Боковые стенки корпуса: строение и упражнения
Боковые стенки корпуса состоят из нескольких видов косых мышц, которые раcположены слоями. Они работают вместе с передней стенкой корпуса, и тренировать их тоже надо вместе.
1️⃣ Наружные косые мышцы. Как и все мышцы боковой части кора, наружные мышцы живота парные — есть правая и левая мышца. Наружные косые мышцы находятся поверх других косых мышц и идут от боковых частей нижних ребер к тазовым костям. Волокна мышц идут снаружи внутрь и сверху вниз. Когда сокращается правая или левая мышца, корпус наклоняется в сторону. Когда сокращаются обе мышцы, они помогают наклонять корпус вниз: например, делать скручивания.
Каждая мышца состоит из двух частей:
🔸 Мышечная часть. В районе ребер расположена мышечная часть наружной мышцы. Она состоит из красных волокон, которые могут сокращаться.
🔸 Сухожильная часть. Ближе к пупку мышца превращается в сухожильную пластину.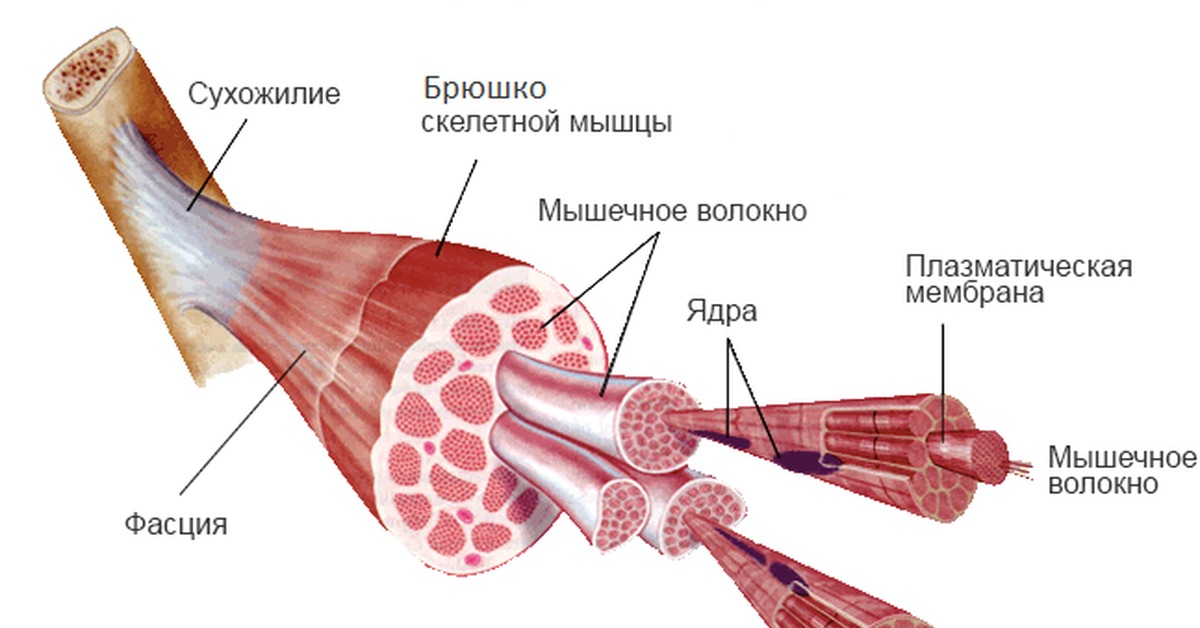
2️⃣ Внутренние косые мышцы. Внутренние косые мышцы также парные и находятся внутри — под наружными. Они устроены наоборот — идут снизу вверх, от тазовых костей к ребрам. Волокна также направлены в обратную сторону. А вот функция у них такая же: наклонять корпус в сторону работой одной мышцы или вниз работой обеих мышц.
3️⃣ Поперечные мышцы живота.
🐌 Особенность косых мышц в том, что они только наполовину состоят из мышечной ткани.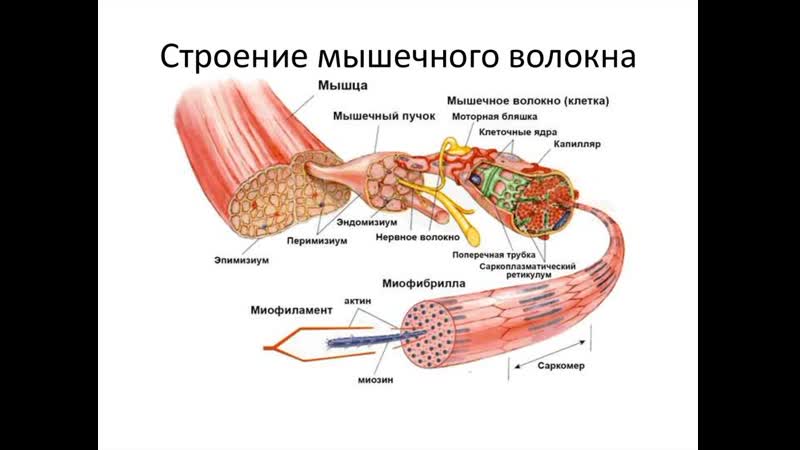 Вторая половины мышцы состоит из сухожильной пластины, которое помогает нам выдерживать нагрузку. Косые мышцы похожи на панцирь — они поддерживают внутрибрюшное давление и защищают внутренние органы от повреждений. Поэтому косые мышцы не способны сокращаться так долго или мощно, как мышцы ног или рук — у них другая задача.
Вторая половины мышцы состоит из сухожильной пластины, которое помогает нам выдерживать нагрузку. Косые мышцы похожи на панцирь — они поддерживают внутрибрюшное давление и защищают внутренние органы от повреждений. Поэтому косые мышцы не способны сокращаться так долго или мощно, как мышцы ног или рук — у них другая задача.
Похожие на ДНК искусственные волокна в 36 раз сильнее сильнее мышц человека | Технологии
Исследователи из Университета Вуллонгонга (UOW) в Австралии создали искусственные мышцы, вдохновленные сверхспирализацией нитей ДНК.
ДНК — один из самых впечатляющих примеров силы сжатия в природе. Именно она позволяет скрутить нити длиной около 2 метров и поместить их в одну человеческую клетку. Для этого ДНК использует процесс суперспирализации. Напомним, сверхспирализация — явление перекручивания топологически замкнутых цепей ДНК, в результате которого ось двойной спирали ДНК сама закручивается в спираль более высокого порядка.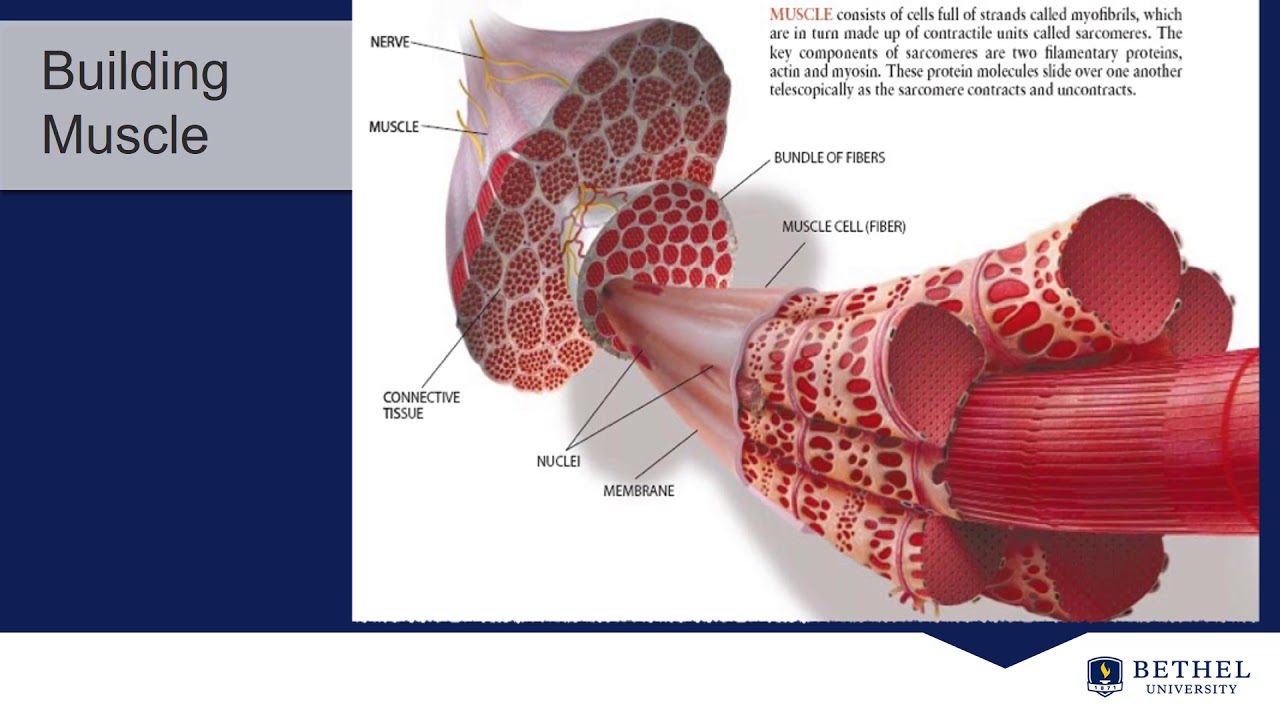
В рамках нового исследования ученые UOW воспроизвели это явление при создании искусственных мышц. Биологи сделали их из композитных полиэфирных волокон, покрытых гидрогелем, который набухает при намокании. Их скрутили в форму спирали ДНК, а затем погрузили в воду для набухания.
Обычно это приводит к распутыванию волокон. Однако ученые выяснили, что, если зажать их концы, то они подвергаются сверхспирализации. В результате нити создают относительно большое количество механической силы.
В эксперименте суперспиральные волокна сократились до 10% от своей первоначальной длины, генерируя эквивалент 1 Джоуля энергии на грамм. Механическая работа, которую может выполнять мышца из таких нитей, в 36 раз выше, чем у сопоставимой скелетной мышцы человека.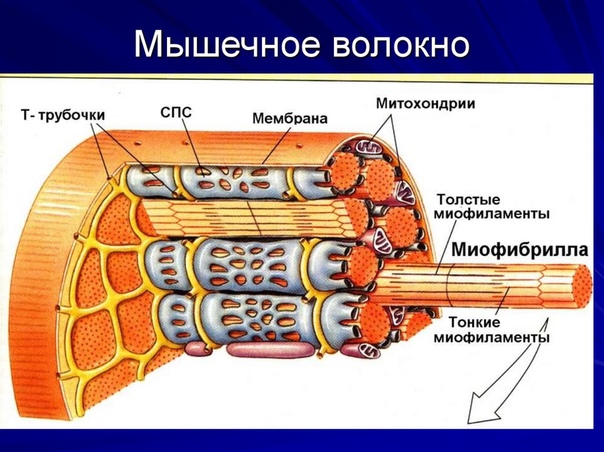
Тем не менее, пока образцы двигаются довольно медленно из-за механизма действия гидрогеля. Однако ученые уверяют, что процесс можно ускорить, изменив материалы или методы производства искусственных мышц, оставив сверхспирализацию волокн.
Читать далее
Илон Маск: первые туристы на Марс погибнут
Создана первая точная карта мира. Что не так со всеми остальными?
Обнаружена мертвая звезда, вращающаяся вокруг своей оси за секунду
Бактерии «научили» вырабатывать мышечные волокна — Наука
ТАСС, 30 августа. Ученые создали трансгенных бактерий, которые могут производить длинные волокна титина – одного из основных белковых компонентов мышц человека и других млекопитающих.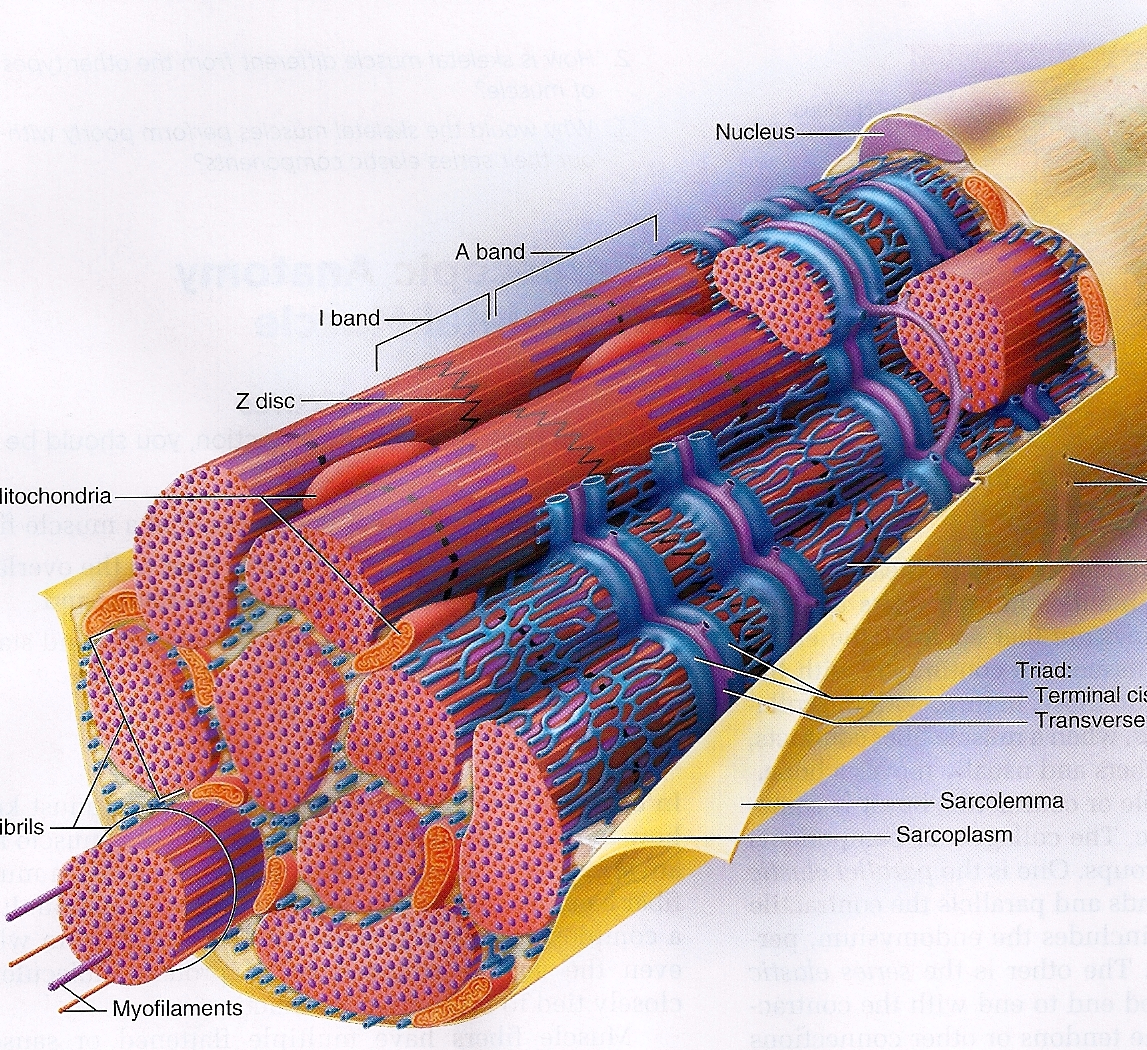
«Производство этих волокон отличается низкой стоимостью и его можно легко расширить. Их можно будет использовать в самых разных целях, в том числе для создания бронежилетов и мягких роботов, а также использования в других областях науки и техники, в которых подобные природные материалы раньше не применялись», – рассказал один из авторов работы, профессор Вашингтонского университета в Сент-Луисе Чжан Фучжун.
В последние годы ученые активно исследуют, как работают мышечные волокна и как протекает процесс их «сборки». Помимо лечения дистрофии мышц и прочих болезней опорно-двигательного аппарата, благодаря этому можно создать искусственные аналоги мышечной ткани, пригодные для использования в роботах и прочих устройствах, где необходимы высокая гибкость и сила.
Чжан и его коллеги открыли относительно простой и дешевый метод выращивания волокон титина – одного из ключевых компонентов мышц и одного из самых крупных белков человеческого организма. Его большие размеры, как отмечают биологи, выступали в прошлом главным препятствием для создания микробов, способных производить большие количества титина.
Его большие размеры, как отмечают биологи, выступали в прошлом главным препятствием для создания микробов, способных производить большие количества титина.
Бактериальная фабрика мускулов
Американские исследователи обошли эту проблему. Они модифицировали одиночные звенья титина таким образом, что теперь их могут соединять друг с другом специальные бактериальные ферменты. Они отвечают за «склеивание» сложных белковых молекул, состоящих из нескольких обособленных частей.
«Инструкции» по сборке подобных одиночных звеньев мышечного белка ученые вставили в геном кишечной палочки, после чего проследили, как будет протекать процесс сборки нитей титина внутри этих микробов. Как оказалось, бактерии успешно справились с задачей и начали производить сложные цепочки белка, примерно в 50 раз превосходящие по размерам типичные белки бактерий.
Подготовив большое число подобных молекул титина, ученые переплели их особым образом и использовали для создания искусственного аналога мышечных нитей.
Подобные свойства бактериального титина позволяют применять его для производства искусственных мышц для роботов, бронежилетов, а также хирургических нитей, не вызывающих воспалений и полностью совместимых с организмом человека. Кроме того, аналогичный подход может применяться для производства других крупных белков, потенциально полезных для практического использования, подытожили Чжан и его коллеги.
Типы волокон скелетных мышц
Типы волокон скелетных мышц
Волокна скелетных мышц не одинаковы по своим механическим и метаболическим особенностям. Типы волокон различаются на основе следующих характеристик:
— в зависимости от максимальной скорости укорочения — быстрые волокна и медленные волокна ;
— в зависимости от главного пути образования АТФ —
оксидативные волокна
и
гликолитические волокна
.
Быстрые и медленные мышечные волокна содержат изоферменты миозина , которые расщепляют АТФ с разной максимальной скоростью; этому соответствует различная максимальная скорость рабочего цикла поперечных мостиков и, следовательно, укорочения волокна. Высокая АТФазная активность миозина свойственна быстрым волокнам, более низкая — медленным волокнам. Хотя в быстрых волокнах скорость рабочего цикла примерно в четыре раза выше, чем в медленных, поперечные мостики обоих типов генерируют одинаковую силу.
Другой подход к классификации волокон скелетных мышц основан на различиях
ферментативных механизмов синтеза АТФ. В некоторых волокнах много
митохондрий
, и, следовательно, обеспечивается высокий уровень
окислительного фосфорилирования
; это
оксидативные волокна
. Количество образующейся в них АТФ зависит от снабжения мышцы кровью, с
которой поступают молекулы кислорода и богатых энергией соединений. Волокна
этого типа окружены многочисленными капиллярами.
В гликолитических волокнах , наоборот, мало митохондрий, но высокое содержание ферментов гликолиза и большие запасы гликогена . Эти волокна окружены относительно небольшим числом капилляров, и миоглобина в их ткани немного, что соответствует ограниченному использованию кислорода. Вследствие недостатка миоглобина гликолитические волокна выглядят светлыми и получили название белых мышечных волокон.
На основании двух рассмотренных характеристик (скорость укорочения и тип метаболизма) можно выделить три типа волокон скелетных мышц.
—
Медленные оксидативные волокна (тип I)
— низкая активность миозиновой АТФазы и высокая окислительная
способность.
— Быстрые оксидативные волокна (тип IIа) — высокая активность миозиновой АТФазы и высокая окислительная способность.
— Быстрые гликолитические волокна (тип IIб) — высокая активность миозиновой АТФазы и высокая гликолитическая способность.
Отметим, что не обнаружен четвертый теоретически возможный вариант — медленные гликолитические волокна.
