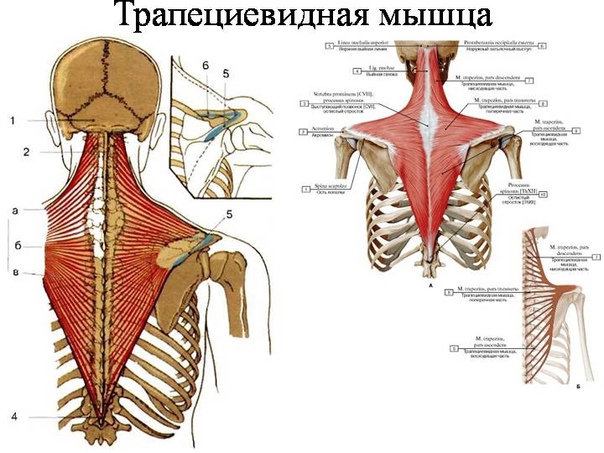
Кроме того, у абсолютного большинства людей точки возникают в одних и тех же местах; и таких мест – семь. Это — семь классических триггерных точек трапециевидной мышцы. При этом одни из них могут вызывать боль только в спине и шее, а другие — помимо спины и шеи – ещё и головную боль. Но, давайте перечислим все симптомы синдрома трапециевидной мышцы.
Перечисление симптомов трапециевидной мышцы начнём с шеи. Ведь синдром трапециевидной мышцы является основным источником шейных болей. Чаще всего, боль идёт по заднебоковой поверхности и доходит до черепа. С шеи боль может переходить на висок и боковую поверхность головы, а также — ощущаться позади глазницы или распространяться в затылок. Ещё миофасциальный синдром трапециевидной мышцы может вызвать отраженные боли в ушной раковине (но не в глубине уха), а также — рефлекторное головокружение и вегето-сосудистые кризы.
Следующий симптом трапециевидной мышцы – лицевая боль. Обычно она локализуется в углу нижней челюсти, области жевательных мышц и нижних зубов. Этот синдром хорошо знаком стоматологам.
Нередко боль от трапециевидной мышцы шеи активирует сателлитные триггерные точки, расположенные в других шейных мышцах. Это вызывает головную боль напряжения. Вообще, активация сателлитных точек весьма характерна для трапециевидной мышцы. Чаще всего это происходит с лестничными мышцами. А если активируются точки в мышце поднимающей лопатку и ременной мышце, то возникает симптом «неподвижной шеи», при котором невозможно повернуть голову.
Рассказывая о триггерных точках трапециевидной мышцы нужно обратить особое внимание на очень коварную точку, находящуюся внизу, между позвоночником и лопаткой. Коварство её заключается в том, что она способна вызывать повторное формирование триггерных точек наверху — в трапециевидной мышце шеи. На этой проблеме часто «спотыкаются» неопытные врачи. Устранив все триггеры в шее и верхней части спины, такой врач никак не может понять, почему боль не проходит. А причина, чаще всего, именно в этой точке.
Устранив все триггеры в шее и верхней части спины, такой врач никак не может понять, почему боль не проходит. А причина, чаще всего, именно в этой точке.
Также для синдрома трапециевидной мышцы характерны боли в области надплечий и верхушки плечевого сустава. И тогда, всё, что давит на плечи, причиняет особую боль и неудобства. Это и бретельки бюстгальтера, и тяжелая верхняя одежда, и сумка через плечо и т.д. Также нужно отметить, что в этой области расположена точка, вызывающая неприятные ощущения по типу «гусиной кожи». Ещё одним симптомом трапециевидной мышцы является жгучая боль в межлопаточной области или вдоль позвоночного края лопатки.
Вообще, нужно сказать, что многие люди искренне не понимают, почему у них болит трапециевидная мышца. Ведь, по их мнению, для этого нет никаких причин. Чтобы разъяснить этот вопрос, нужно сказать, что боль в трапециевидной мышце часто является следствием наших привычек. Ведь то, что мы считаем обычной привычкой, на поверку оказывается хронической микротравмой. Например, при синдроме трапециевидной мышцы симптомы могут появиться из-за привычки удерживать телефон ухом или спать на животе, повернув голову в одну и ту же сторону. Привычно располагая руки на вершине руля или сопровождая свою речь активной жестикуляцией, мы тоже вызываем перегрузку мышцы. Такая же незаметная перегрузка формирует симптомы трапециевидной мышцы из-за привычки постоянно откидывать волосы со лба резким движением головы. А подпирая руками подбородок, мы перегружаем трапециевидную мышцу спины. В общем, капля за каплей — вода камень точит. Так и хроническая микротравма — коварна своей незаметностью. Кстати, для этого и существует профилактика — чтобы вовремя устранять накопившиеся перегрузки.
Например, при синдроме трапециевидной мышцы симптомы могут появиться из-за привычки удерживать телефон ухом или спать на животе, повернув голову в одну и ту же сторону. Привычно располагая руки на вершине руля или сопровождая свою речь активной жестикуляцией, мы тоже вызываем перегрузку мышцы. Такая же незаметная перегрузка формирует симптомы трапециевидной мышцы из-за привычки постоянно откидывать волосы со лба резким движением головы. А подпирая руками подбородок, мы перегружаем трапециевидную мышцу спины. В общем, капля за каплей — вода камень точит. Так и хроническая микротравма — коварна своей незаметностью. Кстати, для этого и существует профилактика — чтобы вовремя устранять накопившиеся перегрузки.
Касаясь вопросов статистики, нужно отметить, что слева трапециевидная мышца болит чаще, чем справа. Хотя, если следовать логике – должно быть наоборот. Ведь у большинства людей рабочая рука правая. Следовательно, и болеть должно чаще справа. Однако факт остаётся фактом. Скорее всего, всё дело в компенсаторной перегрузке, которая возникает в противовес работающей конечности. Так часто бывает – компенсирующий участок перегружается сильнее основного. Что касается медицины, тут нет никакой разницы — болит трапециевидная мышца справа или слева – методы лечения одни и те же.
Лечение трапециевидной мышцы
Лечение трапециевидной мышцы можно разделить на основное и вспомогательное. Основным – является мягкая мануальная терапия. Она кардинально отличается от обычной мануальной не только своей мягкостью и безопасностью, но и более высокой эффективностью. И это неудивительно, ведь мягкая мануальная терапия на 90% состоит из мышечных и фасциальных техник. Даже при лечении грыжи диска и других заболеваний позвоночника, мягкая мануальная терапия действует не как обычная – резко и с хрустом вправляя позвонки, а, через, так называемый, мягкий мышечный «рычаг», который полностью исключает любую опасность.
Вспомогательным лечением трапециевидной мышцы служат физиопроцедуры, массаж и лекарственная терапия, которая, по нашему мнению, уместна лишь в крайних случаях острой боли. И, в завершение темы, несколько слов о профилактике.
И, в завершение темы, несколько слов о профилактике.
Выполняйте один раз в 3-6 месяцев по одному профилактическому сеансу мягкой мануальной терапии, и вы забудете, как болит трапециевидная мышца.
Трапеция — где она находится и как ее укрепить
Автор статьи: Fitness Guru
У нас есть три мышцы трапеции в теле. Две на спине и третья вокруг тазобедренного сустава. Вы хотите получить полную информацию? Опишем их все!
В теле человека три трапециевидных мышцы. Первая расположена на верхней части спины, вторая – на задней части бедра, а третья – в нижней части спины. Ниже вы найдете самую важную информацию обо всех этих мышцах.
Трапеция вложения
Латинское название этой мышцы – трапеция. Это ровная мышца, расположенная с обеих сторон позвоночника. Обе трапеции образуют форму ромба или четырехугольника и являются наиболее поверхностно расположенными мышцами верхней части спины.
Начальные трейлеры расположены на:
- верхний шейный и затылочный отдел,
- связка шеи,
- остистые отростки позвонков C7-Th22.

Конечные прицепы расположены на:
- плечевой конец ключицы,
- плечевой аппендикс и гребень лопатки,
- медиальная часть верхнего гребня лопатки.
Функции мышцы трапеции
Верхняя часть мышцы поднимает лопатку или наклоняет голову назад (когда лопатка стабильна). Средняя часть приближает лопатку к позвоночнику, а нижняя часть опускает лопатку или поднимает туловище (со стабилизированной лопаткой).
Взаимодействие верхней и нижней частей позволяет руке подняться под углом 90 градусов. Вся мышца прижимает лопатку к груди.
Упражнения для мышц трапеции
Если вы хотите укрепить трапецию, вам нужно выполнять движения, за которые отвечают эти мышцы. Вот некоторые из наших советов:
- поднимайте руки с гантелями выше линии плеча,
- поднимайте руки с гантелями вбок с изгибом туловища вперед,
- опускайте руки, держа штангу с сопротивлением,
- подтягивание на турнике,
- гребля.
Что делать, если трапеция болит
Перегрузка верхней трапеции довольно распространена.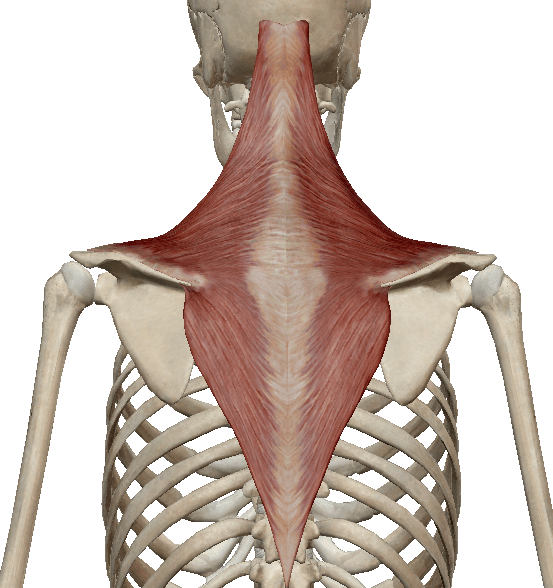 Это происходит как с людьми, которые работают физически, так и с теми, кто работает за компьютером. Чаще всего возникает чувство чрезмерного напряжения шеи, боли (при прикосновении и без) или жжения. Облегчение принесет:
Это происходит как с людьми, которые работают физически, так и с теми, кто работает за компьютером. Чаще всего возникает чувство чрезмерного напряжения шеи, боли (при прикосновении и без) или жжения. Облегчение принесет:
- регулярные физические нагрузки,
- шейные упражнения позвоночника (растяжение и укрепление),
- холодные компрессы (кубики льда в ткани),
- массаж.
Если боль не проходит, стоит обратиться к опытному физиотерапевту.
Трейлеры и функция трапеции
Эта мышца называется по-латыни musculus quadratus femoris. Это маленькая, прямоугольная и ровная мышца. 99% людей имеют по одной трапециевидной мышце в каждом тазобедренном суставе, более конкретно в задней и нижней части этих суставов. Один процент людей вообще не имеет этих мышц!
Quadratus femoris прикрепляется к боковому краю седалищной кости и к межпозвонковому бедренному гребню. На его поверхности проходит седалищный нерв, вырезающий вертикальную борозду. Он относится к мышцам, называемым брюшной группой мышц обода нижней конечности.
В его функции входит: отведение, приведение и внешнее вращение бедра.
Упражнения для трапециевидных мышц бедер
Вы не можете «изолировать» эти мышцы или заставить их работать. Так что, если они у вас есть, вы их включите в работу, во время которой выполняете движения внешнего вращения бедер. Многие мышцы участвуют в этих движениях. Ниже приведены примеры упражнений, во время которых они работают:
- поднимая ноги назад, лежа на животе,
- поднимая ноги, согнутые в коленях,
- приседания.
Прицепы и трапециевидная мышца поясницы
Musculus quadratus lumborum расположен с обеих сторон позвоночника, в нижней части спины. Ее поверхностный слой простирается от ребер позвонков L2-L5 до нижнего края 12-го ребра и тела Th22 позвонка. Задний слой расположен между подвздошным гребнем и подвздошно-поясничной связкой, ребрами позвонков L1-L4 и нижним краем 12-го ребра.
Трапециевидная мышца опускает 12-е ребро и изгибает туловище в сторону, когда оно работает. Когда обе мышцы работают одновременно, они стабилизируют поясничный отдел позвоночника
Когда обе мышцы работают одновременно, они стабилизируют поясничный отдел позвоночника
Упражнения для трапециевидных мышц поясницы
Поскольку эти мышцы работают при боковом наклоне туловища, в этом положении они могут быть как укреплены, так и растянуты. Растяжение происходит на противоположной стороне наклона.
Боковые наклоны туловища влево укрепляет четырехугольную мышцу на стороне, противоположной уклону, то есть справа.
Просмотров: 820
Фитнес на трапеции
Если вы пообещали себе, что в новом году займетесь спортом и придете в форму, а ваш запал уже начал угасать, то дополнительный стимул вам может придать перспектива повисеть вниз головой. Trapfit – новое слово в аэробике, которое сочетает упражнения для укрепления мышц с занятиями в воздухе на трапеции.
Раскачивание из стороны в сторону, вися вниз головой – пожалуй, одно из самых веселых спортивных упражнений.
И хотя с первого взгляда это может выглядеть как детское дурачество, упражнения на самом деле представляют собой профессиональную программу занятий на трапеции. Для их выполнения необходима определенная физическая форма, нужно хорошо переносить нагрузки на сердце и обладать хорошей растяжкой.
Джошуа Майкл, создатель программы, объясняет: «Trapfit объединяет в себе воздушную акробатику и фитнес. Программа направлена на укрепление мышц, в частности, речь идет о мышцах плечевого пояса, мышцах рук и верхней части брюшного пресса».