Волокна варьируются не только по своим биохимическим особенностям, но и по
размерам: у гликолитических волокон диаметр существенно больше, чем у
оксидативных (
рис. 30.28
). Это сказывается на величине развиваемого ими
напряжения
. Число толстых и тонких
филаментов
на единицу площади поперечного сечения примерно одинаково для всех типов
скелетных мышечных волокон. Таким образом, чем значительнее диаметр
волокна, тем большее число параллельно задействованных толстых и тонких
филаментов участвует в генерировании силы и тем больше, наконец,
максимальное напряжение мышечного волокна.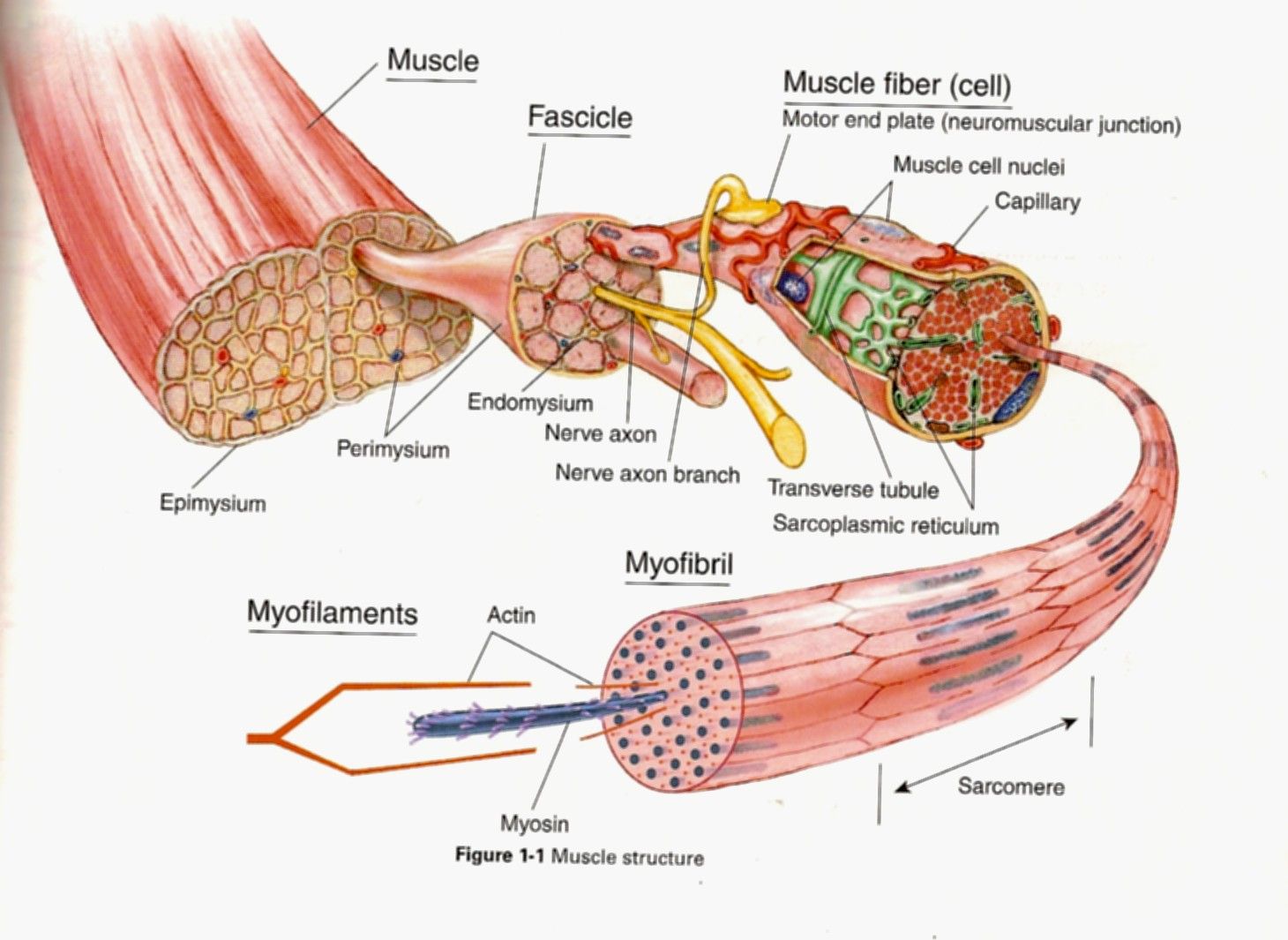 Отсюда следует, что
гликолитическое волокно
, имеющее больший диаметр, развивает в среднем более значительное
напряжение по сравнению с напряжением
оксидативного волокна
.
Отсюда следует, что
гликолитическое волокно
, имеющее больший диаметр, развивает в среднем более значительное
напряжение по сравнению с напряжением
оксидативного волокна
.
Кроме того, рассмотренные три типа мышечных волокон характеризуются разной устойчивостью к утомлению . Быстрые гликолитические волокна утомляются через короткое время, тогда как медленные оксидативные волокна очень выносливы, что позволяет им длительно поддерживать сократительную активность практически при постоянном уровне напряжения. Быстрые оксидативные волокна занимают промежуточное место по способности противостоять развитию утомления ( рис. 30.29 ).
Характеристики трех типов волокон скелетных мышц обобщены в табл. 30.3 .
Резюме.
— Различают три типа скелетных мышечных волокон в зависимости от
максимальной скорости укорочения и преобладающего способа образования АТФ:
медленные оксидативные, быстрые оксидативные и быстрые гликолитические.
— Разная максимальная скорость укорочения быстрых и медленных волокон обусловлена различиями АТФазы миозина: высокой и низкой АТФазной активности соответствуют быстрые и медленные волокна.
— Быстрые гликолитические волокна имеют в среднем больший диаметр, чем оксидативные, и потому развивают более значительное напряжение, но быстрее утомляются.
— Все мышечные волокна одной двигательной единицы принадлежат к одному и тому же типу; большинство мышц содержат все три типа двигательных единиц.
— Характеристики трех типов скелетных мышечных волокон обобщены в табл. 30.3 .
Ссылки:
10 вещей, которые нужно знать о них
Повышение тонуса или улучшение очертания мышц — одна из наиболее частых целей людей, начинающих программу упражнений. Термин «тонус» происходит от «тонуса» — технического термина, обозначающего состояние сокращения мышечного волокна. Если все волокна в мышце сокращаются, это создает форму, обычно ассоциируемую с четко определенной мышцей.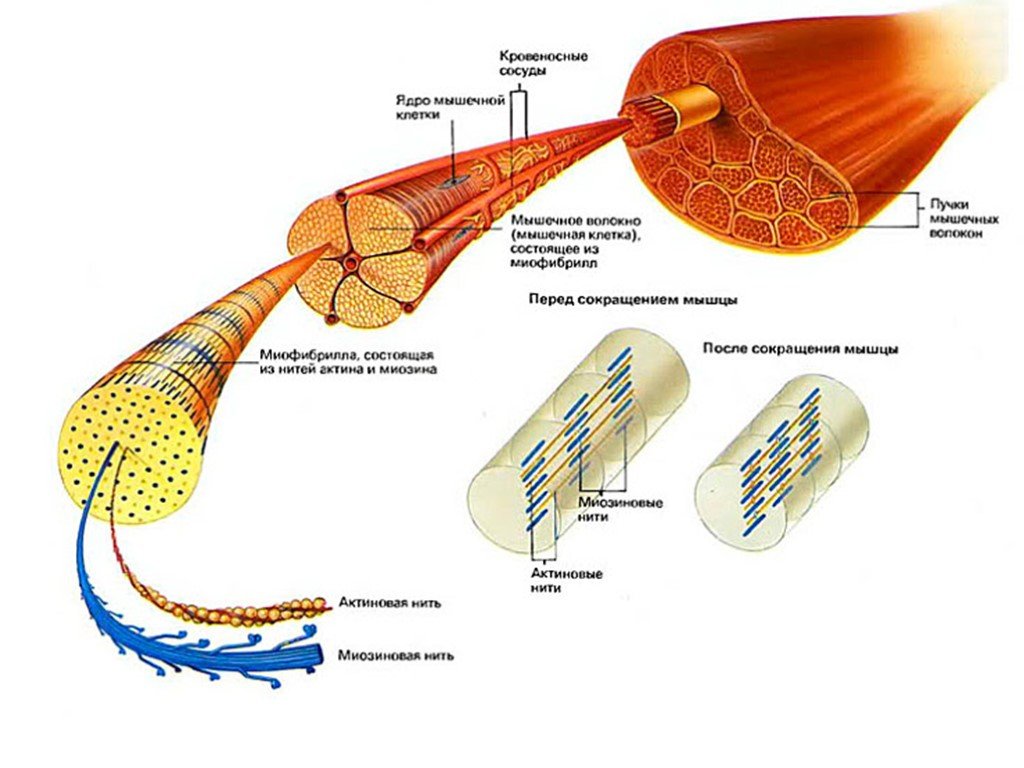
Основное назначение мышечных волокон — контролировать физические силы, движущиеся по телу.Действия по укорачиванию мышц могут генерировать силу для перемещения сопротивления; например, при переходе из положения сидя в положение стоя квадрицепсы и большая ягодичная мышца укорачиваются, чтобы помочь телу противостоять силе тяжести. Действия по удлинению мышц могут применяться для контроля и замедления силы; например, квадрицепсы и ягодицы удлиняются, чтобы контролировать движения тела, когда оно возвращается в сидячее положение. Если вы заинтересованы в разработке программ упражнений, которые приносят результат, важно хорошо понимать, как работают мышечные волокна и как они реагируют на упражнения.
Чтобы более эффективно помогать вашим клиентам в достижении их целей в фитнесе, помните следующие важные факты о мышечных волокнах и о том, как на них влияют упражнения:
- Мышечные волокна приспосабливаются к определенному типу физических нагрузок, возникающих во время тренировки.
 Механический стресс относится к физическим нагрузкам, возникающим во время тренировки с отягощениями, которые вызывают микротравмы мышечных волокон. Поскольку волокна повреждаются в результате упражнений, они сигнализируют о биохимической реакции по производству новых клеток-сателлитов, ответственных за восстановление механической структуры мышечной клетки и создание новых мышечных белков.
Механический стресс относится к физическим нагрузкам, возникающим во время тренировки с отягощениями, которые вызывают микротравмы мышечных волокон. Поскольку волокна повреждаются в результате упражнений, они сигнализируют о биохимической реакции по производству новых клеток-сателлитов, ответственных за восстановление механической структуры мышечной клетки и создание новых мышечных белков. - Мышечные волокна активируются двигательным нейроном, который является связующим звеном между центральной нервной системой и конкретной мышцей, необходимой для выполнения определенной деятельности. Мышечно-двигательная единица — это двигательный нейрон и прикрепленные к нему мышечные волокна. Думайте о двигательной единице как о выключателе света для мышцы — поскольку мышца необходима для создания силы, двигательные единицы «загораются», чтобы стимулировать волокна к сокращению, чтобы произвести эту силу. Существует ряд различных типов мышечных единиц, которые обычно делятся на три общие категории: тип I, тип IIA и тип IIB .
- Согласно теории «все или ничего» моторная единица либо активна, либо неактивна.Когда ему дают сигнал к сокращению, он активирует все прикрепленные к нему мышечные волокна. Медленно сокращающиеся двигательные единицы имеют низкий порог активации и низкие скорости проводимости и прикреплены к мышечным волокнам I типа. Быстро сокращающиеся двигательные единицы имеют более высокий порог активации, прикреплены к мышечным волокнам типа II и способны передавать сигналы с более высокими скоростями, что приводит к увеличению мышечной силы.
- Волокна типа I известны как аэробные волокна. Эти волокна имеют более высокую плотность митохондрий, которые эффективны в аэробном метаболизме (процессе создания энергии для подпитки мышечной активности кислородом).Митохондрии придают клетке более темный цвет, поэтому они известны как красные мышечные волокна. Волокна типа I используют кислород для создания энергии для низкоинтенсивных, долгосрочных и ориентированных на выносливость действий, таких как ходьба, бег, плавание, езда на велосипеде или стояние в течение длительных периодов времени.
- Волокна типа IIB известны как анаэробные мышечные волокна. Волокна типа IIB накапливают энергию, которая выделяется при коротких, взрывных и чрезвычайно интенсивных занятиях. Волокна типа IIB не имеют митохондрий и имеют бесцветный вид, поэтому они известны как белые волокна.Поскольку у них ограниченный запас накопленной энергии и, следовательно, они быстро утомляются, волокна типа IIB используются для силовых и силовых упражнений, требующих большого количества силы за короткий период времени.
- Волокна типа IIA имеют митохондрии, поэтому они могут участвовать в аэробной деятельности; однако их также можно использовать для быстрого создания силы во время действий, требующих большого количества силы или мощности. Быстро сокращающиеся мышечные волокна также имеют больший диаметр, чем волокна типа I, и играют более значительную роль в гипертрофии (технический термин, обозначающий рост мышц).Вовлечение и иннервация мышечных волокон типа II требует создания достаточной механической перегрузки, чтобы утомить задействованную мышцу к концу подхода
- Мышечные волокна работают по принципу размера. Поскольку мышце требуется сила, она начнется с активации меньших двигательных единиц I типа. Когда эти двигательные единицы типа I не могут обеспечить необходимую силу (или утомление), для выполнения работы задействуются более крупные двигательные единицы типа II и мышечные волокна. Мышца имеет конечное количество двигательных единиц, и двигательные единицы типа II с более высоким порогом не «включаются», если не требуется высокий уровень силы.Наиболее распространенный способ увеличить активацию двигательных единиц — это поднимать более тяжелые веса, потому что повышенная нагрузка на мышцу заставляет большее количество двигательных единиц активировать больше волокон для создания силы, необходимой для преодоления сопротивления. Вот почему ваши мышцы дрожат, когда вы впервые пытаетесь поднять тяжелый вес — мышечные двигательные единицы, которые ранее не использовались, «просыпаются» и приводятся в действие.
- Мышечные волокна испытывают два определенных типа гипертрофии. Миофибриллярная гипертрофия относится к увеличению размера или толщины отдельных актиновых и миозиновых белковых нитей, что может улучшить способность отдельных волокон создавать силу. Миофибриллярная гипертрофия не приводит к увеличению мышц; скорее, это приводит к образованию более толстых мышечных волокон, способных генерировать больше силы. Саркоплазматическая гипертрофия — это увеличение объема полужидкого межфибриллярного вещества, окружающего отдельное мышечное волокно. Эта жидкость содержит белки, которые способствуют восстановлению и росту тканей.Мышечный «насос», над которым работают бодибилдеры, на самом деле является саркоплазматической гипертрофией — поперечное сечение мышечных волокон будет увеличиваться, но большая часть увеличенного размера мышц происходит из-за увеличенного объема саркоплазмы и несокращающихся белков, не участвующих напрямую. производство силы.
- Одним из долгосрочных способов адаптации мышц к тренировкам с отягощениями является увеличение поперечной ширины мышечных волокон. По мере увеличения площади поперечного сечения волокна имеют большее поверхностное натяжение и становятся способными создавать большее количество силы. Мышцы с большей площадью поперечного сечения отдельных мышечных волокон способны создавать большее количество силы. Помимо того, что они отвечают за создание силы, необходимой для динамических движений, мышечные волокна типа II имеют больший диаметр (поперечную ширину), чем волокна типа I, и отвечают за гипертрофию или увеличение размера конкретной мышцы.
- Мышечные волокна типа IIA и IIB отвечают за движение, а также за размер и определение мышц.Обе классификации мышечных волокон типа II создают более высокие уровни силы для движения человека и известны как фазовые мышцы. Волокна типа I отвечают за поддержание осанки и стабильности суставов и могут быть отнесены к тонизирующим мышцам.
 Механический стресс относится к физическим нагрузкам, возникающим во время тренировки с отягощениями, которые вызывают микротравмы мышечных волокон. Поскольку волокна повреждаются в результате упражнений, они сигнализируют о биохимической реакции по производству новых клеток-сателлитов, ответственных за восстановление механической структуры мышечной клетки и создание новых мышечных белков.
Механический стресс относится к физическим нагрузкам, возникающим во время тренировки с отягощениями, которые вызывают микротравмы мышечных волокон. Поскольку волокна повреждаются в результате упражнений, они сигнализируют о биохимической реакции по производству новых клеток-сателлитов, ответственных за восстановление механической структуры мышечной клетки и создание новых мышечных белков.

 Поскольку мышце требуется сила, она начнется с активации меньших двигательных единиц I типа. Когда эти двигательные единицы типа I не могут обеспечить необходимую силу (или утомление), для выполнения работы задействуются более крупные двигательные единицы типа II и мышечные волокна. Мышца имеет конечное количество двигательных единиц, и двигательные единицы типа II с более высоким порогом не «включаются», если не требуется высокий уровень силы.Наиболее распространенный способ увеличить активацию двигательных единиц — это поднимать более тяжелые веса, потому что повышенная нагрузка на мышцу заставляет большее количество двигательных единиц активировать больше волокон для создания силы, необходимой для преодоления сопротивления. Вот почему ваши мышцы дрожат, когда вы впервые пытаетесь поднять тяжелый вес — мышечные двигательные единицы, которые ранее не использовались, «просыпаются» и приводятся в действие.
Поскольку мышце требуется сила, она начнется с активации меньших двигательных единиц I типа. Когда эти двигательные единицы типа I не могут обеспечить необходимую силу (или утомление), для выполнения работы задействуются более крупные двигательные единицы типа II и мышечные волокна. Мышца имеет конечное количество двигательных единиц, и двигательные единицы типа II с более высоким порогом не «включаются», если не требуется высокий уровень силы.Наиболее распространенный способ увеличить активацию двигательных единиц — это поднимать более тяжелые веса, потому что повышенная нагрузка на мышцу заставляет большее количество двигательных единиц активировать больше волокон для создания силы, необходимой для преодоления сопротивления. Вот почему ваши мышцы дрожат, когда вы впервые пытаетесь поднять тяжелый вес — мышечные двигательные единицы, которые ранее не использовались, «просыпаются» и приводятся в действие. Миофибриллярная гипертрофия не приводит к увеличению мышц; скорее, это приводит к образованию более толстых мышечных волокон, способных генерировать больше силы. Саркоплазматическая гипертрофия — это увеличение объема полужидкого межфибриллярного вещества, окружающего отдельное мышечное волокно. Эта жидкость содержит белки, которые способствуют восстановлению и росту тканей.Мышечный «насос», над которым работают бодибилдеры, на самом деле является саркоплазматической гипертрофией — поперечное сечение мышечных волокон будет увеличиваться, но большая часть увеличенного размера мышц происходит из-за увеличенного объема саркоплазмы и несокращающихся белков, не участвующих напрямую. производство силы.
Миофибриллярная гипертрофия не приводит к увеличению мышц; скорее, это приводит к образованию более толстых мышечных волокон, способных генерировать больше силы. Саркоплазматическая гипертрофия — это увеличение объема полужидкого межфибриллярного вещества, окружающего отдельное мышечное волокно. Эта жидкость содержит белки, которые способствуют восстановлению и росту тканей.Мышечный «насос», над которым работают бодибилдеры, на самом деле является саркоплазматической гипертрофией — поперечное сечение мышечных волокон будет увеличиваться, но большая часть увеличенного размера мышц происходит из-за увеличенного объема саркоплазмы и несокращающихся белков, не участвующих напрямую. производство силы. Мышцы с большей площадью поперечного сечения отдельных мышечных волокон способны создавать большее количество силы. Помимо того, что они отвечают за создание силы, необходимой для динамических движений, мышечные волокна типа II имеют больший диаметр (поперечную ширину), чем волокна типа I, и отвечают за гипертрофию или увеличение размера конкретной мышцы.
Мышцы с большей площадью поперечного сечения отдельных мышечных волокон способны создавать большее количество силы. Помимо того, что они отвечают за создание силы, необходимой для динамических движений, мышечные волокна типа II имеют больший диаметр (поперечную ширину), чем волокна типа I, и отвечают за гипертрофию или увеличение размера конкретной мышцы. Если у вашего клиента есть фитнес-цель, связанная с улучшением мышечной массы или наращиванием силы, единственный способ достичь этого — активировать двигательные единицы типа II и мышечные волокна. Существует распространенное заблуждение, что развитие мышечного тонуса («повышение тонуса») лучше всего достигается за счет использования более легких весов для более частых повторений.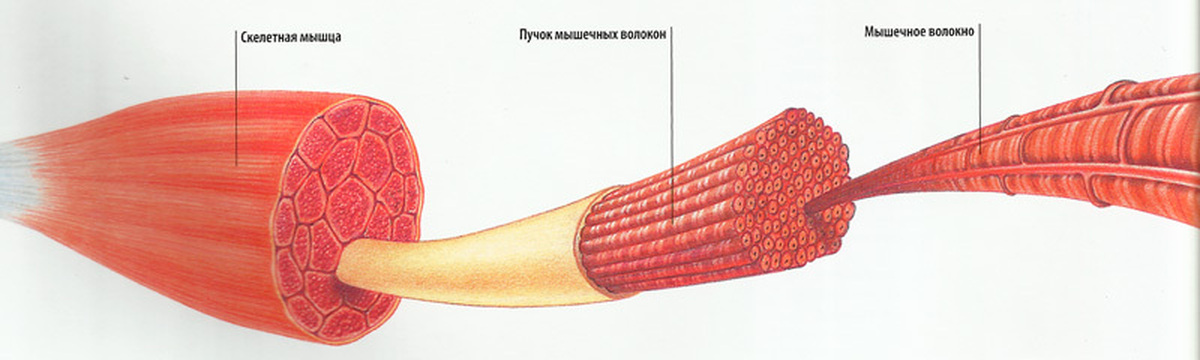 Хотя можно добиться четкости с более легкими весами, для этого требуется много повторений. Использование легких весов для большого количества повторений может улучшить аэробную эффективность волокон типа I, но если упражнение не выполняется до утомления, которое возникает только после набора волокон типа II, оно не приведет к достижению желаемой цели определения. Однако, если время является проблемой (как для многих людей), использование более тяжелого веса для меньшего количества повторений является эффективным методом стимулирования роста и определения волокон типа II.Важно отметить, что упражнения с более высоким диапазоном повторений приведут к росту мышц из-за саркоплазматической гипертрофии, в то время как использование более тяжелых нагрузок для меньшего количества повторений вызывает гипертрофию миофибрилл, которая увеличивает силу, но не обязательно приводит к значительному увеличению мышечного роста.
Хотя можно добиться четкости с более легкими весами, для этого требуется много повторений. Использование легких весов для большого количества повторений может улучшить аэробную эффективность волокон типа I, но если упражнение не выполняется до утомления, которое возникает только после набора волокон типа II, оно не приведет к достижению желаемой цели определения. Однако, если время является проблемой (как для многих людей), использование более тяжелого веса для меньшего количества повторений является эффективным методом стимулирования роста и определения волокон типа II.Важно отметить, что упражнения с более высоким диапазоном повторений приведут к росту мышц из-за саркоплазматической гипертрофии, в то время как использование более тяжелых нагрузок для меньшего количества повторений вызывает гипертрофию миофибрилл, которая увеличивает силу, но не обязательно приводит к значительному увеличению мышечного роста.
Получение человеческих мышечных волокон и сателлитных клеток из плюрипотентных стволовых клеток человека in vitro
Chal, J. и другие. Дифференциация плюрипотентных стволовых клеток в мышечные волокна для моделирования мышечной дистрофии Дюшенна. Nat. Biotechnol. 33 , 962–969 (2015).
и другие. Дифференциация плюрипотентных стволовых клеток в мышечные волокна для моделирования мышечной дистрофии Дюшенна. Nat. Biotechnol. 33 , 962–969 (2015).
CAS PubMed Google ученый
Rohwedel, J. et al. Дифференцировка мышечных клеток эмбриональных стволовых клеток отражает миогенез in vivo : онтогенетически регулируемая экспрессия генов миогенной детерминации и функциональная экспрессия ионных токов. Dev. Биол. 164 , 87–101 (1994).
CAS PubMed Google ученый
Вобус, А. Возможности эмбриональных стволовых клеток. Мол. Аспекты Мед. 22 , 149–164 (2001).
CAS PubMed Google ученый
Greco, T.L. и другие. Анализ рудиментарной мутации хвоста показывает, что дозировка гена Wnt-3a регулирует осевое развитие мышей. Genes Dev.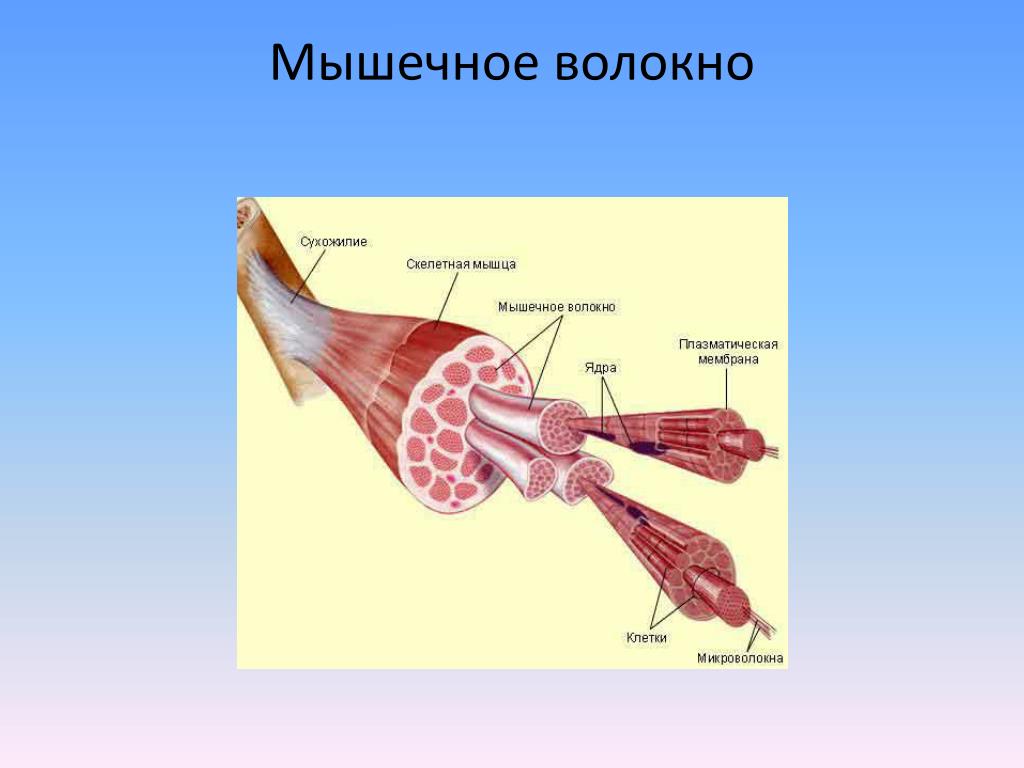 10 , 313–324 (1996).
10 , 313–324 (1996).
CAS PubMed Google ученый
Aulehla, A. et al. Градиент бета-катенина связывает системы часов и волнового фронта при сегментации эмбрионов мыши. Nat. Cell Biol. 10 , 186–193 (2008).
CAS PubMed Google ученый
Данти, W.C. Jr. et al. Передача сигналов Wnt3a / beta-catenin контролирует развитие задней части тела путем координации формирования и сегментации мезодермы. Разработка 135 , 85–94 (2008).
CAS PubMed Google ученый
Yamaguchi, T.P., Takada, S., Yoshikawa, Y., Wu, N. & McMahon, A.P. T (Brachyury) является прямой мишенью Wnt3a во время спецификации параксиальной мезодермы. Genes Dev. 13 , 3185–3190 (1999).
CAS PubMed PubMed Central Google ученый
Чал, Дж. И Pourquie, О. Формирование рисунка и дифференциация позвоночника. Cold Spring Harb. Монография 41–116 http://dx.doi.org/10.1101/087969825.53.41 (2009).
И Pourquie, О. Формирование рисунка и дифференциация позвоночника. Cold Spring Harb. Монография 41–116 http://dx.doi.org/10.1101/087969825.53.41 (2009).
Yoshikawa, Y., Fujimori, T., McMahon, A.P. & Takada, S. Доказательства того, что отсутствие передачи сигналов Wnt-3a способствует нейрализации вместо развития параксиальной мезодермы у мышей. Dev. Биол. 183 , 234–242 (1997).
CAS PubMed Google ученый
Чепмен, Д.Л., Агульник, И., Хэнкок, С., Сильвер, Л.М., Папайоанну, В. Tbx6, ген T-Box мыши, участвующий в формировании параксиальной мезодермы при гаструляции. Dev. Биол. 180 , 534–542 (1996).
CAS PubMed Google ученый
Galceran, J., Farinas, I., Depew, MJ, Clevers, H. & Grosschedl, R. Wnt3a — / — подобный фенотип и дефицит конечностей у Lef1 (- / -) Tcf1 (- / — ) мышей. Genes Dev. 13 , 709–717 (1999).
13 , 709–717 (1999).
CAS PubMed PubMed Central Google ученый
Линдсли, Р.С., Гилл, Дж. Г., Киба, М., Мерфи, Т.Л. И Мерфи, К. Каноническая передача сигналов Wnt необходима для развития мезодермы, происходящей из эмбриональных стволовых клеток. Разработка 133 , 3787–3796 (2006).
CAS PubMed Google ученый
Гадуэ, П., Хубер, Т.Л., Паддисон, П.Дж. и Келлер, Г.М. Передача сигналов Wnt и TGF-бета необходима для индукции модели in vitro образования примитивной полоски с использованием эмбриональных стволовых клеток. Proc. Natl. Акад. Sci. США 103 , 16806–16811 (2006).
CAS PubMed Google ученый
Sumi, T., Tsuneyoshi, N., Nakatsuji, N. & Suemori, H. Определение спецификации ранних клонов человеческих эмбриональных стволовых клеток с помощью согласованного баланса канонического Wnt / бета-катенина, Activin / Nodal и BMP сигнализация. Разработка 135 , 2969–2979 (2008).
Разработка 135 , 2969–2979 (2008).
CAS PubMed Google ученый
Nakanishi, M. et al. Направленная индукция передней и задней примитивной полоски с помощью Wnt из эмбриональных стволовых клеток, культивируемых в бессывороточной среде определенного химического состава. FASEB J. 23 , 114–122 (2009).
CAS PubMed Google ученый
Сюй, К.и другие. Система культивирования эмбрионов рыбок данио определяет факторы, которые способствуют миогенезу позвоночных у разных видов. Cell 155 , 909–921 (2013).
CAS PubMed PubMed Central Google ученый
Борчин, Б., Чен, Дж. И Барбери, Т. Получение и FACS-опосредованная очистка предшественников скелетных мышц PAX3 + / PAX7 + из плюрипотентных стволовых клеток человека. Stem Cell Rep. 1 , 620–631 (2013).
1 , 620–631 (2013).
CAS Google ученый
Shelton, M. et al. Получение и распространение PAX7-позитивных мышечных предшественников из эмбриональных стволовых клеток человека и мыши. Stem Cell Rep. 3 , 516–529 (2014).
CAS Google ученый
Hwang, Y. et al. WNT3A способствует миогенезу эмбриональных стволовых клеток человека и усиливает приживление in vivo и . Sci. Отчетность 4 , 5916 (2014).
CAS PubMed PubMed Central Google ученый
Aulehla, A. et al. Wnt3a играет главную роль в часах сегментации, контролирующих сомитогенез. Dev. Ячейка 4 , 395–406 (2003).
CAS PubMed Google ученый
Гита-Логанатан, П., Ниммагадда, С., Скааль, М. , Хуанг Р. и Крист Б. Передача сигналов Wnt в развитии сомитов. Ann. Анат. 190 , 208–222 (2008).
, Хуанг Р. и Крист Б. Передача сигналов Wnt в развитии сомитов. Ann. Анат. 190 , 208–222 (2008).
CAS PubMed Google ученый
Tonegawa, A., Funayama, N., Ueno, N. & Takahashi, Y. Подразделение мезодермы вдоль медиолатеральной оси у цыплят, контролируемое различными концентрациями BMP-4. Разработка 124 , 1975–1984 (1997).
CAS PubMed Google ученый
Хирсингер, Э.и другие. Noggin действует ниже Wnt и Sonic Hedgehog, чтобы противодействовать BMP4 в формировании паттерна птичьего сомита. Разработка 124 , 4605–4614 (1997).
CAS PubMed Google ученый
McMahon, J.A. и другие. Опосредованный Noggin антагонизм передачи сигналов BMP необходим для роста и формирования паттерна нервной трубки и сомита. Genes Dev. 12 , 1438–1452 (1998).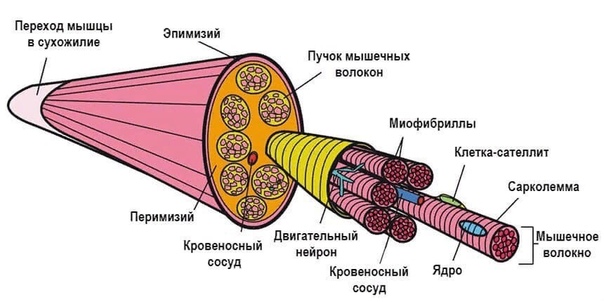
CAS PubMed PubMed Central Google ученый
Решеф Р., Марото, М., Лассар, А. Регуляция судьбы дорсальных сомитных клеток: BMP и Noggin контролируют время и характер экспрессии миогенного регулятора. Genes Dev. 12 , 290–303 (1998).
CAS PubMed PubMed Central Google ученый
Streit, A. & Stern, C.D. Формирование паттерна мезодермы и формирование сомитов во время регрессии узла: дифференциальные эффекты хордина и ноггина. мех. Dev. 85 , 85–96 (1999).
CAS PubMed Google ученый
Miura, S., Davis, S., Klingensmith, J. & Mishina, Y. Передача сигналов BMP в эпибласте необходима для правильного рекрутирования предполагаемой параксиальной мезодермы и развития сомитов. Разработка 133 , 3767–3775 (2006).
CAS PubMed Google ученый
Села-Доненфельд, Д.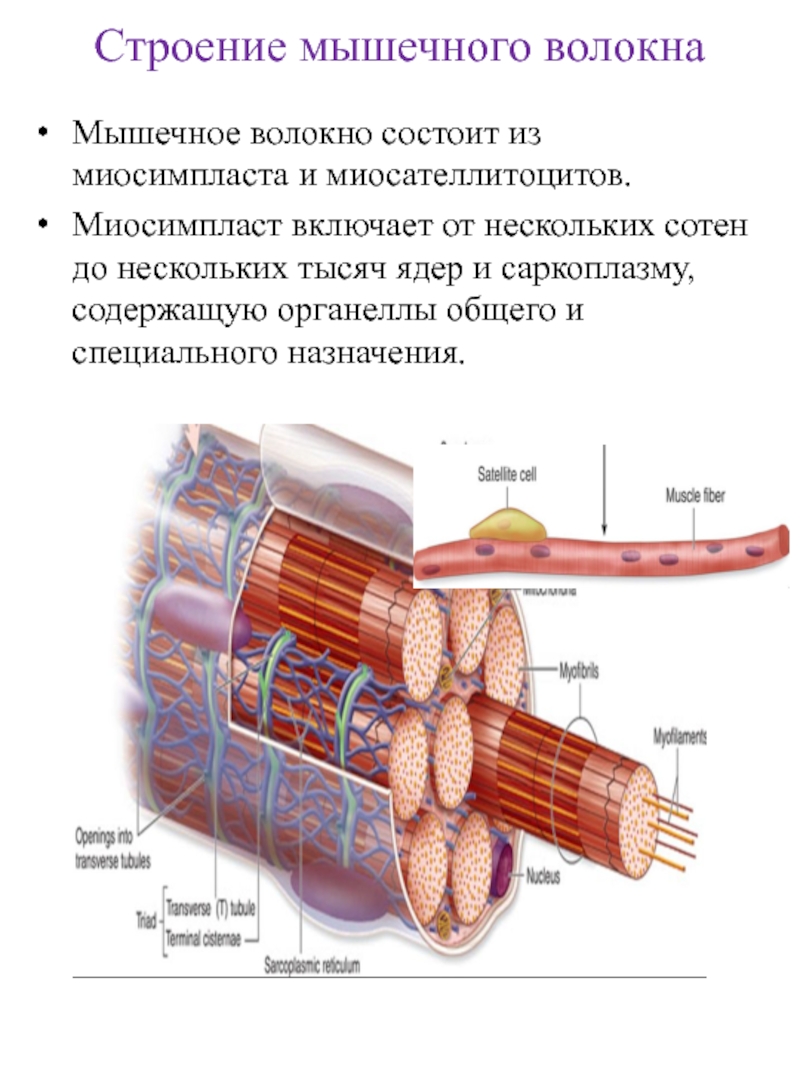 И Kalcheim, C. Локализованные взаимодействия BMP4-noggin генерируют динамическое формирование паттерна экспрессии noggin в сомитах. Dev. Биол. 246 , 311–328 (2002).
И Kalcheim, C. Локализованные взаимодействия BMP4-noggin генерируют динамическое формирование паттерна экспрессии noggin в сомитах. Dev. Биол. 246 , 311–328 (2002).
CAS PubMed Google ученый
Morizane, R. et al. Органоиды нефрона, полученные из плюрипотентных стволовых клеток человека, моделируют развитие и повреждение почек. Nat. Biotechnol. 33 , 1193–1200 (2015).
CAS PubMed PubMed Central Google ученый
Умеда, К.и другие. Хондрогенная параксиальная мезодерма человека, направленная спецификация и предполагаемое выделение из плюрипотентных стволовых клеток. Sci. Отчет 2 , 455 (2012).
PubMed PubMed Central Google ученый
Adelman, C.A., Chattopadhyay, S. & Bieker, J.J. Путь BMP / BMPR / Smad управляет экспрессией эритроид-специфичных факторов транскрипции EKLF и GATA1 во время дифференцировки эмбриоидных тел в бессывороточной среде. Разработка 129 , 539–549 (2002).
Разработка 129 , 539–549 (2002).
CAS PubMed Google ученый
Ностро, M.C., Cheng, X., Keller, G.M. & Gadue, P. Передача сигналов Wnt, активина и BMP регулирует различные стадии пути развития от эмбриональных стволовых клеток до крови. Cell Stem Cell 2 , 60–71 (2008).
CAS PubMed PubMed Central Google ученый
Ленгерке, К.и другие. BMP и Wnt определяют гематопоэтическую судьбу путем активации пути Cdx-Hox. Cell Stem Cell 2 , 72–82 (2008).
CAS PubMed Google ученый
Орлова В.В., Чува де Соуза Лопес, С. и Валдимарсдоттир, Г. Передача сигналов BMP-SMAD: от плюрипотентных стволовых клеток к сердечно-сосудистой функции. Cytokine Growth Factor Rev. 27 , 55–63 (2016).
CAS PubMed Google ученый
Виварелли, Э. И Коссу, Г. Нейронный контроль ранней миогенной дифференцировки в культурах сомитов мышей. Dev. Биол. 117 , 319–325 (1986).
И Коссу, Г. Нейронный контроль ранней миогенной дифференцировки в культурах сомитов мышей. Dev. Биол. 117 , 319–325 (1986).
CAS PubMed Google ученый
Cossu, G., Kelly, R., Di Donna, S., Vivarelli, E. & Buckingham, M. Дифференциация миобластов во время сомитогенеза у млекопитающих зависит от эффекта сообщества. Proc. Natl. Акад. Sci. USA 92 , 2254–2258 (1995).
CAS PubMed Google ученый
Buffinger, N. & Stockdale, F.E. Миогенная спецификация сомитов опосредована диффузными факторами. Dev. Биол. 169 , 96–108 (1995).
CAS PubMed Google ученый
Rong, P.M., Teillet, M.A., Ziller, C. & Le Douarin, N.M. Комплекс нервной трубки / хорды необходим для дифференцировки поперечно-полосатых мышц позвоночника, но не конечностей и стенки тела. Разработка 115 , 657–672 (1992).
Разработка 115 , 657–672 (1992).
CAS PubMed Google ученый
Миллер, Дж. Б., Эверитт, Э. А., Смит, Т. Х., Блок, Н. Э. И Доминов, Я. Клеточное и молекулярное разнообразие в развитии скелетных мышц: новости из in vitro и. Bioessays 15 , 191–196 (1993).
CAS PubMed Google ученый
Невилл, К., Розенталь, Н., МакГрю, М., Богданова, Н. и Хаушка, С. Культуры скелетных мышц. Methods Cell Biol. 52 , 85–116 (1997).
CAS PubMed Google ученый
Дановиз, М.Е. и Яблонка-Реувени, З. Сателлитные клетки скелетных мышц: предпосылки и методы выделения и анализа в системе первичного культивирования. Methods Mol. Биол. 798 , 21–52 (2012).
CAS PubMed PubMed Central Google ученый
Миллер, Дж. Б. и Стокдейл, Ф.Э. Истоки развития волокон скелетных мышц: клональный анализ миогенных клеточных линий, основанный на экспрессии быстрых и медленных тяжелых цепей миозина. Proc. Natl. Акад. Sci. USA 83 , 3860–3864 (1986).
Б. и Стокдейл, Ф.Э. Истоки развития волокон скелетных мышц: клональный анализ миогенных клеточных линий, основанный на экспрессии быстрых и медленных тяжелых цепей миозина. Proc. Natl. Акад. Sci. USA 83 , 3860–3864 (1986).
CAS PubMed Google ученый
Rutz, R. & Hauschka, S. Клональный анализ миогенеза позвоночных. VII. Наследование типа мышечной колонии посредством последовательных субклональных пассажей in vitro . Dev. Биол. 91 , 103–110 (1982).
CAS PubMed Google ученый
Lian, X. et al. Эффективная дифференцировка плюрипотентных стволовых клеток человека в эндотелиальные предшественники посредством низкомолекулярной активации передачи сигналов WNT. Stem Cell Rep. 3 , 804–816 (2014).