Занятие начинается с 20-минутной разминки на полу. Затем следует 20-минутная разминка в воздухе с использованием тяжелой палки из алюминия и железа весом 5,5 кг. Далее – упражнения в воздухе на трапеции.
По словам Майкла, балка активизирует мышцы спины и предплечий.
«Многие приходят на занятие не потому, что хотят стать стройнее, а потому что видят, как весело оно проходит. Не все осознают, как сильно задействованы мышцы верхней части тела в повседневной жизни. Для выполнения простых каждодневных дел необходимо быть сильным. На наших занятиях мы не только укрепляем свое тело, но и от души веселимся. Одно простое подтягивание или раскачивание на трапеции требует больших усилий. Я уже не говорю о мышцах брюшного пресса во время раскачивания на трапеции».
Во второй половине занятий посетителям предоставляется возможность от души подурачиться.
Актриса Вероника Мендоза из Калифорнии говорит, что занятие требует определенной подготовки: нужно быть в хорошей спортивной форме. А во время упражнений она чувствует себя ребенком.
«Это очень тяжело прежде всего для рук. Но когда я там, я веселюсь словно ребенок. Мне кажется, что я на качелях. Однако я чувствую, что определенно тренирую свое тело. Поэтому я так люблю эти занятия. Я как будто оказываюсь на детской площадке».
Танцовщица Остин Уэйти из Калифорнии, говорит, что адреналин заставляет забыть о сложности упражнений.
«Это ни с чем нельзя сравнить. Ощущение полета. Вдруг вы чувствуете мышцы, о которых и не подозревали ранее. У меня слабые руки и плечи, и на следующий день после занятий я просыпаюсь и понимаю, над чем мне нужно поработать, понимаю, где мне нужна помощь, чтобы увереннее себя чувствовать. И я снова иду на занятия. Это очень сложно, но и весело, особенно часть, когда можно просто качаться на трапеции. Такое чувство, что ты летаешь».
Мануальный терапевт из Голливуда доктор Кен Бест считает, что Trapfit – упражнение для всего тела.
«Один из плюсов упражнений на трапеции – адреналин. Но при этом обратите внимание, что для их выполнения нужно иметь сильное тело, иначе вы не сможете висеть на балке. По сравнению с обычными упражнениями с весом, когда мышцы тренируются по одной схеме, Trapfit для всех работает по-разному. Это другой тип упражнений на пресс, руки и плечи».
Доктор Бест предупреждает, что Trapfit не для фитнес-новичков. Прежде чем пойти на занятия необходимо потренироваться в спортзале и поработать с весом.
«Упражнения на трапеции – стресс для суставов. А, следовательно, создается большая вероятность их повреждения. Поэтому очень важна предварительная подготовка. Особенно это касается верхней части туловища».
Trapfit — отличный способ познакомится с новыми людьми, потому что некоторые упражнения выполняются в парах.
«Настоящее Время»
врач-невролог: трапециевидная мышца — врата мозга — Английского языка жемчуга
вот увидел фото девушки с развитой трапецией и вспомнил, что будучи подростком я завидовал людям с развитым плечевым поясом, было в этом что-то чарующее. сам я был тогда чем-то средним между колобком и сосиской. потом вспомнил, что у меня трапеция и плечи тоже не слабо развиты.после этого вспомнил, что упражнение «дровосек» на трапецию — мое любимое после приседаний * Омоложение с помощью всего одного упражнения — приседаний, мнение врача * и потом всплыло в памяти, что у меня есть статья врача-невролога А.А.Пономаренко, изложена ниже…
Серьёзные изменения в работе головного мозга и вегетативной нервной системы после болевого раздражения трапециевидной мышцы, заставили искать объяснение этим явлениям. Многочисленные наблюдения, на протяжении 13 лет привели меня к выводу, что тесная анатомическая и функциональная связь ядер добавочного нерва и ядер ретикулярной формации в стволе головного мозга может быть использована в целях лечения. А именно, для воздействия на подкорковые структуры и кору мозга. На мой взгляд, трапециевидная мышца в связи с особенностями своей иннервации (нервного управления) представляет собой уникальную площадку, для доступа к любым мозговым центрам.
В отличие от всех мышц тела эти мышцы управляются из коры, как одноимённого, так и противоположного полушарий. Также её особенность в том, что в формировании добавочного нерва принимают участие ядра шейного отдела спинного мозга и ядра ствола, где и залегает ретикулярная формация. Кроме того, ядра добавочного нерва тесно связаны с ядрами блуждающего нерва, являющегося центром парасимпатической системы. Несмотря на то, что добавочный нерв чисто двигательный, связь его с блуждающим настолько тесна, что они имеют даже общее ядро. Кроме того, эти мышцы имеют довольно большую площадь, и удобное для манипуляций расположение. Удобство в том, что в месте их нахождения нет близко лежащих нервов или органов, которые можно было бы повредить при манипуляциях.
Ретикулярная формация (в переводе с латыни — сетчатое образование) представляет собой часть головного мозга, которая выполняет многочисленные жизненно важные функции. А именно, она, проникая из ствола во все без исключения отделы головного и спинного мозга, объединяет их в единое целое в анатомическом и функциональном смыслах. Все внешние воздействия и сигналы от органов и систем самого тела проходят через эту глобальную сеть. Сигналы эти анализируются, изменяются и поступают в соответствующие отделы мозга для дальнейшего анализа и формирования адекватной ответной реакции.
Кроме того, в ведении этой системы находятся дыхательный, температурный и сосудодвигательный (управляет артериальным давлением и частотой сердцебиения) центры. Эта сеть оказывает нисходящее и восходящее влияния, как тормозящее, так и активирующее (преимущественно), из-за чего её ещё называют восходящей активирующей системой. То есть, это именно та структура, которая отслеживает всё и отвечает за всё, посредством перераспределения активности между различными отделами нервной системы.
Существование такой системы связей, особенностей трапециевидных мышц (описанных выше), а также появление болезненных участков с повышенным тонусом на этих мышцах при различных болезненных состояниях, не может не подталкивать к определённым догадкам. Исчезновение же всевозможных симптомов, относящихся к нарушениям в различных системах организма, после кратковременного интенсивного болевого раздражения таких участков и вовсе приводит к выводу, что мышцы эти являются своеобразной картой головного мозга. Болезненные участки на этой карте соответствуют мозговым центрам, изменившим свою нормальную активность на патологически повышенную или пониженную.
* сегодня покупал рубашку и ещё раз подметил: промышленность создает товары не для нормально развитых людей: воротник рубашки просто врезается в шею. какую бы маечку ни покупал, всегда есть эта проблема.
Что такое нижняя трапеция?
Нижняя трапеция является одним из трех отделов трапециевидной мышцы в верхней части спины. Большая мышца, растягивающаяся от задней части шеи наружу до плеч и посередине спины, трапеция выполняет ряд функций в верхней части тела, в зависимости от того, какая часть мышцы активируется. Нижняя или нижняя часть отвечает за медиальное вращение и депрессию или поворот внутрь и вытягивание лопаток вниз.
Хотя это одна непрерывная мышца, трапеция имеет три четких участка: верхний, средний и нижний трапеции. Он имеет четырехугольную или ромбовидную форму с треугольником верхней трапеции, прикрепленным к затылочной кости у основания черепа и распространяющимся наружу к вершине лопатки. Средняя трапеция простирается горизонтально между верхним грудным позвонком и верхними наружными краями лопаток. Под ним находится перевернутый треугольник нижней трапеции, который проходит по диагонали по обе стороны от позвоночника от остистых отростков среднего-нижнего грудного позвонка до нижних внутренних краев лопаток.
Поскольку его волокна идут наклонно, начиная с середины спины и простираясь вверх и наружу для прикрепления к любому из лопаток, нижняя трапеция расположена так, чтобы тянуть вниз и внутрь на лопатках. При этом он поворачивает нижний угол лопатки, которая является самой нижней точкой треугольной лопатки, внутрь к позвоночнику. Он также работает против верхней трапеции, которая является разделением мышц, работа которых заключается в том, чтобы поднять лопатки или пожать плечами. Верхняя трапеция часто чрезмерно развита, особенно у людей, которые работают перед компьютером, поэтому укрепление нижних волокон этой мышцы является рекомендуемой стратегией для расслабления плотных верхних волокон.
Чтобы укрепить нижнюю трапецию и опустить плечи вниз и назад, рекомендуется выполнить несколько упражнений. Одним из таких упражнений является настенная горка, которая работает как нижних, так и средних волокон. Чтобы выполнить скольжение по стене, нужно стоять, прижав голову, лопатки и копчик к стене, а ноги на расстоянии 10 или 12 дюймов от стены. Сжимая лопатки назад и вместе, он должен твердо прижать локти к стене по обе стороны от тела руками вверх и костяшками пальцев, касающимися стены. Поддерживая это втянутое и вдавленное положение лопатки, он должен затем медленно сдвинуть локти вверх по стене до уровня плеч и полностью вниз по направлению к его бокам, никогда не опуская руки вперед и никогда не снимая напряжения в лопатках.
Другое рекомендуемое упражнение для нижней трапеции — подбородок, который использует нижние волокна, чтобы подтянуть лопатки вниз и вместе, когда тело тянет вверх. Для любого, кто не может выполнить подбородок без посторонней помощи, вспомогательная машина подбородка, которая встречается в большинстве клубов здоровья, является приемлемой заменой. Как и в случае с настенной горкой, необходимо выполнить от двух до четырех подходов и до 12 повторений для повышения прочности.
ДРУГИЕ ЯЗЫКИ
Тренировка трапеций — как правильно качать трапециевидные мышцы
Трапеции — основа массивности верхней части туловища. Регулярные тренировки трапециевидных мышц укрепляют мускулатуру позвоночного столба, формируя спортивную осанку — а также обеспечивают возможность работы с тяжелым весом при выполнении базовых упражнений со штангой.
Однако при прокачке трапеций необходимо соблюдать правильную технику и осознанно вовлекать в работу целевую мышцу — кроме этого, выполнять упражнения нужно в определенном диапазоне повторений. Анатомия трапециевидных мышц и три лучших упражнения — в материале ниже.
// Мышцы трапеций — анатомия
Трапециевидная мышца располагается в верхней части спины. Она состоит из трех пучков, каждый из которых выполняет определенную функцию. Например, верхний отдел трапеции стабилизирует шею, поднимает плечевой пояс и участвует в движении лопаток.
Средняя часть трапеции обеспечивает сведение лопаток и находится между ними. В свою очередь, нижний пучок трапециевидной мышцы участвует в опускании и вращательных движениях лопаток. Общая функция трапеций – поддержание туловища в прямом положении.
В силовых тренировках трапециевидная активно участвует в тяговых упражнениях (становая тяга, тяга к поясу и тп), махах, шрагах, а также при удержании веса. Важно и то, что большие и прокаченные трапеции позволяют комфортнее выполнять приседания со штангой (придерживая гриф на спине в области лопаток).
// Читать дальше:
Как качать правильно?Трапеция – сильная и выносливая мышца. Для ее эффективной тренировки необходимо выполнять упражнения с достаточно большим рабочим весом, чтобы обеспечить нужную нагрузку и стимулировать мышечный рост. Именно поэтому техника должна быть безупречной.
Отдых между подходами – 30-60 сек, в зависимости от способностей восстановления организма. Число повторений в подходе — от 10 до 15. Чтобы точнее определить нужное число повторений, ориентируйтесь на ощущение в мышцах. Завершающие 2-3 повтора в подходе должны проходить с ощущением характерного жжения в трапециях.
Совет для повышения интенсивности тренировки трапеций — делать паузы в пиковой точке сокращения мышцы (то есть, в верхней точке упражнения).
// Читать дальше:
Упражнения для тренировки трапециевидных мышц в тренажерном залеТрапециевидная мышца участвует во всех базовых движениях при тренировке спины и плеч. Однако есть еще несколько эффективных упражнений, которые позволяют отдельно прокачать трапецию. Рекомендуем выполнять их в день тренировки спины или плеч.
// Тренировка для спины
1. Тяга к подбородку
В этом упражнении прорабатываются трапециевидные и дельтовидные мышцы. В положении стоя держите штангу или гантели узким хватом, чтобы между кистями оставалось расстояние 15-20 см. Поднимайте вес вдоль тела до уровня подбородка. Локти — направлены вверх. Чтобы акцентировать нагрузку именно на трапециях, в верхней точке локти должны быть выше уровня плеч.
Если взять вес широким хватом, то большая часть нагрузки придется на средний пучок дельты — то есть, тяга к подбородку эффективно подойдет как для прокачки трапеции, так и для тренировки дельтовидных мышц.
2. Махи с гантелями в короткой амплитуде
Обычно махи выполняют для тренировки плеч. Но при определённой технике это упражнение даёт ощутимую нагрузку на верхнюю часть трапеции.
Исходное положение — стоя прямо, держите гантели в руках. Бедра и ягодичные мышцы должны быть напряжены. Спина зафиксирована в прямом положении, голова направлена вперед. Поднимайте гантели через стороны и тянитесь локтями вверх.
В самой верхней точке кисти должны оказаться чуть выше головы. Затем опустите гантели только до уровня плеч. Движение в такой короткой амплитуде направлено именно на прокачку трапециевидной мышцы. Она постоянно находится в напряжении, испытывая статодинамическую нагрузку.
В этом упражнении вес гантелей не важен. Подберите такое отягощение, чтобы выдержать запланированное время под нагрузкой. Выполняйте махи 4 подхода по схеме: 40 секунд работы / 40 секунд отдыха.