CAS Google ученый
Tesar, P.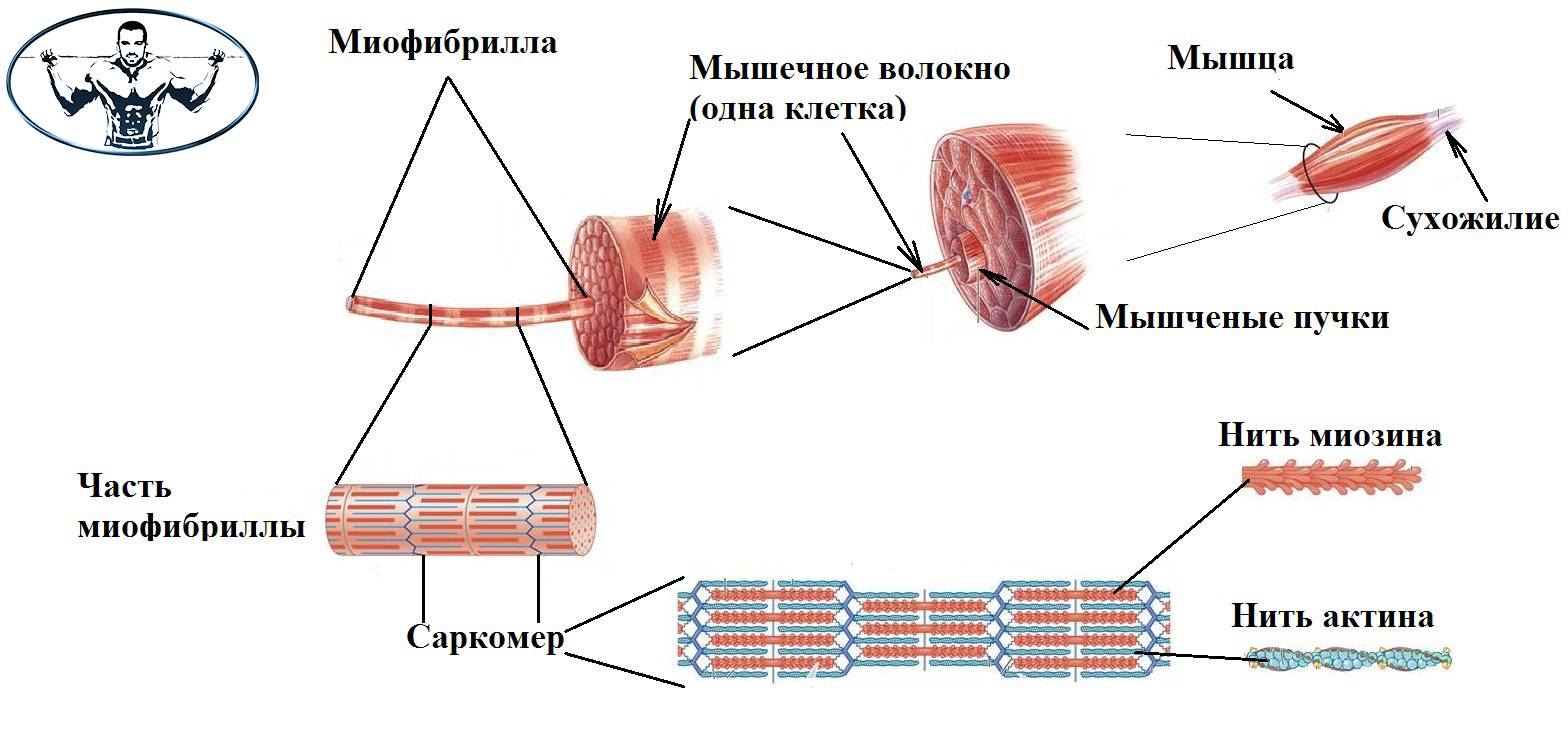 J. et al. Новые клеточные линии эпибласта мыши имеют общие черты с эмбриональными стволовыми клетками человека. Природа 448 , 196–199 (2007).
J. et al. Новые клеточные линии эпибласта мыши имеют общие черты с эмбриональными стволовыми клетками человека. Природа 448 , 196–199 (2007).
CAS PubMed PubMed Central Google ученый
Шелтон, М., Кочарян, А., Лю, Дж., Скерянц, И.С. И Стэнфорд, W.L. Устойчивое создание и распространение предшественников скелетных мышц и миоцитов из плюрипотентных стволовых клеток человека. Методы 101 , 73–84 (2015).
PubMed Google ученый
Махмуд, А., Харкнесс, Л., Шредер, Х.Д., Абдаллах, Б.М. & Kassem, M. Усиленная дифференцировка человеческих эмбриональных стволовых клеток в мезенхимальные предшественники путем ингибирования передачи сигналов TGF-бета / активин / узел с использованием SB-431542. J. Bone Miner. Res. 25 , 1216–1233 (2010).
CAS PubMed Google ученый
Хосояма, Т. , МакГиверн, Дж. В., Ван Дайк, Дж. М., Эберт, А. Д. и Сузуки, М. Получение миогенных предшественников непосредственно из плюрипотентных стволовых клеток человека с использованием сферической культуры. Stem Cells Пер. Med. 3 , 564–574 (2014).
, МакГиверн, Дж. В., Ван Дайк, Дж. М., Эберт, А. Д. и Сузуки, М. Получение миогенных предшественников непосредственно из плюрипотентных стволовых клеток человека с использованием сферической культуры. Stem Cells Пер. Med. 3 , 564–574 (2014).
CAS PubMed PubMed Central Google ученый
Maffioletti, S.M. и другие. Эффективное получение и индуцируемая дифференцировка расширяемых скелетных миогенных клеток из человеческих ES и индивидуальных iPS-клеток. Nat. Protoc. 10 , 941–958 (2015).
CAS PubMed Google ученый
Shoji, E., Woltjen, K. & Sakurai, H. Управляемая миогенная дифференцировка индуцированных человеком плюрипотентных стволовых клеток. Methods Mol. Биол. 1353 , 89–99 (2016).
CAS PubMed Google ученый
Альбини, С. и Пури, П.L. Генерация миосфер из чЭСК путем эпигенетического репрограммирования. J. Vis. Exp. 88 , e51243 (2014).
и Пури, П.L. Генерация миосфер из чЭСК путем эпигенетического репрограммирования. J. Vis. Exp. 88 , e51243 (2014).
Google ученый
Герли М.Ф., Маффиолетти С.М., Миллет, К. и Тедеско, Ф.С. Трансплантация индуцированных плюрипотентных стволовых клеток мезоангиобластоподобных миогенных предшественников на мышиных моделях мышечной регенерации. J. Vis. Exp. 83 , e50532 (2014).
Google ученый
Дараби, Р.И Perlingeiro, R.C. Получение скелетных миогенных предшественников из плюрипотентных стволовых клеток человека с использованием условной экспрессии PAX7. Methods Mol. Биол. 1357 , 423–439 (2016).
CAS PubMed Google ученый
Ставропулос М.Е., Менгарелли И. и Барбери Т. Дифференциация мультипотентных мезенхимальных предшественников и скелетных миобластов от эмбриональных стволовых клеток человека.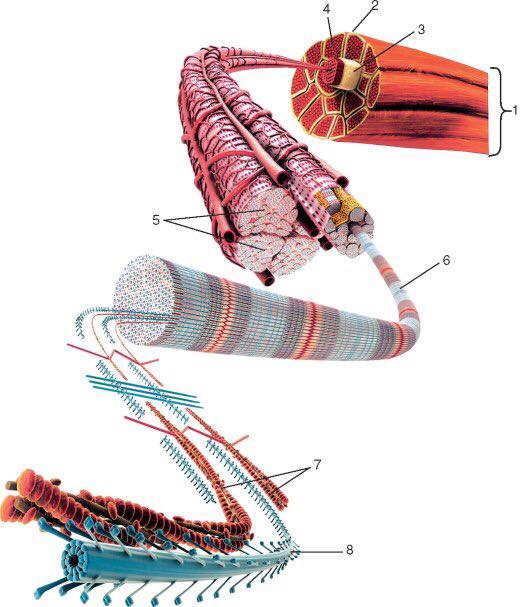 Curr.Protoc. Stem Cell Biol. Глава 1 , Раздел 1F 8 (2009).
Curr.Protoc. Stem Cell Biol. Глава 1 , Раздел 1F 8 (2009).
PubMed Google ученый
Filareto, A. et al. Подход генной терапии ex vivo для лечения мышечной дистрофии с использованием индуцибельных плюрипотентных стволовых клеток. Nat. Commun. 4 , 1549 (2013).
PubMed PubMed Central Google ученый
Кватрочелли, М.и другие. Внутренняя память клетки усиливает миогенную приверженность ИПСК, происходящих из перицитов. J. Pathol. 223 , 593–603 (2011).
CAS PubMed Google ученый
Barberi, T. et al. Получение приживляемых скелетных миобластов из эмбриональных стволовых клеток человека. Nat. Med. 13 , 642–648 (2007).
CAS PubMed Google ученый
Beers, J. и другие. Пассирование и размножение колоний плюрипотентных стволовых клеток человека путем бесферментной диссоциации в химически определенных условиях культивирования. Nat. Protoc. 7 , 2029–2040 (2012).
и другие. Пассирование и размножение колоний плюрипотентных стволовых клеток человека путем бесферментной диссоциации в химически определенных условиях культивирования. Nat. Protoc. 7 , 2029–2040 (2012).
CAS PubMed PubMed Central Google ученый
Baharvand, H., Salekdeh, G.H., Taei, A. & Mollamohammadi, S. Эффективный и простой в использовании протокол криоконсервации ES- и iPS-клеток человека. Nat. Protoc. 5 , 588–594 (2010).
CAS PubMed Google ученый
Marti, M. et al. Характеристика плюрипотентных стволовых клеток. Nat. Protoc. 8 , 223–253 (2013).
CAS PubMed Google ученый
Schwartz, P.H., Brick, D.J., Nethercott, H.E. И Стовер А.Е. Традиционная культура эмбриональных стволовых клеток человека. Methods Mol. Биол. 767 , 107–123 (2011).
Биол. 767 , 107–123 (2011).
CAS PubMed PubMed Central Google ученый
Лин С. и Талбот П. Методы культивирования эмбриональных стволовых клеток мыши и человека. Methods Mol. Биол. 690 , 31–56 (2011).
CAS PubMed Google ученый
van den Brink, S.C. et al. Нарушение симметрии, спецификация зародышевого листка и осевая организация в агрегатах эмбриональных стволовых клеток мыши. Разработка 141 , 4231–4242 (2014).
CAS PubMed PubMed Central Google ученый
Gouti, M. et al. In vitro поколение нейромезодермальных предшественников обнаруживает различные роли передачи сигналов wnt в спецификации идентичности спинного мозга и параксиальной мезодермы. PLoS Biol. 12 , e1001937 (2014).
PubMed PubMed Central Google ученый
Rohwedel, J. , Гуан К., Хегерт К. и Вобус А.М. Эмбриональные стволовые клетки как модель in vitro для исследований мутагенности, цитотоксичности и эмбриотоксичности: современное состояние и перспективы на будущее. Токсикология In vitro 15 , 741–753 (2001).
, Гуан К., Хегерт К. и Вобус А.М. Эмбриональные стволовые клетки как модель in vitro для исследований мутагенности, цитотоксичности и эмбриотоксичности: современное состояние и перспективы на будущее. Токсикология In vitro 15 , 741–753 (2001).
CAS PubMed Google ученый
Giobbe, G.G. и другие. Функциональная дифференциация плюрипотентных стволовых клеток человека на чипе. Nat. Методы 12 , 637–640 (2015).
CAS PubMed Google ученый
Park, D., Lim, J., Park, J.Y. И Ли, С. Краткий обзор: микросреда стволовых клеток на чипе: современные технологии тканевой инженерии и биологии стволовых клеток. Stem Cells Пер. Med. 4 , 1352–1368 (2015).
CAS PubMed PubMed Central Google ученый
Островидов, С.и другие. Инженерия ткани скелетных мышц: методы формирования скелетных мышечных трубок и их применение. Tissue Eng. Часть B Ред. 20 , 403–436 (2014).
Tissue Eng. Часть B Ред. 20 , 403–436 (2014).
PubMed PubMed Central Google ученый
Cheng, C.W., Solorio, L.D. И Альсберг, Э. Децеллюляризованные ткани и внеклеточные матриксы клеточного происхождения в качестве каркасов для ортопедической тканевой инженерии. Biotechnol. Adv. 32 , 462–484 (2014).
CAS PubMed PubMed Central Google ученый
Деннис, Р.Г. И Косник, П. II. Возбудимость и изометрические сократительные свойства конструкций скелетных мышц млекопитающих, сконструированных in vitro . Клетка in vitro. Dev. Биол. Anim. 36 , 327–335 (2000).
CAS PubMed Google ученый
Bian, W.И Bursac, Н. Сконструированные сети ткани скелетных мышц с управляемой архитектурой. Биоматериалы 30 , 1401–1412 (2009).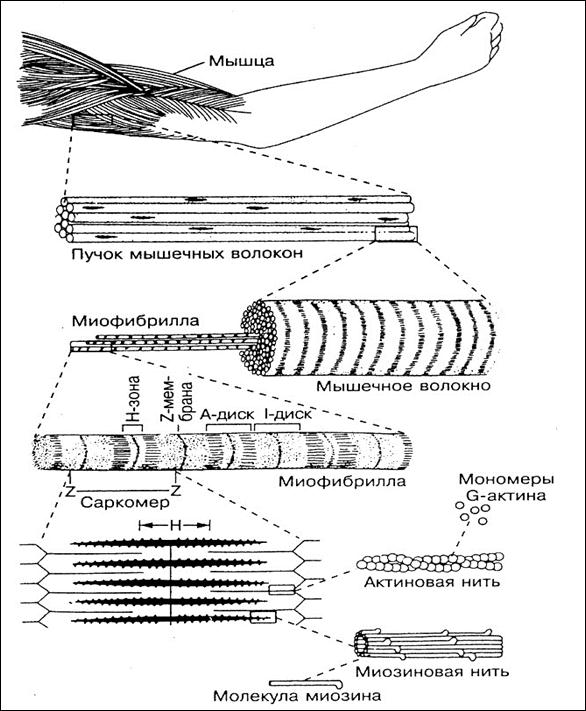
CAS PubMed Google ученый
Юхас М. и Бурзак Н. Инженерное восстановление скелетных мышц. Curr. Opin. Biotechnol. 24 , 880–886 (2013).
CAS PubMed PubMed Central Google ученый
Сакар, М.S. et al. Формирование и оптогенетический контроль инженерных 3D-биоактуаторов скелетных мышц. Лабораторный чип 12 , 4976–4985 (2012).
CAS PubMed PubMed Central Google ученый
Neal, D., Sakar, M.S., Ong, L.L., Harry Asada, H. Формирование вытянутых трехмерных тканей в виде пучков, состоящих из высокоплотных выровненных клеток, с использованием жертвенного внешнего формования. Лабораторный чип 14 , 1907–1916 (2014).
CAS PubMed Google ученый
Grosberg, A.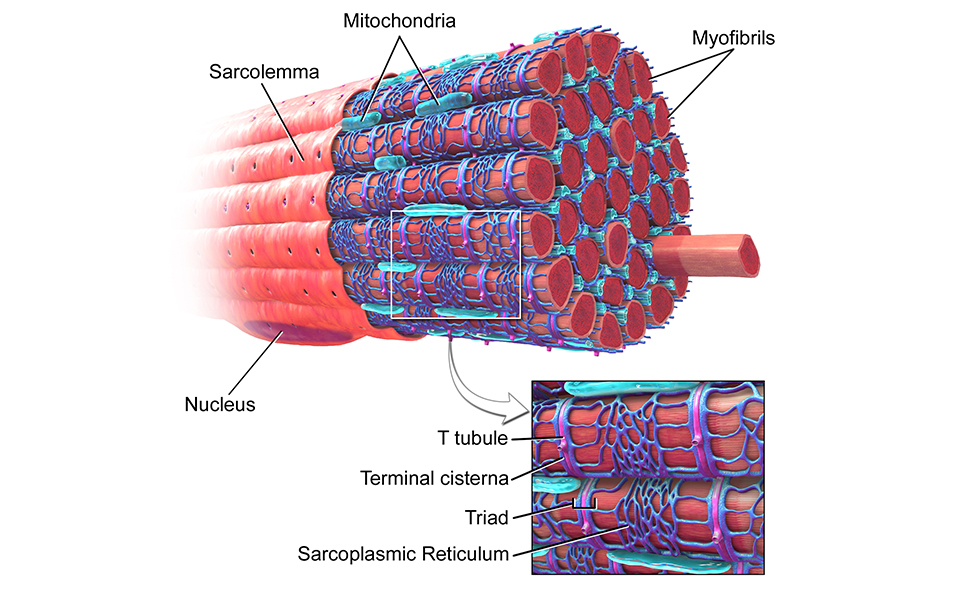 et al. Мышца на чипе: in vitro анализ сократимости гладких и поперечно-полосатых мышц. J. Pharmacol. Toxicol. Методы 65 , 126–135 (2012).
et al. Мышца на чипе: in vitro анализ сократимости гладких и поперечно-полосатых мышц. J. Pharmacol. Toxicol. Методы 65 , 126–135 (2012).
CAS PubMed PubMed Central Google ученый
Benam, K.H. и другие. Разработаны модели in vitro болезни. Ann. Преподобный Патол. 10 , 195–262 (2015).
CAS Google ученый
Ванденбург, Х. Функциональная оценка и дизайн ткани скелетных мышц. Ann. NY Acad. Sci. 961 , 201–202 (2002).
PubMed Google ученый
Ванденбург, Х., Шански, Дж., Дель Татто, М. и Хромиак, Дж. Органогенез скелетных мышц в культуре тканей. Methods Mol. Med. 18 , 217–225 (1999).
CAS PubMed Google ученый
Vandenburgh, H. et al. Автоматический скрининг лекарств с сократительной мышечной тканью, созданной из дистрофических миобластов. FASEB J. 23 , 3325–3334 (2009).
et al. Автоматический скрининг лекарств с сократительной мышечной тканью, созданной из дистрофических миобластов. FASEB J. 23 , 3325–3334 (2009).
CAS PubMed PubMed Central Google ученый
Ванденбург, Х.и другие. Платформа для скрининга лекарств, основанная на сократимости тканевой мышцы. Мышечный нерв 37 , 438–447 (2008).
CAS PubMed Google ученый
Demestre, M. et al. Формирование и характеристика нервно-мышечных соединений между мотонейронами, происходящими от hiPSC, и мышечными трубками. Stem Cell Res. 15 , 328–336 (2015).
CAS PubMed Google ученый
Дас, М., Рамси, Дж. У., Бхаргава, Н., Станческу, М., Хикман, Дж. Дж. Инженерия ткани скелетных мышц: модель созревания, способствующая долгосрочному выживанию мышечных трубок, структурному развитию аппарата сопряжения возбуждения и сокращения и экспрессии тяжелой цепи миозина новорожденных. Биоматериалы 30 , 5392–5402 (2009).
Биоматериалы 30 , 5392–5402 (2009).
CAS PubMed PubMed Central Google ученый
Дас, М., Рамси, Дж. У., Бхаргава, Н., Stancescu, M. & Hickman, J.J. Определенная долговременная in vitro тканевая инженерная модель нервно-мышечных соединений. Биоматериалы 31 , 4880–4888 (2010).
CAS PubMed PubMed Central Google ученый
Guo, X. et al. Формирование нервно-мышечного соединения между мотонейронами, полученными из стволовых клеток человека, и скелетными мышцами крысы в определенной системе. Tissue Eng. Часть C Методы 16 , 1347–1355 (2010).
PubMed PubMed Central Google ученый
Askanas, V. et al. Накопление CK-MM нарушается в иннервируемых и сокращающихся культивируемых мышечных волокнах у пациентов с мышечной дистрофией Дюшенна. Life Sci. 41 , 927–933 (1987).
Life Sci. 41 , 927–933 (1987).
CAS PubMed Google ученый
Смит, А.С., Лонг, С.Дж., Пироцци, К. и Хикман, Дж.J. Функциональная система для высокопроизводительного скрининга нервно-мышечных соединений. Технологии 1 , 37–48 (2013).
CAS PubMed PubMed Central Google ученый
Thomson, S.R. и другие. Морфологические характеристики мотонейронов не определяют их относительную предрасположенность к дегенерации на мышиной модели тяжелой спинальной мышечной атрофии. PloS One 7 , e52605 (2012).
CAS PubMed PubMed Central Google ученый
Puttonen, K.A. и другие. Создание функциональных нервно-мышечных соединений из линий плюрипотентных стволовых клеток человека. Фронт. Клетка. Neurosci. 9 , 473 (2015).
PubMed PubMed Central Google ученый
Rohwedel, J. et al. Формирование постсинаптических мембран во время дифференцировки эмбриональных стволовых клеток in vitro . Exp. Cell Res. 239 , 214–225 (1998).
et al. Формирование постсинаптических мембран во время дифференцировки эмбриональных стволовых клеток in vitro . Exp. Cell Res. 239 , 214–225 (1998).
CAS PubMed Google ученый
Костроминова, Т.Ю., Кальве, С., Арруда, Е.М. и Ларкин, Л.М.Ультраструктура мышечно-сухожильных соединений в конструкциях сухожилий и скелетных мышц, сконструированных in vitro . Histol. Histopathol. 24 , 541–550 (2009).
PubMed PubMed Central Google ученый
Луи, П.P. Технология стволовых клеток для регенерации сухожилий: текущее состояние, проблемы и направления будущих исследований. Клонирование стволовых клеток 7 , 163–174 (2015).
Google ученый
Juhas, M., Engelmayr, G.C. Младший, Фонтанелла, А.Н., Палмер, Г.М. И Бурсак, Н. Биомиметически сконструированные мышцы, способные к интеграции сосудов и функциональному созреванию. Proc. Natl. Акад. Sci. США 111 , 5508–5513 (2014).
Proc. Natl. Акад. Sci. США 111 , 5508–5513 (2014).
CAS PubMed Google ученый
Голобова Д. и др. Формирование эндотелиальной сети в тканевых скелетных мышцах человека. Tissue Eng. Часть A 21 , 2548–2558 (2015).
CAS PubMed PubMed Central Google ученый
Carosio, S. et al. Получение eX vivo — васкуляризированной мышечно-инженерной ткани (X-MET). Sci. Отчет 3 , 1420 (2013).
PubMed PubMed Central Google ученый
Levenberg, S. et al. Инженерия васкуляризированной ткани скелетных мышц. Nat. Biotechnol. 23 , 879–884 (2005).
CAS PubMed Google ученый
Додсон, М.В., Вирк, Дж. Л., Хосснер, К. Л., Бирн, К., и Макнамара, Дж. P. Развитие и использование определенной системы совместного культивирования мышц и жира. Tissue Cell 29 , 517–524 (1997).
P. Развитие и использование определенной системы совместного культивирования мышц и жира. Tissue Cell 29 , 517–524 (1997).
CAS PubMed Google ученый
Shoji, E. et al. Ранний патогенез мышечной дистрофии Дюшенна смоделирован на индуцированных пациентами плюрипотентных стволовых клетках. Sci. Отчетность 5 , 12831 (2015).
CAS PubMed PubMed Central Google ученый
Абуджарур, Р.и другие. Миогенная дифференцировка плюрипотентных стволовых клеток, индуцированных специфической мышечной дистрофией, для использования в открытии лекарств. Stem Cells Пер. Med. 3 , 149–160 (2014).
CAS PubMed PubMed Central Google ученый
Ясуно Т. и др. Функциональный анализ миоцитов, полученных из ИПСК пациента с дефицитом карнитин-пальмитоилтрансферазы II. Biochem. Биофиз. Res. Commun. 448 , 175–181 (2014).
Biochem. Биофиз. Res. Commun. 448 , 175–181 (2014).
CAS PubMed Google ученый
Tanaka, A. et al. Эффективная и воспроизводимая миогенная дифференцировка из iPS-клеток человека: перспективы моделирования миопатии Миёси in vitro . PloS One 8 , e61540 (2013).
CAS PubMed PubMed Central Google ученый
Kawagoe, S. et al. Генерация индуцированных плюрипотентных стволовых (iPS) клеток, полученных из мышиной модели болезни Помпе, и дифференцировка клеток Pompe-iPS в клетки скелетных мышц. Мол. Genet. Метаб. 104 , 123–128 (2011).
CAS PubMed Google ученый
Young, C.S. et al. Единая стратегия делеции CRISPR-Cas9, нацеленная на большинство пациентов с МДД, восстанавливает функцию дистрофина в мышечных клетках, происходящих от hiPSC. Стволовая клетка 18 , 533–540 (2016).
Стволовая клетка 18 , 533–540 (2016).
CAS PubMed PubMed Central Google ученый
Тран, Т., Андерсен, Р., Шерман, С.П. и Пайл, А.Д. Понимание развития скелетных мышц и их применения в регенеративной медицине. Внутр. Rev. Cell Mol. Биол. 300 , 51–83 (2013).
CAS PubMed Google ученый
Li, H.L. et al. Точная коррекция гена дистрофина у пациента с мышечной дистрофией Дюшенна, индуцированная плюрипотентными стволовыми клетками с помощью TALEN и CRISPR-Cas9. Stem Cell Rep. 4 , 143–154 (2015).
CAS Google ученый
Йокота, Т., Пистилли, Э., Дадди, В. и Нагараджу, К. Потенциал опосредованной олигонуклеотидами терапии с пропуском экзонов при мышечной дистрофии Дюшенна. Мнение эксперта. Биол. Ther. 7 , 831–842 (2007).
CAS PubMed Google ученый
Kazuki, Y. et al. Полная генетическая коррекция iPS-клеток мышечной дистрофии Дюшенна. Мол. Ther. 18 , 386–393 (2010).
CAS PubMed Google ученый
van Deutekom, J.C. et al. Восстановление локального дистрофина с помощью антисмыслового олигонуклеотида PRO051. New Engl. J. Med. 357 , 2677–2686 (2007).
CAS PubMed Google ученый
Skuk, D. & Tremblay, J.P. Внутримышечная трансплантация клеток как потенциальное лечение миопатий: клинические и доклинические данные. Мнение эксперта. Биол. Ther. 11 , 359–374 (2011).
PubMed Google ученый
Островидов С. и др. Дифференциация стволовых клеток в сторону миогенного происхождения для регенерации мышечной ткани: основное внимание уделяется мышечной дистрофии. Stem Cell Rev. 11 , 866–884 (2015).
Stem Cell Rev. 11 , 866–884 (2015).
CAS Google ученый
Бурзак, Н., Джухас, М. и Рандо, Т.А. Синергия инженерии и биологии для лечения и моделирования травм и заболеваний скелетных мышц. Ann. Преподобный Биомед. Англ. 17 , 217–242 (2015).
CAS Google ученый
Ляо, Х. и Чжоу, G.Q. Развитие и прогресс инженерии скелетно-мышечной ткани. Tissue Eng. Часть B Ред. 15 , 319–331 (2009).
PubMed Google ученый
Конинг, М., Harmsen, M.C., van Luyn, M.J., Werker, P.M. Текущие возможности и проблемы в инженерии тканей скелетных мышц. J. Tissue Eng. Regen. Med. 3 , 407–415 (2009).
CAS PubMed Google ученый
Туровская А., Фигероа-Мазот X. и Фольч А. Долгосрочные микрофлюидные культуры микрочипов миотрубок для высокопроизводительной фокальной стимуляции. Nat. Protoc. 1 , 1092–1104 (2006).
Долгосрочные микрофлюидные культуры микрочипов миотрубок для высокопроизводительной фокальной стимуляции. Nat. Protoc. 1 , 1092–1104 (2006).
CAS PubMed PubMed Central Google ученый
Reimann, J. et al. Распределение Pax7 в биоптатах скелетных мышц человека и культурах миогенных тканей. Cell Tissue Res. 315 , 233–242 (2004).
PubMed Google ученый
Коттлорс М. и Киршнер Дж. Повышенное количество сателлитных клеток при мышечной дистрофии Дюшенна. Cell Tissue Res. 340 , 541–548 (2010).
PubMed Google ученый
Леппер, К., Партридж, Т.А. И Фан, К. Абсолютная потребность в Pax7-положительных сателлитных клетках при регенерации скелетных мышц, вызванной острым повреждением. Разработка 138 , 3639–3646 (2011).
CAS PubMed PubMed Central Google ученый
Самбасиван, р.и другие. Экспрессирующие Pax7 сателлитные клетки незаменимы для регенерации скелетных мышц взрослых. Разработка 138 , 3647–3656 (2011).
CAS PubMed Google ученый
Мерфи, М.М., Лоусон, Дж. А., Мэтью, С.Дж., Хатчесон, Д.А. & Кардон, Г. Сателлитные клетки, фибробласты соединительной ткани и их взаимодействия имеют решающее значение для регенерации мышц. Разработка 138 , 3625–3637 (2011).
CAS PubMed PubMed Central Google ученый
Рока И., Рекена Дж., Эдель М.Дж. и Альварес-Паломо А.Б. Миогенные предшественники iPS-клеток для заместительной терапии клеток скелетных мышц. J. Clin. Med. 4 , 243–259 (2015).
CAS PubMed PubMed Central Google ученый
Mendell, J.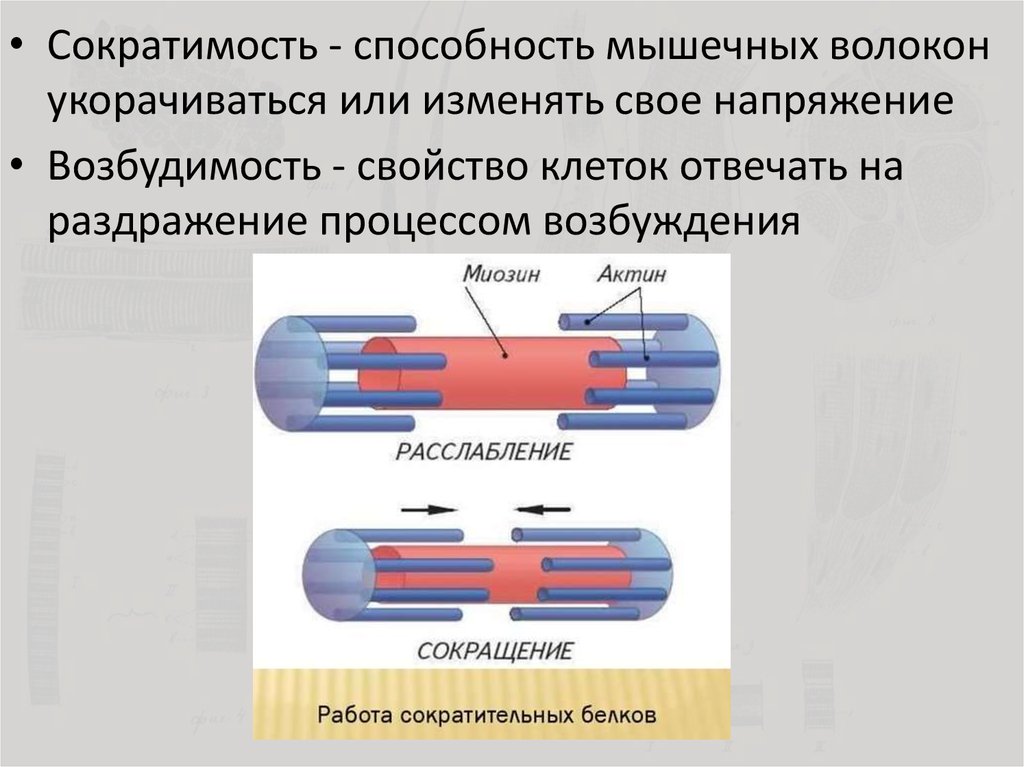 R. et al. Перенос миобластов в лечении мышечной дистрофии Дюшенна. New Engl. J. Med. 333 , 832–838 (1995).
R. et al. Перенос миобластов в лечении мышечной дистрофии Дюшенна. New Engl. J. Med. 333 , 832–838 (1995).
CAS PubMed Google ученый
Рандо, Т.А., Павлат, Г.К. И Блау, Х. Судьба миобластов после трансплантации в зрелую мышцу. Exp. Cell Res. 220 , 383–389 (1995).
CAS PubMed Google ученый
Гуссони, Э., Блау, Х.М. и Кункель, Л.М. Судьба отдельных миобластов после трансплантации в мышцы пациентов с МДД. Nat. Med. 3 , 970–977 (1997).
CAS PubMed Google ученый
Montarras, D. et al. Прямая изоляция сателлитных клеток для регенерации скелетных мышц. Наука 309 , 2064–2067 (2005).
CAS PubMed Google ученый
Коллинз, К.A. et al. Функция стволовых клеток, самообновление и поведенческая неоднородность клеток из ниши взрослых мышечных сателлитных клеток. Cell 122 , 289–301 (2005).
Функция стволовых клеток, самообновление и поведенческая неоднородность клеток из ниши взрослых мышечных сателлитных клеток. Cell 122 , 289–301 (2005).
CAS PubMed Google ученый
Sacco, A., Doyonnas, R., Kraft, P., Vitorovic, S. & Blau, H.M. Самовосстановление и разрастание одиночных пересаженных мышечных стволовых клеток. Nature 456 , 502–506 (2008).
CAS PubMed PubMed Central Google ученый
Болдрин, Л.И Морган, Дж. Э. Сателлитные клетки человека: идентификация мышечных волокон человека. PLoS Curr. 3 , РРН1294 (2011).
Google ученый
Marg, A. et al. Сателлитные клетки человека обладают регенеративной способностью и поддаются генетическим манипуляциям. J. Clin. Инвестировать. 124 , 4257–4265 (2014).
CAS PubMed PubMed Central Google ученый
Сюй, Х.и другие. Трансплантация и регенерация сателлитных клеток человека из различных скелетных мышц. Stem Cell Rep. 5 , 419–434 (2015).
CAS Google ученый
Гилберт, П.М. и другие. Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре. Наука 329 , 1078–1081 (2010).
CAS PubMed PubMed Central Google ученый
Тирни, М.T. et al. Автономное ремоделирование внеклеточного матрикса контролирует прогрессивную адаптацию регенеративной способности мышечных стволовых клеток во время развития. Cell Rep. 1–13 (2016).
Charville, G.W. и другие. Ex vivo экспансия и in vivo самообновление мышечных стволовых клеток человека. Stem Cell Rep. 5 , 621–632 (2015).
Stem Cell Rep. 5 , 621–632 (2015).
CAS Google ученый
Менг, Дж., Adkin, C.F., Xu, S.W., Muntoni, F. & Morgan, J.E. Вклад мышечных клеток человека в регенерацию скелетных мышц у мышей-хозяев с дистрофией. PloS One 6 , e17454 (2011).
CAS PubMed PubMed Central Google ученый
Таджбахш, С. Стволовые клетки скелетных мышц в онтогенетическом и регенеративном миогенезе. J. Intern. Med. 266 , 372–389 (2009).
CAS PubMed Google ученый
Бейкер, Р.К. И Лайонс, Г. Эмбриональные стволовые клетки и in vitro, мышечное развитие. Curr. Верхний. Dev. Биол. 33 , 263–279 (1996).
CAS PubMed Google ученый
Дараби, Р., Сантос, Ф. Н. И Perlingeiro, R.C. Терапевтический потенциал эмбриональных и взрослых стволовых клеток для регенерации скелетных мышц. Stem Cell Rev. 4 , 217–225 (2008).
Н. И Perlingeiro, R.C. Терапевтический потенциал эмбриональных и взрослых стволовых клеток для регенерации скелетных мышц. Stem Cell Rev. 4 , 217–225 (2008).
CAS PubMed Google ученый
Salani, S. et al. Получение клеток скелетных мышц из эмбриональных и индуцированных плюрипотентных стволовых клеток в качестве модели in vitro и для терапии мышечных дистрофий. J. Cell. Мол. Med. 16 , 1353–1364 (2012).
CAS PubMed PubMed Central Google ученый
Вилкин, Дж.Т. Превращение патологических клеток в терапевтические: одиссея через плюрипотентность. Мол. Ther. 20 , 2012–2014 (2012).
CAS PubMed PubMed Central Google ученый
Swierczek, B., Ciemerych, M.A., Archacka, K. От плюрипотентности к миогенезу: многоступенчатый процесс в блюде. J. Muscle Res. Cell Motil. 36 , 363–375 (2015).
CAS PubMed PubMed Central Google ученый
Грабовская, И., Archacka, K., Czerwinska, A.M., Krupa, M. & Ciemerych, M.A. Плюрипотентные стволовые клетки мыши и человека и средства их миогенной дифференцировки. Результаты Пробл. Cell Differ. 55 , 321–356 (2012).
CAS PubMed Google ученый
Вильшут, К.Дж., Линг, В.Б. И Бернштейн, H.S. Краткий обзор: терапия мышечных дистрофий стволовыми клетками. Stem Cells Пер. Med. 1 , 833–842 (2012).
CAS PubMed PubMed Central Google ученый
Abujarour, R. & Valamehr, B. Генерация клеток скелетных мышц из плюрипотентных стволовых клеток: достижения и проблемы. Фронт. Cell Dev. Биол. 3 , 29 (2015).
PubMed PubMed Central Google ученый
Декель, И., Магал, Ю., Пирсон-Уайт, С., Эмерсон, К.П. и Шани М. Условное преобразование ES-клеток в скелетные мышцы экзогенным геном MyoD1. New Biol. 4 , 217–224 (1992).
CAS PubMed Google ученый
Шани М. и др. Последствия конститутивной экспрессии MyoD1 в ES-клетках и эмбрионах мыши. Symp. Soc. Exp. Биол. 46 , 19–36 (1992).
CAS PubMed Google ученый
Дэвис, Р.Л., Вайнтрауб, Х. и Лассар, А. Экспрессия одной трансфицированной кДНК превращает фибробласты в миобласты. Cell 51 , 987–1000 (1987).
CAS PubMed Google ученый
Комай, Г. и Таджбахш, С. Молекулярная и клеточная регуляция скелетного миогенеза. Curr. Верхний. Dev. Биол. 110 , 1–73 (2014).
CAS PubMed Google ученый
Дараби, Р.и другие. Функциональная регенерация скелетных мышц из дифференцирующихся эмбриональных стволовых клеток. Nat. Med. 14 , 134–143 (2008).
CAS PubMed Google ученый
Рао, Л. и др. Высокоэффективное получение скелетных мышечных трубок из эмбриональных стволовых клеток человека. Stem Cell Rev. 8 , 1109–1119 (2012).
Google ученый
Goudenege, S.и другие. Миобласты, полученные из нормальных hESC и дистрофических hiPSC, эффективно сливаются с существующими мышечными волокнами после трансплантации. Мол. Ther. 20 , 2153–2167 (2012).
CAS PubMed PubMed Central Google ученый
Albini, S. et al. Эпигенетическое перепрограммирование эмбриональных стволовых клеток человека в клетки скелетных мышц и создание сократительных миосфер. Cell Rep. 3 , 661–670 (2013).
CAS PubMed PubMed Central Google ученый
Дараби Р. и др. Миогенные предшественники ES- и iPS человека восстанавливают дистрофин и улучшают сократительную способность после трансплантации дистрофическим мышам. Стволовая клетка 10 , 610–619 (2012).
CAS PubMed PubMed Central Google ученый
Fougerousse, F.и другие. Экспрессия Six и Eya во время сомитогенеза человека и активация семейства генов MyoD. J. Muscle Res. Cell Motil. 23 , 255–264 (2002).
CAS PubMed Google ученый
Goulding, M.D., Chalepakis, G., Deutsch, U., Erselius, J.R. & Gruss, P.Pax-3, новый ДНК-связывающий белок мыши, экспрессируемый на раннем этапе нейрогенеза. EMBO J. 10 , 1135–1147 (1991).
CAS PubMed PubMed Central Google ученый
Йостес, Б., Walther, C. & Gruss, P. Мышиный парный бокс-ген, Pax7, экспрессируется специфически во время развития нервной и мышечной системы. мех. Dev. 33 , 27–37 (1990).
CAS PubMed Google ученый
Gerard, M. et al. Экспрессия PAX-генов во время эмбрионального развития человека, предварительный отчет. C R Acad Sci III 318 , 57–66 (1995).
CAS PubMed Google ученый
Терзич, Дж.И Сарага-Бабич, М. Характер экспрессии генов PAX3 и PAX6 во время эмбриогенеза человека. Внутр. J. Dev. Биол. 43 , 501–508 (1999).
CAS PubMed Google ученый
Skoglund, G. et al. Физиологические и ультраструктурные особенности индуцированных человеком плюрипотентных и полученных из эмбриональных стволовых клеток скелетных миоцитов in vitro . Proc. Natl. Акад. Sci. США 111 , 8275–8280 (2014).
CAS PubMed Google ученый
Армулик, А., Дженов, Г. и Бетсхольц, К. Перициты: развитие, физиологические и патологические перспективы, проблемы и перспективы. Dev. Ячейка 21 , 193–215 (2011).
CAS PubMed PubMed Central Google ученый
Peault, B. et al. Стволовые клетки и клетки-предшественники в развитии, поддержании и лечении скелетных мышц. Мол. Ther. 15 , 867–877 (2007).
CAS PubMed Google ученый
Dellavalle, A. et al. Перициты скелетных мышц человека являются миогенными предшественниками, отличными от сателлитных клеток. Nat. Cell Biol. 9 , 255–267 (2007).
CAS PubMed Google ученый
Tedesco, F.S. и другие. Трансплантация генетически скорректированных предшественников ИПСК человека мышам с мышечной дистрофией конечностей. Science Transl. Med. 4 , 140ra189 (2012).
Google ученый
Моретти А., Лаугвиц К.Л., Дорн Т., Синнекер Д. и Маммери С. Модели сердечных заболеваний человека с плюрипотентными стволовыми клетками. Колд-Спринг-Харбор: перспективы в медицине 3 , a014027 (2013).
PubMed PubMed Central Google ученый
Робертсон, К., Тран, Д. И Джордж С.С. Краткий обзор: фазы созревания кардиомиоцитов, полученных из плюрипотентных стволовых клеток человека. Стволовые клетки 31 , 829–837 (2013).
CAS PubMed Google ученый
Bock, C. et al. Справочные карты вариаций ES- и iPS-клеток человека позволяют проводить высокопроизводительную характеристику линий плюрипотентных клеток. Cell 144 , 439–452 (2011).
CAS PubMed PubMed Central Google ученый
Боултинг, г.L. et al. Функционально охарактеризованный набор тестов индуцированных человеком плюрипотентных стволовых клеток. Nat. Biotechnol. 29 , 279–286 (2011).
CAS PubMed PubMed Central Google ученый
Halme, D.G. И Кесслер, Д.А. Регулирование FDA терапии на основе стволовых клеток. New Engl. J. Med. 355 , 1730–1735 (2006).
CAS PubMed Google ученый
Ванденбург, Х.Скрининг с высоким содержанием лекарственных средств с использованием искусственно созданных тканей опорно-двигательного аппарата. Tissue Eng. Часть B Ред. 16 , 55–64 (2010).
CAS PubMed Google ученый
Desbordes, S.C. & Studer, L. Адаптация плюрипотентных стволовых клеток человека к высокопроизводительному скринингу с высоким содержанием. Nat. Protoc. 8 , 111–130 (2013).