3. Выпады в сторону с гантелями
Упражнение в большей степени нацелено на проработку мышц ног. Однако статичное удержание веса создает ощутимую нагрузку для трапециевидной мышцы и предплечий. Также в этом упражнении сбалансированно работают мышцы пресса и ягодицы. Они стабилизируют позвоночник в прямом положении.
Сделайте широкий шаг в сторону и подконтрольно переносите вес тела на выставленную ногу, сгибая колено. Другая нога остается прямой и поддается легкому растяжению. Старайтесь держать спину ровной, слегка наклонив корпус вперед.
При желании выпады в сторону можно заменить на «прогулку фермера». Смысл упражнения в том, чтобы передвигаться короткими шагами, держа в руках гантели.
// Читать дальше:
Как тренировать трапецию в домашних условиях?Домашние тренировки с весом собственного тела тоже помогут укрепить трапециевидную мышцу. Например, тренировать трапецию можно в упоре лежа.
Для этого примите упор лежа, как при классических отжиманиях от пола. Расслабьте трапециевидную мышцу, чтобы плечи слегка прогнулись назад, а лопатки сжались. Затем напрягите трапецию — лопатки должны расправиться. В этом упражнении количество повторений не имеет значения. Выполнение одного подхода должно занимать примерно 45-60 секунд.
Продвинутым атлетам тренировать трапецию можно в стойке на руках у стены. Здесь тот же принцип мышечной работы. Стоя на руках, расслабьте трапецию, чтобы плечи “провалились” вниз. Затем напрягите трапецию и верните плечи в исходное положение.
// Читать дальше:
***
🍒 Новые материалы на Фитсевен — 5 раз в неделю! Следите за обновлениями!Для получения максимального эффекта от тренировки трапециевидной мышцы необходимо ее прокачивать вместе с остальными мышечными группами. Только в этом случае можно добиться видимого и ощутимого результата в увеличении трапеции.
Автор: Роман Кононов
В продолжение темы
Дата последнего обновления материала — 5 февраля 2021
Трапециевидное тело — обзор
3.32.4.6.1 Медиальное ядро трапециевидного тела
Нейроны MNTB часто сравнивают с кустистыми клетками VCN, потому что они имеют несколько общих черт, включая сильный, многоузловой, афферентный синапс (чашечка Held), и они обладают характеристиками зажигания и проводимостью, зависящей от напряжения, которые напоминают проводимость густых ячеек (Banks, MI и Smith, PH, 1992; Forsythe, ID, и Barnes-Davies, M., 1993; Brew, HM and Forsythe, ID. , 1995). Однако есть и заметные отличия.Например, нейроны MNTB редко запускают более одного потенциала действия при деполяризации (Banks, MI и Smith, PH, 1992; Forsythe, ID и Barnes-Davies, M., 1993), тогда как кустистые нейроны VCN обычно запускают один, но до три потенциала действия на ступень деполяризующего тока. Однако у мышей при температуре 22-25 ° C сообщалось о множественных потенциалах действия (Brew, H. M. et al. , 2003). Во-вторых, высота потенциала действия, измеренная в срезах мозга при 33–34 ° C, немного больше в нейронах MNTB (∼50 мВ; Banks, M.I. and Smith, P.H., 1992), чем в кустистых нейронах, где он составляет в среднем 30-40 мВ (Oertel, D., 1983; Francis, H. W. and Manis, P.B., 2000).
Как и густые клетки VCN, главные нейроны MNTB обладают калиевым током, активируемым низким напряжением, и высокопороговым выпрямителем с задержкой. Ток, активируемый низким напряжением, снижается 4-AP и блокируется DTX-I (Бэнкс, Мичиган и Смит, PH, 1992; Форсайт, И. Д. и Барнс-Дэвис, М., 1993; Брю, HM и Форсайт. , ID, 1995; Додсон, П.D. et al. , 2002). Детальное фармакологическое исследование токов у крыс (Dodson, P. D. et al. , 2002) показало, что блокирование с помощью DTX-I является относительно полным, вовлекая каналы Kv1 в генерацию проводимости. Ток также в значительной степени блокируется DTX-K, который является избирательным для каналов, содержащих Kv1.1. Титюстоксин-Kα блокирует около половины тока, активируемого низким напряжением, что позволяет предположить, что только субпопуляция каналов содержит Kv1.2. Ноксиустоксин, который также блокирует Kv1.2, а также каналы, содержащие Kv1,3 и Kv 1,7, также блокировали около половины тока. Взятые вместе, эти данные предполагают, что ток, активируемый низким напряжением в нейронах MNTB, может состоять из примерно равной смеси каналов, содержащих Kv1.1 и Kv1.2, или Kv1.1 без Kv1.2, но, возможно, Kv1.6. (Додсон, PD и др. , 2002). Наличие других каналов неясно, хотя соединение, блокирующее каналы Kv1.3, на ток не повлияло. В соответствии с этими данными иммуноокрашивание выявило Kv1.1, Kv1.2, Kv1.6, но не Kv1.4 или Kv1.5, в MNTB (Dodson, PD et al. , 2002), и присутствуют высокие уровни мРНК Kv1.1 и Kv1.2 в MNTB путем гибридизации in situ (Grigg, JJ et al. , 2000).
Дополнительные доказательства вклада Kv1.1 в токи, активируемые низким напряжением в MNTB, были получены в исследовании мышей с нокаутом Kv1.1, проведенным Brew H. M. et al. (2003). У этих мышей ток, активируемый низким напряжением, измеренный около порога, был снижен в нокаутах, но не устранен.DTX-I блокировал около половины тока как у нормальных мышей, так и у мышей с нокаутом; оставшийся ток активизировался с другим временным курсом и не был идентифицирован (хотя, поскольку он не был заблокирован DTX-I, он, похоже, не генерируется каналами семейства Kv1). Тем не менее, в нокаутах наблюдалось существенное увеличение числа потенциалов действия, вызываемых во время прямоугольного импульса тока. Эти результаты подтверждают не только Kv1.1 в нейронах MNTB, но также показывают, что в отсутствие Kv1.1, Kv1.2 и / или Kv1.6 остаются и генерируют ток, активированный низким напряжением. В этом исследовании также был идентифицирован небольшой кратковременный, быстро инактивирующийся ток, хотя он, по-видимому, был в значительной степени инактивирован в состоянии покоя.
Наиболее детальные исследования высокопорогового тока выполнены на нейронах MNTB. Как Kv3.1, так и Kv3.3 присутствуют в этих клетках с помощью иммуноцитохимии (Perney, TM et al. , 1992; Li, W. et al. , 2001) и путем гибридизации in situ (Wang, L .Y. et al. , 1998). Сравнение I HT в нейронах MNTB и гомомультимеров Kv3.1 в клетках CHO (Wang, L. Y. et al. , 1998) показывает, что токи очень похожи. Сходство распространяется на зависимость кинетики активации от напряжения и величину чувствительности к блокированию с помощью ТЕА. В этих клетках ток, подобный Kv3.1, способствует реполяризации потенциала действия и позволяет клеткам быстро срабатывать в ответ на серии импульсов деполяризующего тока (Wang, L.Y. et al. , 1998).
Повторяющееся ингибирование медиального ядра трапециевидного тела у монгольской песчанки (Meriones Unguiculatus)
Abstract
Основные нейроны в медиальном ядре трапециевидного тела (MNTB) получают сильный и точный во времени возбуждающий сигнал от глобулярных кустистых клеток в ядре улитки через чашечку Held. Чрезвычайно большие синаптические токи, производимые чашечкой, иногда приводили к мнению о MNTB как о простом релейном синапсе, который преобразует входящее возбуждение в исходящее торможение.Однако электрофизиологические и анатомические исследования показали дополнительное присутствие тормозных глицинергических токов, которые достаточно велики для подавления потенциалов действия в нейронах MNTB, по крайней мере, в некоторых случаях. Источник (и) глицинергического ингибирования MNTB до конца не изучен. Одним из основных внешних источников глицинергического ингибирующего воздействия на MNTB является вентральное ядро трапециевидного тела. Однако было высказано предположение, что нейроны MNTB получают дополнительные тормозящие входы через внутренние связи (коллатерали глицинергических проекций нейронов MNTB).Хотя некоторые авторы постулировали их присутствие, эти побочные эффекты никогда не исследовались подробно. Здесь мы проверяем гипотезу о том, что коллатерали основных клеток MNTB обеспечивают глицинергическое ингибирование MNTB. Мы вводили краситель в отдельные основные нейроны MNTB, прослеживали их проекции и иммуногистохимически идентифицировали их синапсы. Мы обнаружили, что коллатерали оканчиваются внутри MNTB и обеспечивают дополнительный источник ингибирования для других основных клеток, создавая тормозную микросхему внутри MNTB.Только от четверти до трети нейронов MNTB получают такие побочные входные данные. Эта микросхема может производить ингибирование боковой полосы и улучшать настройку частоты нейронов MNTB в соответствии с физиологическими наблюдениями.
Образец цитирования: Dondzillo A, Thompson JA, Klug A (2016) Повторное ингибирование медиального ядра трапециевидного тела у монгольской песчанки ( Meriones Unguiculatus ). PLoS ONE 11 (8): e0160241. https://doi.org/10.1371 / journal.pone.0160241
Редактор: Идзуми Сугихара, Токийский медико-стоматологический университет, ЯПОНИЯ
Поступила: 21 марта 2016 г .; Одобрена: 15 июля 2016 г .; Опубликован: 4 августа 2016 г.
Авторские права: © 2016 Dondzillo et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Эта работа была поддержана NIH / NIDCD R01 DC 011582 to A.K., и частично грантом Центра неврологических заболеваний Скалистых гор NIH P30NS048154. Эксперименты по визуализации проводились в центре усовершенствованной световой микроскопии медицинского кампуса Университета Колорадо Аншутц при частичной поддержке NIH / NCRR Colorado CTSI Grant Number UL1 RR025780.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Медиальное ядро трапециевидного тела (MNTB) — это ядро слухового ствола мозга, участвующее в пути локализации источника звука, а также в ряде других слуховых цепей [1–4]. Он получает возбуждающий сигнал от глобулярных кустистых клеток (GBC), расположенных в контралатеральном переднем вентральном ядре улитки (aVCN) [5-10]. Аксоны GBC большого диаметра перемещаются вдоль акустической полоски, пересекают среднюю линию внутри трапециевидного тела [10] и оканчиваются на основных клетках MNTB через тип гигантского аксо-соматического терминала чашечки, называемого чашечкой Held [5,11] .Одна основная клетка получает входные данные от одного GBC, но аксоны GBC иногда разветвляются внутри MNTB с образованием нескольких чашечек [5,10,12]. MNTB является основным источником глицинергического ингибирования ипсилатеральных медиальных и латеральных верхних оливковых ядер (MSO, LSO, соответственно), вентрального и дорсального ядер латерального лемниска (VNLL, DNLL, соответственно) и других мишеней [13-15 ]. Окрашивание Гольджи и электронная микроскопия (EM) охарактеризовали три типа нейронов в MNTB: звездчатые, удлиненные и основные клетки ([5], кошка), причем последние представляют большинство (82%) клеток ([16], крысиные ).
Из-за преобладающего глицинергического выброса MNTB, он традиционно считается реле в слуховом пути (рассмотрено в [17], но также см. [18,19]). Однако ряд анатомических и физиологических отчетов предполагает, что клетки MNTB также подвергаются нервному подавлению [1,9,20–24]. В частности, положительная метка по глицину и ГАМК существует в пресинаптических компартментах вне чашечки, оканчивающихся на основной клеточной соме, как продемонстрировано с помощью ЭМ, а также с помощью иммуногистохимии и световой микроскопии [25,26].Вклад ГАМК в тормозящий постсинаптический ток уменьшается с возрастом. Электрофизиологические исследования in vitro срезов ствола мозга MNTB показали, что ингибирующий глицинергический вход производит большие постсинаптические токи с очень быстрым затуханием и имеет потенциал шунтировать глутаматергический вход в чашечку чашечки [20,21,27]. Записи нейронов MNTB in vivo демонстрируют несколько типов глицинергических эффектов, действующих на активацию MNTB по-разному, включая латеральное ингибирование.Это латеральное ингибирование блокировалось применением блокатора рецепторов глицина стрихнина [24], что позволяет предположить наличие глицинергических входов на основные нейроны MNTB.
Недавнее анатомическое исследование с использованием объемного отслеживания нейронов у мышей показало, что вентральное ядро трапециевидного тела (VNTB) является основным внешним источником глицинергического входа в MNTB [20]. Помимо этого внешнего источника, несколько исследований предложили дополнительный внутренний источник тормозящего воздействия на нейроны MNTB.Например, отслеживающие исследования с использованием нейробиотина или пероксидазы хрена (HRP) в регистрирующем электроде показали, что меньшая часть основных клеток MNTB имеет аксональные коллатерали, возвращающиеся обратно в окрестности исходной клетки (песчанка, [9] кошка [23]; крыса, [28]). Хотя это наводит на размышления, эти коллатерали никогда не исследовались более подробно, и вопрос о том, являются ли они действительными синаптическими входами, никогда не проверялся.
В данной работе мы проверяем две основные гипотезы. 1) остаются ли коллатерали основных аксонов клеток MNTB в ядре и 2) если да, то образуют ли они тормозящие синапсы на других основных клетках MNTB?
Материалы и методы
Заявление об этике
Все процедуры с животными были одобрены Комитетом по уходу и использованию животных (IACUC) Медицинского кампуса Университета Колорадо (номер разрешения B-88412 (05) 1D.Кроме того, неукоснительно соблюдались все применимые законы и постановления, а также политика PHS. В этих экспериментах использовалось 37 песчанок мужского и женского пола, выращенных в нашей собственной колонии в Калифорнийском университете в Денвере. Все процедуры с животными были одобрены Комитетом по уходу и использованию животных Школы медицины Университета Колорадо и проводились в соответствии со стандартами Национального института здравоохранения по гуманному обращению с лабораторными животными.
In-vitro Заполнение и запись отдельных клетокПодготовка срезов .Срезы ствола мозга получали из монгольских песчанок ( Meriones unguiculatus ) обоих полов в возрасте от p13 до p17 (все стадии после появления слуха). Животных анестезировали ингаляцией изофлурана (IsoFlo, Abbott Laboratories, США) и декапитировали. Ствол мозга вырезали и разрезали на срезы от 180 до 300 мкм с помощью вибратома (VT1000S, Leica, Wetzlar, Германия) под ледяной диссекцией (125 мМ NaCl, 2,5 мМ KCl, 1 мМ MgCl 2 , 0,1 мМ CaCl 2 , 25 мМ глюкоза, 1.25 мМ NaH 2 PO 4 , 25 мМ NaHCO 3 , 0,4 мМ аскорбиновой кислоты, 3 мМ мио-инозитола, 2 мМ пировиноградной кислоты; все химические вещества от Sigma – Aldrich, Миссури) барботировали в течение не менее 15 минут с 5% CO 2 -95% O 2 . Срезы переносили в инкубационную камеру, содержащую искусственную спинномозговую жидкость (aCSF) (125 мМ NaCl, 2,5 мМ KCl, 1 мМ MgCl 2 , 2 мМ CaCl 2 , 25 мМ глюкозы, 1,25 мМ NaH 2 PO 4 , 25 мМ NaHCO 3 , 0.4 мМ аскорбиновой кислоты, 3 мМ мио-инозитола, 2 мМ пировиноградной кислоты; все химические вещества от Sigma – Aldrich) и барботировали 5% CO 2 -95% O 2 . Срезы инкубировали до 45 мин при 37 ° C, после чего камеру доводили до комнатной температуры. Заполнение краской производили в течение 4–5 часов после нарезки.
Одноклеточная загрузка красителя
Одиночные срезы головного мозга помещали в записывающую камеру и просматривали на микроскопе Zeiss Axioscope II с водно-иммерсионным объективом 40x, который был частью стандартной установки патч-зажима (описанной в [29]).В некоторых экспериментах нейроны, расположенные в MNTB, были визуализированы, и цельноклеточный патч фиксировали стеклянными электродами. Патч-пипетки (2,4–3,2 МОм) извлекали из боросиликатного стекла 1,5 мм (Harvard Instruments, Кент, Великобритания) с помощью универсального съемника DMZ (Zeitz Instruments, Мюнхен, Германия) и заполняли внутренним раствором глюконата калия (113 мМ K-глюконат, 4,5 мМ MgCl 2 , 9 мМ HEPES, 5 мМ EGTA, 14 мМ трис2-фосфокреатин, 4 мМ Na 2 АТФ, 0,3 мМ трис-GTP, 1,5 мМ CaCl 2 ; pH доведен до 7.25 с КОН; 295–300 мОсм; все химические вещества от Sigma – Aldrich). После успешного получения конфигурации цельной ячейки ячейку оставляли прикрепленной к электроду примерно на 20 минут без каких-либо дополнительных электрофизиологических записей, чтобы обеспечить диффузию красителя. Биоцитин (Sigma-Aldrich, кат. B4261) в концентрации 3 мг / мл, растворенный во внутренней пипетке, использовали для мечения нейронов.
В дополнительных экспериментах клетки метили внеклеточной струей раствора красителя, доставленной в область сомы клетки, с последующей оккстеклеточной электропорацией.Для этого метода использовались стеклянные пипетки, аналогичные описанным выше, но они были заполнены aCSF и биоцитином с концентрацией 3 мг / мл. Внеклеточные затяжки подавали с продолжительностью 500 мс и давлением в диапазоне 4–6 фунтов на квадратный дюйм, подавали через Picospritzer III (Parker Hannifin Corp, Кливленд, Огайо, США), который был подсоединен к электроду. Электрические стимулы представляли собой прямоугольные импульсы 5–7 В длительностью 20 мс, повторяемые 20–50 раз, подаваемые с помощью четырехканального стимулятора STG 2004 с компьютерным управлением (Multi Channel Systems, Ройтлинген, Германия) и блока изоляции стимуляции (Iso-flex, AMPI). , Иерусалим, Израиль).Чтобы свести к минимуму вероятность неспецифической метки, юкстаклеточную электропорацию получали под визуальным контролем, в результате чего мы идентифицировали клеточную сому клетки, подлежащей электропорации, под микроскопом с 40-кратным объективом. Затем мы подносили кончик пипетки к клеточной мембране, устанавливали режим прикрепления клеток при сопротивлении 100–600 МОм, аналогичный конфигурации «рыхлого пятна», а затем применяли процедуру электропорации. Благодаря постоянному потоку записывающей ванны через камеру со скоростью 3 мл / мин, краситель вымывался из ванны с постоянной скоростью, что дополнительно уменьшало вероятность маркировки близлежащих клеток.Пипетку оставляли на месте в течение примерно 2 минут после электропорации, чтобы позволить красителю диффузию.
Объем выборки
Мы успешно заполнили 39 нейронов в срезах, приготовленных из 37 песчанок в возрасте P13 –P17 (сводка в таблице 1). 22 из этих заполненных нейронов показали либо очень короткие нейриты, либо отсутствие помеченных нейритов (общая длина нейритов менее 60 мкм). В остальных 17 нейронах общая длина нейритов, меченных в каждом нейроне, составляла не менее 60 мкм, не считая сомы.В некоторых случаях меченые нейриты были извлечены без исходной сомы, которая могла стать пористой для красителя либо во время процедуры зажима пластыря (и снятия электрода), либо в процессе электропорации.
Большая часть анализа была проведена на коронарных срезах головного мозга (35 случаев). Эта плоскость была выбрана потому, что нейриты в значительной степени остаются в пределах разреза в этой плоскости, что означает, что они только иногда переплетаются взад и вперед, и если это так, то это находится в пределах 30 мкм в ростро-каудальной плоскости относительно сомы.Важно отметить, что нейриты не проецируются в ростро-каудальном направлении. Если бы это было так, мы бы увидели точечный узор, образованный мечеными нейритами, вышедшими из плоскости, чего мы не наблюдали. Кроме того, в дополнительных экспериментах с использованием срезов, разрезанных в горизонтальной и парасагиттальной плоскости, нейриты обычно перемещались в медиолатеральном направлении.
Реконструкция меченых клеток
Следующий процесс был использован для реконструкции трехмерной архитектуры помеченных клеток.Монтированные срезы получали на конфокальном микроскопе Olympus FV1000 (Olympus, Waltham, MA) с использованием объектива 60x, 1,2 NA, или 20x, 0,75 N.A., или объектива 10x, 0,4 N.A. Одна лазерная линия TRITC (547 нм) использовалась для возбуждения красителя, который использовался для заполнения ячейки. Для каждого заполнения было записано от 19 до 54 конфокально полученных последовательных оптических срезов (среднее значение = 40 сечений на стопку) и импортировано в Matlab 2014a с установленной программой обработки изображений (Mathworks, Natick MA). Чтобы добиться наилучшей визуализации трехмерной структуры помеченной клетки, мы стремились выделить пиксели, связанные с флуоресцентно помеченной меткой красителя, локализованной в клетке.Чтобы изолировать эти пиксели, мы создали маску изображения контура ячейки, которая была применена к каждой секции стека. Определяемая пользователем маска была нарисована вокруг помеченных компонентов, оставляя границу размером ~ 20 пикселей, а не только на видимых структурах. Этот метод не только исключал источники шума (например, автофлуоресценцию и артефакты), но и позволял полуобъективную визуализацию меченой клеточной архитектуры. Вкратце, для каждого стека изображений максимальная проекция (то есть максимальное значение интенсивности пикселей для каждого положения [x, y] в стеке изображений) вычислялась как единственная матрица для первого канала изображения RGB.Значения интенсивности пикселей максимальной матрицы проекции были нормализованы между 0 и 1 на основе деления значения интенсивности каждого пикселя на максимальное значение пикселя. Нормализованная матрица максимальной проекции была преобразована в двоичное изображение, в котором значения пикселей> 0,75 = 1 и значения пикселей <0,75 = 0. Для дальнейшего уточнения контура ячейки использовался алгоритм обнаружения кластера пикселей (например, функция Matlab Image Processing Toolbox 'bwareaopen' ) был применен к матрице для обнаружения всех смежных кластеров пикселей и исключения кластеров <40 пикселей.На этом этапе были удалены небольшие области артефакта высокой интенсивности пикселей. Из оставшихся смежных кластеров пикселей в двоичном изображении самый большой был связан с контуром помеченной ячейки. Чтобы создать окончательную маску изображения контура ячейки, которая будет применяться к каждому разделу, определяемая пользователем маска многоугольника была обведена вокруг самого большого непрерывного кластера пикселей в максимальном двоичном изображении проекции. Эта маска изображения, созданная из определяемой пользователем области интереса, была применена к каждому отдельному оптическому участку импортированного стека изображений.Наконец, значения пикселей, извлеченные из каждой плоскости стека в пределах определяемой пользователем интересующей области, были использованы для создания трехмерного графика (функция Matlab «patch»).
Иммуногистохимические процедуры
В каждом отделе мозга от 3 до 9 отдельных нейронов были нацелены на маркировку, в зависимости от размера и доступности MNTB в каждой секции. После завершения заполнения ячеек срезы извлекали из записывающей камеры и переносили в 4% параформальдегид (PFA) в фосфатно-солевом буфере (PBS) на срок до 2 часов.Затем срезы либо обрабатывали непосредственно для иммуногистохимии, либо повторно разделяли. Срезы, отобранные для повторного разделения, переносили в 30% раствор сахарозы на срок до 3 дней, повторно разрезали на срезы толщиной 50 мкм на замораживающем микротоме (Leica SM 2010R, Buffalo Grove, IL) и затем обрабатывали для иммуногистохимии.
Для иммуногистохимического мечения срезы инкубировали в 0,3% Triton X-100 (TX100; Sigma-Aldrich) и 5% нормальной козьей сыворотке (NGS; Jackson Immunoresearch Laboratories, West Grove, PA) в PBS в течение 60–120 минут.Первичные антитела разводили в 1% NGS и 0,3% TX100 в PBS, и ткань инкубировали в течение двух дней при 4 ° C. После инкубации первичных антител ткань несколько раз промывали 2% NGS в PBS. Вторичные антитела получали в 1% NGS и 0,3% TX100, и срезы инкубировали при комнатной температуре в течение 2 часов. После последней промывки срезы помещали в среду Fluoromount-G (SouthernBiotech). Установленные срезы получали на конфокальном микроскопе Olympus FV1000 с использованием 60x, 1,2 Н.A., или 20x, 0,75 N.A., или 10x, 0,4 N.A. Для возбуждения красителей использовали три лазерные линии: 488 нм, TRITC (547 нм) и 635 нм. Основываясь на аксиальных размерах вокселей от 0,07 мкм до 0,2 мкм при сканировании с максимальным увеличением иммуноположительных кластеров, мы обосновали сопоставление пресинаптических и постсинаптических структур. Для всех идентифицированных терминалов, в которых мы наблюдали положительный иммуногистохимический сигнал, мы оценивали аппозицию по следующим критериям. Сопоставление требовало положительных сигналов как от биоцитина, так и от иммуногистохимической метки для совместного присутствия как минимум в двух смежных пикселях.
Антитела.
Первичные антитела, использованные в этом исследовании, перечислены в таблице 2. Моноклональные антитела мыши к гефирину mAb7a (Synaptic Systems, номер по каталогу: 147 011, RRID: AB_887717 Goettingen, Германия) были созданы против экстракта сырых синаптических клеток спинного мозга крысы. мембраны [30,31] и показано на Вестерн-блоттинге экстрактов мембран головного мозга крысы для специфического связывания с полосой 93 кДа мембранного белка (гефирина) [32,33]. Специфичность этого антитела также была показана ранее у мышей с нулевым мутантом по гефирину, что не привело к иммуномечению кластеров гефирина [34].
Кроличье антитело против SNAP-25 Sigma-Aldrich (номер по каталогу: S9684, номер партии: 069K4784, RRID: AB_261576) было выделено из фракции IgG антисыворотки против синтетического пептида, соответствующего N-концу человеческого SNAP-25 (аминокислоты 9–29 с лизином, добавленным на С-конце), который идентичен последовательности у крысы, мыши и курицы. Специфичность этого антитела была продемонстрирована путем ингибирования иммуноблоттинга после предварительной инкубации антитела с иммунизирующим пептидом SNAP-25 (аминокислоты 9–29 с С-концевым лизином; данные производителя).
Были использованы следующие вторичные антитела (все от Invitrogen / Molecular Probes): козий антикроличий IgG, конъюгированный с Alexa 647 (каталожный номер: A21245), и козий антимышиный IgG, конъюгированный с Alexa 488 (каталожный номер: A11029). . Оба вторичных антитела использовали в разведениях 1: 1000. Для реакции с биоцитином использовали авидин, конъюгированный с TRITC (конъюгат ExtrAvidin-TRITC; Sigma-Aldrich, кат. E3011), в разведении 1: 1000 в том же растворе, что и вторичные антитела.
Results
Целью нашего исследования было проверить гипотезу о том, что аксоны от нейронов MNTB формируют коллатеральные тормозные входы на соседние нейроны MNTB.Мы проверили эту гипотезу, пометив аксоны одиночных нейронов MNTB, определив потенциальные повторяющиеся коллатерали, охарактеризовав их терминалы и оценив количество нейронов, которые производят такие коллатерали. В то время как предыдущие исследования предполагали их возникновение [9,23,28], в этих исследованиях не выяснялось, как часто возникают эти коллатерали и действительно ли они образуют синаптические окончания.