CAS PubMed Google ученый
Ватанабэ, К.и другие. Ингибитор ROCK обеспечивает выживание диссоциированных эмбриональных стволовых клеток человека. Nat. Biotechnol. 25 , 681–686 (2007).
CAS PubMed Google ученый
Kleinman, H.K. и другие. Комплексы базальной мембраны с биологической активностью. Биохимия 25 , 312–318 (1986).
CAS PubMed Google ученый
Хаушка, С.Д., Кенигсберг, И. Влияние коллагена на развитие мышечных клонов. Proc. Natl. Акад. Sci. USA 55 , 119–126 (1966).
CAS PubMed Google ученый
Kuhl, U., Ocalan, M., Timpl, R. & von der Mark, K. Роль ламинина и фибронектина в отборе миогенных против фиброгенных клеток из клеток скелетных мышц in vitro . Dev. Биол. 117 , 628–635 (1986).
CAS PubMed Google ученый
фон дер Марк, К. и Оджалан, М. Антагонистические эффекты ламинина и фибронектина на экспрессию миогенного фенотипа. Дифференциация 40 , 150–157 (1989).
CAS PubMed Google ученый
Мэйли, М.А., Дэвис, М.Дж. и Граундс, М.Д. Внеклеточный матрикс, факторы роста, генетика: их влияние на пролиферацию клеток и образование мышечной трубки в первичных культурах скелетных мышц взрослых мышей. Exp. Cell Res. 219 , 169–179 (1995).
CAS PubMed Google ученый
Pinset, C. & Whalen, R.G. Для индукции миогенной дифференцировки в бессывороточной среде не требуется синтеза ДНК. Dev. Биол. 108 , 284–289 (1985).
CAS PubMed Google ученый
Гото, С., Миядзаки, К., Funabiki, T. & Yasumitsu, H. Условия бессывороточного культивирования для анализа секреторных протеиназ во время миогенной дифференцировки миобластов C2C12 мыши. Анал. Biochem. 272 , 135–142 (1999).
CAS PubMed Google ученый
Reiss, K. & Korohoda, W. Формирование мышечных трубок в культурах миогенных клеток куриного эмбриона в бессывороточной среде индуцируется импульсной обработкой инсулином. Folia Histochem. Cytobiol. 26 , 133–141 (1988).
CAS PubMed Google ученый
Herrmann, B.G., Labeit, S., Poustka, A., King, T.R. & Lehrach, H. Клонирование гена T, необходимого для образования мезодермы у мышей. Nature 343 , 617–622 (1990).
CAS PubMed Google ученый
Кишперт, А.И Herrmann, B.G. Иммуногистохимический анализ белка Brachyury у эмбрионов мышей дикого типа и мутантных мышей. Dev. Биол. 161 , 179–193 (1994).
PubMed Google ученый
Юн, Дж. К. & Wold, B. Регулятор bHLH pMesogenin1 необходим для созревания и сегментации параксиальной мезодермы. Genes Dev. 14 , 3204–3214 (2000).
CAS PubMed PubMed Central Google ученый
Юн, Дж.К., Мун, Р. & Wold, B. Белок класса bHLH pMesogenin1 может определять фенотип параксиальной мезодермы. Dev. Биол. 222 , 376–391 (2000).
CAS PubMed Google ученый
Sassoon, D. et al. Экспрессия двух миогенных регуляторных факторов миогенина и MyoD1 во время эмбриогенеза мышей. Nature 341 , 303–307 (1989).
CAS PubMed Google ученый
Браун, Т.& Arnold, H.H. ES-клетки, несущие два инактивированных аллеля myf-5, образуют клетки скелетных мышц: активация альтернативного myf-5-независимого пути дифференцировки. Dev. Биол. 164 , 24–36 (1994).
CAS PubMed Google ученый
Яблонька-Реувени, З. & Патерсон, Б.М. Паттерны экспрессии MyoD и миогенина в культурах миобластов эмбрионов и взрослых цыплят. J. Histochem. Cytochem. 49 , 455–462 (2001).
CAS PubMed Google ученый
Lyons, G.E., Ontell, M., Cox, R., Sassoon, D. & Buckingham, M. Экспрессия генов миозина в развивающихся скелетных мышцах эмбриона мыши. J. Cell Biol. 111 , 1465–1476 (1990).
CAS PubMed Google ученый
Силл, П.и другие. Pax7 необходим для спецификации миогенных сателлитных клеток. Cell 102 , 777–786 (2000).
CAS PubMed Google ученый
Заммит, П.С. и другие. Pax7 и миогенная прогрессия в сателлитных клетках скелетных мышц. J. Cell Sci. 119 , 1824–1832 (2006).
CAS PubMed Google ученый
Маккейн, М.L., Agarwal, A., Nesmith, H.W., Nesmith, A.P., Parker, K.K. Микроформованные желатиновые гидрогели для расширенного культивирования сконструированных сердечных тканей. Биоматериалы 35 , 5462–5471 (2014).
CAS PubMed PubMed Central Google ученый
Паскуалини, Ф.С., Шихи, С.П., Агарвал, А., Аратин-Шаус, Ю., и Паркер, К.К. Структурное фенотипирование кардиомиоцитов, полученных из стволовых клеток. Stem Cell Rep. 4 , 340–347 (2015).
CAS Google ученый
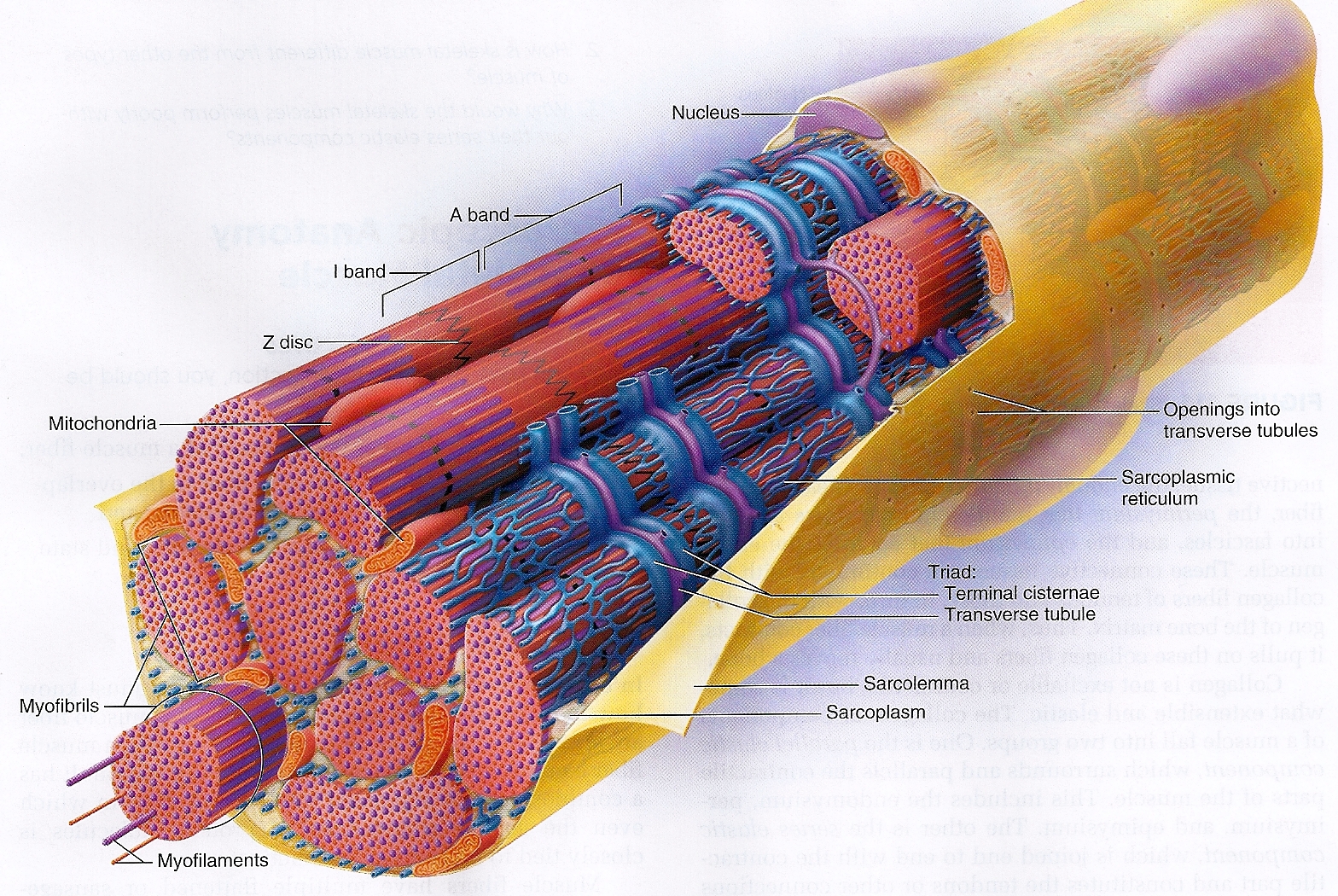
Волокна скелетных мышц | Анатомические отрывки
На изображении выше показано увеличенное изображение скелетно-полосатой мышцы.
Отдельная скелетная мышца состоит из сотен и тысяч длинных параллельных мышечных волокон, связанных вместе.
Есть три типа мышечных волокон; Тип I, Тип IIa и Тип IIb.
Тип I
Они известны как «медленно сокращающиеся» мышечные волокна.У них богатое кровоснабжение, содержащее миоглобин и множество капилляров, и они полагаются на кислород для высвобождения энергии посредством аэробного дыхания. Это делает их более устойчивыми к утомлению и способностью обеспечивать непрерывную энергию в течение более длительного времени ⚡️, но также означает, что они способны создавать лишь небольшое усилие.
Тип II
Они известны как «быстро сокращающиеся» мышечные волокна. Эти мышечные волокна сокращаются быстрее, обеспечивая прилив энергии, но они дышат анаэробно и быстрее утомляются.
Их можно подразделить на:
- «быстрый»: тип IIa
- «очень быстро»: тип IIb
Тип IIa часто называют гибридом типа I и типа II. У них богатое кровоснабжение и много митохондрий, поэтому они дышат аэробно, подобно типу I. Они также обладают способностью быстро гидролизовать АТФ, используя как аэробный, так и анаэробный метаболизм, и поэтому сокращаются быстрее, чем тип I.
Тип IIb практически не использует кислород напрямую; они анаэробны и извлекают энергию из органических молекул, хранящихся в их клетках.В результате в них меньше митохондрий, капилляров и миоглобина.
Естественно, у людей разные пропорции типов волокон в мышцах, и в разных видах спорта используются разные типы волокон. ? ️♀️ ?? Например, спринтеры, метатели и штангисты полагаются на всплески мощности от мышц с высокой долей быстро сокращающихся волокон. ?? Тот, у кого в мышцах много медленно сокращающихся волокон, вряд ли побьет рекорды на дистанции 100 метров, но кому-то с более высокой долей быстро сокращающихся мышечных волокон будет труднее прийти в форму для марафона.
Процент каждого мышечного волокна, присутствующего у человека, определяется тремя факторами: генетикой, уровнем гормонов в крови и уровнем предпринятых тренировок.
Скелетная мышца — одна из 14 моделей микроанатомии на платформе Complete Anatomy. Оцените мельчайшие детали человеческого тела в потрясающем 3D. Попробуйте БЕСПЛАТНО сегодня.
Мышечная ткань — Knowledge @ AMBOSS
Последнее обновление: 23 сентября 2021 г.
Резюме
Мышечная ткань — это мягкая ткань, которая в основном состоит из длинных мышечных волокон.Скоординированное взаимодействие миофиламентов актина и миозина внутри миоцитов дает мышечной ткани способность сокращаться. В зависимости от внутриклеточного расположения этих миофиламентов мышечная ткань классифицируется как поперечно-полосатая (скелетная и сердечная) или гладкая (гладкая). Миофиламенты поперечно-полосатой мышцы расположены в саркомеры, в то время как миофиламенты гладких мышц не имеют определенного расположения. Основные механизмы сокращения (связь возбуждения-сокращения и механизм скользящей нити) аналогичны для всех типов мышц.Скелетные мышцы находятся под произвольным контролем соматической нервной системы. Гладкая мышца находится под непроизвольным контролем вегетативной нервной системы и внешних раздражителей (например, химических, механических). Он обладает большей эластичностью и присутствует в стенках полых органов (например, желудка, мочевого пузыря, матки), стенках сосудов, дыхательных и мочевыводящих путях. Сердечная мышца также находится под непроизвольным контролем кардиостимуляторов и образует стенки камер сердца (миокарда).
Мышечные клетки (миоциты)
Обзор
[1]- Классификация
- Функция: сокращение мышц или напряжение, чтобы двигаться или сопротивляться нагрузке.
- Происхождение: мезодерма
Структуры мышечных клеток
[1]Сарколемма (миолемма)
- Определение: мембрана мышечной клетки, которая содержит мембранные впячивания.
- Характеристики
Саркоплазма
Саркоплазматический ретикулум (SR)
- Определение: эндоплазматический ретикулум мышечных клеток, который образует сеть L-канальцев.
- Характеристики
- Магазины Ca 2+
Миофиламенты
Сравнение типов мышечных клеток
[1]Миофиламенты
Миофиламенты — это белковые волокна, состоящие из толстых (миозин) и тонких (актин) нитей, которые отвечают за сократительные свойства мышечных клеток.
Структура
Функция
Белки тропонина в сердечной мышце имеют другую структуру, чем в скелетных мышцах. При инфаркте миокарда сердечные миоциты повреждаются в результате абсолютного дефицита кислорода и выбрасывают свое внутриклеточное содержимое в кровоток. Таким образом, повышение уровня сердечного тропонина в крови указывает на повреждение ткани сердечной мышцы.
Структура
- Содержит около 250 молекул миозина
- Молекулы мышечного миозина (миозин II) [2]
- Различные изоформы встречаются в разных типах мышц и определяют скорость сокращения
- Состоит из двух тяжелых и двух легких белковых цепей.
- Домены
- Голова: имеет сайт связывания актина и АТФ (который также обладает активностью АТФазы).
- Шея: две легкие белковые цепи прикрепляются к тяжелым белковым цепям, которые служат для регулирования миозиновой головки.
- Хвост: тяжелые цепи миозина намотаны друг на друга в виде спирали, с головками миозина, выступающими с обеих сторон.
- Головки миозина имеют сайт связывания для актина и сайт связывания для АТФ (который также обладает активностью АТФазы).
Функция
Как поперечнополосатые, так и гладкомышечные клетки опосредуют сокращения через актин и миозин.
Мутации в генах, кодирующих цепь тяжелого белка β-миозина или сердечного миозин-связывающего белка C, являются основной причиной семейной аутосомно-доминантной гипертрофической обструктивной кардиомиопатии.
Сравнение сократительных нитей между мышечными клетками
[1]Сокращение миофиламентов
Обзор
- Поперечно-полосатые и гладкие мышцы имеют схожие механизмы сокращения, но с некоторыми важными различиями.
- В этом разделе представлен общий обзор основных принципов мышечного сокращения.
- Подробнее об отдельных типах мышц см. Разделы ниже.
Муфта возбуждения-сжатия
[1] [3]- Определение: процесс, при котором инициирующий стимул (например,g., AP, химический стимул) вызывает повышение внутриклеточного Ca 2+ и последующее укорочение миофиламентов, что приводит к сокращению мышц.
- Типы
- Описание
В состоянии покоя актин и миозин не могут взаимодействовать, потому что они ингибируются регуляторными белками. Инициирующий стимул необходим для взаимодействия между миофиламентами.
Модель скользящей нити
[5] [6]Сравнение стимуляции и сокращения
[1]Энергетическое обеспечение сокращения мышц
- Мышечным клеткам требуется АТФ в качестве источника энергии для: [1]
- Поддержание структуры и ионных градиентов
- Взаимодействие между головкой миозина и актином, которое приводит к сокращению мышц
- Производство АТФ в мышечных клетках [1]
- См. «Пути синтеза АТФ.”
Глюкозо-аланиновый цикл обеспечивает скелетные мышцы глюкозой в качестве источника энергии. Производство мочевины в качестве побочного продукта требует много энергии, поэтому оно менее эффективно, чем цикл молочной кислоты.
Скелетная мышца
Обзор
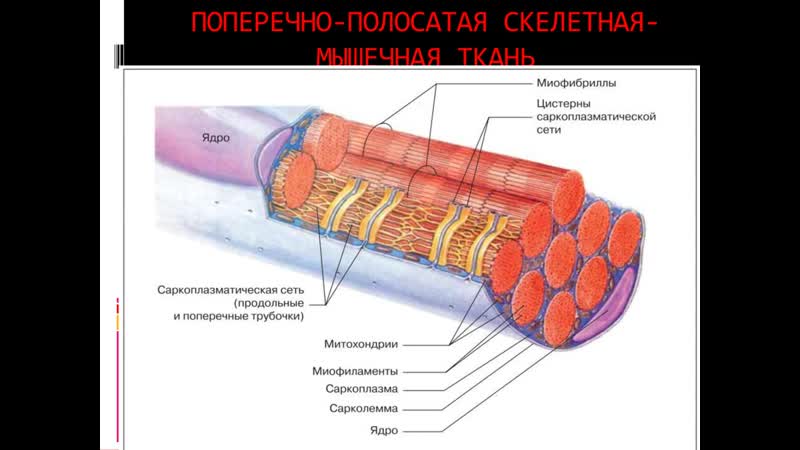
[1]- Тип: поперечно-полосатая мышечная ткань
- Структура
- Соединенные мышечные клетки образуют мышечные волокна, окруженные эндомизием.
- Пучки мышечных волокон образуют пучки, окруженные перимизием.
- Множественные пучки образуют мышцу, окруженную эпимизием.
- Мышцы и эпимизий покрыты фасцией.
- Мышцы прикрепляются к костям с помощью сухожилий.
- Регуляция сокращения
- Под произвольным контролем соматической нервной системы
- Стимуляция происходит на моторной замыкательной пластинке.
- Функция: отвечает за движение скелета
Микроскопическая анатомия
[1]- Макияж
- Регенерация: через миосателлитные клетки, которые расположены между мышечными волокнами
«Слишком (2) быстро, чтобы продолжаться; светлый и белый, лишнего воздуха нет.«Наиболее важными характеристиками волокон скелетных мышц 2 типа являются быстрое сокращение, кратковременная активность, белый цвет и анаэробный гликолиз (отсутствие воздуха).
Саркомеры
[1]Компоненты для электронной микроскопии
- Полоса Z
- Отделяет один саркомер от другого
- Действует как точка крепления
- Полосы Z сближаются во время сжатия
- Полоса M: центр зоны H, к которой с противоположных сторон прикреплены миозиновые нити
- I группа
- Группа;
- Состоит из трех сегментов: бледный центральный сегмент (зона H), окруженный двумя темными внешними сегментами
Полоса I и зона H укорачиваются во время сокращения, тогда как полоса A остается той же длины.
Анкоридж
Связь между возбуждением и сокращением в скелетных мышцах
Сокращение скелетных мышц является результатом притока внутриклеточного кальция из запасов в СР. Это объясняет способность скелетных мышц сокращаться, несмотря на лечение блокаторами кальциевых каналов, которые могут блокировать приток внеклеточного кальция через DHPR, но не могут влиять на способность определения напряжения DHPR или результирующее высвобождение внутриклеточного кальция.
Мутация гена рецептора рианодина поперечно-полосатых мышечных клеток приводит к образованию рецептора рианодина, который может активироваться некоторыми веществами, такими как ингаляционные наркотики (например, изофлуран). Эта активация приводит к неконтролируемому высвобождению Ca 2+ из SR, что приводит к непрерывному сокращению, которое значительно увеличивает потребление энергии и кислорода мышечными клетками. У больных наблюдается лактоацидоз из-за повышенного анаэробного гликолиза и гипертермии из-за повышенного мышечного метаболизма.Это опасное для жизни состояние называется злокачественной гипертермией.
Этапы цикла сокращения (цикл поперечного мостика)
[1]- Образование поперечного мостика: высвобожденный внутриклеточный Ca 2+ связывается с тропонином C и вызывает конформационные изменения → тропомиозин удаляется от сайта связывания миозина актиновая нить → миозиновая головка связывает актин под углом 90 °, образуя поперечный мост
- Силовой удар миозиновой головки: миозиновая головка высвобождает фосфат (P i ) → миозиновая головка наклоняется на 45 °, тянет миозин вдоль актина → мышца укорачивается (сокращается) → высвобождается АДФ
- Ослабление поперечного мостика: новый АТФ связывается с головкой миозина → головка миозина отделяется от актиновой нити → миозин возвращается в исходное положение
- Переориентация миозиновой головки: гидролиз АТФ до АДФ и P i (оба остаются на миозиновой головке) → миозиновая головка изменяет свою конформацию (переходит в «взведенное состояние») → миозин возвращается в исходное положение (готов к снова связываются с актином)
- Репликация цикла
- Если концентрация Ca 2+ в мышечной клетке остается повышенной, новый цикл начинается с образования поперечных мостиков.