Маркировка одиночных нейронов в МНТБ
Мы использовали как цельноклеточную, внутриклеточную диффузию красителя, так и электропорацию отдельных клеток для маркировки отдельных нейронов в MNTB.Мы решили пометить только небольшое количество нейронов на MNTB, чтобы снизить вероятность реконструкции слишком большого количества меченых перекрывающихся нейритов, которые могли бы скрыть происхождение нейритов. Таким образом, было помечено от 3 до 9 нейронов на срез (рис. 1), но обратите внимание, что не каждый эксперимент приводил к восстанавливаемому меченому нейрону. Важно отметить, что в нейронах, которые могли пассивно поглощать краситель из прилипшей к клетке инфузии, мы никогда не наблюдали меченых нейритов. На рис. 1А показаны 8 очень ярких клеточных сомат и несколько других, которые светлее фона, но тусклее восьми ярких клеток.Эти диммерные клетки, скорее всего, являются примером пассивного поглощения красителя из затяжки без присоединенной к клетке электропорации, которая никогда не приводила к маркировке нейритов. Из шести меченых нейронов, расположенных по крайней мере на 2 диаметра клеток друг от друга (рис. 1A), два имеют очень разные нейриты, выходящие из сомы в сторону дорсальной части MNTB. На рис. 1B показаны сомы трех меченых нейронов плюс один длинный нейрит из неидентифицированного клеточного тела, охватывающий медиолатеральную длину MNTB. Более длинные нейриты имеют тенденцию смешиваться и образовывать спирали, по-видимому, друг вокруг друга, что делает очень сложным проследить их пути (стрелка на Рис. 1B, см. Также Рис. 2).Поле на рис. 1B указывает на присутствие отдельного аксонального образования, которое происходит из неидентифицированной клеточной сомы и расположено на другой глубине, чем тела клеток, изображенные здесь.
Рис. 1. Электропорация одной клетки выявляет предполагаемые MNTB к коллатералям MNTB.
A: срез ствола мозга, содержащий MNTB с несколькими нейронами, меченными биоцитином, меченными с помощью метода электропорации одной клетки. Пунктирная линия очерчивает границы MNTB на основе автофлуоресценции ядра.Обозначены шесть нейронов, два из них (стрелки) имеют отдельные нейриты, которые вращаются внутри MNTB. Обратите внимание, что диммерные клетки, вероятно, пассивно помеченные затяжкой, не имеют меченых нейритов. B: Другой пример участка ствола мозга с MNTB и тремя мечеными нейронами, расположенными дальше друг от друга, при этом один нейрит проходит через MNTB между двумя верхними клетками. Красная стрелка указывает место, где нейриты, по-видимому, смешиваются. Красный прямоугольник указывает на область, которая показана в увеличенном масштабе на фиг. 6A.Шкала 100 мкм на обеих панелях.
https://doi.org/10.1371/journal.pone.0160241.g001
Рис. 2. Отслеживание коллатералей основных клеток MNTB требует разреженного мечения нейронов, чтобы избежать перекрытия нейритов.
A: нейрит, происходящий из верхней клетки, движется вниз в непосредственной близости от дендритов нижних нейронов. Другой нейрит, происходящий из той же (верхней) клетки, движется вверх и расщепляется, чтобы вернуться и сформировать терминал в пунктирной рамке.Максимальная проекция на глубину 15,2 мкм (19 виртуальных срезов 0,8 мкм / срез = 15,2 мкм). Пунктирный квадрат увеличен на рис. 5D. B: Увеличение области, показанной в сплошной рамке на панели A, показывающей нейрит верхней клетки (стрелки), спиралевидно закручивающийся рядом с областью дендритного дерева нижнего нейрона (острие стрелки). Средняя проекция на глубину 34,8 мкм (58 виртуальных секций по 0,6 мкм / срез = 34,8 мкм). Шкала на обеих панелях: 20 мкм.
https: // doi.org / 10.1371 / journal.pone.0160241.g002
Мозаичное сканирование, показанное на рис. 2A, представляет собой максимальную проекцию через общую глубину 15 мкм секции ствола мозга P13 MNTB. Две соматы расположены на расстоянии более 40 мкм друг от друга. Предполагаемый аксон, происходящий из верхней клетки, движется вниз в окрестности второй сомы. Увеличенное изображение на рис. 2B показывает, что этот аксон (стрелки) из верхней ячейки поворачивается вверх в область дендрита (стрелки), принадлежащего нижней ячейке.Здесь аксон (стрелки), кажется, закручивается по спирали вокруг нейрита (предполагаемого дендрита) нижней клетки.
Эти находки указывают на плотно упакованные нейриты, происходящие из MNTB, которые могут достигать ~ 40 мкм осевого расстояния. Это свидетельствует о том, что для отслеживания коллатералей одиночных нейронов MNTB важно производить только разреженное мечение клеток MNTB. Кроме того, наши результаты подтвердили, что только те нейроны, которые были подвергнуты электропорации после нанесения краски, стали метиться от сомы к нейритам.В то время как те нейроны, которые были пассивно помечены одной только затяжкой, никогда не проявляли меченых нейритов.
Морфология окончаний аксонов и меченых клеток
Заполненные биоцитином коллатерали, оканчивающиеся внутри MNTB, образуют различные паттерны (Рис. 3). Как правило, мы наблюдали, что терминалы, обернутые вокруг предполагаемых клеточных тел (фиг. 3A и 3C), и более крупные ветви расщепляются на более мелкие ветвления (фиг. 3A и 3C) перед инкапсулированием сомы (стрелки на фиг. 3A и 3C). В некоторых случаях нейриты, по-видимому, обвивают соседние сомы в непосредственной близости от тела клетки, из которого они произошли (стрелки на рис. 3C указывают на нейриты).
Рис. 3. Типы аксональных окончаний, образованных нейритами в MNTB.
A: Структура с утолщениями в форме бутона, образованными на ветвях, которые образуют круглую форму (стрелки). B: Более классические аксональные ветви с узкими и разросшимися утолщениями в виде бутонов (стрелки). C: нейрон, который сформировал терминал (стрелки) на своем коллатерале в непосредственной близости от его исходной сомы. Масштабные линейки: 10 мкм для панелей A, B и 20 мкм для панели C.
https: // doi.org / 10.1371 / journal.pone.0160241.g003
Кроме того, мы также наблюдали окончания аксонов, которые имеют небольшие ответвления с характерными вздутием, предположительно бутоны (Рис. 3B, стрелки), которые разделяются на несколько более мелких ветвей. Эти терминалы также проходят вблизи поверхности сомы клетки, образуя en passant терминалов, а не оборачиваясь вокруг клеток.
Мы проследили заполненные биоцитином сомы и их нейриты и обнаружили, что в 17 случаях, в которых нейриты были успешно помечены на большие расстояния (от 60 мкм до 140 мкм), шесть продуцировали MNTB в коллатерали MNTB.Из них были подготовлены 3D-реконструкции для анализа направления их проекций и глубины, на которой перемещаются их нейриты. Нейриты преимущественно перемещались в медиолатеральном направлении и оставались в основном в корональной плоскости. На рис. 4 показаны два примера, в которых нейриты перемещались преимущественно в медиолатеральном направлении и оставались в пределах 30 мкм от корональной плоскости (рис. 4; ось z представляет ростро-каудальное направление).
Рис. 4. Примеры трехмерных реконструкций двух нейронов, меченных биоцитином.
Реконструкции основаны на порогах интенсивности и алгоритме кластера смежных пикселей (см. Методы) и показывают, что нейриты помеченных нейронов проецировались дорсально (A) или вентрально (B), но оставались в диапазоне от 14 до 30 глубина мкм в корональной плоскости среза (ось z представляет ростро-каудальное направление).
https://doi.org/10.1371/journal.pone.0160241.g004
Для дальнейшего подтверждения наших результатов трехмерного анализа мы провели серию парасагиттальных и горизонтальных срезов и обнаружили, что меченые нейроны действительно проецируются в медиа- боковая плоскость, а не ростро-каудальная плоскость (n = 8, данные не показаны).
Идентичность синапсов в MNTB
Для дальнейшей идентификации предполагаемых окончаний аксонов мы использовали пре- и постсинаптические антитела против SNAP-25 и гефирина. SNAP-25 является маркером пресинаптических окончаний, используемым для идентификации мембраносвязанных белков, участвующих в слиянии мембранных везикул и экзоцитозе [35–37]. Гефирин является постсинаптическим маркером, маркирующим якорный белок для постсинаптических рецепторов GABA , A и глицина [38-40].
На рис. 5A – 5C показано такое же окончание аксона, как показано на рис. 1B (прямоугольник), но с дополнительной маркировкой антител против гефирина.На панелях показана гефириновая метка, локализованная на мембране сомы постсинаптических клеток, что указывает на наличие постсинаптических плотностей (отмечено желтыми стрелками; фиг. 5B). Дополнительное внеядерное окрашивание, вероятнее всего, из-за трансляции белка в клеточной соме, отмечено стрелками (рис. 5B). Наложение биоцитина (пурпурный) с гефирином (голубым) показано на фиг. 5C, что указывает на то, что кластеры гефирина хорошо совпадают с пресинаптическими аксональными элементами коллатералей (фиг. 5C, желтые открытые стрелки).
Рис. 5. Иммуногистохимия в сочетании с меткой индикатора предполагает, что коллатерали MNTB и MNTB образуют функциональные синапсы.
A: компактная круглая структура, помеченная в нейроне, показанном на рис. 1B (прямоугольник). B: Окрашивание гефирином, локализованное на клеточной мембране (стрелки) и окружающей внеядерной области (стрелки). C: Перекрытие двух каналов, показанных на A и B, показывает, что гефирин (голубой) находится рядом с аксоном, меченным биоцитином (пурпурный, пустые стрелки). D: пресинаптическая метка SNAP-25, локализованная по окружности сомы клетки (не маркирована), и E: гефирин (стрелки и наконечники стрелок, как указано выше).F: Наложение гефирина (голубой), SNAP-25 (зеленый) и соответствующего окончания аксона (пурпурный) показывает очень тесную близость нейрита с пресинаптической и постсинаптической плотностью (пустые стрелки). G: Третий терминал, который, кажется, образует рудиментарные фенестрации. H: Окрашивание гефирином показывает круглую форму, которая предполагает тело постсинаптической клетки. I: Гефириновая метка (голубая), по-видимому, сосредоточена вокруг помеченного терминала (пурпурный), который частично инкапсулирует предполагаемую постсинаптическую сому. Масштабные линейки: 10 мкм для всех панелей.
https://doi.org/10.1371/journal.pone.0160241.g005
Мы также подтвердили, что постсинаптические структуры, идентифицированные по метке гефирина, также совпадают с пресинаптической меткой SNAP-25, которая присутствовала в меченных биоцитином аксонах (рис. 5D, 5E и 5F). На рис. 5E показано постсинаптическое мечение гефирина (стрелки), хорошо выровненное с клеточной мембраной, и нецелевое внеядерное окрашивание (стрелки). На фиг.5D показана пресинаптическая метка с антителом к SNAP-25 (стрелки), присутствующая в структуре аксона (фиг.5F).Фиг. 5G представляет собой еще один пример нейрита, меченного биоцитином, который сопоставлен с постсинаптическим гефирином (Фиг. 5H). Терминал пространственно совпадает с высокой концентрацией мечения антителом к гефирину (рис. 5I).
Частота появления залогов MNTB к MNTB
Мы стремились определить частоту коллатералей главных нейронов MNTB; ограничены ли эти коллатерали определенными областями MNTB и проецируются ли они на другие нейроны MNTB в непосредственной близости от их происхождения или на более удаленные нейроны.С этой целью мы проанализировали проекции 17 заполненных нейритов размером 60 мкм и более. Из этих 17 заливок 16 были выполнены в корональных сечениях (рис. 6), а одна — в горизонтальном срезе. Для каждого заполнения венечного среза мы записывали ростро-каудальное расположение сомы и направление движения самого длинного нейрита, выходящего из этой заполненной сомы. На рис. 6А показана микрофотография самого рострального и самого каудального полусечения в корональной плоскости ствола мозга песчанки P18.Ростральная часть соответствует приблизительно -4,9 мм Bregma, а хвостовая часть соответствует приблизительно -5,5 мм Bregma (изменено для молодых животных из Атласа мозга песчанок; Loskota, 1974). Мы проследили расположение заполненных сомат и расположение меченых нейритов и нанесли их на карту в эскизах MNTB (рис. 6B). Рисунки сверху вниз представляют расположение MNTB в порядке от каудального до рострального. Исходные точки стрелок представляют соматы, а направление стрелок представляет направление проекции самого длинного нейрита, выходящего из сомы.Красные стрелки указывают нейроны, у которых есть аксоны с возвращающимися коллатералями в MNTB (5 нейронов). Мы проанализировали перемещение в пределах медиолатеральной оси в корональном срезе MNTB и нормализовали к положению средней линии справа (пунктирная линия).
Рис. 6. Схематическое изображение расположения и направления проекции меченых основных ячеек MNTB.