- В зависимости от активности АТФазы (скорости расщепления АТФ в единицу времени) тяжелой цепи миозина может происходить ~ 10–100 циклов перекрестных мостиков в секунду.
- Чем больше циклов перемычки в единицу времени, тем быстрее и сильнее усадка.
Трупное окоченение — это жесткость мускулов после смерти, вызванная стойким прикреплением актина к миозину из-за недостатка АТФ.
Типы мышечных сокращений
- Изометрическое сокращение: мышца сокращается и генерирует силу, но не укорачивается и не удлиняется.
- Изотоническое сокращение: длина мышцы изменяется, а сила мышц остается постоянной.
- Концентрическое сокращение: мышца укорачивается
- Эксцентрическое сокращение: мышца удлиняется
- Ауксотоническое сокращение: одновременное изменение силы и длины мышц
Рефлексы
Гладкая мышца
Обзор
[1]- Тип: гладкая мышечная ткань
- Расположение
- Регуляция сокращения: под непроизвольным контролем вегетативной нервной системы и внешних раздражителей (например,г., химический, механический)
- Функция
Микроскопическая анатомия
[1]Гладкомышечные клетки не имеют саркомеров.
Муфта возбуждения-сжатия
- Стимулы: контролируют сокращение и расслабление (различаются от органа к органу)
- Клетки кардиостимулятора (например, интерстициальные клетки Кахаля вызывают ритмическую перистальтику в желудочно-кишечном тракте)
- Механические стимулы (например, растяжение)
- Нейротрансмиттеры вегетативной нервной системы (ацетилхолин или норадреналин) возле гладких мышц
- Метаболические стимулы (значение pH, O 2 )
- Гормоны; (е.g., NO, адреналин, гистамин, серотонин, окситоцин, вазопрессин, вазоактивный кишечный полипептид)
Гладкомышечные клетки не имеют моторных замыкательных пластинок.
Шаги сжатия
- Стимул открывает потенциалзависимые каналы Ca 2+ L-типа в сарколемме → приток Ca 2+ из внеклеточного пространства в гладкомышечные клетки (Ca 2+ также высвобождается из SR, дополнительно увеличивая внутриклеточная концентрация Ca 2+ )
- Ca 2+ связывается с кальмодулином в саркоплазме, что приводит к активации кальмодулина. Комплекс
- Ca 2+ -кальмодулин активирует киназу легкой цепи миозина (КЛЦМ).
- КЛЦМ фосфорилирует головку легкой цепи миозинового филамента.
- Активность АТФазы в фосфорилированном миозине способствует образованию перекрестных мостиков с актином, что приводит к сокращению мышц. (см. «Велосипедный спорт по мосту» выше)
Очень медленное прикрепление и отсоединение поперечных мостиков между актином и миозином позволяет гладким мышцам поддерживать длительное тоническое сокращение, потребляя при этом мало АТФ и O 2 .
Концентрация Ca 2+ определяет силу сокращения гладких мышц: чем выше приток Ca 2+ , тем больше силы создается.
Шаги расслабления
- Стимулы
- Повышенная активность фосфатазы легкой цепи миозина (MLCP) и / или снижение активности MLCK
- MLCP дефосфорилирует миозин → миозин больше не взаимодействует с актином → прекращается сокращение
Скольжение нитей и расщепление АТФ миозин-АТФазой происходит в гладких мышцах в 100–1000 раз медленнее, чем в скелетных.Таким образом, максимальная скорость сокращения гладких мышц значительно ниже, чем у скелетных мышц.
КЛЦМ фосфорилирует миозин, что приводит к сокращению гладких мышц. MLCP дефосфорилирует миозин, что приводит к расслаблению гладких мышц.
Клетки гладких мышц контактируют с Ca 2+ и расслабляются, когда НЕТ стресса.
Сердечная мышца
Обзор
[1]Микроскопическая анатомия
[1]Связь возбуждения-сокращения
Сокращение
Клиническое значение
- Мышечные состояния
- Аутоиммунные заболевания
- Новообразования
- Фармакология
- Неврологические симптомы / обследование
- Разное
| Андерсен, 1994 | Мужчины-спринтеры: N = 6 | Группа подготовки к спринтерской тренировке, постсоревновательный сезон (без компаратора) | Тренировки были 2.5–3 часа в день, 6 дней в неделю, и состоял из комбинации силовых и интервальных тренировок в течение 3 месяцев. Силовые тренировки проводились 2,5 дня в неделю и состояли из упражнений с числом повторений от одного до восьми с 80–100% от 1 ПМ. Спринт в гору или интервальный бег выполнялся 2,5 дня в неделю. Один день использовался для прыжков или развлекательных мероприятий, таких как баскетбол. | ВЛ; Биопсии были взяты после 3-недельной разгрузки (которая началась после бегового сезона) и в период после вмешательства. | Изоляция одного волокна. SDS-СТРАНИЦА. | Средние изменения: MHC I: снижение на 10,8% (52,0 +/- 3,0 до 41,2 +/- 4,7) MHC I / IIa: увеличение на 1,2% (от 0,2 +/- 0,2 до 1,4 +/- 1) MHC IIa: увеличение на 17,6% (от 34,7 +/- 6,1 до 52,3 +/- 3,6) MHC IIa / IIx: уменьшение на 7,8% (от 12,9 +/- 5,0 до 5,1 +/- 3,1) MHC IIx: уменьшение на 0,2% (0,2 +/- 0,2 до 0,0) |
| Андерсен, 1994 | Мужчины-футболисты в Датской национальной лиге: N = 14 | Группа силовых тренировок на разгибатели колена с высоким сопротивлением (ST): N = 8 Контрольная группа (без RT): N = 6 | Тренировка в группе ST проводилась в течение 12 недель 3 раза в неделю.Сеансы состояли из четырех подходов по восемь повторений разгибаний колен с использованием нагрузок высокого сопротивления со скоростью 30–50 градусов в секунду. | ВЛ; Биопсии были взяты до и после вмешательства. | Изоляция одного волокна. SDS-СТРАНИЦА. | Средние изменения: Группа ST: MHC I: увеличение на 3,1% (от 55,6 +/- 4,7 до 58,7 +/- 3,2) MHC I / IIa: уменьшение на 1,7% (с 4,4 +/- 1,2 до 2,7 + / −1,0) MHC IIa: уменьшение на 3,3% (от 33,1 +/- 4,9 до 29,8 +/- 3,8) MHC IIa / IIx: 1.Увеличение на 8% (с 6,7 +/- 3,2 до 8,5 +/- 2,2) MHC IIx: увеличение на 0,1% (от 0,2 +/- 0,2 до 0,3 +/- 0,2) Группа CTRL: все изменения менее 3%, кроме: MHC IIa: уменьшение на 6,2% (от 27,7 +/- 5,2 до 21,5 +/- 6,8) MHC IIa / IIx: увеличение на 3,9% (от 7,7 +/- 2,8 до 11,6 +/- 4,9) |
| Williamson, 2000 | Нетренированные пожилые мужчины: N = 7 | Прогрессивные тренировки с отягощениями (PRT) (без компаратора) | Продолжительность вмешательства составила 12 недель, с тренировками 3 раза в неделю.Тренировка состояла из разгибаний колен с частотой 2–3 секунды как для концентрических, так и для эксцентрических упражнений. Испытуемые выполнили три подхода; первые два подхода были по 10 повторений, а последний подход выполнялся до волевого изнеможения. Между подходами давалось около 2–3 минут отдыха. Все сеансы упражнений PRT выполнялись с частотой 80% от максимума однократного повторения (1 RM) и пересматривались каждые 2 недели, чтобы поддерживать заданную интенсивность. | ВЛ; Биопсии были взяты до и после вмешательства. | Изоляция одного волокна.SDS-СТРАНИЦА. | Распределение волокон MHC I значительно увеличилось на 10,4% после PRT, тогда как MHC IIa и IIx остались неизменными. Авторы предположили, что 12-недельный протокол тренировки с отягощениями мог оказаться недостаточным по продолжительности для укрепления и увеличения доли денервированных / реиннервируемых волокон, часто присутствующих в нетренированных пожилых людях; таким образом, волокна типа II адаптировались бы медленнее, чем волокна типа I. |
| Williamson, 2001 | Молодые нетренированные: Мужчины: N = 6 Женщины: N = 6 | Сравнение групп мужчин и женщин с прогрессивной тренировкой с отягощениями (PRT) (тот же протокол, без компаратора) | То же протокол как Уильямсон, 2000. | ВЛ; Биопсии были взяты до и после вмешательства. | Изоляция одного волокна. SDS-СТРАНИЦА. | См. Таблицу 2 Williamson, 2001 для разбивки относительного преобладания гибридных одиночных волокон. Средние изменения: Всего гибридных волокон: уменьшение на 19% у женщин и на 29% у мужчин. MHC IIa увеличился на 24% у мужчин и на 27% у женщин. Очень небольшое изменение типа I в любой группе. |
| Widrick, 2002 | Нетренированные мужчины: N = 6 | Тренировка с отягощениями (без компаратора) | Всего было выполнено 36 тренировок с частотой 3 раза в неделю в непоследовательные дни.Упражнения для нижней части тела состояли из приседаний, разгибания колен, сгибания колен и подъема икр. Упражнения для верхней части тела состояли из жима лежа, опускания широчайших вниз, жима плечами, жима на трицепс, сгибания бицепсов, тяги сидя и упражнения на пресс. Три подхода по 5–10 упражнений, перечисленных выше (разделенные примерно поровну между упражнениями для верхней и нижней части тела). Во время всех занятий тренировочное сопротивление регулировалось таким образом, чтобы испытуемые могли выполнить только указанное количество повторений плюс-минус одно повторение. | ВЛ; Биопсии были взяты до и после вмешательства. | Изоляция одного волокна. SDS-СТРАНИЦА. | Средние изменения: Тип I: снижение на 0% (42–42) I / IIa: уменьшение на 4% (4–0) IIa: увеличение на 25% (30–55) IIa / IIx: уменьшение на 19% ( 22–3) IIx: уменьшение на 3% (3–0) |
| Malisoux, 2005 | Нетренированные мужчины: N = 8 | Программа упражнений цикла сокращения растяжек (SSC) | Программа упражнений SSC, состоящая из из 24 тренировок выполнялось 3 раза в неделю, всего 5228 прыжков.Упражнения включали статический прыжок, прыжок с противодействием вертикали, прыжок с падением (высота 40 см), тройной прыжок на двух ногах, тройной прыжок на одной ноге, прыжок с барьером на двух ногах и прыжок с барьером на одной ноге. Участникам было предложено выполнять все прыжки с максимальным усилием. Количество прыжков постепенно увеличивалось в течение первых 4 недель, так что начальные тренировки длились 20 минут и длились 45 минут в конце тренировочного периода. | ВЛ; Биопсии были взяты до и после вмешательства. | Изоляция одного волокна. SDS-СТРАНИЦА. | Средние изменения: MHC I: снижение на 0,8% (от 30,0 +/- 4,9 до 29,2 +/- 4,1) MHC I / IIa: увеличение на 3,1% (от 1,9 +/- 0,5 до 5,0 +/- 1,4) MHC IIa : Увеличение на 7,2% (от 33,4 +/- 5,2 до 40,6 +/- 4,2) MHC IIa / IIx: уменьшение на 4,7% (от 26,9 +/- 1,9 до 22,2 +/- 4,3) MHC IIx: уменьшение на 4,4% (7,0 + / От −3,0 до 2,6 +/− 1,9) MHC I / IIa / IIx: снижение на 0,4% (0,8 +/− 0,4 до 0,4 +/− 0,2) |
| Trappe, 2006 | Относительно нетренированный: Мужчины: N = 4 Женщины: N = 3 | Марафонские тренировки (без компаратора) | Программа тренировок состояла из 4 дней в неделю, разделенных на две фазы: 13-недельный тренировочный период (увеличение бегового объема на 10% в неделю), затем к 3-недельному периоду снижения.По сравнению с последней неделей тренировок (13 неделя) беговой объем снизился на 25% на 14 неделе, 47% на 15 неделе и 80% за неделю до марафона. | Боковая икроножная мышца; Биопсии взяты перед 16-недельным планом тренировок, после 13 недель беговых тренировок и после 3 недель постепенного снижения нагрузки и марафона. Эксперименты с изоформой MHC одиночного мышечного волокна проводились только перед тренировочной программой и после ее постепенного снижения. | Изоляция одного волокна. SDS-СТРАНИЦА. | Средние изменения: Тип I: увеличение на 8% Тип I / IIa: уменьшение на 5% Тип IIa: увеличение на 2% Тип IIa / x: уменьшение на 4% Тип I / IIax: уменьшение на 5% Всего гибридов: Снижение на 11% |
| Люден, 2011 | Начинающие бегуны: Мужчины: N = 3 Женщины: N = 3 | Марафонская тренировка (без компаратора) | Тот же протокол, что и Trappe, 2006. | VL, Биопсии были взяты перед тренировкой, после 13 недель беговых тренировок и снова после 3 недель сокращенных тренировок после марафона (26,2 мили; 42,2 км). Эти моменты времени обозначаются соответственно как T1, T2 и T3. | Изоляция одного волокна. SDS-СТРАНИЦА. | Soleus: T1 — T3 MHC I: увеличение на 5,1% (от 71,1 +/- 6,1 до 76,2 +/- 4,7) MHC I / IIa: уменьшение на 0,3% (от 6,4 +/- 1,7 до 6,1 +/- 1,6%) MHC IIa: уменьшение на 4% (с 21,4 +/- 4,8 до 17,4 +/- 3,9) MHC IIa / IIx: 0.Снижение на 8% (1,1 +/- 0,8 до 0,3) MHC IIx: не обнаруживается в оба момента времени. VL: T1 — T3 MHC I: увеличение на 6% (от 42,6 +/- 8,6 до 48,6 +/- 7,1) MHC I / IIa: увеличение на 3,1% (от 5,1 +/- 1,3 до 8,2 + −3,0) MHC IIa : Уменьшение на 4,3% (от 40,1 +/- 6,8 до 35,8 +/- 4,4) MHC IIa / IIx: уменьшение на 5,5% (от 11,9 +/- 2,9 до 6,4 +/- 2,4) MHC IIx: увеличение на 1% (от 0 до 1 +/– 1.0 |
Как задействование мышечных волокон влияет на беговую производительность
Большинство бегунов не считают себя ограниченными в мускулах.
Это термин, предназначенный для футболистов, тяжелоатлетов или Чарльза Атласа, когда он пинает песок в лицо тощему бегуну на пляже.
Но на самом деле человеческое тело насчитывает более 600 мускулов. Мы, бегуны, пользуемся большинством из них. Тренировки, призванные задействовать и укрепить наши беговые мышцы, являются ключом к достижению высшей физической формы.
«Если вы научите свое тело задействовать мышечные волокна мощно и эффективно, вы будете бегать быстрее и увеличите свой VO 2 max», — говорит Бивертон, штат Орегон., тренер и ученый Том Шварц, чьи советы на сайте www.therunzone.com помогли множеству бегунов. «Разница между новичком и элитным бегуном в том, что мышцы новичка не были натренированы для выработки достаточной устойчивой силы».
Так что это значит для нас, бегунов на длинные дистанции? Неужели мы проведем лето на Muscle Beach, качая железом, как загорелые, высеченные из гранита бодибилдеры, которые населяют голливудские боевики и дали Калифорнии своего губернатора?
Это отрицательно.Вместо этого нам нужно включить тренировки, нацеленные на мышечные волокна, отвечающие за выносливость и скорость. Хорошая новость заключается в том, что, не осознавая этого, большинство из нас уже используют некоторые из этих тренировок. Хорошая новость заключается в том, что несколько настроек тренировок и пара дополнительных тренировок могут значительно улучшить наши показатели.
Мышечное «волокно» — это то, что мы называем мышечной клеткой. Мышечные клетки сокращаются и расслабляются в ответ на сообщения от нашей центральной нервной системы.
В нашем теле есть три типа мышц.Сердечная мышца находится в нашем сердце. Гладкая мышца контролирует непроизвольные функции, такие как пищеварение и артериальное давление. А скелетная мышца движет нашим телом, превращая ее в мышцу, на которую мы ориентируемся во время тренировки.
Скелетные мышцы составляют более трети массы нашего тела, и их можно разделить на три различных типа волокон:
Медленно сокращающиеся волокна сокращаются медленнее и менее сильно, чем волокна типа II. Бегуны на длинные дистанции любят их, потому что они изобилуют митохондриями, миоглобином и капиллярами — ингредиентами, которые обеспечивают постоянный приток кислорода и энергии, необходимые для аэробного бега.
Промежуточные волокна обладают многими аэробными характеристиками медленных волокон, но они могут сокращаться быстрее и с большей силой. Это делает их идеальными для продолжительного быстрого бега, необходимого для бега на 800 и 1500 метров.
Быстро сокращающиеся волокна — это демоны скорости мышечных клеток. Они сокращаются быстрее и сильнее всех типов волокон. Но они также ограничены неспособностью выполнять аэробные упражнения, обеспечивая только короткие всплески анаэробного бега.
Каждая скелетная мышца (икроножная мышца, подколенное сухожилие и т. Д.) Содержит волокна всех трех типов. Но не у всех бегунов одинаковый процент каждого из них. У элитных бегунов на длинные дистанции высокий процент медленных и средних волокон. Спринтеры богаты на быстроногие. Генетика определяет процентное соотношение типов волокон в наших ногах, но тренировки могут изменить их работу.
Как тренировать мышечные волокна?
Максимальное задействование
Шаблоны набора
Пониженное ингибирование
Конверсия волокна
ПРОЧНОСТЬ
Когда мы бежим, микродарвиновское «выживание сильнейших» «происходит в наших мышечных волокнах. Волокна содержат мелкие миофиламенты, называемые миозином и актином; более слабые миофиламенты повреждаются во время тренировки. Наше тело реагирует, заменяя эти поврежденные миофиламенты более прочными, оставляя нам волокна, которые лучше справляются со стрессом во время бега.
Кроме того, по мере того, как мы увеличиваем интенсивность и объем бега, количество этих миофиламентов увеличивается, вызывая набухание мышечных волокон. Именно это увеличение размера мышечных волокон (так называемая «гипертрофия») приводит к видимому увеличению наших мышц. Комбинация более сильных и крупных волокон позволяет нашим мышцам создавать больше силы, одновременно повышая их устойчивость к повреждениям и усталости.
МАКСИМАЛЬНЫЙ НАБОР
Зная, что мы можем укрепить наши мышечные волокна, понятно, что мы хотели бы укрепить их все.Однако многие бегуны этого не делают. Бегуны на длинные дистанции, которые сосредотачиваются исключительно на объеме, упускают возможность укрепить промежуточные и быстрые волокна. Спринтеры, которые избегают интервальных тренировок, не могут полностью развить промежуточные волокна.
Набор, а затем укрепление всех трех типов мышечных волокон имеет важное значение для достижения наилучшего результата. Конечно, мы должны разумно структурировать это обучение. «Мы не можем тренировать быстрые сокращения каждый день», — говорит Шварц, отмечая, что волокна обладают невысокой адаптивной энергией.«Что касается нижних волокон, вы не можете полностью стимулировать их короткими высококачественными тренировками. Они требуют сочетания низкой интенсивности и большой продолжительности, чтобы получить максимальный тренировочный эффект».
СПОСОБЫ НАБЛЮДЕНИЯ
Психолог Бойд Р. МакКэндлесс написал: «Конечная цель физического двигательного развития — сделать человеческий организм настолько независимым от силы тяжести, насколько это возможно в рамках ограничений человеческого тела».
В детстве мы научились основному контролю над мышцами путем проб и ошибок, подпитываемых нашим внутренним желанием овладеть движением и противостоять гравитации.Мы, бегуны, должны продолжать эту практику, экспериментируя с широким диапазоном темпа, усилия, силы и техники. При этом мы создаем адаптации, которые улучшают функции нашего тела в следующих областях:
Последовательность, в которой мы задействуем мышечные волокна;
Координация набора волокон при определенных темпах и уровнях утомления;
Способность воздействовать на специфические для бега волокна; и
Набор всего спектра полезных волокон.
СНИЖЕНИЕ ИНГИБИЦИИ
Когда одна мышца сокращается, другая должна расслабиться.Но если расслабление не полное (например, если мы сгибаем трицепс, пытаясь согнуть бицепс), движение тормозится. Улучшая нашу способность координировать сокращение и расслабление, мы уменьшаем торможение.
«Мышцы не знают, что делать, — говорит Шварц. «Противоположные мышцы не синхронизированы. Чтобы поднять колени, противоположная мышца должна расслабиться. Важно уменьшить это торможение. Это снижает наши затраты энергии, поэтому мы можем бежать дальше быстрее».
ПРЕОБРАЗОВАНИЕ ВОЛОКНА
Продолжаются споры о том, можно ли полностью преобразовать один тип волокна в другой.Но нет никаких сомнений в том, что более быстрые волокна можно тренировать, чтобы они приобретали характеристики более медленных волокон, что жизненно важно для бегунов на длинные дистанции, стремящихся максимизировать силу и выносливость.