A: два корональных среза MNTB песчанки, помеченные Nissl, представляющие ближний к ростральному и ближний к каудальному концам MNTB (ростральный на –4.9 мм, хвостовой — 5,5 мм относительно Bregma). B: Расположение происхождения и направления проекции всех меченых нейронов, в которых общая длина нейритов составляла не менее 60 мкм, показано в виде стрелок на каждом ростро-каудальном участке MNTB (красные контуры сверху вниз представляют каудально к ростральному) . Стрелки указывают направление движения самого длинного нейрита, происходящего от каждого меченого нейрона, а начало стрелки указывает приблизительное расположение меченого нейрона на медиолатеральной оси MNTB, нормированной по средней линии справа (пунктирная линия) .Красные стрелки указывают нейроны, у которых есть аксоны с возвращающимися коллатералями в MNTB. Представлены 16 случаев, обнаруженных при препарировании коронковой артерии из 17 случаев. Еще один случай (рис. 5G), обнаруженный в горизонтальном разрезе, здесь не показан.
https://doi.org/10.1371/journal.pone.0160241.g006
На рис. 6В показаны данные для 16 нейронов, которые были заполнены в коронарных срезах. Не показан последний случай (показанный на фиг. 5G), обнаруженный при горизонтальном препарировании.Этот нейрон был расположен в латеро-каудальной части вентрального MNTB и проецировался в медиоростральном направлении.
Таким образом, наши данные предполагают, что возвращающиеся коллатерали, происходящие от нейронов MNTB, обеспечивают ингибирующий вход для других нейронов MNTB, расположенных поблизости от исходной клетки. Более того, мы использовали иммуногистохимию для идентификации и характеристики пре- и постсинаптических окончаний, образованных этими заполненными биоцитином коллатералями. Наконец, мы показали, что терминалы состоят из пре- и постсинаптических маркеров, предполагая, что это функциональные синапсы, а не просто случаи, когда нейрит проходит близко к нейрону MNTB.Хотя они явно присутствуют, частота этих коллатералей в MNTB невысока, и только от четверти до трети аксонов MNTB формируют коллатерали по отношению к другим нейронам MNTB.
Обсуждение
Основные выводы
Наши результаты демонстрируют, что субпопуляции нейронов MNTB получают тормозящие входы от других нейронов MNTB. Хотя существование таких коллатералей предполагалось различными другими авторами [9,23,41], мы демонстрируем здесь прямо, что эти коллатерали фактически иннервируют постсинаптические нейроны MNTB посредством функциональных тормозных химических синапсов.В нашем определении «функциональный» синапс состоит из афферентного аксона, заканчивающегося пресинаптическим окончанием, и соответствующей постсинаптической плотности. Мы также обнаружили, что только относительно небольшая часть нейронов MNTB получает этот тип коллатерального торможения.
Наши выводы подтверждаются следующими результатами: 1) мечение биоцитином отдельных нейронов MNTB выявило повторяющиеся коллатерали в MNTB, 2) небольшое количество меченных биоцитином нейронов MNTB на срез мозга позволило точно отследить коллатерали и выявить синаптические терминалы, расположенные близко к клеточным телам нейронов MNTB, 3) иммуногистохимическое мечение против пресинаптического маркера SNAP-25 вместе с мечение против ингибирующего постсинаптического маркера гефирина в сочетании с биоцитин-положительными нейронными коллатералями выявило тормозные синапсы, образованные коллатералями, 4) только примерно на четверть у трети успешно помеченных нейронов были либо возвращающиеся коллатерали, либо сформированные терминалы внутри MNTB.
Мы также обнаружили, что возвращающиеся коллатерали образуют два типа синаптических связей: коллатераль либо 1) разделяется на более мелкие ветви с характерными бутоноподобными вздутием, либо 2) образует округлые и компактные концы. Кроме того, наши данные предполагают, что коллатерали обычно синапсы на соседних нейронах MNTB. Мы также не нашли доказательств того, что синапсы следуют какому-либо тонотопически систематическому распределению, хотя размер нашей выборки может быть слишком мал, чтобы сделать это определение.
Частота появления залогов MNTB — MNTB
Наши выводы о том, что возвращающиеся коллатерали происходят с низкой частотой и что их окончания расположены на соседних клеточных телах в MNTB, согласуются с предыдущими наблюдениями, полученными на нескольких видах: песчанках и летучих мышах [9] и кошках [23,41]. ].Одно из этих исследований отмечает низкую частоту этих коллатералей (от 3 до 30%, [9]), что согласуется с нашими выводами. Кроме того, Guinan и Li [41] наблюдали, используя in vivo электрофизиологических исследований в стволе мозга кошки, небольшую подгруппу нейронов с «своеобразным импульсным поведением», и предположили, что это потенциально может быть объяснено повторяющимся ингибирующим воздействием. Наши результаты вместе с этими предыдущими исследованиями подтверждают существование возвращающихся тормозных коллатералей между основными нейронами в MNTB, хотя эти коллатерали, по-видимому, присутствуют только между небольшим подмножеством нейронов MNTB.
Метод инъекции биоцитина в отдельные нейроны под визуальным контролем позволяет специфично маркировать нейроны, исключает объемную инъекцию, которая обычно создает очень яркую область в месте инъекции, и позволяет инъекцию морфологически идентифицированных отдельных основных нейронов, которые оказались очень эффективными. полезен для поиска нейрональных коллатералей. Мечение нейронов в срезах толщиной 180–300 мкм гарантировало присутствие большинства нейритов внутри среза. Однако дальнейшее повторное нарезание срезов до 50 мкм для облегчения проникновения антител могло привести к некоторой потере нейритов или клеточных тел.Из-за этого наша оценка частоты этих залогов может недооценивать фактическое количество залогов. Однако наши результаты все еще хорошо согласуются с существующей, хотя и скудной литературой по этому вопросу.
Предыдущие анатомические и физиологические исследования показали, что практически каждый нейрон MNTB получает существенное глицинергическое ингибирование [20,21,24,29] и что основным источником этого ингибирования является ипсилатеральный VNTB [20]. Используемые здесь наполнения одиночных клеток специально помечают ингибирование, происходящее только от одиночных нейронов MNTB, тем самым изолируя эти проекции от других возможных ингибирующих входов в нейроны MNTB.Из-за использованной здесь методологии наши результаты не противоречат другим исследованиям, которые показывают, что большинство, если не все основные нейроны MNTB, получают глицинергический вход. Скорее, похоже, что многие, если не все нейроны MNTB получают тормозные входы от VNTB, и, кроме того, меньшее подмножество нейронов MNTB получает второй ингибирующий аминокислотный вход, который исходит изнутри (то есть от другого нейрона MNTB).
Соображения по поводу развития
Иннервация основных клеток MNTB возбуждающими глутаматергическими окончаниями из VCN начинается во время эмбриональной стадии на E17 [42,43].Тонотопная организация VCN достигается на E15.5 [44]. Следовательно, некоторая синаптическая активность может происходить уже на этой ранней стадии развития MNTB. Следующая большая волна изменений в физиологии MNTB совпадает с формированием чашечки около P2 [42,45]. Дальнейшее развитие клеток MNTB происходит в стадии, совпадающие с развитием чашечки Held, в результате чего P14 приобретает зрелые физиологические свойства [42,46]. В то время как о паттернах развития тормозных проекций в главные нейроны MNTB известно меньше, тормозные токи, вызываемые пресинаптическими чашечками, по-видимому, стабильны в развитии около P14 [47].Эффекты MNTB для LSO проходят длительный период созревания, в течение которого происходит некоторое сокращение терминалов, установленных в LSO, на срок до трех недель постнатально [48], хотя прогнозы от MNTB к LSO сами устанавливаются в течение неонатального периода и остаются от в этот раз и далее. Эффекты MNTB к MSO проявляют физиологически зрелые свойства к возрасту P17 [49]. Обсуждаемая здесь работа предполагает, что все проекции между ядрами слухового ствола мозга устанавливаются и стабильны задолго до начала слуха, хотя некоторое сокращение и созревание физиологических свойств все еще происходит в течение первой недели после начала слуха.Важно отметить, что в литературе нет свидетельств того, что целые проекции удаляются после появления слуха.
Результаты, представленные в нашем исследовании, основаны на анализе срезов ствола мозга песчанок в возрасте от P13 до P17 (то есть в течение первой недели после появления слуха (таблица 1)), что делает весьма маловероятным, что описанные коллатерали MNTB-MNTB здесь может быть преходящим с точки зрения развития. Тем не менее, мы не можем исключить возможность того, что описанные здесь терминалы могут еще претерпеть незначительные морфологические изменения.
Значение тормозных входов в нейроны MNTB
Наши данные не касаются функционального значения этих тормозных входов, но мы отмечаем, что практически все нейроны MNTB демонстрируют паттерн тонического возбуждения при стимуляции звуком in-vivo [18,23,50–52], даже если есть высокая вероятность того, что они получат, по крайней мере, внешний тормозной вход от VNTB [20]. Это говорит о том, что этот тормозящий вход может не полностью подавлять возбуждение MNTB, а скорее взаимодействовать с возбуждением чашечки более тонким и специфическим образом.Повторяющийся тормозной коллатераль, как описано здесь, может быть активирован, когда тонически активирующая основная клетка, из которой он возникает, запускает серию потенциалов действия на время предъявления звукового стимула. Затем эти шипы будут перемещаться по исходящему аксону и мгновением позже достичь пресинаптического терминала иннервируемой клетки-мишени. Поскольку эти тормозящие коллатерали иннервируют близлежащие основные клетки, предположительно с аналогичными лучшими характеристическими частотами, мы постулируем, что физиологические свойства коллатералей, описанные здесь, должны включать немного более длительную латентность, чем соответствующее возбуждение чашечек.Кроме того, торможение должно обладать тоническими свойствами, поскольку нейроны MNTB обладают тоническими возбуждающими свойствами и должны быть настроены на частоты либо немного ниже, либо немного выше, чем настроен возбуждающий сигнал основного нейрона. Этим критериям соответствует процесс бокового торможения, который может служить, например, для повышения частотной избирательности. Несколько авторов [9,24,52] описали форму бокового торможения, которая хорошо отображается на свойствах, описанных здесь. Однако мы не уверены, почему только небольшое подмножество нейронов MNTB должно иметь такое латеральное торможение.
В качестве альтернативы, клетки, с которыми связываются возвращающие коллатерали, могут фактически реагировать на одну и ту же звуковую частоту (например, при подавлении наилучшей частоты), а не только на аналогичную звуковую частоту. В этом случае физиологическая роль коллатералей может заключаться в увеличении временной точности звукового кодирования, например, в усилении начальных ответов нейронов MNTB на звуковые стимулы и / или в увеличении фазовой синхронизации ответов на продолжающуюся стимуляцию [53]. Будущие эксперименты in-vivo в сочетании с фармакологическими манипуляциями с ингибированием могут проверить этот сценарий.
Сводка
Таким образом, мы продемонстрировали, что эфферентные аксоны примерно от четверти до трети основных нейронов MNTB образуют возвращающиеся тормозные коллатерали на близлежащие основные нейроны MNTB. Будущие исследования определят функциональное значение этих коллатералей и то, как этот тип ингибирующего входа взаимодействует с ранее продемонстрированным ингибирующим входом от ипсилатерального VNTB.
Вспомогательная информация
S1 Рис. Необработанные данные: конфокальный стек из 54 виртуальных секций, собранных на Olympus FV1000, UPLSAPO obj.60X Вт, 1,2 Н. А.
Разрешение изображения 1024×1024, 16 бит, размер вокселя: x, y, z = 0,207 x 0,207 x 0,4 мкм 3 . Метка биоцитина визуализирована с помощью Extravidin TRITC. Этот вспомогательный файл можно открыть с помощью Fiji (Fiji-win64-20140602), который является дистрибутивом imageJ (NIH) и включает плагин Bio-Formats (http://imagej.net/Fiji/Downloads).
https://doi.org/10.1371/journal.pone.0160241.s001
(ZIP)
S2 Рис. Необработанные данные: конфокальный стек из 48 виртуальных секций, собранных на Olympus FV1000, UPLSAPO obj.60X Вт, 1,2 Н. А.
Разрешение изображения 1024×1024, 16 бит, размер вокселя: x, y, z = 0,09 x 0,09 x 0,6 мкм 3 . Метка биоцитина визуализирована с помощью Extravidin TRITC. Этот вспомогательный файл можно открыть с помощью Fiji (Fiji-win64-20140602), который является дистрибутивом imageJ (NIH) и включает плагин Bio-Formats (http://imagej.net/Fiji/Downloads).
https://doi.org/10.1371/journal.pone.0160241.s002
(ZIP)
S3 Рис. Необработанные данные: конфокальный стек из 19 виртуальных секций, собранных на Olympus FV1000, UPLSAPO obj.60X Вт, 1,2 Н. А.
Разрешение изображения 1024×1024, 16 бит, размер вокселя: x, y, z = 0,207 x 0,207 x 0,7 мкм 3 . Метка биоцитина визуализирована с помощью Extravidin TRITC. Этот вспомогательный файл можно открыть с помощью Fiji (Fiji-win64-20140602), который является дистрибутивом imageJ (NIH) и включает плагин Bio-Formats (http://imagej.net/Fiji/Downloads).
https://doi.org/10.1371/journal.pone.0160241.s003
(TIF)
S4 Рис. Необработанные данные: конфокальный стек из 19 виртуальных секций, собранных на Olympus FV1000, UPLSAPO obj.60X Вт, 1,2 Н. А.
Разрешение изображения 1024×1024, 16 бит, размер вокселя: x, y, z = 0,207 x 0,207 x 0,7 мкм 3 . Первичное антитело мышиное моноклональное антитело против гефирина от Synaptic Systems, каталожный № 147011, визуализированное вторичным антителом козьего антимышиного антитела, конъюгированным с Alexa Fluor 488, Invitrogen / Molecular Probes, кат. № A11029. Этот вспомогательный файл можно открыть с помощью Fiji (Fiji-win64-20140602), который является дистрибутивом imageJ (NIH) и включает плагин Bio-Formats (http: // imagej.net / Fiji / Downloads).
https://doi.org/10.1371/journal.pone.0160241.s004
(TIF)
Благодарности
При поддержке NIH / NIDCD R01 DC 011582 по A.K., и частично грантом Центра неврологических заболеваний Скалистых гор NIH P30NS048154. Эксперименты по визуализации проводились в центре усовершенствованной световой микроскопии медицинского кампуса Университета Колорадо Аншутц при частичной поддержке NIH / NCRR Colorado CTSI Grant Number UL1 RR025780. Мы также хотели бы поблагодарить Dr.Тому Фингеру за полезные обсуждения и доктору Дженнифер Стратфорд за то, что она поделилась своим опытом в области нейроанатомического отслеживания и иммуногистохимических методов. Доктора Элизабет МакКаллаг и Дэниел Толлин предоставили полезные советы по улучшению более ранних версий этой рукописи.
Вклад авторов
- Концептуализация: AD AK.
- Формальный анализ: AD JAT.
- Финансирование: AK.
- Исследование: AD.
- Методология: AD.
- Администрация проекта: AD.
- Ресурсы: АК.
- Программное обеспечение: JAT.
- Проверка: AD.
- Визуализация: AD JAT.
- Написание — первоначальный черновик: AD.
- Написание — просмотр и редактирование: AD JAT AK.
Ссылки
- 1. Томпсон AM, Шофилд BR.Афферентные проекции верхнего оливкового комплекса. Microsc Res Tech. 2000. 51: 330–54. pmid: 11071718
- 2. Grothe B, Pecka M, McAlpine D. Механизмы локализации звука у млекопитающих. Physiol Rev.2010; 90: 983–1012. pmid: 20664077
- 3. Ашида Г., Карр CE. Звуковая локализация: Джеффресс и не только. Текущее мнение в нейробиологии. 2011. С. 745–751. pmid: 21646012
- 4. Borst JGG, Сориа ван Хов Дж. Чашечка удерживаемого синапса: от модельного синапса к слуховой реле.Annu Rev Physiol. 2012; 74: 199–224. pmid: 22035348
- 5. Morest DK. Коллатеральная система медиального ядра трапециевидного тела кошки, ее нейрональная архитектура и отношение к оливо-улитковому пучку. Brain Res. 1968; 9: 288–311. pmid: 5679830
- 6. Варр Брюс. Дегенерация волокон после поражений в области мультиполярных и глобулярных клеток вентрального ядра улитки кошки. Brain Res. 1972; 40: 247–270. pmid: 5027165
- 7.Фриауф Э., Оствальд Дж. Расходящиеся проекции физиологически охарактеризованных нейронов вентрального кохлеарного ядра крысы, как показано внутриаксональной инъекцией пероксидазы хрена. Exp Brain Res. 1988. 73: 263–284. pmid: 3215304
- 8. Спиру Джа, Браунелл В.Е., Зиданик М. Записи трапециевидного тела кошки и маркировка HRP аксонов шаровидных кустистых клеток. J Neurophysiol. 1990; 63: 1169–90. Доступно: http://www.ncbi.nlm.nih.gov/pubmed/2358868 pmid: 2358868
- 9. Кувабара Н., ДиКаприо Р.А, Зук Дж., Кувабара Н., Ди Каприо Ра, Зук Дж. М.. Афференты к медиальному ядру трапециевидного тела и их коллатеральным выступам. J Comp Neurol. 1991; 314: 684–706. pmid: 1816271
- 10. Смит PH, Joris PX, Карни LH, Инь TC. Проекции физиологически охарактеризованных аксонов шаровидных пушистых клеток из ядра улитки кошки. J Comp Neurol. 1991; 304: 387–407. pmid: 2022755
- 11. Held H. Die centrale gehörleitung. Arch Anat Physiol Anat Abt.1893;
- 12. Родригес-Контрерас А., Де Ланге Р.П., Лукассен П.Дж., Борст Дж.Г.Г. Ветвление афферентов чашечки во время постнатального развития в слуховом стволе мозга крыс. J Comp Neurol. 2006; 496: 214–28. pmid: 16538676
- 13. Харрисон Дж. М., Уорр В. Б.. Исследование ядер улитки и восходящих слуховых путей продолговатого мозга. J Comp Neurol. 1962; 119: 341–79. pmid: 13952992
- 14. Elverland HH. Восходящая и внутренняя проекции верхнего оливкового комплекса у кошки.Exp brain Res. 1978; 32: 117–34. pmid: 658183
- 15. Спанглер К.М., Варр В.Б., Хенкель К.К. Проекции основных клеток медиального ядра трапециевидного тела кошки. J Comp Neurol. 1985; 238: 249–262. pmid: 4044914
- 16. Кейси М.А., Фельдман М.Л. Старение в медиальном ядре трапециевидного тела крысы. II. Электронная микроскопия. J Comp Neurol. 1985; 232: 401–13. pmid: 3973099
- 17. Толлин DJ. Боковая верхняя олива: функциональная роль в локализации источника звука.Neurosci. 2003. 9: 127–143.
- 18. Копп-Шейнпфлюг С., Липпе В.Р., Дёрршайдт Г.Дж., Рюбсамен Р. Медиальное ядро трапециевидного тела песчанки — больше, чем реле: сравнение пре- и постсинаптической активности. J Assoc Res Otolaryngol. 2003; 4: 1–23. pmid: 12098017
- 19. Герман Дж., Пека М., фон Герсдорф Х., Гроте Б., Клуг А. Синаптическая передача в чашечке Хелда при уровнях активности, подобных in vivo. J Neurophysiol. 2007; 98: 807–20. pmid: 17507501
- 20.Альбрехт О., Дондзилло А., Майер Ф., Томпсон Дж. А., Клуг А. Тормозящие проекции из вентрального ядра трапециевидного тела в медиальное ядро трапециевидного тела у мыши. Передние нервные цепи. 2014; 8: 83. pmid: 25120436
- 21. Аватрамани Г.Б., Туречек Р., Трассел Л.О. Тормозящее управление синаптическим реле. J Neurosci. 2004. 24: 2643–2647. pmid: 15028756
- 22. Грин JS, Санес DH. Раннее появление ингибирующего входа в MNTB поддерживает бинауральную обработку во время разработки.J Neurophysiol. 2005; 94: 3826–3835. pmid: 16120660
- 23. Смит PH, Joris PX, Yin TC. Анатомия и физиология основных клеток медиального ядра трапециевидного тела (МНТТ) кошки. J Neurophysiol. 1998. 79: 3127–3142. pmid: 9636113
- 24. Копп-Шейнпфлюг С., Демель С., Толнаи С., Дитц Б., Миленкович И., Рюбсамен Р. Глицин-опосредованные изменения надежности начала в центральном синапсе млекопитающих. Неврология. ИБРО; 2008; 157: 432–445.
- 25.Helfert RH, Bonneau JM, Wenthold RJ, Altschuler RA. Иммунореактивность ГАМК и глицина в высшем оливковом комплексе морской свинки. Brain Res. 1989; 501: 269–86. pmid: 2819441
- 26. Лу Т, Рубио МЭ, Трассел ЛО. Глицинергическая передача, сформированная Corelease ГАМК в слуховом синапсе млекопитающих. Нейрон. 2008. 57: 524–535. pmid: 18304482
- 27. Аватрамани Г.Б., Туречек Р., Трассел Л.О. Поэтапное развитие ГАМКергической и глицинергической передачи в MNTB.J Neurophysiol. 2005; 93: 819–828. pmid: 15456797
- 28. Бэнкс М.И., Смит PH. Внутриклеточные записи от нейробиотин-меченых клеток в срезах мозга медиального ядра трапециевидного тела крысы. J Neurosci. 1992; 12: 2819–2837. Доступно: http://www.ncbi.nlm.nih.gov/pubmed/1351938 pmid: 1351938
- 29. Mayer F, Albrecht O, Dondzillo A, Klug A. Глицинергическое ингибирование медиального ядра трапециевидного тела демонстрирует заметное облегчение и может поддерживать высокий уровень постоянной активности.J Neurophysiol. 2014; 112: 2901–15. Доступно: http://www.ncbi.nlm.nih.gov/pubmed/25185813 pmid: 25185813
- 30. Pfeiffer F, Simler R, Grenningloh G, Betz H. Моноклональные антитела и пептидное картирование выявляют структурное сходство между субъединицами глицинового рецептора спинного мозга крысы. Proc Natl Acad Sci U S. A. 1984; 81: 7224–7. pmid: 6095276
- 31. Schmitt B, Knaus P, Becker CM, Betz H. Полипептид Mr 93000 постсинаптического рецепторного комплекса глицина представляет собой белок периферической мембраны.Биохимия. 1987; 26: 805–11. pmid: 3032237
- 32. Becker CM, Hoch W., Betz H. Чувствительный иммуноанализ показывает избирательную ассоциацию периферических и интегральных мембранных белков ингибиторного комплекса рецепторов глицина. J Neurochem. 1989. 53: 124–31. pmid: 2470857
- 33. Кирш Дж, Бец Х. Широко распространенная экспрессия гефирина, предполагаемого линкерного белка рецептора глицина-тубулина, в мозге крысы. Brain Res. 1993; 621: 301–10. pmid: 8242343
- 34. Фишер Ф., Кнейссель М., Тинтруп Х, Хаверкамп С., Рауэн Т., Бец Х. и др.Снижение синаптической кластеризации рецепторов ГАМК и глицина в сетчатке мышей с нулевым мутантом по гефирину. J Comp Neurol. 2000; 427: 634–648. Доступно: http://www.ncbi.nlm.nih.gov/pubmed/11056469 pmid: 11056469
- 35. Ойлер Г.А., Хиггинс Г.А., Харт Р.А., Баттенберг Э., Биллингсли М., Блум Ф.Е. и др. Идентификация нового белка, ассоциированного с синаптосомами, SNAP-25, дифференциально экспрессируемого субпопуляциями нейронов. J Cell Biol. 1989; 109: 3039–52. pmid: 2592413
- 36.Blasi J, Chapman ER, Link E, Binz T, Yamasaki S, De Camilli P и др. Ботулинический нейротоксин А избирательно расщепляет синаптический белок SNAP-25. Природа. 1993; 365: 160–163. pmid: 8103915
- 37. Зёлльнер Т., Уайтхарт С.В., Бруннер М., Эрдьюмент-Бромаж Х., Героманос С., Темпст П. и др. Рецепторы SNAP участвуют в нацеливании и слиянии везикул. Природа. 1993; 362: 318–324. pmid: 8455717
- 38. Триллер А., Клюзо Ф., Пфайфер Ф., Бец Х., Корн Х. Распределение рецепторов глицина в центральных синапсах: исследование с помощью иммуноэлектронной микроскопии.J Cell Biol. 1985. 101: 683–688. pmid: 2991304
- 39. Альтшулер Р.А., Бец Х., Параккал М.Х., Рикс К.А., Вентхольд Р.Дж. Идентификация глицинергических синапсов в ядре улитки посредством иммуноцитохимической локализации постсинаптического рецептора. Brain Res. 1986; 369: 316–320. Доступно: http://www.ncbi.nlm.nih.gov/pubmed/3008938 pmid: 3008938
- 40. Баер К., Вальдфогель Х. Дж., Во время MJ, Снелл Р. Г., Фаулл Р. Л. М., Рис М. И.. Ассоциация гефириновых и глициновых рецепторов в стволе и спинном мозге человека: иммуногистохимический анализ.Неврология. 2003. 122: 773–784. pmid: 14622920
- 41. Guinan JJ, Li RY. Обработка сигналов в слуховых нейронах ствола мозга, которые получают гигантские окончания (чашечки Held) в медиальном ядре трапециевидного тела кошки. Послушайте Res. 1990; 49: 321–34. pmid: 2292504
- 42. Hoffpauir BK, Kolson DR, Mathers PH, Spirou G. a. Созревание синаптических партнеров: функциональный фенотип и синаптическая организация настроены синхронно. J Physiol. 2010. 588: 4365–4385. pmid: 20855433
- 43.Hoffpauir BK, Marrs GS, Mathers PH, Spirou GA. Соединяется ли мозг до того, как периферия сможет управлять? Сравнение трех сенсорных систем у мышей. Brain Res. Elsevier B.V .; 2009; 1277: 115–129.