«Тренируя выносливость, — говорит Шварц, — вы можете превратить даже самые быстрые волокна — с химической точки зрения — в волокна выносливости. Если мы возьмем кого-то, кто никогда раньше не тренировался, а затем тренируем его правую ногу. в течение восьми недель, а не слева, волокна спринта натренированной ноги будут выглядеть и действовать как медленные волокна нетренированной ноги.«
Шварц указывает, что у этого преобразования есть предел. Если Усэйн Болт тренировал свои быстро сокращающиеся волокна на выносливость, эти волокна увеличили бы количество митохондрий, но они никогда не достигли бы количества, равного тому, которое обнаружено в элите. медленно сокращающиеся волокна бегуна на длинные дистанции Тренировки
Понимание мышечных волокон позволяет нам разрабатывать тренировки, нацеленные на определенные типы волокон, тренировать их с правильной интенсивностью и в течение надлежащей продолжительности и обеспечивать максимальную адаптацию.
Следующие ниже тренировки являются неотъемлемой частью хорошо продуманной программы тренировок, независимо от целевой дистанции забега. Они разбиты на целевые волокна, описание тренировки и желаемый эффект. Кроме того, в каждую разбивку входят комментарии Шварца по некоторым физиологическим принципам в игре. Опять же, все это ключ к максимальной производительности на обычных дистанциях. Делайте упор на одних больше, чем на других, чтобы лучше соответствовать требованиям вашего целевого соревнования (больше тренировок, нацеленных на медленные волокна для марафонов, больше на промежуточные волокна на 5 км и т. Д.)
ДЛИННЫЙ БЕГ
ВОЛОКНО: Медленное сокращение, средний
ТРЕНИРОВКА: Долгосрочный бег — основа нашей тренировки на выносливость. Бегайте один или два раза в неделю, его продолжительность варьируется в зависимости от физической подготовки, философии и целей бега. Артур Лидьярд заставлял своих бегунов выполнять изнурительный 22-мильный бег в гору раз в неделю. Тренер ASICS Aggies Джо Рубио рекомендует два длинных бега в неделю, что составляет 20-25 процентов и 15 процентов от общего объема.Мы должны помнить, что многие желательные адаптации происходят после 90-минутной отметки. Темп обычно разговорный, с упором на количество, а не на качество.
ЭФФЕКТ: Продолжая бегать, мы обеспечиваем необходимый стресс для максимальной адаптации наших медленно сокращающихся волокон. Мы отсеивали слабые миофиламенты, заменяя их более сильными. По мере того, как наступает усталость, мы начинаем задействовать промежуточные волокна, повышая их выносливость и координируя их использование вместе с медленными волокнами, генерирующими меньшую силу.
SCHWARTZ: «Мы создаем более прочные медленно сокращающиеся волокна, поэтому нам придется использовать меньше их максимальной емкости при заданной скорости. Это позволяет сэкономить как на топливе, так и на кислороде. Кроме того, медленное волокно может стрелять быстрее на 50 процентов и более после программы марафонских тренировок ».
ДЛИННЫЕ ИНТЕРВАЛЫ
ВОЛОКНА: Медленно-сокращающиеся, средние
ТРЕНИРОВКА: Эти повторения варьируются от 2 до 5 минут (до 6 минут для бегунов в хорошей форме) и могут выполняться на беговой дорожке, дороге , или след.Темп имитирует гонку от 3 до 15 км. Восстановление может равняться продолжительности повторения на ранних занятиях, а затем сокращается по мере того, как мы становимся в более хорошей форме.
ЭФФЕКТ: Увеличивается сила как в медленных, так и в промежуточных волокнах, поскольку слабые миофиламенты заменяются более сильными. Медленно сокращающиеся волокна вынуждены работать с максимальной скоростью сокращения. А поскольку задействуются все наши медленно сокращающиеся и промежуточные волокна, мы развиваем более эффективные рабочие отношения между двумя типами волокон.
SCHWARTZ: «[Большинство бегунов] не бегают дольше 3–5 минут, потому что наша скорость упадет, и в этот момент мы тренируем более медленные и не набираем промежуточных до того же уровня. Интервалы средней длины сосредоточьтесь на силе и мощи «.
КОРОТКИЕ ИНТЕРВАЛЫ
ВОЛОКНА: Промежуточные, быстро сокращающиеся
ТРЕНИРОВКА: Эти более короткие повторения длятся от 30 до 90 секунд и могут выполняться на беговой дорожке, траве или тропинках.Темп основан на времени забега на дистанциях от 800 м до 3 км. Эффект: укрепляются как промежуточные, так и быстро сокращающиеся волокна. Два наших самых быстрых волокна учатся более эффективно взаимодействовать, уменьшая активацию ненужных волокон.
SCHWARTZ: «Короткие интервалы помогают нам развить чувство ритма нашей гонки — как нервно-мышечного, так и когнитивного. Наше тело полагается на сенсорную обратную связь для определения темпа бега: давление земли, напряжение мышц и визуально то, что мы» снова вижу «.
ПОВТОРЕНИЯ НА ДЛИННОМ ГОРУ
ВОЛОКНА: Медленное сокращение, промежуточная тренировка: продолжительностью от 30 до 90 секунд, эти повторения выполняются на умеренно крутом холме.Достаточно от четырех до 10 повторений, с восстановлением не менее 2 минут для более коротких повторений и до 5 минут для более длинных. Шварц предлагает, чтобы темп был эквивалентен тому, который мы могли бы пробежать изо всех сил в течение 3 минут в гору. Я инструктирую бегунов, которых тренирую, найти уровень усилий, позволяющий им завершить тренировку с заправкой бензина; гипотетически они должны быть способны выполнить еще одно-два повторения, если этого требует тренировка.
ЭФФЕКТ: Поскольку сила, а не скорость, создает силу, эта тренировка более эффективна, чем интервалы на треке, тропах и т. Д., при укреплении промежуточных волокон. Сохраняя наши усилия на правильном уровне, мы создаем максимальную нагрузку на наши медленно сокращающиеся и промежуточные волокна. Слишком быстрый бег заставляет больших мальчиков — наши быстро сокращающиеся волокна — выполнять тяжелую работу, что, в свою очередь, приводит к быстрой утомляемости, меньшей нагрузке на целевые волокна и более длительному периоду восстановления, прежде чем мы снова сможем усердно тренироваться.
SCHWARTZ: «Промежуточное волокно сочетает в себе умеренно высокую выработку мощности со средней продолжительностью работы.В то время как один тотальный холм в течение 60 секунд временно утомляет промежуточное волокно, он не истощает гликоген, достаточный для полного эффекта. Вы должны исчерпать примерно 75 процентов гликогена в мышечных волокнах. Это можно сделать с помощью повторений на длинных холмах ».
ПОЯВЛЕНИЯ НА КОРОТКИХ ВЫСОТАХ
ВОЛОКНА: Быстрая тренировка: короткие и крутые повторы в гору продолжительностью менее 10 секунд. Наши усилия должны достигать 90-95 процентов от максимума, а не тотальный спринт, но близко.Две-три минуты — хороший период восстановления.
ЭФФЕКТ: Эта тренировка укрепляет все типы наших волокон, но мы используем ее, чтобы сосредоточиться на быстром сокращении, для активации которого требуется 90-95 процентов усилий. Поскольку диапазон движений, необходимых для спринта в гору, больше, чем на ровной поверхности, мы задействуем более полный набор всех типов волокон. Это также одна из лучших тренировок для снижения торможения, поскольку она ограничивает активацию волокон, которые не влияют на нашу скорость.
SCHWARTZ: «Сила, а не скорость, определяет количество активированных волокон двигательных единиц. Даже если вы быстрее переворачиваете ноги на ровной поверхности, скорость не определяет активацию волокон. большой.»
TEMPO
ВОЛОКНА: Slow-Twitch, Intermediate
ТРЕНИРОВКА: Убер-тренер Джек Дэниэлс описал темп как «комфортно тяжелое» усилие — примерно на 25-30 секунд медленнее, чем темп гонки на 5 км — в течение 20 минут.Пит Пфитцингер изменяет это правило для супер-подготовленных марафонцев, предлагая пробегать до 9 миль в полумарафоне на марафонский темп.
ЭФФЕКТ: Медленно сокращающиеся волокна достигают максимальной скорости набора и сокращения примерно при 80% VO2 max — другими словами, в темповом темпе. Это означает, что все медленно сокращающиеся волокна укрепляются. Промежуточные волокна также будут задействованы, помогая нашему организму установить структуру набора для двух типов волокон в полумарафонском и марафонском темпе.
SCHWARTZ: «Бегун, у которого невысокая скорость ног — у которого нет большого количества быстро сокращающихся волокон, — не захочет выполнять большую скоростную работу.Единственный способ добиться успеха для этого бегуна — это выполнять больше работы с примерно 80% VO 2 макс. «
ПРОГРЕССИЯ
ВОЛОКНА: Медленно-сокращающийся, средний
ТРЕНИРОВКА: Мы начинаем бег легким усилием, затем снижайте темп на 10-15 секунд на каждую милю до тех пор, пока мы не сможем снова снизить темп, не достигнув стресса, характерного для гонок. Большинство бегунов достигают максимума в гоночном темпе около 5 км.
ЭФФЕКТ: Как и в случае с темпе, прогрессивные бега обеспечивают максимальную активацию медленно сокращающихся волокон.Они постепенно привлекают промежуточные волокна, улучшая координацию между типами волокон. Быстрый набор персонала на последних милях может способствовать конверсии волокна.
SCHWARTZ: «Бегун с 40-процентным VO 2 max в темпе 8:00 / милю продолжает добавлять медленно сокращающиеся волокна до тех пор, пока он не достигнет максимального набора в 7-минутном темпе, или 60-процентном максимальном VO2. В этот момент происходит «перекрытие», и промежуточные волокна прыгают внутрь. На протяжении всего пути до 85 процентов VO 2 max, медленно сокращающиеся волокна работают все быстрее, и мы постепенно увеличиваем нагрузку на наши промежуточные волокна в течение выдержанное время.Мы улучшаем как выносливость, так и силу — хотя в основном выносливость ».
ТЕХНИЧЕСКИЕ УПРАВЛЕНИЯ
ВОЛОКНА: Все волокна
ТРЕНИРОВКА: Эта тренировка, также называемая« формовыми упражнениями », включает в себя вариации пропусков. скачки, марш, прыжки, быстрые подъемы коленей, быстрые движения ног и любое количество действий, призванных имитировать, преувеличивать или усиливать наше беговое движение. Для наилучшего эффекта между упражнениями выполняются шаги 60-80 м, а бег на короткие дистанции — это завершено позже.
ЭФФЕКТ: Упражнения, такие как прыжки и удары прикладом, имитируют эффекты динамического растяжения. Ограничение и подпрыгивание более плиометрические, что влияет как на модели набора, так и на силу. Марширующие и быстрые колени уменьшают заторможенность. Чередование каждого упражнения с шагом помогает интегрировать желаемый эффект в нашу механику бега. Короткий пробег от 5 до 7 миль после упражнений закрепляет эти нервно-мышечные достижения в нашем движении.
SCHWARTZ: «Упражнения помогают нам бегать расслабленно и с большим диапазоном движений, улучшая координацию и уменьшая нервное торможение.Плиометрические упражнения влияют на двигательно-нейронную последовательность и стрельбу, что многие люди называют «отдачей» или «упругой отдачей». Улучшены некоторые более мелкие элементы нервного компонента бега, такие как рефлекс растяжения сухожилий Гольджи ».
Суммируя преимущества
Тренировка определенных типов волокон для достижения большей силы, координации набора персонала и преобразования в зависимости от гонки дает множество преимуществ в беге , в том числе:
мышечных волокон, которые могут создавать больше силы;
мышечных волокон, которые могут работать с большей скоростью;
повышенной эффективности шага;
повышенной энергоэффективности;
повышенной скорости; и
повышенной выносливости
Но это еще не все.Тренируя наши мышечные волокна, мы также увеличиваем VO 2 max, запасы гликогена в мышцах, плотность митохондрий и производство энергии (АТФ).
Мы, бегуны, склонны рассматривать развитие нашей сердечно-сосудистой системы как ключ к достижению лучшей физической формы. Но правда в том, что высокооктановое топливо не может превратить Prius в Porsche. И никакое количество кислорода — ни Gatorade, ни PowerBars, ни сила воли — не улучшит беговых характеристик, пока мы не построим более качественное беговое тело.
ОПРЕДЕЛЕНИЕ VO 2 MAX PACE
Поскольку у большинства спортсменов нет доступа к сложным тестам VO 2 , Том Шварц предлагает использовать полевые испытания.После разминки запустите гонку на время на 2400 м (2000 м для менее подготовленных бегунов) на беговой дорожке длиной 400 м.
Ваш темп в гонке на время сравним со скоростью бега при VO 2 max. Например, бегун, который завершает свою гонку на время из шести кругов за 7:50, имеет темп примерно 5:00 на милю при VO 2 max.
Для расчета своего VO 2 максимальный темп:
»Запустите гонку на время на 2400 м (2000 м, если вы относительно неопытны или возвращаетесь к реальным тренировкам).
»Разделите время финиша на 6, чтобы рассчитать темп на круге.
»Умножьте темп на круге на 4, чтобы установить темп в миле при 100% VO 2 макс.
Согласно Шварцу, ваш темп в миле для гонки на время — это ваша максимальная аэробная скорость (MAS). Шварц предлагает использовать MAS в качестве основы для определения темпа во время определенных тренировок. Его рекомендации:
Дистанция — 70% от темпа MAS — 80% от MAS
Длинные интервалы — 90% от MAS Короткие интервалы — 100% от MAS
Старший писатель ПИТ МАГИЛЛ держит три американских рекорда в возрастной группе на дистанциях от 3000 до 10 000 м.
Этот контент создается и поддерживается третьей стороной и импортируется на эту страницу, чтобы помочь пользователям указать свои адреса электронной почты. Вы можете найти больше информации об этом и подобном контенте на сайте piano.io.
Синтетический микробный процесс производит мышечные волокна, которые прочнее, чем кевлар
Источник: © gekaskr / FotoliaВы бы носили одежду или, скажем, шнурки или пояс из мышечных волокон? Что, если бы эти волокна могли выдержать больше энергии, прежде чем разорваться, чем хлопок, шелк, нейлон или даже кевлар, и производились бы без вреда для животных?
Исследователи из инженерной школы Маккелви Вашингтонского университета в Сент-Луисе.Луи разработал технологию синтетической химии, которая могла бы сделать это возможным. Этот метод позволяет им полимеризовать белки внутри сконструированных микробов. Используя эту технологию, команда разработала микробиологическое производство высокомолекулярного мышечного белка тайтина, который затем был превращен в волокна. Тесты показали, что волокна превосходят многие синтетические и натуральные полимеры.
«Прелесть системы в том, что это действительно платформа, которую можно применять где угодно», — сказал Кэмерон Сарджент, аспирант отделения биологических и биомедицинских наук.«Мы можем брать белки из разных природных контекстов, затем помещать их на эту платформу для полимеризации и создавать более крупные и длинные белки для различных материалов с большей устойчивостью». Сарджент и Кристофер Боуэн, доктор философии, который работал в отделе энергетики, окружающей среды и химической инженерии, а сейчас является старшим научным сотрудником Pfizer, являются первыми авторами статьи группы в Nature Communications , которая озаглавлена «Производство микробов. мегадальтон-тайтина дает волокна с выгодными механическими свойствами », в котором они заключили:« Эти волокна имеют потенциальное применение в областях от биомедицины до текстиля, и разработанный подход в сочетании с пониманием структуры-функции обещает ускорить дальнейшие инновации в микробиологическом производстве. из высокоэффективных материалов.”
Биология является отличным источником вдохновения для разработки материалов, поскольку природа может эффективно производить множество высокоэффективных, биоразлагаемых материалов из возобновляемых источников и с использованием только низкоэнергетических процессов, отметили авторы. Исключительно прочные шелка насекомых и подводный липкий виссус, производимый мидиями, — это лишь несколько примеров. И во многих случаях, как отмечали исследователи, «… эти натуральные материалы могут превзойти лучшие доступные альтернативы на основе нефти.”
Не всегда возможно получить такие материалы из их природных источников, и ученые не всегда могут создать синтетические методы, которые имитируют естественные процессы биосинтеза в больших масштабах. «… Необходимы разработанные стратегии микробиологического производства, чтобы облегчить практическое использование и разработку этих высокоэффективных возобновляемых материалов», — продолжила команда.
Синтетический мышечный белок, произведенный в лаборатории Фучжун Чжана, доктора философии, профессора кафедры энергетики, окружающей среды и химической инженерии, теперь произвел синтетический мышечный белок, тайтин, который является одним из трех основных белковых компонентов мышечной ткани.Критичным для механических свойств тайтина является его большой молекулярный размер. «Это самый крупный из известных белков в природе», — сказал Сарджент. По словам Чжана, мышечные волокна вызывают интерес уже давно. Исследователи пытались разработать материалы со свойствами, аналогичными свойствам мышц, для различных применений, например, в мягкой робототехнике. «Мы задавались вопросом:« Почему бы нам просто не создать синтетические мышцы? »- сказал он. «Но мы не собираемся собирать их у животных, мы будем использовать для этого микробы».
Технические микробы могут быть использованы для масштабируемого производства некоторых низкомолекулярных соединений, но прямое микробное производство полимеров с высокими механическими характеристиками ограничено, поскольку многие высокоэффективные природные материалы основаны на белках сверхвысокой молекулярной массы (UHMW) с очень повторяющиеся аминокислотные последовательности.Эти UHMW-повторяющиеся белки, как отметили ученые, «чрезвычайно трудно продуцировать в микробах из-за генетической нестабильности, низкой эффективности трансляции и метаболической нагрузки».
Чтобы обойти некоторые из проблем, которые обычно мешают бактериям продуцировать большие белки, исследовательская группа разработала бактерии, которые объединяют меньшие сегменты белка тайтина в полимеры сверхвысокого молекулярного веса размером около двух мегадальтон, что примерно в 50 раз больше среднего размера. бактериальный белок.Затем они использовали процесс мокрого прядения, чтобы превратить белки в волокна диаметром около десяти микрон, что составляет одну десятую толщины человеческого волоса.
Работая с сотрудниками Янг Шин Джун, доктор философии, профессор кафедры энергетики, окружающей среды и химической инженерии, и Синан Кетен, доктор философии, профессор факультета машиностроения Северо-Западного университета, группа затем проанализировала структуру этих волокон, чтобы идентифицировать молекулярные механизмы, обеспечивающие уникальное сочетание исключительной прочности, прочности и демпфирующей способности или способности рассеивать механическую энергию в виде тепла.Структурный анализ показал, что эти UHMW волокна тайтина содержат выровненные в осевом направлении, бок о бок пары Ig-подобных доменов. «Структурный анализ и молекулярное моделирование предполагают, что эти свойства происходят из уникальной межцепочечной кристаллизации свернутых иммуноглобулиноподобных доменов, которые сопротивляются проскальзыванию между цепями, позволяя разворачиваться внутри цепей», — пишут исследователи.
Исследователи из инженерной школы Маккелви при Вашингтонском университете в Сент-Луисе разработали метод синтетической химии для полимеризации белков внутри созданных микробов.Это позволило микробам производить высокомолекулярный мышечный белок тайтин, который затем превращался в волокна. В будущем из такого материала можно будет сделать одежду или даже защитное снаряжение. [Вашингтонский университет в Сент-Луисе]Они говорят, что, насколько им известно, это достижение представляет собой первый пример инженерного материала макроуровня, произведенного из тайтина. «Используя биосинтетическую силу микробов, эта работа позволила получить новый высокоэффективный материал, который восстанавливает не только наиболее желательные механические свойства натуральных мышечных волокон (т.е., высокая демпфирующая способность и быстрое механическое восстановление), но также и высокая прочность и ударная вязкость, даже выше, чем у многих искусственных и натуральных высококачественных волокон », — пишут они.
«… результаты моделирования показывают, что прекрасные механические свойства микробных волокон тайтина СВМП могут происходить из уникального межфибрильного спаривания свернутых Ig-подобных доменов. Такое межцепочечное нековалентное сшивание через складчатые, растяжимые домены редко исследовалось ни в органических полимерных материалах, ни в других волокнах, полученных микробиологическим путем.”
Помимо потенциального использования в модной одежде или защитной броне, Сарджент указал, что этот материал также может иметь биомедицинское применение. Поскольку он почти идентичен белкам, содержащимся в мышечной ткани, этот синтетический материал предположительно является биосовместимым и поэтому может быть отличным материалом для швов, тканевой инженерии и так далее. «Очень желательное сочетание механических свойств, устойчивого производственного процесса и способности к биологическому разложению волокна делают его отличным кандидатом для экологически безопасного применения в различных областях, от биомедицины до коммерческого текстиля (например.g., антибаллистические материалы, сетки, швы и тканевая инженерия) », — заявила команда.
«Его производство может быть дешевым и масштабируемым», — добавил Чжан. «Это может позволить использовать многие приложения, о которых люди раньше думали, но с натуральными мышечными волокнами». Исследовательская группа Чжана не собирается останавливаться на синтетических мышечных волокнах. В будущем, вероятно, будет больше уникальных материалов, которые станут возможными благодаря их стратегии микробного синтеза.