- 44. Маклад А., Фрич Б. Развитие вестибулярных афферентных проекций в задний мозг и их центральные цели. Brain Res Bull. 2003. 60: 497–510. pmid: 12787869
- 45. Кандлер К., Фриауф Э. Пре- и постнатальное развитие эфферентных связей ядра улитки у крысы.J Comp Neurol. 1993; 328: 161–184. pmid: 8423239
- 46. Ташенбергер Х., фон Герсдорф Х. Точная настройка слухового синапса для обеспечения скорости и точности: онтогенетические изменения пресинаптической формы волны, кинетики EPSC и синаптической пластичности. J Neurosci. 2000; 20: 9162–9173. 20/24/9162 [pii] pmid: 11124994
- 47. Турецек Р., Трассел Л.О. Взаимная регуляция развития пресинаптических ионотропных рецепторов. Proc Natl Acad Sci U S. A. 2002; 99: 13884–9. pmid: 12370408
- 48.Санес Д.Х., Сиверлс В. Развитие и специфичность тормозных терминальных ветвлений в центральной нервной системе. J Neurobiol. 1991; 22: 837–54. pmid: 1663990
- 49. Магнуссон А.К., Капфер С., Гроте Б., Кох У. Созревание глицинергического ингибирования в медиальной верхней оливе песчанки после появления слуха. J Physiol. 2005; 568: 497–512. pmid: 16096336
- 50. Sommer I, Lingenhöhl K, Friauf E. Основные клетки медиального ядра трапециевидного тела крысы: внутриклеточное исследование их физиологии и морфологии in vivo.Exp brain Res. 1993; 95: 223–39. pmid: 8224048
- 51. Lorteije J a M, Rusu SI, Kushmerick C, Borst JGG. Надежность и точность чашечки синапса Хельда мыши. J Neurosci. 2009. 29: 13770–13784. pmid: 19889989
- 52. Koka K, Tollin DJ. Линейное кодирование сложных звуковых спектров по скорости разряда в нейронах медиального ядра трапециевидного тела (MNTB) и его входов. Передние нервные цепи. 2014; 8: 144. pmid: 25565971
- 53. Толлин DJ, Инь TCT.Интеравуральная чувствительность по фазе и разности уровней в низкочастотных нейронах латеральной верхней оливы. J Neurosci. 2005. 25: 10648–57. pmid: 16291937
Перинатальное развитие медиального ядра трапециевидного тела
Шобхана ШиварамакришнанШобхана Сиварамакришнан, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Отделение отоларингологии HNS, Университет Западной Вирджинии
Эшли БрандебураЭшли Брандебура, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Аспирантура по биохимии и молекулярной биологии; Кафедра биохимии, Университет Западной Вирджинии
Пол ХолкомбПол Холкомб, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Аспирантура по неврологии, Университет Западной Вирджинии
Дэниел ХеллерДаниэль Хеллер, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Аспирантура по неврологии, Университет Западной Вирджинии
Дуглас КолсонДуглас Колсон, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии, Университет Западной Вирджинии,
Дакота ДжексонДакота Джексон, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Аспирантура по неврологии, Университет Западной Вирджинии
Питер Х.MathersПитер Х. Мазерс, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Кафедра биохимии; Отделение отоларингологии HNS; Кафедра офтальмологии, Университет Западной Вирджинии
Джордж А. СпируДжордж А. Спиру, Институт неврологии Бланшетт Рокфеллер; Исследовательский центр сенсорной неврологии; Отделение отоларингологии HNS, Университет Западной Вирджинии
Для доступа к полному контенту Oxford Handbooks Online требуется подписка или покупка.Общедоступные пользователи могут искать на сайте и просматривать аннотации и ключевые слова для каждой книги и главы без подписки.
Пожалуйста, подпишитесь или войдите, чтобы получить доступ к полному тексту.
Если вы приобрели печатное издание, содержащее токен доступа, просмотрите этот токен для получения информации о том, как зарегистрировать свой код.
По вопросам доступа или устранения неполадок, пожалуйста, ознакомьтесь с нашими часто задаваемыми вопросами, а если вы не можете найти там ответ, свяжитесь с нами.
31 июля, 2017 г. Дипра Рэй — генеральный директор mPort
Трапециевидная форма тела считается идеальной формой мужского тела.Благодаря широким плечам, массивным рукам и узким бедрам тело хорошо сложено и часто встречается у спортсменов.
Типичные характеристики мужского трапециевидного тела
- Верхняя часть туловища больше нижней
- Плечи и грудь широкие
- Талия и бедра средние узкие
- Тело правильные пропорции
Цели мужчины трапециевидной формы тела
Ваш тип телосложения — один из самых простых в одежде, вы можете выбирать из самого большого разнообразия стилей.
Вы всегда можете поискать идеи стиля у знаменитостей, которые разделяют вашу трапециевидную форму тела: Хью Джекмана, Мэтью МакКонахи, Герада Батлера, Дэниела Крейга.
Не забывайте учитывать вертикальную форму тела, а также другие особенности, такие как возраст, вес, форму лица, длину шеи и т. Д.
Костюмы для мужчин трапециевидной формы
Если у вас трапециевидная фигура и к тому же у вас довольно высокий рост, вам следует выбрать пиджак с тремя пуговицами, так как он будет гармонировать с вашей первоначальной формой и ростом, длина этого пиджака будет соответствовать естественным пропорциям вашего тела.
В связи с тем, что вам не нужен пиджак, чтобы создать впечатление стройной фигуры, вы можете выбрать пиджак с одним отверстием или без него, это зависит от вашего личного вкуса.
Также, поскольку у вас довольно длинные ноги, вам не нужны штаны, которые создают иллюзию того, что вы выше, вместо этого вы хотите предотвратить это. Поэтому вам следует выбрать брюки с регулярной посадкой, это расстояние между поясом и промежностью, и это уравновесит вашу длинную ногу и туловище, чтобы ваше тело выглядело пропорционально.
Дополнительные советы о том, как подобрать одежду для вашей фигуры, можно найти в статье ниже:
Подходит для вашей фигуры — что подходит лучше всего.
Узнайте форму своего тела с помощью mPort
Познакомьтесь со своим телом с помощью трехмерной карты тела mPort. Это самый простой способ видеть свои измерения и следить за своим здоровьем. Найдите ближайшую к вам капсулу для картирования тела на сайте mPort.com.
Подробнее о:
Для получения дополнительной информации о формах тела.
Какая у вас форма тела?
Теперь очередь парней узнать свой тип телосложения. Для дам ознакомьтесь с нашим первым постом о типе телосложения, чтобы узнать, на какую форму или формы может напоминать ваше тело.
Мужское тело отличается от женского по мышечной массе и пропорциям. Как правило, у мужчин плечи шире, а бедра уже, чем у женщин, хотя, как и во всем, что связано с типами телосложения, всегда есть исключения.
Знание формы своего тела может помочь вам научиться правильно тренироваться с максимальной пользой.Это также может помочь вам подобрать одежду, которая отлично смотрится и подходит к футболке.
Господа, заводите двигатели
Встаньте перед вертикальным зеркалом в полный рост и обратите внимание на взаимосвязь между вашими плечами, талией и бедрами как на ориентир для вашей истинной формы.
Примечание: идеального телосложения не существует. Цель каждого тела — сбалансировать элементы. В зависимости от того, какие уникальные элементы вы принесете на стол, разная одежда и тренировки подойдут вам больше, чем другие.Ознакомьтесь со следующими типами и узнайте больше о личном теле, которое принадлежит вам!
1. Трапеция: люди с трапециевидным типом телосложения — это пропорциональные люди со средней или узкой талией и бедрами, а также широкими плечами и грудью. Верхняя часть туловища обычно больше, чем нижняя часть туловища, что можно продемонстрировать при поднятии тяжестей. Этот тип может выбирать из множества типов одежды, так как широкий выбор мужских стилей хорошо подойдет к этому телосложению.
2. Перевернутый треугольник: эта форма часто ассоциируется с бодибилдерами (вспомните Губернатора около 1980 года) и включает в себя широкую грудь и широкие плечи, сочетающиеся с развитыми мышцами груди, рук и плеч.Талия и бедра у этого типа обычно узкие. Ключевым моментом является создание пропорционального телосложения для физического комфорта и хорошо подогнанная одежда, поэтому обязательно включайте упражнения для нижней части тела, такие как приседания и выпады с отягощением, чтобы помочь вам нарастить мышцы нижней части тела.
3. Прямоугольник: самый простой для описания, этот тип включает грудь, талию и бедра примерно одинаковой ширины в сочетании с прямым туловищем. Во время тренировки сосредоточьтесь на верхней части тела и подчеркните плечи, чтобы улучшить свой обтекаемый вид.
4. Треугольник: Мужчины с такой формой имеют больший вес в нижней половине и имеют грудь, которая уже, чем их бедра. Хорошая новость заключается в том, что мужчины с таким типом телосложения могут в полной мере воспользоваться яркими модными вещами, такими как жакеты с квадратными плечами. Тренировки для верхней части тела могут помочь вам сбалансировать вашу фигуру, но не забудьте также проработать ноги, чтобы в полной мере использовать силу нижней части тела.
5. Овальный: Если ваша внешность имеет тенденцию быть круглой, с ярко выраженным животом, ваш типаж — овал.Многие люди в процессе похудения имеют овальную форму и трансформируются в одну из вышеперечисленных форм после того, как значительно похудеют. Другие сохраняют такую форму при любом размере.
Во время тренировки сосредоточьтесь на укреплении кора, чтобы уменьшить давление, которое большой живот оказывает на спину. Ожидайте более четких форм брюшного пресса, избавляясь от жира, чтобы раскрыть свои с трудом заработанные мышцы, и обязательно наслаждайтесь широким спектром упражнений для верхней части тела. Одна из лучших тренировок для овальных форм в пилатесе, которая удлиняет мышцы для эффекта удлинения.
Великий Сократ провозгласил, что нужно «познать самого себя». Большая часть познания самих себя — это понимание нашего физического, умственного и эмоционального состояния. Понимая, как пропорционально ваше тело, вы сможете оценить свои успехи в похудании еще больше, имея реалистичные ожидания относительно того, как ваша фигура будет выглядеть как в тренажерном зале, так и в одежде. Удачи и приятного времяпровождения!
Сравнение пре- и постсинаптической активности
| Копп-Шейнпфлюг, Конни; Липпе, В.Р.; Dörrscheidt, G.J .; Рюбсамен, Р. (2003): Медиальное ядро трапециевидного тела песчанки — это больше, чем просто реле: сравнение пре- и постсинаптической активности. В: Журнал Ассоциации исследований в области отоларингологии, Vol. 4, No. 1: с. 1-23 |
Полный текст недоступен в Open Access LMU.
Abstract
Медиальное ядро трапециевидного тела (MNTB) играет важную роль в обработке межзубных различий интенсивности, что является критическим для локализации источников звука.Обычно считается, что MNTB функционирует в первую очередь как пассивный ретранслятор при преобразовании возбуждающего сигнала, исходящего от контралатерального ядра улитки (CN), в тормозящий сигнал на ипсилатеральную боковую верхнюю оливу. Однако исследования, показывающие, что сам MNTB также является мишенью для ингибирующего воздействия, предполагают, что MNTB может выполнять больше, чем функцию преобразования знаков. Чтобы проверить точность передачи сигнала в синапсе CN-MNTB, пресинаптические потенциалы чашечки («препотенциалы»), отражающие возбуждающий вход в нейрон MNTB, и постсинаптические потенциалы действия одновременно отслеживались с помощью одного и того же электрода во время внеклеточных записей in vivo с MNTB песчанки.Пресинаптическая активность отличалась от постсинаптической активности в нескольких отношениях: (1) частота спонтанных и вызванных звуком разрядов была больше пресинаптически, чем постсинаптически. (2) Постсинаптическая перестройка частоты была более резкой, чем пресинаптическая. (3) Терминалы чашечки и нейроны MNTB показали фазико-тонические паттерны ответа на тональную стимуляцию, но продолжительность начального ответа и уровень тонического компонента были постсинаптически сокращены. (4) Фазовая синхронизация звуковых частот до 1 кГц была больше постсинаптически, чем пресинаптически.(5) Скоростно-интенсивные характеристики пре- и постсинаптической активности достоверно отличались друг от друга в половине нейронов MNTB. Чтобы проверить гипотезу о том, что акустически вызванное ингибирование нейронов MNTB способствовало относительно более низкому уровню постсинаптического разряда, была применена двухтональная стимуляция, при которой ответ на один тональный сигнал, установленный на характеристической частоте нейрона, может быть уменьшен путем добавления второй «тормозной» тон. Тормозной тон вызывал гораздо большее снижение пост-, чем пресинаптической активности, указывая на акустически вызванное тормозящее влияние непосредственно на единицы MNTB.Эти находки показывают, что передача в синапсе CN-MNTB не происходит фиксированным однозначным образом и что ответ нейронов MNTB отражает интеграцию их возбуждающих и тормозных входов.
Тело в форме трапеции — исследования науки и экспертов
электрические мембранные свойства
Трапеция Тело нейронов слухового ствола головного мозга крысы сохраняются в органотипических культурах срезовЖурнал нейробиологии, 1998 г.
Соавторы: Стефан Лорке, Мартин Кунгель, Экхард Фриауф
Абстрактный:
Медиальное ядро трапеции Тело (MNTB) представляет собой заметную структуру в слуховом стволе мозга млекопитающих.Это главный компонент высшего оливкового комплекса и участвует в локализации звука. Недавно были введены органотипические препараты культуры срезов верхнего оливкового комплекса для исследования развития тормозных и возбуждающих проекций (Sanes and Hafidi, 1996; Lohmann et al., 1998). В настоящей статье мы дополнительно оценили органотипичность нашей системы культивирования (Lohmann et al., 1998) и изучили электрические свойства мембран нейронов MNTB, экспрессируемых в условиях культивирования.Для этого нейроны MNTB от крыс в раннем постнатальном периоде (P3–5) изучали через 3–6 дней in vitro (DIV) с помощью записи патч-кламп целых клеток. Их средний потенциал покоя составлял -59 мВ, входное сопротивление составляло в среднем 171 МОм, а средняя постоянная времени составляла 3 мс. При записи с фиксацией напряжения наблюдались четыре типа активируемых напряжением проводимости. Все клетки экспрессировали чувствительный к тетродотоксину (ТТХ) натриевый ток. Можно охарактеризовать два типа калиевых токов: чувствительность к тетраэтиламмонию (TEA) и чувствительность к 4-аминопиридину (4-AP), оба из которых состоят из переходного и длительного компонентов.Наконец, был обнаружен ток выпрямления внутрь, активируемый ступенями гиперполяризационного напряжения. При записи с зажимом тока деполяризующие импульсы тока обычно вызывают единичный потенциал действия. Однако в присутствии 4-АР эти импульсы тока вызывали серию потенциалов действия. Продолжительность потенциалов действия увеличивалась с помощью 4-AP, а постгиперполяризация уменьшалась.